

Abstract
Kainate receptors (KARs) are a subfamily of ionotropic glutamate receptors (iGluRs) that mediate excitatory synaptic transmission, regulate neurotransmitter release, and show a remarkably selective distribution in the brain. Compared to other iGluRs, the precise contribution of KARs to brain function is less understood. Unlike recombinant KARs, native KARs exhibit characteristically slow channel kinetics. The underlying explanation for this dissimilar kinetics has remained elusive until recently. New research has identified Neto1 and Neto2 as KAR auxiliary subunits that determine unique properties of synaptic KARs, including their slow kinetics and high affinity for agonist. Whether these auxiliary subunits regulate KAR trafficking and targeting at the synapse is less clear. By regulating channel gating, Neto1 and Neto2 can increase the diversity of KAR functional properties. These auxiliary subunits may represent a starting point for a better understanding of the role played by neuronal KARs under normal and pathological conditions, but also, they may provide an alternative target for the development of new drugs regulating KARs and brain function.

Susumu Tomita (left) is Associate Professor at Yale University School of Medicine and Pablo Castillo (right) is Professor of Neuroscience at the Albert Einstein College of Medicine. Their research backgrounds are in molecular biology and electrophysiology, respectively. S.T. received his PhD from the University of Tokyo in the field of Alzheimer's disease. During his postdoctoral training with David Bredt and Roger Nicoll at UCSF, he studied the role of AMPA receptor auxiliary subunits regulating receptor function and trafficking. P.C. received his MD/PhD degrees from the University of the Republic in Uruguay and did his PhD thesis work on hippocampal synaptic plasticity and kainate receptor-mediated transmission in Roger Nicoll's laboratory at UCSF in collaboration with Robert Malenka. S.T. and P.C. share a common interest in ionotropic glutamate receptors – kainate receptors in particular – and in molecular mechanisms underlying synaptic transmission and plasticity.
Introduction
Excitatory synaptic transmission in the central nervous system is largely mediated by the neurotransmitter glutamate, which binds to three functionally and structurally distinct classes of ionotropic glutamate receptors (iGluRs), i.e. amino-3-hydroxy-5-methylisoxazole-4-propionic acid receptors (AMPARs), N-methyl-d-aspartic acid receptors (NMDARs), and kainate receptors (KARs). While expression of recombinant receptors in heterologous systems can form ligand-gated ion channels, recombinant and native channels often differ in their channel properties. Increasing evidence indicates that native iGluRs do not operate in isolation but interact with auxiliary subunits that regulate key receptor properties such as their gating, subcellular localization and pharmacology (Jackson & Nicoll, 2011; Yan & Tomita, 2012). Given that AMPARs support the bulk of fast excitatory transmission in the brain, it is not surprising that most studies have focused on AMPAR auxiliary proteins, including the transmembrane AMPAR regulatory proteins (TARPs), the cornichon-like proteins (CNIHs), and others (Diaz, 2010; Kato et al. 2010; Jackson & Nicoll, 2011; Straub & Tomita, 2012). However, recent studies indicate that other iGluRs can also interact with, and be modulated by, distinct auxiliary proteins. In this review article, we summarize current evidence in support of neuropilin and tolloid like proteins (Netos) as auxiliary subunits determining key functional properties of native KARs.
Early studies showed that kainate, a natural product from seaweeds and an analogue of glutamate, induces hyper-excitation of neurons and neurological disorders including epilepsy and neuronal cell death (Johnston, 1973; Olney et al. 1974; Nadler et al. 1978). In addition, a highly selective distribution of high-affinity KARs was shown by radio-labelled [3H]kainate binding pattern of brain sections, with a particularly strong [3H]kainate signal observed in hippocampal stratum lucidum, striatum and the cerebellar granule cell layer (Foster et al. 1981; Monaghan & Cotman, 1982). In the last two decades, a wealth of studies has established that neuronal KARs can mediate synaptic transmission, regulate neurotransmitter release and control cell excitability (Lerma, 2006; Pinheiro & Mulle, 2006; Contractor et al. 2011). Consistent with the selective receptor distribution observed in the brain, KAR-mediated effects are remarkably restricted to specific synapses and neurons.
KARs are tetrameric combinations of five subunits: GluK1–5 (previously termed GluR5–7, KA1/2) (Fig. 1) (Wisden & Seeburg, 1993; Hollmann & Heinemann, 1994; Traynelis et al. 2010). Low-affinity GluK1–3 subunits can form homomeric channels, whereas high-affinity GluK4/5 subunits do not, and only participate in heteromeric receptors. While GluK1–3 can be expressed at the cell surface, GluK4/5 requires GluK1–3 for surface expression (Christensen et al. 2004; Ruiz et al. 2005; Nasu-Nishimura et al. 2006). GluK4/5 have a significantly higher affinity for kainate than GluK1–3 (Hollmann & Heinemann, 1994), and at least one of the high-affinity GluK4/5 subunits is required for normal ionotropic function of neuronal KARs (Fernandes et al. 2009).
Figure 1. Kainate receptor subunits and Neto auxiliary subunits.
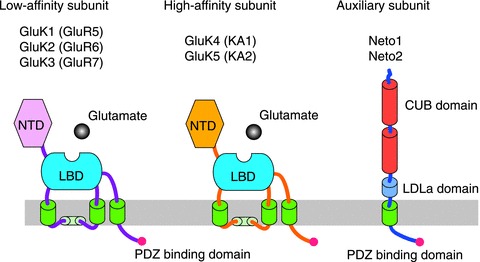
Kainate receptor subunits are classified in two classes, low- and high-affinity subunits. Low-affinity subunits, GluK1–3, can form homomeric channels, whereas high-affinity subunits GluK4/5 require low-affinity subunits to form heteromeric channels. The KAR auxiliary subunits Neto1 and Neto2 are type-1 transmembrane proteins containing two CUB domains and one LDLa domain in the extracellular domain. All three proteins have potential PDZ binding motifs at their C-terminal. NTD, N-terminal domain; LBD, ligand-binding domain.
Neto1 and Neto2 interact with native KARs
In addition to the GluK1–5 channel forming subunits, KARs in the brain also interact with the auxiliary subunits, Neto1/2 (Lerma, 2011; Yan & Tomita, 2012). Neto1/2 were co-purified with GluK2/3 from rodent brains (Zhang et al. 2009; Straub et al. 2011a; Tang et al. 2011). Both Neto1 and Neto2 share an identical and unique domain structure representing a new subfamily of transmembrane proteins containing CUB (complement C1r/C1s, Uegf, Bmp1) and LDLa (low-density lipoprotein receptor class A) domains (Fig. 1). While Neto1 and Neto2 do not interact with AMPARs, it has been reported that Neto1 interacts with NMDARs and by this means could regulate the synaptic abundance of the GluN2A subunit (Ng et al. 2009), although the interaction of Neto1 with NMDARs has not been confirmed by another study (Straub et al. 2011a). Interestingly, Neto1 and Neto2 mRNA expression display a complementary profile in the mouse brain. Neto2 mRNA is abundantly expressed in cerebellar granule cells and cortical neurons, whereas Neto1 mRNA is highly expressed in the CA3 pyramidal neurons (Michishita et al. 2003, 2004; Ng et al. 2009). Further, robust Neto1 labelling is observed by immunohistochemistry in the CA3 s. lucidum (Straub et al. 2011a; Tang et al. 2011), the mf-CA3 synaptic field where high-affinity KARs are highly expressed (Foster et al. 1981; Monaghan & Cotman, 1982).
Kainate receptor function is regulated by Neto1/2
Neuronal KARs mediate a characteristically slow excitatory postsynaptic current (KAR-EPSC) that was originally identified at the mossy fibre to CA3 pyramidal cell (mf-CA3) synapse (Fig. 2A) (Castillo et al. 1997; Vignes & Collingridge, 1997). Similar slow KAR-EPSCs have also been reported in hippocampal interneurons (Cossart et al. 1998; Frerking et al. 1998), cerebellar Golgi cells (Bureau et al. 2000), basolateral amygdala neurons (Li & Rogawski, 1998), thalamic relay neurons (Miyata & Imoto, 2006), neocortical neurons (Kidd & Isaac, 1999; Ali, 2003; Wu et al. 2005), off-bipolar cells of the retina (DeVries & Schwartz, 1999), and dorsal horn neurons (Li et al. 1999). The slow kinetics of native KARs clearly contrasts with the fast activation, deactivation and desensitization of recombinant KARs. While synaptic KARs display both slow rise times (5–10 ms) and decay kinetics (30–150 ms time constant) (Castillo et al. 1997; Kidd & Isaac, 1999; Cossart et al. 2002; Wu et al. 2007), recombinant KARs desensitize/deactivate in a few milliseconds (Erreger et al. 2004; Contractor et al. 2011). Despite the comparatively small KAR-EPSC amplitude, the distinct slow time course of KAR-mediated synaptic responses allows significantly large charge transfer and temporal summation of the synaptic responses, in particular during repetitive activation of presynaptic inputs (Frerking & Ohliger-Frerking, 2002). In this way, synaptic KARs can contribute significantly to spike generation (Miyata & Imoto, 2006; Sachidhanandam et al. 2009) and network activity (Cunningham et al. 2006; Goldin et al. 2007).
Figure 2. Distinct kinetics of native KARs in the brain.
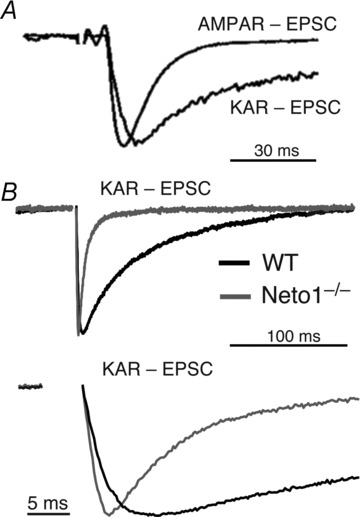
A, in CA3 pyramidal cells of the hippocampus, mossy fibre activation elicits a fast kinetics AMPAR-EPSC and a distinct slow kinetics of KAR-EPSC. These responses have been scaled to the peak of the AMPAR-EPSC. The KAR-EPSC was recorded in the presence of the AMPAR selective antagonist GYKI53655. From Castillo et al. (1997); reprinted by permission from Macmillan Publishers Ltd: Nature©1997. B, in Neto1 knockout mice, both rise time and decay kinetics of mf-CA3 KAR-EPSCs are substantially accelerated. Normalized KAR-EPSCs from wild-type and Neto1 knockout mice are superimposed and depicted at a slow (top panel) and fast (bottom panel) time base. From Straub et al. (2011a); reprinted by permission from Macmillan Publishers Ltd: Nature Neuroscience©2011.
The underlying explanation for the dissimilar kinetics of synaptic and recombinant KARs has remained elusive until recently. It was initially thought that KARs might be located extrasynaptically and be activated by glutamate spillover, which can occur during presynaptic repetitive activity. In this scenario, the slow KAR-EPSC kinetics would be due to glutamate diffusing from the synaptic cleft. The evidence against this possibility is twofold: first, manipulations that increase extrasynaptic glutamate do not affect the KAR-EPSC time course (Castillo et al. 1997; Kidd & Isaac, 1999), and second, KARs can be activated by quantal release and the resulting synaptic response (i.e. miniature KAR-EPSC) has the typical slow kinetics (Cossart et al. 2002). Another possibility is that the high-affinity GluK4/5 subunits could modify the GluK1–3 intrinsic channel gating. Indeed, GluK5, which forms heteromeric receptors with GluK2 in the brain, slows down the decay kinetics of GluK2 homomer channels in heterologous cells (Barberis et al. 2008). Consistent with this observation, KAR-EPSCs become slightly faster in GluK5 knockout mice (Contractor et al. 2003), but not as fast as the homomeric KAR-mediated currents in heterologous cells, suggesting that some other, more fundamental mechanism is involved.
Native iGluRs form macromolecular complexes with scaffolding proteins, enzymes, trafficking chaperone or adaptor proteins and, therefore, the slow kinetics of synaptic KARs could be due to some regulatory/auxiliary protein. Such a protein should fulfil the following requirements: First, it should be expressed by synapses showing slow KAR-EPSCs; second, its co-expression in heterologous cells should slow down the kinetics of recombinant KAR-mediated responses; third, removal of this protein from synapses (or interfering with its interaction with KARs) should accelerate the time course of KAR-EPSCs. A number of KAR-interacting proteins were found, some of which have been implicated in regulating channel kinetics. For example, SAP90/PSD-95, which was first identified as an interacting protein of recombinant GluK2 subunits, alters receptor function by reducing channel desensitization (Garcia et al. 1998). However, subsequent studies using outside-out patch membranes reported that SAP90/PSD-95 does not modulate the rate at which receptors desensitize, but rather accelerates recovery from desensitization (Bowie et al. 2003). The KAR interacting protein for GluR6/GluK2 (KRIP6) was also shown to modify channel kinetics in heterologous cells. KRIP6 reduces peak-current amplitude and steady-state desensitization, but does not significantly alter decay kinetics (Laezza et al. 2007). The role of these KAR-interacting proteins in modulating native KARs remains unexplored.
More recently, the newly identified KAR interacting proteins Neto1 and Neto2 have emerged as auxiliary subunits that determine key properties of native KARs, including their slow channel kinetics. Remarkably, both Neto1 and Neto2 slow deactivation and desensitization of GluK1–3 homomers and GluK2/5 heteromers in heterologous systems, and accelerate recovery from desensitization (Zhang et al. 2009; Copits et al. 2011; Straub et al. 2011a,b). At the single channel level, Neto2 increases the open probability of KARs (Zhang et al. 2009). Moreover, loss of Neto1, but not Neto2, accelerates the decay kinetics of KAR-EPSCs at mf-CA3 synapses (Fig. 2B) (Straub et al. 2011a; Tang et al. 2011). In fact, the time course of KAR-EPSCs and AMPAR-EPSCs (both rise time and decay) becomes indistinguishable in Neto1 knockout mice (Straub et al. 2011a). These findings indicate that the auxiliary subunit Neto1 determines the slow kinetics of synaptic KARs. While Neto2 can regulate KAR function in cultured neurons (Zhang et al. 2009; Copits et al. 2011; Straub et al. 2011a,b), whether this auxiliary subunit can also regulate synaptic KARs in the brain awaits confirmation.
Neto auxiliary subunits determine high agonist affinity of kainate receptors
While AMPARs and NMDARs distribute virtually everywhere in the brain, KARs show a very distinct distribution, as indicated by strong, nanomolar range [3H]kainate labelling in hippocampal s. lucidum, cerebellar granule cell layer, and striatum (Fig. 3A) (Foster et al. 1981; Monaghan & Cotman, 1982). Another long-standing question in the field has been how this distinct distribution of high-affinity KARs in the brain is defined. The [3H]kainate signal is abolished in GluK2 knockout mice (Mulle et al. 1998), suggesting that this signal is a true marker of neuronal KARs. The selective [3H]kainate-binding pattern could be simply due to the presence of the high-affinity subunits GluK4/5. However, given that some mismatch between [3H]kainate binding and GluK4/5 expression patterns exists (Bahn et al. 1994), another mechanism is likely to be implicated. Indeed, the [3H]kainate binding is strongly reduced in Neto1 knockout mice (Fig. 3B), and importantly, this change occurs in the absence of any reduction of GluK2/GluK5 expression at synapses as shown by immunohistochemistry of GluK2/GluK5 in s. lucidum (Fig. 3B) and biochemical PSD fractionation (Straub et al. 2011a). Consistent with these observations, as well as the Neto1-mediated actions on KAR channel gating, Neto1 increases KAR affinity for agonist both in the brain and in heterologous cells expressing GluK2 and GluK5 (Straub et al. 2011a). Regardless, most evidence indicates that by increasing KAR affinity for agonist, Neto1 determines the classical high-affinity [3H]kainate-binding pattern in the brain, while not affecting the synaptic localization of KARs. Whether Neto1-mediated increase in KAR affinity for agonist is accompanied by changes in KAR subunit composition remains to be tested.
Figure 3. Distinct distribution of KARs in the brain.
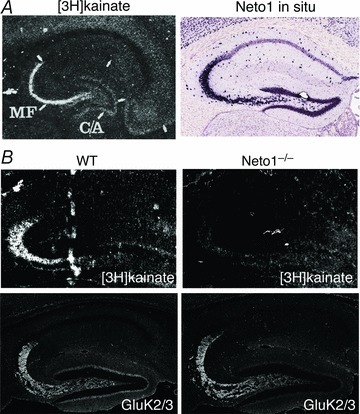
A, [3H]kainate signal shows distinct distribution of high-affinity kainate receptors in the hippocampal stratum lucidum, where mossy fibre (MF) and CA3 pyramidal cells form synapses Modified from Monaghan & Cotman (1982); by permission from Elsevier: Brain Research©1982. Neto1 mRNA is strongly expressed in CA3 pyramidal cells. Image from Allen Brain Atlas. B, [3H]kainate signal was reduced in Neto1 knockout mice (Neto1−/−), whereas GluK2/3 localized at synapses, suggesting that Neto1 only modulates [3H]kainate affinity, but not KAR synaptic localization. Modified from Straub et al. (2011a); reprinted by permission from Macmillan Publishers Ltd: Nature Neuroscience©2011.
Do Neto1/2 regulate KAR trafficking?
The role of AMPAR auxiliary subunits in regulating AMPAR trafficking and targeting is well established (Jackson & Nicoll, 2011; Opazo & Choquet, 2011; Straub & Tomita, 2012). However, whether Neto1/2 play a similar role for KARs is less clear. The mechanism controlling KAR surface expression has been linked to specific motifs encoded in KAR subunit amino acid sequences, as well as post-translational modifications of KAR subunits by phosphorylation, palmitoylation and SUMOylation (Isaac et al. 2004; Jaskolski et al. 2005; Contractor et al. 2011). Several observations argue against Neto1/2 playing a significant role in KAR trafficking. Neto2 robustly enhances glutamate-evoked GluK2 currents without increasing the surface expression of KARs in cRNA-injected oocytes (Zhang et al. 2009). In addition, neither GluK2/3 surface expression (e.g. assessed by surface biotinylation), nor GluK2/3 PSD expression and synaptic localization is altered in the hippocampus of Neto1 KO mice (Straub et al. 2011a). On the other hand, another study has shown that expression of GluK2/5 is reduced at hippocampal PSD of Neto1 KO but not Neto2 KO mice (Tang et al. 2011). The reason(s) for this apparent discrepancy is unclear. Finally, Neto2, but not Neto 1, was recently found to enhance the surface expression of the GluK1 subunit in transfected HEK cells, although this enhancement does not seem to fully account for the observed increase in agonist-mediated response (Zhang et al. 2009; Copits et al. 2011; Straub et al. 2011a,b). Future studies will be necessary to determine the precise role of Neto1/2 in regulating trafficking and targeting of native KARs.
Concluding remarks and future questions
It is now clear that Neto1 and Neto2 are auxiliary subunits of native KARs that have significant effects on their functional properties. However, several important questions remain unanswered. How exactly do Neto subunits regulate KAR function? Addressing this question will require a better understanding of the KAR-Neto interaction at the structural level. It will also be important to know the stoichiometry of this interaction and whether a change in the number of Neto subunits in a single KAR complex could determine the magnitude of Neto-mediated effects on KAR function. Do Neto subunits equally interact with all KAR subunits, or are there some interactions that are more favourable than others? Furthermore, it would be interesting to see whether the Neto-KAR interaction could be physiologically regulated and whether Neto subunits could be involved in activity-dependent plasticity of KAR-mediated transmission.
The generality of the Neto-KAR interaction and its role in synaptic KAR function throughout the brain remain to be investigated. To date, the effects of Neto1/2 have been examined at the mf-CA3 synapse only (Straub et al. 2011a; Tang et al. 2011) but KARs have been observed at several other synapses throughout the brain (see above). Intriguingly, fast KAR-EPSCs (i.e. with a similar fast decay kinetics to AMPAR-EPSCs) have been reported at unitary connections in rat motor cortex (Ali, 2003), Layer V pyramidal neurons in rat somatosensory cortex (Eder et al. 2003), and at Purkinje neurons (Huang et al. 2004), raising the possibility that at some synapses KARs could operate in a Neto1/2-independent manner. Thus far, most evidence indicates that Neto1/2 target postsynaptic but not presynaptic KARs. At the mf-CA3 synapse, where both types of KARs have been identified, presynaptic function is normal in Neto1/2 knockout mice (Straub et al. 2011a; Tang et al. 2011). This observation is entirely consistent with the much stronger Neto1/2 in situ profile in CA3 pyramidal cells compared to dentate granule neurons, which give rise to mossy fibres (Michishita et al. 2003, 2004; Ng et al. 2009). Future studies will have to determine whether Neto proteins can be found at presynaptic terminals and if they play a role in controlling neurotransmitter release. While most attention on Neto1/2 has been focused towards the ionotropic function of KARs, neuronal KARs also mediate unconventional metabotropic (G protein-mediated) signalling (Rodriguez-Moreno et al. 1997; Melyan et al. 2002). Whether Neto subunits can regulate this form of signalling remains untested. Finally, it will be interesting to see if Neto subunits can also regulate extrasynaptic KARs (Eder et al. 2003).
KARs have an important role in neuronal excitability and network activity, and have been implicated in epilepsy and other neuropsychiatric conditions (Bowie, 2008; Vincent & Mulle, 2009). Given the functional impact of Neto1/2 on KAR function, these auxiliary proteins could offer an interesting therapeutic target. For example, a drug disrupting the Neto-KAR interaction, by suppressing the functional contribution of KARs, could suppress network excitability, thereby reducing the threshold for seizure development. Such a drug could make a significant addition to our rather limited pharmacological toolbox for KARs.
Acknowledgments
The authors thank members of the Castillo lab and Tomita lab for helpful discussions, and David Hunt for critical reading of the manuscript. S.T. and P.E.C. are supported by grants from the US National Institutes of Health/National Institute of Mental Health (R01 MH085080 and R01 MH081935, respectively).
References
- Ali AB. Involvement of post-synaptic kainate receptors during synaptic transmission between unitary connections in rat neocortex. Eur J Neurosci. 2003;17:2344–2350. doi: 10.1046/j.1460-9568.2003.02677.x. [DOI] [PubMed] [Google Scholar]
- Bahn S, Volk B, Wisden W. Kainate receptor gene expression in the developing rat brain. J Neurosci. 1994;14:5525–5547. doi: 10.1523/JNEUROSCI.14-09-05525.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barberis A, Sachidhanandam S, Mulle C. GluR6/KA2 kainate receptors mediate slow-deactivating currents. J Neurosci. 2008;28:6402–6406. doi: 10.1523/JNEUROSCI.1204-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowie D. Ionotropic glutamate receptors & CNS disorders. CNS Neurol Disord Drug Targets. 2008;7:129–143. doi: 10.2174/187152708784083821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowie D, Garcia EP, Marshall J, Traynelis SF, Lange GD. Allosteric regulation and spatial distribution of kainate receptors bound to ancillary proteins. J Physiol. 2003;547:373–385. doi: 10.1113/jphysiol.2002.033076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bureau I, Dieudonne S, Coussen F, Mulle C. Kainate receptor-mediated synaptic currents in cerebellar Golgi cells are not shaped by diffusion of glutamate. Proc Natl Acad Sci U S A. 2000;97:6838–6843. doi: 10.1073/pnas.97.12.6838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castillo PE, Malenka RC, Nicoll RA. Kainate receptors mediate a slow postsynaptic current in hippocampal CA3 neurons. Nature. 1997;388:182–186. doi: 10.1038/40645. [DOI] [PubMed] [Google Scholar]
- Christensen JK, Paternain AV, Selak S, Ahring PK, Lerma J. A mosaic of functional kainate receptors in hippocampal interneurons. J Neurosci. 2004;24:8986–8993. doi: 10.1523/JNEUROSCI.2156-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Contractor A, Mulle C, Swanson GT. Kainate receptors coming of age: milestones of two decades of research. Trends Neurosci. 2011;34:154–163. doi: 10.1016/j.tins.2010.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Contractor A, Sailer AW, Darstein M, Maron C, Xu J, Swanson GT, Heinemann SF. Loss of kainate receptor-mediated heterosynaptic facilitation of mossy-fiber synapses in KA2-/- mice. J Neurosci. 2003;23:422–429. doi: 10.1523/JNEUROSCI.23-02-00422.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Copits BA, Robbins JS, Frausto S, Swanson GT. Synaptic targeting and functional modulation of GluK1 kainate receptors by the auxiliary neuropilin and tolloid-like (NETO) proteins. J Neurosci. 2011;31:7334–7340. doi: 10.1523/JNEUROSCI.0100-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cossart R, Epsztein J, Tyzio R, Becq H, Hirsch J, Ben-Ari Y, Crepel V. Quantal release of glutamate generates pure kainate and mixed AMPA/kainate EPSCs in hippocampal neurons. Neuron. 2002;35:147–159. doi: 10.1016/s0896-6273(02)00753-5. [DOI] [PubMed] [Google Scholar]
- Cossart R, Esclapez M, Hirsch JC, Bernard C, Ben-Ari Y. GluR5 kainate receptor activation in interneurons increases tonic inhibition of pyramidal cells. Nat Neurosci. 1998;1:470–478. doi: 10.1038/2185. [DOI] [PubMed] [Google Scholar]
- Cunningham MO, Pervouchine DD, Racca C, Kopell NJ, Davies CH, Jones RS, Traub RD, Whittington MA. Neuronal metabolism governs cortical network response state. Proc Natl Acad Sci U S A. 2006;103:5597–5601. doi: 10.1073/pnas.0600604103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeVries SH, Schwartz EA. Kainate receptors mediate synaptic transmission between cones and ‘Off’ bipolar cells in a mammalian retina. Nature. 1999;397:157–160. doi: 10.1038/16462. [DOI] [PubMed] [Google Scholar]
- Diaz E. Regulation of AMPA receptors by transmembrane accessory proteins. Eur J Neurosci. 2010;32:261–268. doi: 10.1111/j.1460-9568.2010.07357.x. [DOI] [PubMed] [Google Scholar]
- Eder M, Becker K, Rammes G, Schierloh A, Azad SC, Zieglgansberger W, Dodt HU. Distribution and properties of functional postsynaptic kainate receptors on neocortical layer V pyramidal neurons. J Neurosci. 2003;23:6660–6670. doi: 10.1523/JNEUROSCI.23-16-06660.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erreger K, Chen PE, Wyllie DJ, Traynelis SF. Glutamate receptor gating. Crit Rev Neurobiol. 2004;16:187–224. doi: 10.1615/critrevneurobiol.v16.i3.10. [DOI] [PubMed] [Google Scholar]
- Fernandes HB, Catches JS, Petralia RS, Copits BA, Xu J, Russell TA, Swanson GT, Contractor A. High-affinity kainate receptor subunits are necessary for ionotropic but not metabotropic signaling. Neuron. 2009;63:818–829. doi: 10.1016/j.neuron.2009.08.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Foster AC, Mena EE, Monaghan DT, Cotman CW. Synaptic localization of kainic acid binding sites. Nature. 1981;289:73–75. doi: 10.1038/289073a0. [DOI] [PubMed] [Google Scholar]
- Frerking M, Malenka RC, Nicoll RA. Synaptic activation of kainate receptors on hippocampal interneurons. Nat Neurosci. 1998;1:479–486. doi: 10.1038/2194. [DOI] [PubMed] [Google Scholar]
- Frerking M, Ohliger-Frerking P. AMPA receptors and kainate receptors encode different features of afferent activity. J Neurosci. 2002;22:7434–7443. doi: 10.1523/JNEUROSCI.22-17-07434.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garcia EP, Mehta S, Blair LA, Wells DG, Shang J, Fukushima T, Fallon JR, Garner CC, Marshall J. SAP90 binds and clusters kainate receptors causing incomplete desensitization. Neuron. 1998;21:727–739. doi: 10.1016/s0896-6273(00)80590-5. [DOI] [PubMed] [Google Scholar]
- Goldin M, Epsztein J, Jorquera I, Represa A, Ben-Ari Y, Crepel V, Cossart R. Synaptic kainate receptors tune oriens-lacunosum moleculare interneurons to operate at theta frequency. J Neurosci. 2007;27:9560–9572. doi: 10.1523/JNEUROSCI.1237-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hollmann M, Heinemann S. Cloned glutamate receptors. Annu Rev Neurosci. 1994;17:31–108. doi: 10.1146/annurev.ne.17.030194.000335. [DOI] [PubMed] [Google Scholar]
- Huang YH, Dykes-Hoberg M, Tanaka K, Rothstein JD, Bergles DE. Climbing fiber activation of EAAT4 transporters and kainate receptors in cerebellar Purkinje cells. J Neurosci. 2004;24:103–111. doi: 10.1523/JNEUROSCI.4473-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Isaac JT, Mellor J, Hurtado D, Roche KW. Kainate receptor trafficking: physiological roles and molecular mechanisms. Pharmacol Ther. 2004;104:163–172. doi: 10.1016/j.pharmthera.2004.08.006. [DOI] [PubMed] [Google Scholar]
- Jackson AC, Nicoll RA. The expanding social network of ionotropic glutamate receptors: TARPs and other transmembrane auxiliary subunits. Neuron. 2011;70:178–199. doi: 10.1016/j.neuron.2011.04.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jaskolski F, Coussen F, Mulle C. Subcellular localization and trafficking of kainate receptors. Trends Pharmacol Sci. 2005;26:20–26. doi: 10.1016/j.tips.2004.11.008. [DOI] [PubMed] [Google Scholar]
- Johnston GA. Convulsions induced in 10-day-old rats by intraperitional injection of monosodium glutamate and related excitant amino acids. Biochem Pharmacol. 1973;22:137–140. doi: 10.1016/0006-2952(73)90267-0. [DOI] [PubMed] [Google Scholar]
- Kato AS, Gill MB, Yu H, Nisenbaum ES, Bredt DS. TARPs differentially decorate AMPA receptors to specify neuropharmacology. Trends Neurosci. 2010;33:241–248. doi: 10.1016/j.tins.2010.02.004. [DOI] [PubMed] [Google Scholar]
- Kidd FL, Isaac JT. Developmental and activity-dependent regulation of kainate receptors at thalamocortical synapses. Nature. 1999;400:569–573. doi: 10.1038/23040. [DOI] [PubMed] [Google Scholar]
- Laezza F, Wilding TJ, Sequeira S, Coussen F, Zhang XZ, Hill-Robinson R, Mulle C, Huettner JE, Craig AM. KRIP6: a novel BTB/kelch protein regulating function of kainate receptors. Mol Cell Neurosci. 2007;34:539–550. doi: 10.1016/j.mcn.2006.12.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lerma J. Kainate receptor physiology. Curr Opin Pharmacol. 2006;6:89–97. doi: 10.1016/j.coph.2005.08.004. [DOI] [PubMed] [Google Scholar]
- Lerma J. Net(o) excitement for kainate receptors. Nat Neurosci. 2011;14:808–810. doi: 10.1038/nn.2864. [DOI] [PubMed] [Google Scholar]
- Li H, Rogawski MA. GluR5 kainate receptor mediated synaptic transmission in rat basolateral amygdala in vitro. Neuropharmacology. 1998;37:1279–1286. doi: 10.1016/s0028-3908(98)00109-9. [DOI] [PubMed] [Google Scholar]
- Li P, Wilding TJ, Kim SJ, Calejesan AA, Huettner JE, Zhuo M. Kainate-receptor-mediated sensory synaptic transmission in mammalian spinal cord. Nature. 1999;397:161–164. doi: 10.1038/16469. [DOI] [PubMed] [Google Scholar]
- Melyan Z, Wheal HV, Lancaster B. Metabotropic-mediated kainate receptor regulation of IsAHP and excitability in pyramidal cells. Neuron. 2002;34:107–114. doi: 10.1016/s0896-6273(02)00624-4. [DOI] [PubMed] [Google Scholar]
- Michishita M, Ikeda T, Nakashiba T, Ogawa M, Tashiro K, Honjo T, Doi K, Itohara S, Endo S. A novel gene, Btcl1, encoding CUB and LDLa domains is expressed in restricted areas of mouse brain. Biochem Biophys Res Commun. 2003;306:680–686. doi: 10.1016/s0006-291x(03)01035-0. [DOI] [PubMed] [Google Scholar]
- Michishita M, Ikeda T, Nakashiba T, Ogawa M, Tashiro K, Honjo T, Doi K, Itohara S, Endo S. Expression of Btcl2, a novel member of Btcl gene family, during development of the central nervous system. Brain Res Dev Brain Res. 2004;153:135–142. doi: 10.1016/j.devbrainres.2004.06.012. [DOI] [PubMed] [Google Scholar]
- Miyata M, Imoto K. Different composition of glutamate receptors in corticothalamic and lemniscal synaptic responses and their roles in the firing responses of ventrobasal thalamic neurons in juvenile mice. J Physiol. 2006;575:161–174. doi: 10.1113/jphysiol.2006.114413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Monaghan DT, Cotman CW. The distribution of [3H]kainic acid binding sites in rat CNS as determined by autoradiography. Brain Res. 1982;252:91–100. doi: 10.1016/0006-8993(82)90981-7. [DOI] [PubMed] [Google Scholar]
- Mulle C, Sailer A, Perez-Otano I, Dickinson-Anson H, Castillo PE, Bureau I, Maron C, Gage FH, Mann JR, Bettler B, Heinemann SF. Altered synaptic physiology and reduced susceptibility to kainate-induced seizures in GluR6-deficient mice. Nature. 1998;392:601–605. doi: 10.1038/33408. [DOI] [PubMed] [Google Scholar]
- Nadler JV, Perry BW, Cotman CW. Intraventricular kainic acid preferentially destroys hippocampal pyramidal cells. Nature. 1978;271:676–677. doi: 10.1038/271676a0. [DOI] [PubMed] [Google Scholar]
- Nasu-Nishimura Y, Hurtado D, Braud S, Tang TT, Isaac JT, Roche KW. Identification of an endoplasmic reticulum-retention motif in an intracellular loop of the kainate receptor subunit KA2. J Neurosci. 2006;26:7014–7021. doi: 10.1523/JNEUROSCI.0573-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ng D, Pitcher GM, Szilard RK, Sertie A, Kanisek M, Clapcote SJ, Lipina T, Kalia LV, Joo D, McKerlie C, Cortez M, Roder JC, Salter MW, McInnes RR. Neto1 is a novel CUB-domain NMDA receptor-interacting protein required for synaptic plasticity and learning. PLoS Biol. 2009;7:e41. doi: 10.1371/journal.pbio.1000041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Olney JW, Rhee V, Ho OL. Kainic acid: a powerful neurotoxic analogue of glutamate. Brain Res. 1974;77:507–512. doi: 10.1016/0006-8993(74)90640-4. [DOI] [PubMed] [Google Scholar]
- Opazo P, Choquet D. A three-step model for the synaptic recruitment of AMPA receptors. Mol Cell Neurosci. 2011;46:1–8. doi: 10.1016/j.mcn.2010.08.014. [DOI] [PubMed] [Google Scholar]
- Pinheiro P, Mulle C. Kainate receptors. Cell Tissue Res. 2006;326:457–482. doi: 10.1007/s00441-006-0265-6. [DOI] [PubMed] [Google Scholar]
- Rodriguez-Moreno A, Herreras O, Lerma J. Kainate receptors presynaptically downregulate GABAergic inhibition in the rat hippocampus. Neuron. 1997;19:893–901. doi: 10.1016/s0896-6273(00)80970-8. [DOI] [PubMed] [Google Scholar]
- Ruiz A, Sachidhanandam S, Utvik JK, Coussen F, Mulle C. Distinct subunits in heteromeric kainate receptors mediate ionotropic and metabotropic function at hippocampal mossy fiber synapses. J Neurosci. 2005;25:11710–11718. doi: 10.1523/JNEUROSCI.4041-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sachidhanandam S, Blanchet C, Jeantet Y, Cho YH, Mulle C. Kainate receptors act as conditional amplifiers of spike transmission at hippocampal mossy fiber synapses. J Neurosci. 2009;29:5000–5008. doi: 10.1523/JNEUROSCI.5807-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Straub C, Hunt DL, Yamasaki M, Kim KS, Watanabe M, Castillo PE, Tomita S. Distinct functions of kainate receptors in the brain are determined by the auxiliary subunit Neto1. Nat Neurosci. 2011a;14:866–873. doi: 10.1038/nn.2837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Straub C, Tomita S. The regulation of glutamate receptor trafficking and function by TARPs and other transmembrane auxiliary subunits. Curr Opin Neurobiol. 2012 doi: 10.1016/j.conb.2011.09.005. (in press) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Straub C, Zhang W, Howe JR. Neto2 modulation of kainate receptors with different subunit compositions. J Neurosci. 2011b;31:8078–8082. doi: 10.1523/JNEUROSCI.0024-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tang M, Pelkey KA, Ng D, Ivakine E, McBain CJ, Salter MW, McInnes RR. Neto1 is an auxiliary subunit of native synaptic kainate receptors. J Neurosci. 2011;31:10009–10018. doi: 10.1523/JNEUROSCI.6617-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Traynelis SF, Wollmuth LP, McBain CJ, Menniti FS, Vance KM, Ogden KK, Hansen KB, Yuan H, Myers SJ, Dingledine R, Sibley D. Glutamate receptor ion channels: structure, regulation, and function. Pharmacol Rev. 2010;62:405–496. doi: 10.1124/pr.109.002451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vignes M, Collingridge GL. The synaptic activation of kainate receptors. Nature. 1997;388:179–182. doi: 10.1038/40639. [DOI] [PubMed] [Google Scholar]
- Vincent P, Mulle C. Kainate receptors in epilepsy and excitotoxicity. Neuroscience. 2009;158:309–323. doi: 10.1016/j.neuroscience.2008.02.066. [DOI] [PubMed] [Google Scholar]
- Wisden W, Seeburg PH. Mammalian ionotropic glutamate receptors. Curr Opin Neurobiol. 1993;3:291–298. doi: 10.1016/0959-4388(93)90120-n. [DOI] [PubMed] [Google Scholar]
- Wu LJ, Ko SW, Zhuo M. Kainate receptors and pain: from dorsal root ganglion to the anterior cingulate cortex. Curr Pharm Des. 2007;13:1597–1605. doi: 10.2174/138161207780765864. [DOI] [PubMed] [Google Scholar]
- Wu LJ, Zhao MG, Toyoda H, Ko SW, Zhuo M. Kainate receptor-mediated synaptic transmission in the adult anterior cingulate cortex. J Neurophysiol. 2005;94:1805–1813. doi: 10.1152/jn.00091.2005. [DOI] [PubMed] [Google Scholar]
- Yan D, Tomita S. Defined criteria for auxiliary subunits of glutamate receptors. J Physiol. 2012;590:21–31. doi: 10.1113/jphysiol.2011.213868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang W, St-Gelais F, Grabner CP, Trinidad JC, Sumioka A, Morimoto-Tomita M, Kim KS, Straub C, Burlingame AL, Howe JR, Tomita S. A transmembrane accessory subunit that modulates kainate-type glutamate receptors. Neuron. 2009;61:385–396. doi: 10.1016/j.neuron.2008.12.014. [DOI] [PMC free article] [PubMed] [Google Scholar]