

Abstract
Understanding how novel complex traits originate involves investigating the time of origin of the trait, as well as the origin of its underlying gene regulatory network in a broad comparative phylogenetic framework. The eyespot of nymphalid butterflies has served as an example of a novel complex trait, as multiple genes are expressed during eyespot development. Yet the origins of eyespots remain unknown. Using a dataset of more than 400 images of butterflies with a known phylogeny and gene expression data for five eyespot-associated genes from over twenty species, we tested origin hypotheses for both eyespots and eyespot-associated genes. We show that eyespots evolved once within the family Nymphalidae, approximately 90 million years ago, concurrent with expression of at least three genes associated with early eyespot development. We also show multiple losses of expression of most genes from this early three-gene cluster, without corresponding losses of eyespots. We propose that complex traits, such as eyespots, may have originated via co-option of a large pre-existing complex gene regulatory network that was subsequently streamlined of genes not required to fulfill its novel developmental function.
Author Summary
Butterfly eyespots play an essential role in natural and sexual selection, yet the evolutionary origins of eyespots and of their underlying gene regulatory network remain unknown. By scoring phenotypes and wing expression of five genes in 399 and 21 nymphalid species, respectively, we tested when eyespots and expression of their associated genes evolved. We found that the origin of eyespots was concurrent with the origin of the gene expression patterns, approximately 90 million years ago. Following this event, many genes expressed in eyespot development were lost in some lineages without a corresponding loss of eyespots, indicating substantial evolution in the cluster of genes associated with eyespots. This finding suggests that complex traits such as butterfly eyespots may initially evolve by re-deploying pre-existing gene regulatory networks, which are subsequently trimmed of genes that are unnecessary in the novel context.
Introduction
One of the most conspicuous novelties in the animal world, the eyespot, has received considerable attention regarding its underlying developmental gene regulatory network [1]–[8], but the origin of this “novel” complex trait remains unknown [9]. At least twelve genes are known to be expressed in the future eyespot centers of developing wings in at least one of two model species, Bicyclus anynana (Butler) and Junonia coenia Hübner [6], [7], [10], including the transcription factors Antennapedia (Antp), spalt (sal), engrailed (en), and Distal-less (Dll) and the trans-membrane receptor Notch [2], [4], [6], [7]. However, of these twelve genes expressed in focal centers, at least six (Antp, sal, Dll, Notch, patched, and hedgehog) are not expressed in all species with eyespots [7], [8]. This variation in gene expression among species suggests eyespots may not be homologous within Nymphalidae and prompts further examination into the evolutionary origins of eyespots.
Unraveling the origins of this complex trait requires a broad comparative framework where the trait and the genes associated with the trait are investigated simultaneously. If eyespots evolved multiple times within the Nymphalidae, the developmental gene regulatory networks governing their production may not necessarily be homologous. In contrast, if nymphalid eyespots arose once, the observed variation in gene expression argues for lineage-specific changes in the developmental pathway generating eyespots. Here we integrate morphological, phylogenetic, and developmental data to evaluate the untested assumption that nymphalid eyespots are homologous. We discover that from both morphological and developmental perspectives of homology [11]–[13], nymphalid eyespots and an associated gene cluster arose a single time, early in the evolution of the Nymphalidae. From this single origin, multiple losses of gene expression have occurred, suggesting a novel means in which complex traits originate: from an initial gene regulatory network co-option followed by streamlining of extraneous network elements.
Results/Discussion
For a morphological assessment of homology we used Mayr's definition where “a feature is homologous in two or more taxa if it can be traced back to the same feature in the presumptive common ancestor.” (p. 45, [11]). If eyespots are homologous, there should be a single origin of this trait; in contrast, multiple origins of eyespots within the Nymphalidae would demonstrate that the traits are not homologous [14]. We reconstructed the history of eyespot evolution using adult wing characters for all 399 representative species included in a phylogenetic hypothesis of relationships of most nymphalid genera ([15] and Table S1). Ancestral state estimates indicate eyespots evolved once, twice, or three times in the history of the clade (Figure S1). To compare the likelihood of each of these scenarios, we performed likelihood ratio tests among the one, two, and three origin hypotheses. In all comparisons, the single-origin hypothesis provided a significantly better fit than the two- or three-origin hypotheses (Figure 1A and Table S2), demonstrating that nymphalid eyespots fulfill the phylogenetic homology criterion. Eyespots probably evolved close to the base of the Nymphalidae, after the split of the basal nymphalid subfamily Libytheinae and either before or after the split of the subfamily Danainae, during a relatively short 10 MY time window (red bars in Figure 1).
Figure 1. Origins of eyespots and associated gene expression.

(A) Origin of eyespots inferred from 399 nymphalid and 29 outgroup species from phylogeny in [13]. (B) Origin of expression in eyespot centers inferred from gene expression profiles of 23 species. Presence or absence of expression of genes in future eyespot centers indicated by black and white boxes, respectively, and grey boxes indicate species/gene combinations for which expression data are unavailable. Green bars indicate two independent origins of eyespot-associated Antp expression. In both (A) and (B), divergence times (in millions of years) are from [13], [14]; red bars on the phylogeny indicate the possible locations of the single origin of eyespots, while gold bars indicate possible locations for the single origin of gene expression for sal, Notch, Dll, and possibly en in the eyespot centers. Asterisks (*) indicate species for which expression data are from [7], [8].
We next used gene expression profiles of 21 nymphalid and two outgroup species with eyespots (Figure S2) to determine if the gene regulatory networks associated with eyespot development are homologous within nymphalids and across butterfly lineages. Networks are considered homologous in two or more taxa if all the genes and their regulatory interactions can be traced back to the same network in the most recent common ancestor [12]. We addressed the first portion of this homology assessment by testing for gene expression in the most recent common ancestor of eyespot-bearing nymphalid species. When focal expression is reconstructed on the history of Nymphalidae, expression of sal, Notch, and Dll in future eyespot centers are all estimated to have arisen once, approximately 90 million years ago (Figure 1B). Ancestral state estimates for two genes, Antp and en, were ambiguous, with one or two origins of focal expression possible. Single-origin models had the highest likelihood for en, while Antp had a maximum likelihood estimate of two origins; likelihood ratio tests on these two genes show a better fit of the model espousing a two origins for Antp expression (green bars in Figure 1B), but cannot discern between one (identical to sal, Notch, and Dll) or two independent, more recent origins for en expression (Figure 1A, Figure S3, and Table S3). These results demonstrate that a majority of eyespot associated genes investigated here (sal, Notch, Dll and potentially en) have a single origin of expression in the eyespot centers, lending support to homology of the eyespot gene regulatory network across Nymphalidae. Eyespots in the closely related Lycaenidae (Figure S2V) and the more distantly related Papilionidae [8] do not express any surveyed genes at their center, suggesting an independent and developmentally distinct origin. Cursory examination of these and other butterfly lineages (Riodinidae and Pieridae) suggests that eyespots are rare in these clades and are more likely to have evolved multiple times recently, rather than once, early in the clades' evolution, as shown here for nymphalids; however, a more thorough comparative examination of eyespot evolution in these clades should be done in future.
As a final assessment of homology, we tested whether the temporal patterns of gene expression within the network were conserved across taxa. Homologous gene regulatory networks should show similar relative temporal patterns of expression, while non-homologous networks would not be expected to show genes expressed in the same order across taxa [13]. Previous work in J. coenia and B. anynana suggested that eyespot focal genes are expressed in the following order: Antp→sal→Notch→en→Dll [5], [7], [16]. We measured establishment of focal expression in four taxa (B. anynana, Colobura dirce (Linnaeus), Vanessa cardui (Linnaeus), and J. coenia), spanning approximately 85 million years of divergence, and tested for conservation in the relative timing of expression in the future eyespot centers. Conserved timing of focal expression was highly supported in B. anynana, C. dirce, and V. cardui (Figure 2 and Figure S4). Although two of three comparisons in J. coenia were not significant, the relative timing of focal expression was qualitatively consistent with timing observed in the other three taxa. Similar patterns of conserved temporal dynamics were observed in co-stainings of pairs of genes in additional species (Figure S5). This conservation of temporal dynamics of gene expression in future eyespot centers further supports the hypothesis that the regulatory networks of eyespots are homologous.
Figure 2. Onset of gene expression in future eyespot centers shows conserved temporal dynamics among nymphalids.
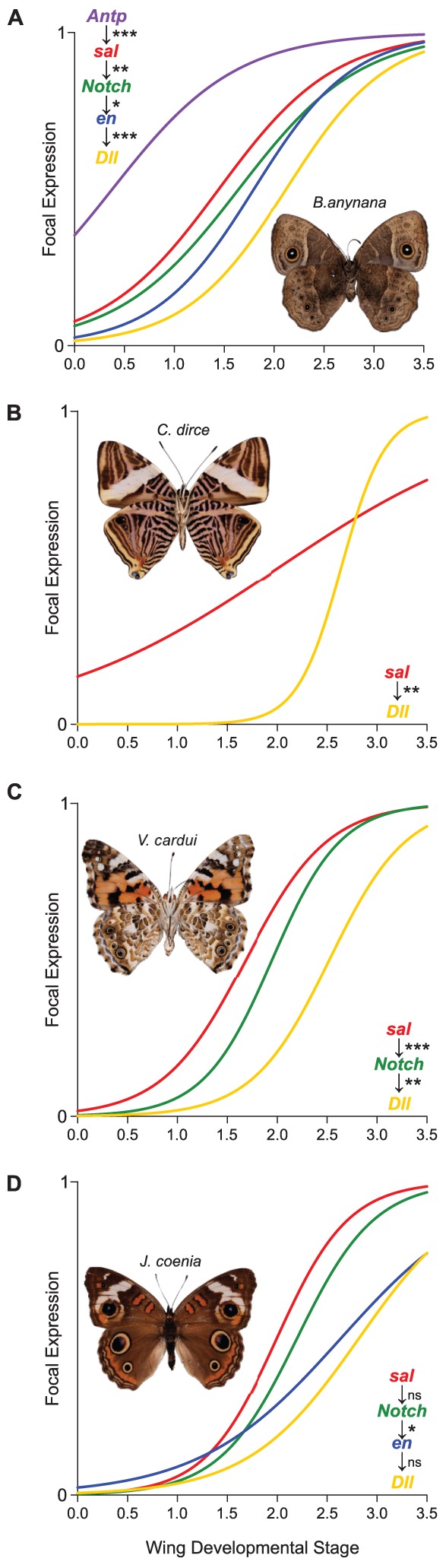
Differences in relative timing of expression in the eyespot center between genes are consistent across (A) B. anynana, (B) C. dirce, (C) V. cardui, and (D) J. coenia. ***P<0.0001, **P<0.01, *P<0.05, ns = not significant at 0.05 level.
The widespread expression of Distal-less and spalt in future eyespots of nymphalids suggests a conserved and functional role of these transcription factors in wing pattern development. Dll and sal were expressed in the future eyespot centers of all but one and two surveyed nymphalid species, respectively (Figure 1 and Figure S2). Sal expression is also associated with non-eyespot patterns in two species: in Consul fabius (Cramer), sal is expressed in larval wing discs in locations where crescent-shaped patterns develop (Figure S2F); in Siproeta stelenes (Linnaeus), wing expression of sal is associated with patches of black scales that develop on the ventral hind wings (Figure S2R).
Similarly, black patches of scales are associated with pupal stage expression of Dll and sal in B. anynana and with sal expression in the distantly related butterfly Pieris rapae (Linnaeus) (Pieridae) [6], [13]. Recent transgenic experiments suggest a functional role of Dll and sal in black scale development in B. anynana during the pupal stages of development (X. Tong, in review). Functional and comparative expression data together suggest that Dll and sal may have had a prior role in wing color pattern development, before they became co-opted into the eyespot center's regulatory network. The putative previous function of Dll and sal in color patterning wings, combined with the novel genetic background provided by other co-opted genes, may have facilitated the rapid appearance of an eyespot pattern.
With the exception of Antp, genes initially associated with eyespots demonstrate clear evidence of evolutionary lability. The genes estimated to have evolved concurrently with eyespots (sal, Notch, Dll, and possibly en) are not expressed in all future eyespot centers (Figure 1B and Figure S3). Previous studies have shown how loss of gene expression in complex regulatory networks is associated with the loss of a trait [17], but nymphalid butterflies show a pattern of network reduction without concurrent losses of eyespots. Although there is variation in the number of genes expressed in different species and the number of rings of colors in adult eyespots, we found no relationship between the two quantities (F1,5 = 1.56, p = 0.267). This widespread variation among species in the number of genes expressed during eyespot development suggests some elements of the gene regulatory network, in some species, either no longer play an essential role in eyespot development, or never played a role in the first place.
The correspondence of eyespot origins with an origin of expression of at least three of the five genes examined (Figure 1) is consistent with the hypothesis that eyespots originated from a gene network co-option event (Figure 3) [9]. This hypothesis posits a complex gene regulatory network involved in differentiating some other trait in a butterfly's body became expressed, in its entirety, in the future eyespot centers, and was subsequently rewired to generate the novel eyespot patterns. Subsequent network simplification is likely to happen when genes co-opted into the novel context are not functional in producing the novel trait. Loss of gene expression in the eyespot context may happen once network genes or their cis-regulatory elements duplicate, allowing the sub-functionalization and specialization of each copy for a different function [18]. This process of duplication and specialization provides for losses of expression in novel contexts (e.g. the eyespot), while expression is retained in the original context. Alternatively, genes from the original network may be secondarily co-opted to function in the development of the novel trait due to their fortuitous expression there. If different lineages undergo different paths of secondary co-option, this mechanism may provide an explanation for the phenomenon of developmental system drift, where networks diverge between lineages despite conservation of the final phenotype [19].
Figure 3. Regulatory network simplification in a complex trait.

Following the origin of a complex trait and its underlying developmental gene regulatory network, genes that are non-functional or unnecessary may be subsequently removed from the network (genes 2 and 3), without eliminating the trait. Genes expressed in homologous traits of all taxa may represent a ‘core network’ of regulatory elements (genes 1 and 4) that are necessary for the development of the novel trait.
The relevance of this co-option hypothesis to eyespot evolution requires additional verification from multiple lines of evidence. First, it will be key to discover the identity and interactions of all genes in the eyespot developmental network, ideally exploiting comparative transcriptomic approaches. This may allow identification of putative ancestral gene regulatory networks originally involved in the co-option event. The existence of ‘source ancestral networks’ would provide additional evidence against the alternative hypothesis that the eyespot developmental network was built gradually, gene by gene. Transcriptomic data for eyespot-bearing species in the lycaenids and riodinids, which together form the sister group to Nymphalidae, may demonstrate a completely independent origin of the eyespot gene regulatory network or identify homologous parts of the eyespot network shared with nymphalids. Functional gene knock-downs that show parallel effects in similar clusters of target genes in more than one developmental context would also support the co-option scenario. In addition, cis-regulatory elements of genes co-opted as part of a larger network to the eyespot location should be pleiotropic and drive gene expression in the eyespots and in the original developmental context, whereas no such pleiotropic elements are expected if the eyespot gene regulatory network originated de novo [9]. Finally, comparative functional data will be necessary to understand how variation in eyespot gene regulatory networks translates to variation in morphology among butterfly species.
In summary, this study highlights the utility of the comparative approach in understanding the origins and evolution of complex traits. The differences in gene expression in eyespot centers among nymphalid species suggest considerable cryptic developmental variation in a homologous trait. This type of broad comparative survey should prove useful in identifying candidates for future functional studies within and across taxa: genes expressed in all or a majority of species likely play a necessary role in the development of a complex trait and should be the primary targets of functional experiments. Future comparative work in other systems will allow for additional tests of the co-option hypothesis, to determine how often complex traits originate via bursts of complexity in gene expression, followed by genetic streamlining of unnecessary elements.
Materials and Methods
Gene Expression
We dissected final instar larval wing discs from captive reared individuals and stained for gene products using the protocol described in [20]. Bicyclus anynana larvae were collected from the Yale colony established from Malawi. Vanessa virginiensis (Drury), Polygonia interrogationis (Fabricius), and Lycaena phlaeas (Linnaeus) larvae were collected in New Haven, CT. Vanessa cardui and Danaus plexippus (Linnaeus) larvae were purchased from Educational Science (League City, TX, USA). Junonia coenia larvae were provided by L. Grunert and H.F. Nijhout at Duke University. Larvae of all other species were obtained from The Butterfly Farm at Costa Rica Entomological Supply (La Guacima, Alajuela, Costa Rica) and from surrounding farms. Wings were stained for gene products of Antp (4C3 mouse monoclonal anti-Antp at 1∶400 concentration; Developmental Studies Hybridoma Bank), sal (GP66-1 guinea pig polyclonal anti-sal at 1∶20000), Notch (C17.9C6-s mouse monoclonal anti-Notch at 1∶20), en (4F11 mouse monoclonal anti-en at 1∶5, a gift from Nipam Patel), or Dll (rabbit polyclonal anti-Dll at 1∶200, a gift from Grace Boekhoff-Falk). We used donkey anti-mouse (Jackson Immunoresearch #715-095-150), goat anti-guinea pig (Molecular Probes #A11076), and goat anti-rabbit (Molecular Probes #T-2767) secondary antibodies at a concentration of 1∶200. All wings were mounted with ProLong Gold (Invitrogen, Carlsbad, CA, USA) and images of gene expression were captured on a Nikon 90i microscope using the NIS-Elements software (Nikon Instruments, Melville, NY, USA). To confirm that focal expression was not absent due to a failed immunostaining, we looked for presence of gene expression in other areas of the wing: Dll – along the margin and in the mid-line of most wing compartments; en – in the posterior compartment; Notch – along the veins; and by co-staining wings in the same session with those of B. anynana where focal expression is present for all genes. These antibodies are cross-reactive outside the Lepidoptera suggesting that the targeted protein epitopes are extremely conserved [21]–[24]. In addition, those antibodies raised against Antp, sal, and Dll, are polyclonal, further suggesting that absence of expression of these genes is unlikely to be due to molecular evolution of the epitope sequence as there are many possible epitopes that can be targeted by the antibody. All images will be archived at http://www.lepdata.org/monteiro/lepdata.html. Expression data for seven additional taxa (Inachus io, Melitaea cinxia, Melanargia galathea, Pararge aegeria, Lasiommata megera, Heteropsis iboina, Parnassius apollo) were taken from previously published works [7], [8].
Origins of Eyespots
We reconstructed ancestral states of eyespots on the current estimate of nymphalid relationships and divergence times, and included all 29 Papilionoidea species selected as outgroups in the same phylogenetic estimate [15]. The outgroup species include representatives of all three subfamilies of Riodinidae and six of the seven families of Lycaenidae; these two families form the sister clade to the Nymphalidae. Each species was scored for presence or absence of eyespots at any location in adult wings. Eyespots were scored as any wing pattern element that (1) was roughly circular or oval and contained at least two concentric rings of color or (2) had a central pupil and a disc of color around it. All species were scored from the Yale Peabody Entomology Museum's image archive (http://www.lepdata.org/monteiro/lepdata.html). Ancestral state estimation was performed in Mesquite [25], using a two-parameter asymmetrical model of evolution, to allow different rates in gains versus losses of eyespots. Values for rate parameters were simultaneously optimized with ancestral state estimations. Divergence times within the nymphalid clade are from [15], while divergence times among families are from [26].
To test among one, two, or three origin hypotheses for the evolution of eyespots, we used BayesTraits [27] to compare likelihoods among hypotheses. In total, we compared six different models of evolution by calculating the likelihood of each model by fixing states at ancestral nodes corresponding to each hypothesis (Figure S1 and Table S2). Models were considered significantly different if log likelihood values differed by two or more log likelihood units [28].
Origins of Eyespot Focal Gene Expression
Based on estimated nymphalid relationships [15] and relationships among butterfly families [26], we estimated the ancestral state of eyespot-associated focal expression using 21 nymphalid species, one lycaenid species, and one papilionid species (all of which have eyespots on adult wings) for which expression data were available. Divergence times between two pairs of species (Vanessa cardui/V. virginiensis and Hamadryas amphinome (Linnaeus)/H. februa (Hübner)) were not available in [15], so we based divergence times on cytochrome oxidase subunit I mitochondrial DNA sequences available from GenBank and the Barcode of Life Database, using an estimate of 2.3% sequence divergence expected per million years [29]. The two Vanessa species differed, on average, by 5.45%, yielding a divergence time of 2.37 mya, and the average pairwise difference between the two Hamadryas species was 5.71%, for a divergence time of 2.48 mya. The expression data include those presented in this study, as well as expression data for Antp, sal, Notch, and Dll for seven additional species from [7], [8]. We scored each gene for each species for presence or absence of focal expression in future eyespot centers (Figure S2). We estimated ancestral states for each gene separately in Mesquite [25], using a unique two-parameter model for each gene. As with analyses of morphological data, rate parameter values were simultaneously optimized with ancestral state estimations. The expression of three genes, sal, Notch, and Dll, was unambiguously reconstructed as evolving once, while two genes, Antp and en, required explicit tests of origins to distinguish between one or two origin hypotheses.
To distinguish among models of evolution for focal expression of Antp and en, we again used likelihood ratio tests in BayesTraits [27]. Fixing states at ancestral nodes corresponding to each hypothesis, we calculated model likelihoods and compared the log likelihoods among models (Figure S3 and Table S3). We rejected models that differed by two or more log likelihood units from the best fit model [28].
Relative Timing of Eyespot Focal Gene Expression
Developmental stages of wing discs of B. anynana, C. dirce, V. cardui, and J. coenia were measured using the protocol of [5]. Wing compartments for each disc were recorded as having no focal expression (0) or focal expression (1) (Figure S6). Only those compartments that consistently displayed eyespots in adult wings were included for subsequent timing analyses. This resulted in the following compartments being analyzed: nine in B. anynana (forewing M1 and Cu1 and hindwing Rs, M1, M2, M3, Cu1, Cu2 and Pc); seven in C. dirce (forewing R5 and M1 and hindwing Rs, M1, M2, M3, and Cu1); six in V. cardui (forewing R5 and M1 and hindwing M1, M2, M3, and Cu1); and four in J. coenia (forewing M1 and Cu1 and hindwing M1 and Cu1). We first combined data across wing compartments for each species and analyzed data for each species without regard to compartment identity (e.g. data from all four compartments of J. coenia were combined in a single matrix of developmental stage and focal expression). Then, within each species, we fit a logistic curve to our data, where developmental stage was the predictor of focal expression. Data for each gene were modeled separately. To determine if timing of focal expression differed between individual genes, we used a method adapted from mRNA expression studies [30], which presents a null hypothesis of identical temporal expression between a pair of genes. The observed data from each gene are then a noisy representation of a single underlying relationship between developmental stage and focal gene expression. The difference in the temporal expression profiles between two genes is measured as the difference in the areas under each genes' logistic expression curve, δobs. To assess significance of this difference, we compared δobs to a distribution of δ generated via bootstrapping from the observed data. Briefly, to compare the temporal expression profile of gene A to gene B, we first calculated δobs. We then generated a distribution of differences (δnull) based on curves fitted to bootstrapped data, based on original sampling efforts of gene A and gene B. These bootstrapped samples are gene A, which is based on nA/2 samples observed for gene A and nA/2 samples observed for gene B, and gene B, based on nB/2 samples observed for gene A and nB/2 samples observed for gene B, where nA and nB are sample sizes for gene A and gene B, respectively. Under the null hypothesis, δobs should fall within the bounds of the δnull distribution. We performed the following comparisons, based on 10,000 bootstrap replicates in each case: B. anynana: Antp vs. sal, sal vs. Notch, Notch vs. en, and en vs. Dll; C. dirce: sal vs. Dll; V. cardui: sal vs. Notch and Notch vs. Dll; J. coenia sal vs. Notch, Notch vs. en, and en vs. Dll. Curve-fitting and bootstrapping analyses were performed in the R software package [31].
Correlation between Network Complexity and Eyespot Complexity
We tested for a correlation between the number of genes expressed in the eyespot center (network complexity) and the maximum number of different colored rings in adult eyespot pattern (trait complexity) for six nymphalid species for which complete staining profiles were available (Figure 1B). We used linear regression in the R software package [31] to determine if network complexity (number of genes) predicted eyespot complexity (number of rings).
Supporting Information
Schematic of relationships used for likelihood ratio tests of eyespot origins within Nymphalidae. Analyses conducted on tree of 399 nymphalid species+29 outgroup species (in this figure, clades of each nymphalid subfamily and outgroup family are collapsed for ease of viewing). Letters at nodes indicate nodes used for fixing ancestral states in likelihood ratio tests (see Table S2). All clades except Libytheinae and Calinaginae include at least one species with eyespots on adult wings.
(PDF)
Gene expression profiles of 21 nymphalid species and 3 outgroup species. Gene expression in larval wing discs of (A) Tithorea tarricina hindwings, (B) Danaus plexippus forewings, (C) Morpho peleides forewings, (D) Caligo memnon forewings, (E) Bicyclus anynana hindwings, (F) Consul fabius hindwings, (G) Hypna clytemenstra forewing, (H) Dryadula phaetusa hindwings, (I) Hamadryas amphinome hindwings, (J) Hamadryas februa hindwings, (K) Catonephele numilia hindwings, (L) Nessaea aglaura hindwings, (M) Myscelia cyaniris hindwings, (N) Vanessa virginiensis forewings, (O) Vanessa cardui forewings, (P) Polygonia interrogationis hindwings, (Q) Colobura dirce hindwings, (R) Siproeta stelenes hindwings, (S) Anartia fatima hindwings, (T) Chlosyne janais hindwings, (U) Junonia coenia forewings, and outgroups (V) Lycaena phlaeas (Lycaenidae) forewings, (W) Pieris rapae (Pieridae) forewings, (X) Papilio anchisiades (Papilionidae) hindwings. Expression in eyespot centers is indicated by plus signs (‘+’). Genes that displayed no elevated expression in future eyespot centers are indicated by minus signs (‘−’).
(PDF)
Relationships among taxa used for inferring history of gene expression in future eyespot centers. Divergence times (in millions of years) within Nymphalidae from [13] and divergence times among families from [14]. Numbered nodes were used for fixing ancestral states in likelihood ratio tests (see Table S3). Expression for each gene indicated as: (−) no central expression, (+) central expression; taxon/gene combinations missing symbols indicate data not available. Asterisks (*) indicate species for which expression data are from [7], [8].
(PDF)
Temporal dynamics of gene expression in future eyespot centers of four nymphalid species. Graphs show the relationship between wing developmental stage and eyespot central expression (present vs. absent). Lines show best-fit logistic curves for each gene and sizes of points indicate relative number of samples observed at each developmental stage. See Figure S6 for central expression category examples.
(PDF)
Co-stains showing temporal differences in expression of pairs of genes in the eyespot centers. (A–C) Antp before sal; (D–E) Antp before dll; (F–I) sal before Notch; (J) sal before en; (K–L) sal before Dll; (M–N) Notch before Dll; and (O–P) en before Dll.
(PDF)
Eyespot central expression categories for temporal expression analyses. Images show anti-Sal antibody stains in the Cu1 compartment of B. anynana hindwings. Gene expression was categorized as zero (central expression absent) when no pattern of up-regulation was evident (A) or when expression was only detected in the midvein area (B). Gene expression was categorized as one (central expression present), when a clear cluster of central cells was detected, either in the presence (C) or absence (D) of expression in the midvein area.
(PDF)
Data matrix for presence or absence of eyespots in 399 nymphalid taxa and 17 outgroup species.
(DOC)
Model comparisons for the number of eyespot origins in Nymphalidae. Node states refer to ancestral state (0 = eyespots absent and 1 = eyespots present) assigned to nodes as lettered in Figure S1. Differences in log likelihoods are relative to the best-fit model of a single origin, after divergence of Danainae; bold ΔlnL values indicate models providing a significantly worse fit than the single origin model.
(DOC)
Model comparisons of expression evolution of en and Antp in eyespot centers. Node states refer to ancestral state (0 = no central expression and 1 = expression in future eyespot centers) assigned to nodes as numbered in Figure S3. Differences in log likelihoods are relative to the best-fit model for each gene (the single origin model in en and the two origin, recent gain in Biblidinae model in Antp); significantly worse models are indicated by bold ΔlnL values.
(DOC)
Acknowledgments
We thank P.S. Shamble, M. Sullivan, and A. Burg for help photographing specimens; R. Tunney for assisting with immunostainings; P. Vargas for invaluable help acquiring farmed Costa Rican material and mediating export of tissues; J. Brinckerhoff for access to facilities at the Butterfly Farm in Costa Rica; and three anonymous reviewers for suggestions on an earlier draft of this manuscript. Permission to export butterfly wing tissues was provided by the Dirección General de Vida Silvestre del Ministerio de Ambiente y Energia. Images used in morphological analyses were obtained from specimens in the Yale Peabody Museum of Natural History and the American Museum of Natural History.
Funding Statement
This work was supported by a grant to AM from NSF (IOS 0818731). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Carroll SB, Gates J, Keys DN, Paddock SW, Panganiban GEF, et al. (1994) Pattern formation and eyespot determination in butterfly wings. Science 265: 109–114. [DOI] [PubMed] [Google Scholar]
- 2. Brakefield PM, Gates J, Keys D, Kesbeke F, Wijngaarden PJ, et al. (1996) Development, plasticity and evolution of butterfly eyespot patterns. Nature 384: 236–242. [DOI] [PubMed] [Google Scholar]
- 3. Keys DN, Lewis DL, Selegue JE, Pearson BJ, Goodrich LV, et al. (1999) Recruitment of a hedgehog regulatory circuit in butterfly eyespot evolution. Science 283: 532–534. [DOI] [PubMed] [Google Scholar]
- 4. Beldade P, Brakefield PM (2002) The genetics and evo-devo of butterfly wing patterns. Nat Rev Genet 3: 442–452. [DOI] [PubMed] [Google Scholar]
- 5. Reed RD, Serfas MS (2004) Butterfly wing pattern evolution is associated with changes in a Notch/Distal-less temporal pattern formation process. Curr Biol 14: 1159–1166. [DOI] [PubMed] [Google Scholar]
- 6. Monteiro A, Glaser G, Stockslager S, Glansdorp N, Ramos D (2006) Comparative insights into questions of lepidopteran wing pattern homology. BMC Dev Biol 6: 52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Saenko SV, Marialva MSP, Beldade P (2011) Involvement of the conserved Hox gene Antennapedia in the development and evolution of a novel trait. EvoDevo 2: 9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Shirai LT, Saenko SV, Keller RA, Jeronimo MA, Brakefield PM, et al. (2012) Evolutionary history of the recruitment of conserved developmental genes in association to the formation and diversification of a novel trait. BMC Evol Biol 12: 21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Monteiro A, Podlaha O (2009) Wings, horns, and butterfly eyespots: how do complex traits evolve? PLoS Biol 7: e1000037 doi:10.1371/journal.pbio.1000037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Monteiro A, Prudic KL (2010) Multiple approaches to study color pattern evolution in butterflies. Trends Evol Biol 2: e2. [Google Scholar]
- 11.Mayr E (1982) The Growth of Biological Thought. Cambridge, MA: Harvard Univ Press.
- 12.Abouheif A (1999) Establishing homology criteria for regulatory gene networks: prospects and challenges. In: Bock GR, Cardew G, editors. Homology. Novartis Foundation Symposium 222. Chichester: Wiley. pp. 207–225. [DOI] [PubMed]
- 13. Monteiro A (2012) Gene regulatory networks reused to build novel traits: Co-option of an eye-related gene regulatory network in eye-like organs and red wing patches on insect wings is suggested by optix expression. Bioessays 34: 181–186. [DOI] [PubMed] [Google Scholar]
- 14. Wake DB, Wake MH, Specht CD (2011) Homoplasy: from detecting pattern to determining process and mechanism of evolution. Science 331: 1032–1035. [DOI] [PubMed] [Google Scholar]
- 15. Wahlberg N, Leneveu J, Kodandaramaiah U, Peña C, Nylin S, et al. (2009) Nymphalid butterflies diversify following near demise at the Cretaceous/Tertiary boundary. Proc R Soc B 276: 4295–4302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Reed RD, Chen P-H, Nijhout HF (2007) Cryptic variation in butterfly eyespot development: the importance of sample size in gene expression studies. Evol Dev 9: 2–9. [DOI] [PubMed] [Google Scholar]
- 17. Abouheif E, Wray GA (2002) Evolution of the gene network underlying wing polyphenism in ants. Science 297: 249–252. [DOI] [PubMed] [Google Scholar]
- 18. Force A, Lynch M, Pickett FB, Amores A, Yan Y, et al. (1999) Preservation of duplicate genes by complementary, degenerative mutations. Genetics 151: 1531–1545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. True JR, Haag ES (2001) Developmental system drift and flexibility in evolutionary trajectories. Evol Dev 3: 109–119. [DOI] [PubMed] [Google Scholar]
- 20. Brunetti CR, Selegue JE, Monteiro A, French V, Brakefield PM, et al. (2001) The generation and diversification of butterfly eyespot color patterns. Curr Biol 11: 1578–1585. [DOI] [PubMed] [Google Scholar]
- 21. Hayward DC, Patel NH, Rehm EJ, Goodman CS, Ball EE (1995) Sequence and expression of grasshopper Antennapedia: comparison to Drosophila. Dev Biol 172: 452–465. [DOI] [PubMed] [Google Scholar]
- 22. Fehon RG, Kooh PJ, Rebay I, Regan CL, Xu T, et al. (1990) Molecular interactions between the protein products of the neurogenic loci Notch and Delta, two EGF-homologous genes in Drosophila. Cell 61: 523–534. [DOI] [PubMed] [Google Scholar]
- 23. Patel NH, Martin-Blanco E, Coleman KG, Poole SJ, Ellis MC, et al. (1989) Expression of engrailed proteins in arthropods, annelids, and chordates. Cell 58: 955–968. [DOI] [PubMed] [Google Scholar]
- 24. Panganiban G, Sebring A, Nagy L, Carroll S (1995) The development of crustacean limbs and the evolution of arthropods. Science 270: 1363–1366. [DOI] [PubMed] [Google Scholar]
- 25.Maddison WP, Maddison DR (2011) Mesquite: a modular system for evolutionary analysis, version 2.75. Available: http://mesquiteproject.org. Accessed 3 October 2011.
- 26. Heikkilä M, Kaila L, Mutanen M, Peña C, Wahlberg N (2012) Cretaceous origin and repeated tertiary diversification of the redefined butterflies. Proc R Soc B 279: 1093–1099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Pagel M, Meade A (2007) BayesTraits, version 1.0. Available: http://www.evolution.reading.ac.uk/BayesTraits.html. Accessed 28 July 2011.
- 28. Pagel M (1999) The maximum likelihood approach to reconstructing ancestral character states of discrete characters on phylogenies. Syst Biol 48: 612–622. [Google Scholar]
- 29. Brower AVZ (1994) Rapid morphological radiation and convergence among races of the butterfly Heliconius erato inferred from patterns of mitochondrial DNA evolution. Proc Natl Acad Sci U S A 91: 6491–6495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Minas C, Waddell SJ, Montana G (2011) Distance-based differential analysis of gene curves. Bioinformatics 27: 3135–3141. [DOI] [PubMed] [Google Scholar]
- 31.R Development Core Team (2011) R: a language and environment for statistical computing, Available: http://www.R-project.org.Accessed 20 December 2011.
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Schematic of relationships used for likelihood ratio tests of eyespot origins within Nymphalidae. Analyses conducted on tree of 399 nymphalid species+29 outgroup species (in this figure, clades of each nymphalid subfamily and outgroup family are collapsed for ease of viewing). Letters at nodes indicate nodes used for fixing ancestral states in likelihood ratio tests (see Table S2). All clades except Libytheinae and Calinaginae include at least one species with eyespots on adult wings.
(PDF)
Gene expression profiles of 21 nymphalid species and 3 outgroup species. Gene expression in larval wing discs of (A) Tithorea tarricina hindwings, (B) Danaus plexippus forewings, (C) Morpho peleides forewings, (D) Caligo memnon forewings, (E) Bicyclus anynana hindwings, (F) Consul fabius hindwings, (G) Hypna clytemenstra forewing, (H) Dryadula phaetusa hindwings, (I) Hamadryas amphinome hindwings, (J) Hamadryas februa hindwings, (K) Catonephele numilia hindwings, (L) Nessaea aglaura hindwings, (M) Myscelia cyaniris hindwings, (N) Vanessa virginiensis forewings, (O) Vanessa cardui forewings, (P) Polygonia interrogationis hindwings, (Q) Colobura dirce hindwings, (R) Siproeta stelenes hindwings, (S) Anartia fatima hindwings, (T) Chlosyne janais hindwings, (U) Junonia coenia forewings, and outgroups (V) Lycaena phlaeas (Lycaenidae) forewings, (W) Pieris rapae (Pieridae) forewings, (X) Papilio anchisiades (Papilionidae) hindwings. Expression in eyespot centers is indicated by plus signs (‘+’). Genes that displayed no elevated expression in future eyespot centers are indicated by minus signs (‘−’).
(PDF)
Relationships among taxa used for inferring history of gene expression in future eyespot centers. Divergence times (in millions of years) within Nymphalidae from [13] and divergence times among families from [14]. Numbered nodes were used for fixing ancestral states in likelihood ratio tests (see Table S3). Expression for each gene indicated as: (−) no central expression, (+) central expression; taxon/gene combinations missing symbols indicate data not available. Asterisks (*) indicate species for which expression data are from [7], [8].
(PDF)
Temporal dynamics of gene expression in future eyespot centers of four nymphalid species. Graphs show the relationship between wing developmental stage and eyespot central expression (present vs. absent). Lines show best-fit logistic curves for each gene and sizes of points indicate relative number of samples observed at each developmental stage. See Figure S6 for central expression category examples.
(PDF)
Co-stains showing temporal differences in expression of pairs of genes in the eyespot centers. (A–C) Antp before sal; (D–E) Antp before dll; (F–I) sal before Notch; (J) sal before en; (K–L) sal before Dll; (M–N) Notch before Dll; and (O–P) en before Dll.
(PDF)
Eyespot central expression categories for temporal expression analyses. Images show anti-Sal antibody stains in the Cu1 compartment of B. anynana hindwings. Gene expression was categorized as zero (central expression absent) when no pattern of up-regulation was evident (A) or when expression was only detected in the midvein area (B). Gene expression was categorized as one (central expression present), when a clear cluster of central cells was detected, either in the presence (C) or absence (D) of expression in the midvein area.
(PDF)
Data matrix for presence or absence of eyespots in 399 nymphalid taxa and 17 outgroup species.
(DOC)
Model comparisons for the number of eyespot origins in Nymphalidae. Node states refer to ancestral state (0 = eyespots absent and 1 = eyespots present) assigned to nodes as lettered in Figure S1. Differences in log likelihoods are relative to the best-fit model of a single origin, after divergence of Danainae; bold ΔlnL values indicate models providing a significantly worse fit than the single origin model.
(DOC)
Model comparisons of expression evolution of en and Antp in eyespot centers. Node states refer to ancestral state (0 = no central expression and 1 = expression in future eyespot centers) assigned to nodes as numbered in Figure S3. Differences in log likelihoods are relative to the best-fit model for each gene (the single origin model in en and the two origin, recent gain in Biblidinae model in Antp); significantly worse models are indicated by bold ΔlnL values.
(DOC)