

INTRODUCTION: CHEMOKINES AND CHEMOKINE RECEPTORS
Chemokines are small 70 amino acid long soluble proteins that chemoattract a variety of mononuclear cell types to sites of inflammation or secondary lymphoid organs by interacting with chemokine receptors. 1 They are divided into four subfamilies of CC, CXC, CX3C, and C chemokines based on the positions of two conserved cysteine residues in their amino (N)-termini. 2,3 The CXC chemokine receptor 4 (CXCR4) is an attractive target for therapeutic intervention because of its involvement in the pathogeneses of a wide range of infectious, inflammatory, and other diseases. CXCR4 is a transmembrane (TM) protein that belongs to the superfamily of G-protein-coupled receptors (GPCRs). 2,4,5 It possesses seven TM helices and transmits signals from its natural chemokine ligand, stromal cell-derived factor (SDF)-1α/CXCL12, to intracellular biological pathways via heterotrimeric G-proteins.
The activation of CXCR4 by SDF-1α can trigger different downstream signaling pathways that result in a variety of physiological responses, such as chemotaxis, cell survival and proliferation, intracellular calcium flux, and gene transcription (Fig. 1). 6-15 These normal physiological responses also share several downstream effectors with multiple pathological processes, including tumor cell metastasis, and autoimmune and inflammatory diseases. For instance, CXCR-mediated chemotaxis and cell survival involves PI3 kinase (PI3K) which also plays a major role in cancer cell survival, proliferation, and metastasis. 10 Whereas cancer cell proliferation requires the activation of Akt (serine/threonine protein kinase) via the PI3K pathway, physiologically occurring cell survival can activate Bcl-2-associated death promoter (BAD) via both MEK (MAP kinase kinase) and PI3K pathways, which leads to the inhibition of the proapoptotic protein Bcl-2. 6 Similarly, although Janus kinase (JAK)/Signal Transducer and Activator of Transcription (STAT) pathway allows a G-protein independent signaling pathway via CXCR4, the receptor phosphorylation by JAK2 and JAK3 leads to the activation and nuclear translocation of a variety of STAT proteins, which leads to cancer cell survival and proliferation.16
Figure 1.

CXCR4 intracellular signaling pathways. CXCR4 activation by SDF-1α can trigger a variety of physiological responses, such as chemotaxis, cell survival and proliferation, intracellular calcium flux, and gene transcription, whereas CXCR4 antagonists fail to do so. These normal physiological responses also share several downstream effectors with multiple pathological processes, including tumor cell metastasis, HIV-associated dementia (induced by HIV-1 gp120), and autoimmune and inflammatory diseases. 6-15
The structures of multiple chemokines have been determined by NMR or X-ray crystallography, including those of SDF-1α, 17,18 viral macrophage inflammatory protein (vMIP)-II, 19,20 macrophage inflammatory protein (MIP)-1β, 21 and ‘regulated on activation, normal T-cell expressed and secreted’ (RANTES). 22 These structures demonstrate the highly conserved three-dimensional structures of all chemokines, including a flexible N-terminus, a three-stranded anti-parallel β-sheet, and a C-terminal α-helix. 23 In the typical structure, the first two cysteine residues are situated close together near the amino (N)-terminus, with the third cysteine residue residing in the center of the molecule, and the fourth cysteine residue located close to the carboxyl (C)-terminal end. 24 An “N-loop” of approximately ten amino acids follows the first two cysteine residues. Following the N-loop, there is a single-turn “310-helix,” a β-sheet with three β-strands, and a C-terminal α-helix, connected by turns called “30s,” “40s,” and “50s” loops. The third and fourth cysteine residues are located in the 30s and 50s loops, respectively.
Due to its involvement in a wide range of physiological and pathologic processes, there has been intensive biological, chemical, and pharmaceutical research to understand the molecular mechanisms of chemokine–receptor interactions and the modulation of chemokine–receptor functions. The ultimate goal is to translate these discoveries into novel treatment strategies for clinical applications. This review describes and discusses some of the recent advances in medicinal chemistry and drug discovery that involve CXCR4, which is implicated in human immunodeficiency virus (HIV)-1 infection, normal hematopoietic and neural stem cell migration, cancer–stromal cell interaction, solid tumors, and inflammation and autoimmune diseases such as rheumatoid arthritis and allergic asthma.
CXCR4 ANTAGONISTS AGAINST HIV-1 ENTRY
HIV-1 enters target cells through a fusion process in which the HIV-1 gp120 envelope glycoprotein binds to CD4, the main receptor for HIV-1 on the target cell surface. 25-28 However, CD4 alone is not sufficient for HIV-1 fusion, and the chemokine receptors CXCR4 and CCR5 act as coreceptors for syncytium-inducing and non-syncytium-inducing HIV-1 strains, respectively (Fig. 2). The initial binding of HIV-1 gp120 to CD4 results in conformational changes in gp120 and CD4. 29-31 The gp120–CD4 complex then interacts with a chemokine coreceptor such as CXCR4 or CCR5 to form a heterotrimeric complex of gp120–CD4–coreceptor. 32-34 During the asymptomatic stage of disease, macrophage (M)-tropic strains of HIV-1 (also known as R5-tropic) primarily use CCR5 as the entry coreceptor. 26-28 However, in 40–50% of HIV-1-infected individuals, T-cell (T)-tropic strains (also known as X4-tropic), which predominantly use CXCR4, eventually replace M-tropic strains, resulting in rapid disease progression. 35-37 Dualtropic strains are those HIV-1 isolates that are capable of using both CXCR4 and CCR5. CXCR4-using variants are believed to evolve from CCR5-using variants by way of transitional intermediate variants. Natural chemokines that bind CXCR4 or CCR5 can inhibit HIV-1 infection, 38,39 most likely by blocking common binding sites on the chemokine receptors that are also required for gp120 interaction, and/or by inducing receptor internalization. 40,41
Figure 2.

Representative CXCR4 interactions with several different classes of CXCR4 ligands, including SDF-1α (natural agonist), vMIP-II (natural antagonist), HIV-1 gp120 (viral ligand), AMD3100 (small synthetic organic molecule), and DV1/9 (modified peptides). Orange arrows represent disease and drug related pathways, and blue arrows indicate physiologic functions of CXCR4.
Viral resistance to CCR5 inhibitors is well documented. Most often, it involves selection of pre-existing minor HIV-1 variants that can use CXCR4 for entry. 42-44 Also selection of mutations that change the coreceptor use from CCR5 to CXCR4 has been demonstrated in vitro. 45 More recently, Mosier’s group has demonstrated that coreceptor switching may be the only route to resistance for macromolecular CCR5 inhibitors, and that this pathway requires more mutations and encounters more fitness obstacles than development of resistance to small molecule CCR5 inhibitors. 46 Although the use of CXCR4 inhibitors may not have a wide window of opportunity for the treatment of HIV-infected individuals in comparison to CCR5 inhibitors because the X4-tropic virus appears at the late stage of HIV infection, the development of CXCR4 inhibitors is still urgently needed to overcome the viral drug resistance against CCR5 inhibitors and to reduce the occurrence of transitional intermediate variants that can successfully switch to CXCR4-using variants.
The discovery of distinct chemokine coreceptors for different HIV-1 strains has important clinical implications for developing new therapeutic strategies, and it has presented many new directions for Acquired Immune Deficiency Syndrome (AIDS) research. One of these new important areas of research is the biochemical and biophysical characterization of the interactions of chemokine receptors with HIV-1, natural ligands, and de novo designed inhibitors.47-55 A combined approach that integrates structural biology, molecular biology, and synthetic chemistry has been used to define precise regions involved in the molecular interactions of chemokine receptors with their natural ligands and with HIV-1 gp120. More recently, crystal structures of CXCR4 bound to an antagonist small molecule (IT1t) and to a cyclic peptide (CVX15) have been reported at a resolution of 2.5 to 3.2 angstroms. 56 Compared with previous GPCR structures, these recent structures indicate a more open and larger binding pocket of CXCR4 that is located closer to the extracellular surface. Taken together, these studies have provided structural and chemical insights into receptor–ligand interactions and a foundation for the design of therapeutic agents that can prevent HIV-1 entry.
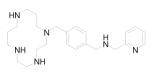
When compared to other anti-HIV-1 agents targeting components of the rapidly mutating virus population (such as reverse transcriptase and protease inhibitors), strategies based on HIV-1 coreceptors, CXCR4 and CCR5, have appeal because they target invariant host determinants.40,53,57-69 The fact that individuals with CCR5 mutations are highly resistant to HIV-1 infection also demonstrates the feasibility of this approach for inhibiting actual viral infection, 3,70 although it has been recently found that CCR5 deficiency is a potential risk factor for both early and late clinical manifestations after West Nile Virus infection. 71 Early chemical screening and medicinal chemistry efforts identified several CXCR4 antagonistic peptides and organic compounds that are structurally unrelated to natural chemokines. 53,63-66,68 Among these, N-α-acetyl-nona-D-arginine amide (ALX40-4C) and plerixafor (AMD3100) were the first CXCR4 antagonists to enter clinical trials (Table 1). 53,72,73 1 (ALX40-4C) is a highly basic peptide designed originally as an arginine mimic of the HIV Tat protein. It inhibits HIV-1 infection exclusively by blocking direct virus-CXCR4 interactions. 72,73 With a net charge of +9, 1 can bind to the negatively charged domains of CXCR4, where it competes with the V3 loop of HIV-1 gp120 to prevent virus entry. 53,72 AMD3100 is a bicyclam containing two cyclam rings connected by a p-phenylenebis(methylene) linker. It is a highly potent and selective anti-HIV-1 inhibitor that acts by binding to CXCR4 (Fig. 2). 65 Mutation of Asp171 or Asp262 (located in TM domains IV and VI of CXCR4, respectively) to alanine also suggested that the negatively charged aspartate residues at positions 171 and 262 of CXCR4 may represent crucial sites for electrostatic interaction of the positively charged bicyclam rings.74 Similarly, molecular modeling suggested that one cyclam ring of AMD3100 interacts with Asp171 in the TM domain IV of CXCR4, whereas the other ring is sandwiched between the carboxylate groups of Asp262 and Glu288 from TM helices VI and VII, respectively. 75 More recently, using homology modeling and molecular docking, Kawatkar et al demonstrated that several small CXCR4 antagonists, including AMD3100, have a distinct binding site that is different from the region occupied by the N-terminus of SDF-1α. This suggested the co-existence of an antagonistic ligand-binding site with a SDF-1α signaling site in CXCR4. 76
Table 1.
List of CXCR4 Inhibitors
| Name of CXCR4 Inhibitors or Analog Designation |
Molecular Type | Amino Acid Sequences, Modification Diagrams, or Chemical Structures |
Biological Activities | Stage of Development |
References |
|---|---|---|---|---|---|
| 1 | d-Peptide | Ac-NH-(d-Arg)9-COOH | Block HIV-1 entry | Phage I | 53,72,73 |
| AMD3100 | Small Orgnic Molecule |
|
(1) Block HIV-1 entry; (2) promote stem cell migration; (3) inhibit stromal–leukemia cell interactions; and (4) anti-RA and allergic asthma activities |
Approved in 2008 for patients with lymphoma and multiple myeloma as a source of hematopoietic stem cells for transplantation |
57, 200,111, 117,113,125 |
| 2 | Small Orgnic Molecule |
|
CXCR4 Antagonist | Pre-clinical | 77 |
| AMD3465 | Small Orgnic Molecule |
|
Block HIV-1 entry | Pre-clinical | 78 |
| 3 | Small Orgnic Molecule |
|
Block HIV-1 entry | Phase I | 80 |
| 4 | Small Orgnic Molecule |
|
Block HIV-1 entry | Phase I | 84 |
| 5 | Small Orgnic Molecule |
|
Block HIV-1 entry | Pre-clinical | 85,86 |
| 6 | Small Orgnic Molecule |
|
Block HIV-1 entry | Pre-clinical | 87 |
| T22 | Peptide |

|
(1) Block HIV-1 entry; and (2) anti-RA activities |
Pre-clinical | 58 |
| T140 | Peptide |
|
(1) Block HIV-1 entry; (2) promote stem cell migration; and (3) anti-RA activities |
Pre-clinical | 88,89,201 |
| TC14012 | Peptide |
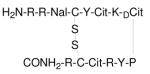
|
Block HIV-1 entry | Pre-clinical | 88 |
| 7 | Peptide |
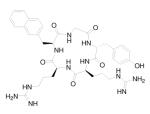
|
(1) Block HIV-1 entry; and (2) anti-RA activities |
Pre-clinical | 89 |
| 8 | Small Orgnic Molecule |
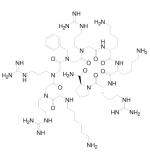
|
Block HIV-1 entry | Pre-clinical | 93 |
| R3G | Small Orgnic Molecule |
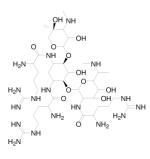
|
Block HIV-1 entry | Pre-clinical | 203 |
| SDF-1α | Peptide |
KPVSLSYRCPCRFFESHVARANVK HLKILNTPNCALQIVARLKNNNRQ VCIDPKLKWIQEYLEKALNK |
Block HIV-1 entry | Pre-clinical | 59,60,67,69 |
| CTCE-9908 | Peptide | KGVSLSYRKRYSLSVGK | Inhibit cancer growth and metastases |
Phase II | 128-130 |
| vMIP-II | Peptide |
LGASWHRPDKCCLGYQKRPLPQV LLSSWYPTSQLCSKPGVIFLTKRGR QVCADKSKDWVKKLMQQLPVTA R |
Block HIV-1 entry | Pre-clinical | 19,20 |
| V1 | Peptide |
 LGASWHRPDKCCLGYQ KRPLP |
Block HIV-1 entry | Pre-clinical | 61 |
| DV1 | d-Peptide |
 LGASWHRPDKCCLGYQ KRPLP |
Block HIV-1 entry | Pre-clinical | 64 |
| 9 |
d-Amino Acid Containing Peptide |
LGASWHRPDKCCLGYQKRPLPQVL LSSWYPTSQLCSKPGVIFLTKRGR QVCADKSKDWVKKLMQQLPVTA R |
(1) Block HIV-1 entry; and (2) inhibit stromal–leukemia cell interactions |
Pre-clinical | 98,99,124 |
D-amino acids are shown in italic.
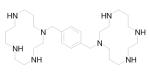
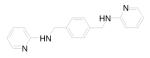
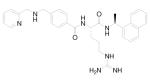
Based on this proposed molecular mechanism for AMD3100, Zhan et al designed and synthesized a series of candidate compounds by replacing the cyclam moieties of AMD3100 with N-containing basic centers that are hypothesized to bind to the acidic residues in CXCR4. 77 The synthesis of a series of small molecular CXCR4 antagonists with two aromatic amine moieties connected by a para-xylylene group ultimately led to the discovery of N1,N4-di-2-pyridinyl-1,4-benzenedimethanamine (WZ811), a potent compound that can block CXCR4 at sub-nanomolar concentrations (Table 1). 2 (WZ811) was also more effective than AMD3100 in inhibiting CXCR4/SDF-1α-mediated modulation of cyclic adenosine monophosphate (cAMP) levels and SDF-1α-induced Matrigel invasion. However, clinical applications of AMD3100 and its derivatives are limited due to their lack of oral bioavailability, which is related to their high positive charge at physiological pH. To improve these characteristics, analogous compounds with fewer basic amine groups have been developed, including monocyclam derivatives (AMD3465) 78,79 and non-cyclam derivatives (AMD070) 80 (Table 1). AMD3465, a monocyclam with a pyridinylmethylene amine moiety, is comparable to AMD3100 in its antiviral activity, and it is capable of inhibiting the replication of virus strains that are resistant to other anti-HIV drugs such as zidovudine. 78 It targets the side-chains of Asp171 and Asp262 as well as Glu288 on CXCR4. It also binds to His281 of the target protein. 3 (AMD070) is a potent orally available CXCR4 antagonist that strongly inhibits virus infectivity via CXCR4. 80,81 It works in an additive or synergistic fashion when combined with other HIV inhibitors such as reverse transcriptase and protease inhibitors. More recently, Schols’ group further modified 3 by generating novel heterocyclic analogs of 3 82 and also by (1) sequentially replacing the benzimidazole ring with a substituted pyridine ring; (2) opening up the tetrahydroquinoline ring; and (3) combining both of these modifications to afford open chain analogs (4), while still maintaining potent HIV-1 inhibition. 83
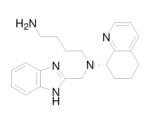
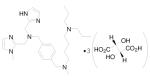
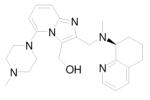
Oral availability still remains one of the key limitations to be overcome for these compounds to be effective as anti-HIV drugs, because retroviral infection continues throughout the life of the infected person. More recent advances in this area of research have led to the development of two additional orally active CXCR4 antagonists. N-{(S)-4-guanidino-1-[(S)-1-naphthalen-1-yl-ethylcarbamoyl]butyl}-4-{[(pyridin-2-yl-methyl)amino]methyl}benzamide (KRH-1636, Table 1) is a potent small nonpeptide CXCR4 inhibitor derived from a small cyclopentapeptide, cyclo(-Nal-Gly-D-Tyr-Arg-Arg-) (FC131). 84 Although this compound was efficiently absorbed from the rat duodenum, it could not be further developed as an anti-HIV-1 agent because of its poor oral bioavailability. Continuous efforts to find more potent CXCR4 antagonists that are bioavailable when administered orally led to the development of KRH-3955 (Table 1), a 4 (KRH-1636) derivative, which has a much more potent anti-HIV-1 activity than either AMD3100 or 4. 85,86 5 (KRH-3955) showed oral bioavailability of 25.6% in rats, and its oral administration blocked T-tropic HIV-1 replication in human peripheral blood lymphocytes and in a severe combined immunodeficiency mouse model. It was also active against recombinant T-tropic HIV-1 containing resistant mutations in reverse transcriptase, protease, and envelope with enfuvirtide resistance mutations. Experiments that examined the effects of 5 on the binding of several anti-CXCR4 monoclonal antibodies (MAbs) suggested that the binding sites of 5 are located in all three extracellular domains of CXCR4. Similarly, Jenkinson et al recently reported an orally bioavailable and highly selective CXCR4 antagonist, ([5-(4-methyl-1-piperazinyl)-2-({methyl[(8S)-5,6,7,8-tetrahydro-8-quinolinyl]amino}methyl)imidazo[1,2-a]pyridin-3-yl]methanol) (GSK812397, Table 1), that was derived from a chemical class distinct from either 3 or 5. 87 The pharmacokinetic properties of 6 (GSK812397) compare favorably with those of 5, as its bioavailability was found to be similar across several species, including rat (21%), dog (21%), and monkey (17%).
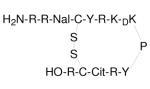
An 18 amino acid synthetic polyphemusin (T22) 58 and its downsized analogs (T140 and TC14012 (14 amino acid residues)) also inhibit CXCR4-mediated HIV-1 entry 88 (Table 1). Based on the finding that the critical pharmacophore of T140 is comprised of Arg2, L-3-(2-naphthyl)alanine (Nal)3, Tyr5, and Arg14, screening of pentapeptide libraries containing these critical residues resulted in the identification of the cyclic pentapeptide 7 (FC131), that has similar anti-HIV activity to T140 (Table 1). 89 More recently, smaller cyclic peptides based on the structure of T140 90,91 as well as nonpeptide analogs 92 have been reported. The nonpeptide analogs have 50% inhibitory concentrations that are at least two orders of magnitude higher than that of T140. Other studies have reported a hybrid peptoid/peptide oligomer of nine residues (CGP64222), tri-arginine derivative of gentamicin C (R3G), and neomycin B-hexaarginine conjugate (NeoR) as Arg-mimic and cationic CXCR4 antagonists 93 (Table 1). 8 (CGP64222), a basic peptide oligomer of nine residues, inhibits the replication of a wide range of laboratory HIV strains through selective interaction with CXCR4. It specifically binds to the second extracellular loop (ECL2) of CXCR4, and it retains its full activity against the azidothymidine (AZT)-resistant HIV-1 strain. 94
An alternative route for designing specific CXCR4 inhibitors is to use natural chemokine ligands as design templates and to mimic specific regions of these chemokine ligands. 59-64,67-69,95 First, using SDF-1α sequence as the design template 59,60,67,69 (Table 1), a series of peptides derived from the N-terminus of SDF-1α were generated. The studies of these peptides demonstrated that the N-terminus of SDF-1α is essential for CXCR4 recognition, signal transduction, and antiviral activity. However, these peptides were less potent than the native SDF-1α. 67,69 Noting the earlier observation that peptides and organic compound inhibitors of CXCR4, including 1, 53 AMD3100, 57 and T22, 58 have highly positive charges, the attachment of positively charged residues to the N-terminal peptide sequence of SDF-1α enhanced its ability to bind to CXCR4. 59
CXCR4 can also be recognized by vMIP-II which recognizes a variety of CC and CXC chemokine receptors, including CXCR4, CCR5 and CCR2 96,97 (Table 1; Fig. 2). Studies of synthetic peptides derived from the N-terminus of vMIP-II demonstrated that the N-terminus of vMIP-II is the major determinant for CXCR4 recognition. 61 Only the 1-21 residue peptide of vMIP-II (V1) showed CXCR4 binding, and it selectively prevented CXCR4 signal transduction and its coreceptor function in mediating the entry of T-and dual-tropic HIV-1 isolates. Despite having a dramatically different conformation of side-chain groups, an all-D-amino acid analog (DV1) of V1 peptide displayed higher binding affinity for CXCR4 and significant antiviral activity as an inhibitor of the replication of CXCR4-dependent HIV-1 strains 64 (Fig. 2). The use of unnatural D-peptides can be advantageous for therapeutic development as such peptides are highly stable and resistant to proteolytic degradation.
In addition to short peptides derived from natural chemokines, a systematic strategy based on the full length chemokine sequences was developed for the synthesis of a new family of unnatural chemokines called SMM (synthetically and modularly modified)-chemokines that, unlike natural chemokines, have high receptor binding selectivity. 98 This strategy was applied successfully to transform vMIP-II, a very nonselective chemokine, into a series of new analogs with significantly enhanced selectivity and potency for CXCR4 or CCR5, through modification of only a small N-terminal module of 10 residues. Two representative SMM-chemokines, RCP168 and RCP188, selective for CXCR4 and CCR5, respectively, showed similar or significantly enhanced binding affinities for their corresponding target receptors, whereas they have decreased or even completely abolished cross-binding activities for other receptors (Table 1). 9 (RCP168) was also more effective in inhibiting HIV-1 infection than SDF-1α, and its anti-HIV activity was comparable to that of Fuzeon (T-20, also known as enfuvirtide), a currently marketed drug targeting HIV-1 entry (Fig. 2). More interestingly, 9 did not interfere with the Ca2+ signaling induced by SDF-1α at its effective CXCR4 binding concentration and only showed its anti-Ca2+ signaling effects at concentrations over 20 times higher than the CXCR4 binding concentration. Moreover, 9 significantly inhibited HIV-1 entry, in contrast to its much weaker activity in interfering with SDF-1α signaling. Thus, these disparate inhibitory activity profiles of 9 in differentiating HIV-1 coreceptor function versus the normal function of CXCR4 may prove to be advantageous in clinical applications, as 9 may not induce unwanted Ca2+ signaling or interfere with SDF-1α signaling important for the normal physiological functions at the concentrations used for inhibiting HIV-1 infection. The mechanistic basis for the disparate activities of 9 was further demonstrated by mutational mapping analysis of the binding sites of 9, which revealed that 9 binding sites on CXCR4 overlap significantly with HIV-1 but differ from SDF-1α. 99 These novel and potent CXCR4 peptide antagonists will serve as leads for the development of new therapeutic agents for HIV-1 infection and other diseases affecting the immune system.
It remains to be further studied whether the blocking of the normal CXCR4–SDF-1α functions in HIV-1 therapy may cause potential undesirable side effects, given that CXCR4 100,101 or SDF-1α 102 knockout mice die during embryogenesis with evidence of hematopoietic, cardiac, vascular, and cerebellar defects, 103 which indicates the essential role of CXCR4 in development. However, it should be noted that since CXCR4 antagonists are given to adults, not neonates, it is unlikely that these potential side effects will be a clinical issue, unless given to pregnant women. To avoid affecting the normal function of CXCR4, Sachpatzidis et al took the approach of allosteric agonists and identified two CXCR4 peptide ligands (RSVM and ASLW) that are insensitive to the CXCR4 antagonists, AMD3100 and T140, or the monoclonal antibodies, 12G5 and 44717.111. This was achieved by screening a semi-randomized 17-mer library in a yeast strain that expressed a functional CXCR4 receptor. 68 In chemotaxis assays, 10 (RSVM) behaves as a partial agonist, while 11 (ASLW) behaves as a superagonist that displays a chemotactic index greater than the maximum observed in SDF-1α. Allosteric agonists that have different binding sites from CXCR4 antagonists may be therapeutically useful when combined with small molecule antagonists for anti-HIV therapy, since they could allow maintenance of essential receptor functions. The data also illustrated that other binding sites may exist for non-physiological agonists.
CXCR4 ANTAGONISTS AND AGONISTS THAT MODULATE STEM CELL AND CANCER CELL MIGRATION AND THEIR POTENTIAL APPLICATIONS IN THE TREATMENT OF HEMATOPOIETIC AND SOLID CANCERS
Adult bone marrow (BM) is the primary source of hematopoietic stem cells (HSCs) that initiate hematopoiesis. 104 The elevation of intermediates of HSCs, namely hematopoietic progenitor cells (HPCs), in blood of patients recovering from chemotherapy or radiation 105,106 led to the realization that the HSCs and HPCs are able to exit the bone marrow, and that these mobilized cells could be collected and would re-home to marrow and restore hematopoiesis. Currently, mobilized peripheral blood is the primary source of hematopoietic cells used for hematopoietic transplantation. However, HPCs and HSCs circulate in blood at frequencies too low to allow efficient collection of sufficient numbers for transplantation. Therefore, an increase in the number of HSCs in blood following mobilization would improve the efficiency of transplantation and would have the potential to shorten recovery from cytopenia, to enhance immune reconstitution, and to reduce morbidity and mortality.
Granulocyte colony-stimulating factor (G-CSF) is the predominant mobilizer of hematopoietic cells used clinically, but it results in a broad individual variability in mobilization responsiveness, 107 frequently requiring repeated dosing, often with side effects. Although the longer acting G-CSF, Pegfilgrastim, may address the multi-dosing requirement of conventional G-CSF, improved methods to mobilize and collect HSCs and HPCs for hematopoietic rescue are still warranted. Recent studies have indicated that the interactions between CXCR4 and SDF-1α modulate hematopoietic cell mobilization and engraftment. 108,109 CXCR4 is extensively expressed in HSCs and HPCs, and its interaction with SDF-1α promotes the retention of CXCR4 expressing hematopoietic cells in the BM microenvironment (Fig. 2). Consequently, the disruption of CXCR4–SDF-1α interactions has been hypothesized to result in the mobilization of hematopoietic cells. 110 Indeed, the blockade of CXCR4 with an antagonist, such as AMD3100 111 or 4F-benzoyl-TN14003 (T140), 112 results in the mobilization of hematopoietic cells (Table 1; Fig. 2). AMD 3100 (also known as Mozobil (plerixafor)) was finally approved in 2008 for patients with lymphoma, non-Hodgkin’s lymphoma (NHL), multiple myeloma as a source of HSCs for transplantation. 113-115
Another approach involves the use of the lipopeptide GPCR modulator, called pepducin, which is composed of a peptide motif derived from the amino acid sequence of one of the intracellular (IC) loops of a target GPCR coupled to a lipid tether. 116 Acting as a CXCR4 functional antagonist when administered systemically by I.V. bolus, ATI-2341 was identified as a potent mobilizer of bone marrow polymorphonuclear neutrophils (PMNs), HSCs, and HPCs. The successful generation of inhibitory CXCR4 nanobodies (VHH-based single variable domains) has also been reported, and these can induce HSC mobilization as rapidly and as nearly as effectively as AMD3100. 117 More recently, Polyphor (POL)6326, a selective and reversible CXCR4 inhibitor designed based on Polyphor’s proprietary Protein Epitope Mimetics (PEM) Technology, has been reported to induce HSC mobilization as well. 118,119 In fact, 12 (POL6326) has successfully completed Phase I clinical trials and is currently being investigated as a stand-alone therapy in a Phase II clinical trial for its efficacy in autologous transplantation of HSCs in multiple myeloma patients. Taken together, these results suggest that antagonizing the interactions of SDF-1α produced by BM stromal cells with CXCR4 expressed on hematopoietic cells could be an effective strategy for mobilizing HSC.
In addition to mediating the release and homing of CXCR4 expressing hematopoietic cells from and to the SDF-1α-producing BM microenvironment, CXCR4–SDF-1α interaction has also emerged as a critical mediator of stromal–leukemia cell interactions (Fig. 2). CXCR4–SDF-1α interaction not only protects chronic lymphocytic leukemia (CLL) cells from apoptosis, 120 but also allows the spontaneous migration of malignant cells beneath bone marrow stromal cells. This suggests that CLL cells might use this mechanism to infiltrate the marrow. 121 In fact, the BM microenvironment is known to provide a sanctuary for subpopulations of acute myelogenous leukemia (AML), 122 chronic lymphocytic leukemia (CLL), 120 and acute lymphoblastic leukemia (ALL) 123 to evade chemotherapy-induced apoptosis and acquire a drug-resistant phonotype. As such, CXCR4 inhibition has been hypothesized to interfere with stromal–leukemia cell interactions, thereby increasing the sensitivity of CLL, AML, or ALL cells to chemotherapy. Indeed, inhibition of CXCR4 by 9, a potent CXCR4 antagonist, enhanced the sensitivity of CLL and AML cells to chemotherapy by reducing the protection conferred by stromal cells 124 (Table 1; Fig. 2). Similar results were obtained with small molecule CXCR4 inhibitors, such as AMD3100 125 and AMD3465, 124 further suggesting that CXCR4–SDF-1α interaction contributes to the resistance of leukemia cells to chemotherapy-induced apoptosis. Thus, disruption of this interaction by CXCR4 inhibitors represents a novel strategy for targeting leukemia cells within the BM microenvironment.
Besides hematopoietic cell lines, CXCR4 is widely expressed on multiple solid tumors, including breast, lung, colon, ovarian, bone, brain, and skin cancer cells. The CXCR4–SDF-1α axis is involved in cancer cell progression, angiogenesis, metastasis, and survival. For instance, Zlotnik’s group has demonstrated that CXCR4 is highly expressed in human breast cancer cells, malignant breast tumors, and metastasis, and that malignant melanoma, which has a similar metastatic pattern as breast cancer, also shows high expression level of CXCR4. 126 In fact, the over-expression of CXCR4/SDF-1α in breast cancer has been correlated with increased metastasis to brain, lymph node, lung, liver, and bone marrow. 127 In this regard, Chemokine Therapeutics Corp (CTC)E-9908, a small peptide analog derived from the N-terminus of SDF-1α, emerged as a new generation of anti-cancer drug candidates (Table 1), as it was found to inhibit cancer growth and metastasis in several animal models by targeting CXCR4 that is present on most of these solid tumors. 128-130
Similar to hematopoietic cells, neural stem cells (NSCs) also have the capacity to migrate; in this case, into distant areas of neurodegeneration. This has been shown in a number of experimental models of central nervous system (CNS) disease, including at ages where extensive migration has conventionally been deemed to be limited. 131-135 Because stem cell engagement with a degenerating environment is the first critical step in regeneration, 95,135 understanding the mechanisms underlying stem cell mobilization during injury is crucial for the full realization of the therapeutic promise of NSCs. In this regard, CXCR4 expressing human NSCs (hNSCs) were previously shown to migrate in vivo toward an infarcted area (a representative CNS injury), where local astrocytes and endothelium upregulate SDF-1α 131-135 (Fig. 2). Exposure of hNSCs to SDF-1α and subsequent induction of CXCR4-mediated signaling triggers a series of intracellular processes that lead to the survival, proliferation, and most importantly, migration of neural stem cells for injury repair. These steps include rapid and sustained phosphorylation of p38MAPK kinase (implicated in the regulation of cytokine-induced cell migration 136-138); phosphorylation of the ribosomal S6 kinase (p90RSK, implicated in the phosphorylation of cytoskeletal molecules involved in neurite outgrowth 139,140); phosphorylation of c-Jun (a kinase involved in migratory responses 141); rapid activation of extracellular response kinase (involved in proliferative responses), and rapid phosphorylation of paxilin (a scaffold molecule critical for migration 142). As such, CXCR4–SDF-1α interaction implicated in the promotion of stem cell migration may serve as important targets for development of novel stem cell-based therapeutic for CNS diseases.
The extensive exploration of CXCR4–HIV-1 antagonism facilitated the investigation of highly potent and selective CXCR4 agonists capable of mediating NSC migration for injury repair. Recently, we developed a panel of “dual-moiety” chemokine analogs by linking a highly potent and selective CXCR4 binding moiety to a critical CXCR4-activating moiety of SDF-1α (unpublished results), based on the rationale that the newly created analogs may have signaling activities and capability to mobilize CXCR4-expressing hNSCs. Indeed, these “dual-moiety” chemokine analogs were found to promote the migration of NSCs in vitro and in vivo. As such, the effective mobilization of hNSCs with de novo designed agonists may lead to new translational therapeutics for the clinical repair of CNS injuries and other neurodegenerative conditions.
Since SDF-1α was identified as a potential stem cell homing factor, a large number of other studies have focused on CXCR4–SDF-1α interaction as a potential therapeutic target. For instance, many of these studies confirmed the significance of CXCR4–SDF-1α axis in mobilization of BM-derived stem cells to sites of myocardial infarction and ischemic injury to induce cardiac regeneration. 143,144 Different approaches have been described to modify CXCR4–SDF-1α interaction. An indirect approach by using G-CSF in a rabbit ischemic reperfusion model demonstrated that G-CSF can increase SDF-1α levels and induce rapid mobilization of CXCR4-positive cells towards infarcted areas, thus leading to decreased scar size.145 Also acute blockade of the interaction between CXCR4 and SDF-1α by a single dose of AMD3100 146 was found to induce a rapid and transient hematopoietic cell mobilization. In fact, short-term administration of AMD3100 showed significant improvement of blood restoration and angiogenesis during the acute phase of ischemia in mouse models. 147
CXCR4 ANTAGONISTS AGAINST INFLAMMATION AND AUTOIMMUNE DISEASES: RHEMATOID ARTHRITIS AND ALLERGIC ASTHMA
Rheumatoid arthritis (RA), an autoimmune disease affecting about 1% of the world population, is characterized by chronic inflammation of multiple joints, proliferation of the synovial cells, massive infiltration of leukocytes into inflamed joints, and destruction of the cartilage and bone of the affected joints. 148-150 A large number of CD4+ memory T-cells infiltrate the inflamed synovia, 151-157 and the pathogenesis of RA is then invoked by the action of inflammatory cytokines, including interleukin (IL)-1, IL-6, interferon (IFN)-γ, and tumor necrosis factor (TNF)-α and -β. 151,158 Accumulating evidence also suggests the involvement of CXCR4–SDF-1α interaction in RA pathogenesis. Nanki et al reported that CD4+ T-cells express high levels of CXCR4, and that the concentration of SDF-1α is also highly elevated in the synovia of RA patients, 152 suggesting that CXCR4 is important for T-cell retention in RA-affected synovial tissues (Fig. 2). This is further supported by the observation that T helper 1 (Th1) cells, which are believed to be at least partly involved in RA pathogenesis, are attracted by RA synovial fluid, and that chemotaxis can be inhibited in vitro by anti-SDF-1α antibody in vitro. 159 Moreover, SDF-1α inhibits T-cell apoptosis in addition to promoting the migration of memory T-cells. Taken together, these results indicate that the development of inhibitors that block CXCR4–SDF-1α interaction might lead to promising RA therapies. Indeed, several CXCR4 antagonists, including AMD3100, 160,161 T22 (an 18-mer peptide), 162 T140 (a 14-mer), 162 and 7 (a cyclic pentapeptide), 89,163 are known to have anti-RA activities (Table 1; Fig. 2).
CXCR4 is also associated with the pathophysiology of allergic asthma. Allergic asthma has a number of key characteristics, including airway inflammation of different cell types, airway hyper-responsiveness (AHR), mucus production, and variable airflow. 164,165 The initiating phase is characterized by the appearance of IgE and mast cells. In the propagation phase, Th2-polarized T-lymphocytes and eosinophils infiltrate the airways, initiating a chronic inflammatory state. The effector phase is characterized by the production of spasmogenic substances, AHR, and mucus hypersecretion. Steroid drugs have been the standard treatment, but development of resistance and potent side effects that are attributed to the long-term steroid use have promoted a search for alternatives for reducing the allergic lung inflammation and airway hyperreactivity seen in allergic asthma. One approach has been to explore anti-cytokine therapy for the treatment of allergic asthma, as interleukin (IL)-4, IL-5, and IL-13 are fundamentally important in the pathophysiology of allergic asthma. 166 Treatment of human asthmatics with MAbs against IL-5 demonstrated a decrease in the number of eosinophils, although no major change in the AHR was observed. 166 Considerable efforts have also been made to reduce the Th2-type responses that are detrimental to the asthmatic condition. The Th2 subsets express CCR3, CCR4, CCR8, and CXCR4, but only CXCR4 and its ligand SDF-1α have been found to be relevant during Th2-type allergic airway responses 167 (Fig. 2). The finding that Th2-type lymphocytes express CXCR4 on their cell surface, whereas Th1-type lymphocytes have very little or no expression of CXCR4, 168,169 adds further support. A cockroach allergen-induced animal model of asthma was used to show that blocking CXCR4 by AMD3100 significantly reduced airway hyperreactivity, peribronchial eosinophilia, and the overall inflammatory responses (Table 1; Fig. 2). 170 In fact, analysis of total leukocytes indicated an 80% reduction in recruited leukocytes over background numbers in AMD3100-treated animals compared to control animals treated with saline. Similarly, AMD3465 (Table 1) inhibited eosinophil recruitment during type-2 pulmonary granuloma formation after both Mycobacteria bovis purified protein derivative and Schistosoma mansoni egg antigen challenge, suggesting potential therapeutic application for chronic hypersensitivity diseases. 171 The blockade of CXCR4 as a means of controlling allergic asthma may lead to effective new therapeutics for down-regulating the inflammation and pathophysiology of allergen-induced response.
FUTURE DIRECTIONS AND PERSEPECTIVES
Drug discovery research targeting CXCR4, a GPCR, has made tremendous progress in the past 15 years since the 1996 report of CXCR4 as an important coreceptor for HIV-1 entry 25 and gone well beyond the focus of this receptor on HIV-1 infection to other human diseases and treatments including cancer, stem cell mobilization, and inflammation and autoimmune diseases. Given the diverse functions of this important GPCR and many potential therapeutic molecules discovered and reported thus far, some of which are described in this review, research on CXCR4 will continue to be intensive in the future. Some of the future research questions and areas of further investigations, which are not meant to be comprehensive but only reflect those familiar to us, are outlined below in order to highlight some selected examples of research that may be directly or indirectly relevant to the advancement of therapies targeting this important receptor.
STRUCTURE-BASED DRUG DISCOVERY USING THE CXCR4 CRYSTAL STRUCTURES
The determination of the crystal structures of CXCR4 bound to an antagonistic small molecule 56 provides the first high-resolution structure of a chemokine receptor bound to a synthetic ligand. This result is significant since (1) it allows the verification of previous structure–function studies of CXC4–ligand interactions and drug design work (i.e., checking on the accuracy of the predicted mechanisms of CXCR4 interactions with HIV-1, natural ligands, and de novo designed inhibitors); and more importantly, (2) it provides a new foundation for future drug design and discovery based on these crystal structures, such as the use of a receptor-based virtual screening approach (RBVS). 172 Prior to the publication of the CXCR4 crystal structures, drug discovery research that targets CXCR4 (and other GPCRs) mostly involved chemical screening and medicinal chemistry efforts 53,63-66,68 or use of the natural ligands as design templates. 98,99,173,174 With the new knowledge of the crystal structures of CXCR4, RBVS approaches can be used to exploit the molecular recognition between a ligand and CXCR4 to select out chemical entities that bind strongly to the active sites of CXCR4. This strategy has been successfully employed for identifying hits within a wide range of target proteins with experimentally determined or homology built structures, such Bcl-2, 175 acetylcholinesterase, 176 metalloenzymes, 177 phosphotriesterase, 177 and GPCRs. 178 As such, these crystal structures of CXCR4 will provide a more reliable basis for receptor structure-based drug design.
DRUG DISCOVERY RESEARCH THAT WILL BE MORE SPECIFIC TO DIFFERENT FUNCTIONS OF CXCR4
In addition to its roles in a wide range of human disease pathologies (i.e., HIV-1 entry, cancer metastasis, and inflammation and autoimmune diseases), the CXCR4–SDF-1α interaction has essential physiological functions in immunomodulation, organogenesis, hematopoiesis, and cerebellar neuron migration 100,102,179 (Figs. 1 and 2). This is further demonstrated by the death of CXCR4 or SDF-1α knockout mice, which succumb to hematopoietic, cardiac, vascular, and cerebellar defects during embryogenesis. 100-102 This highlights the importance of selective disruption of a particular targeted disease function of this important receptor without compromising its other functions. In this regard, some evidence shows that HIV-1 gp120 and SDF-1α interact with functionally distinct sites in CXCR4, 55 and that certain synthetic CXCR4 inhibitors such as SMM-chemokines interact with many residues on CXCR4 TM and extracellular domains that are important for HIV-1 entry, but they do not affect SDF-1α binding or signaling. 85 More research will be needed to determine the mechanistic and structural basis for the development of new inhibitory agents that may affect only the pathological responses of CXCR4, but not the physiological ones. In addition, specific functions of CXCR4 may be modulated by different signaling pathways, as studies showed that different signaling pathways are involved in neural stem cell migration versus inflammation. 180 Using SMM-chemokines as probes, we also found seemingly distinct signaling pathways of neuronal apoptosis involved in HIV-associated dementia (HAD) (Fig. 1), where these pathways were activated by different agonistic or antagonistic ligands of CXCR4 or CCR5. 181 In this regard, future research should focus on the dissection of distinct pathways responsible for different functions (pathological and physiological) of CXCR4 and the discovery of novel therapeutic agents that impact only the selective pathways, thereby leading to desirable effects on the targeted functions or responses. For example, it will be a question of significant clinical interest whether one may develop novel synthetic agonists that promote CXCR4-mediated stem cell movement towards injury sites for regenerative repairs without provoking unwanted inflammation that also involves the same receptor. The development of such disease function and pathway specific drugs may be conceivable once we understand more of the cellular, molecular, and structural basis underling the various functions of CXCR4.
DESIGN AND DISCOVERY OF CXCR4 AGONISTS
Most of the drug discovery targeting CXCR4 has focused on the development of CXCR4 antagonists for HIV-1 infection and cancer. Nevertheless, recent studies suggest that the activation of CXCR4 signaling may represent a new avenue for therapeutic development; for example, to mobilize hematopoietic stem cells or to direct stem cell migration towards brain or cardiac injury sites for regenerative repairs. 131-135,180,182 Exposure of NSCs to SDF-1α and subsequent induction of CXCR4-mediated signaling triggers a series of intracellular processes that lead to survival, proliferation, and migration of neural stem cells for injury repair. Indeed, we recently developed a panel of “dual-moiety” selective CXCR4 agonists that are capable of mediating NSC migration for injury repair in the brain, which points to the feasibility of this approach (unpublished results). It has also been reported that CXCR4 agonists, such as pepducin 116 and CTCE-0021, 182 can mobilize bone marrow hematopoietic cells. In addition to synthetic agonists, the natural agonist SDF-1α can be delivered by adenovirus to activate CXCR4. For instance, adenovirus-mediated SDF-1α gene transfer has been utilized to improve cardiac structure and function after experimental cardiac infarction through the combined effects of angiogenesis of CD133+ stem cells and anti-apoptosis.183,184 More recently, Lefrancois et al synthesized two CXCR4 agonists with chemotactic potencies only one log unit less that SDF-1α by linking the agonistic N-terminus of SDF-1α to the inverse agonist cyclopeptide T140. 185 As such, more interesting discoveries in the development of CXCR4 agonists are anticipated in the near future.
STRUCTURE–FUNCTION STUDIES OF CXCR4 –LIGAND BINDING AND SIGNALING MECHANISMS
The mechanism of ligand binding and activation of CXCR4 has been of immense basic importance and practical value for designing both antagonistic and agonistic ligands of CXCR4. Prior to the recent publication of the crystal structures of CXCR4 by Stevens’ group, 56 several groups sought to characterize CXCR4 interactions with HIV-1, natural ligands, and de novo designed inhibitors using molecular modeling, receptor chimeras, and site-specific mutagenesis. These studies have been exploited to validate the accuracy of a two-site model that was initially developed for the C5a chemoattractant and its receptor. 186 This model has the chemokine core domain as the “site one” docking domain and the chemokine N-terminus as the “site two” signaling trigger. According to this model, the motif composed of amino acids 12–17 of SDF-1α, RFFESH loop, first docks onto the N-terminal domain of CXCR4, and this contact allows the subsequent interaction of the flexible N-terminus of SDF-1α with the receptor groove formed by TM domains, thereby triggering the receptor function, whereas the globular portion of the chemokine binds to the extracellular loops. 17,186,187 The N-terminus of SDF-1α, being relatively flexible and unstructured in solution, has been confirmed as essential for CXCR4 recognition and signal transduction. 17,18,67 We incorporated this concept and further hypothesized a two-pocket model of CXCR4. 55,99 According to the model shown in Figure 3, the natural CXCR4 chemokine antagonist vMIP-II and its synthetic modified analogs DV1 and 9 (Table 1) are speculated to recognize a binding pocket in CXCR4, whereas the natural CXCR4 chemokine agonist SDF-1α reaches more deeply into another different signaling pocket. 188 The recently published CXCR4 crystal structures in complex with small molecule antagonists may allow consideration of more structure-based experiments to examine these hypotheses. In particular, new crystal structures of CXCR4 in complex with agonists, either natural or synthetic, and with natural antagonists and/or synthetic analogs derived from these natural templates (such as DV1) should shed more insight into CXCR4–ligand binding and signaling mechanisms and allow the testing of the above described two-site or two-pocket model for CXCR–ligand interaction.
Figure 3.

Two pocket model as demonstrated by distinct binding and signaling pockets in CXCR4. As described in our earlier publication, 188 these structural models show the superimposition of the average structures obtained from computational molecular dynamics simulations of 9–CXCR4 (A) and SDF-1α–CXCR4 complexes (B). The superimposition displays the difference in the binding of the N-termini of ligands. In the case of 9 (green), the N-terminus populates site A (red circle, denoted as binding pocket), whereas in the case of SDF-1α (red), the N-terminus populates a hydrophobic region in site B (blue circle, denoted as signaling pocket) in CXCR4. The first N-terminal residue for either ligand is displayed in a stick model.
STRUCTURE–FUNCTION STUDIES OF CXCR4 DIMERIZATION
Structure–function studies of CXCR4 also need to include the study of the role of CXCR4 dimerization, which can provide new opportunities for drug design. Five independent crystal structures of CXCR4 56 have each revealed consistent homodimers with an interface, including TM helices V and VI, which are consistent with previous biological studies that suggested the importance of CXCR4 homodimerization or heterodimerization (with other GPCRs) in CXCR4 function. 189-192 Another piece of evidence for CXCR4 dimerization involves the “warts, hypogammaglobulinemia, infections and myelokathexis” (WHIM) syndrome. 191 WHIM is primarily a heterozygous disease in which truncated CXCR4 is co-expressed with the wild-type receptor. Dimerization has been proposed as the most likely mechanism to explain the dominance of mutant CXCR4 over the wild-type receptor. Whether synthetic bivalent or multivalent ligands can be used to probe CXCR4 dimerization or polymerization represents an interesting area of research. Since the early 1980s, bivalent ligands which combine two identical pharmacophores, linked by a spacer, into a single molecule have been synthesized for many different GPCRs as specific pharmacological tools. Some of these molecules that contain two identical pharmacophores showed higher binding affinity, potency, or selectivity over their respective monomeric counterparts. 193-197 Our recent work on a synthetic novel bivalent ligand designated as DV1 dimer also suggests the feasibility of this approach for studying CXCR4 dimerization (unpublished results). Similarly, Tanaka et al recently designed and synthesized novel CXCR4 bivalent ligands consisting of two molecules of a 7 analogue, [cyclo(-D-Tyr-Arg-Arg-Nal-D-Cys-)] [Nal ) L-3-(2-naphthyl)alanine], connected by a poly(L-proline) or a PEGylated poly(L-proline) linker, and they determined the distance between two binding sites of ligands consisting of CXCR4 dimers. 197 The findings suggest that this type of “molecular ruler” approach could be utilized in the design of bivalent ligands for any GPCRs. This line of research will certainly be expanded in the future, not only to study CXCR4 dimerization or oligomerization, but also to understand how CXCR4 interacts with other GPCRs. Indeed, Sohy et al used a combination of luminescence complementation and bioluminescence resonance energy transfer assays to demonstrate the existence of hetero-oligomeric complexes composed of at least three chemokine receptors (CCR2, CCR5, and CXCR4). 198 Similarly, Sierro et al also demonstrated that CXCR7 plays a specialized role in endothelial biology and heart valve development by forming functional heterodimers with CXCR4 and enhancing SDF-1α-induced signaling. 199 More recently, Demmer et al constructed symmetric dimers with excellent antagonistic activity using a derivative of a cyclic pentapeptide as monomer. 202
THE FLEXIBLE NATURE OF CXCR4–LIGAND INTERFACE
The structural and stereochemical flexibility of CXCR4 is one interesting feature of CXCR4–ligand interactions that deserves further investigation. We previously reported a number of D-amino acid containing compounds such as DV1 and 9 (Table I). They represent various short peptides and full length protein analogs of vMIP-II that can bind to CXCR4 with potency and selectivity comparable to or higher than their L-peptide and protein counterparts (such as V1 and vMIP-II) (Table 1).98,99,173,188 This was surprising because of the profoundly different side chain topologies between D- and L-enantiomers, as they have mirror image conformations. This finding also suggested a marked stereochemical or conformational flexibility of CXCR4–peptide or CXCR4–protein interface, which appears to be distinctive from other known GPCR–ligand interactions that require a stricter stereochemistry. For instance, from the same studies of vMIP-II, it has been demonstrated that CCR5 binding has a strict structural requirement and involves multiple sites other than the N-terminus of vMIP-II. The recently reported CXCR4 crystal structures show a larger, more flexible, and open ligand binding pocket located closer to the extracellular surface, when compared to the binding sites of other GPCR structures noted in the past. 56 This supports our original findings and might explain the stereochemical flexibility seen in our studies of D-peptide ligands. One may further speculate that the flexibility of the CXCR4–ligand interface might be a feature of CXCR4 that HIV-1 capitalizes on by allowing sequence and conformational variations to occur in the V3 loop of the HIV-1 gp120. If the resulting changes in the gp120 V3 loop are accommodated by the flexible binding surface of CXCR4, thus promoting coreceptor switching from CCR5 to CXCR4, but not by that of an antibody, the virus can then evade neutralizing antibodies, especially after the host immunity is exhausted at the symptomatic phase of HIV-1 infection, while maintaining a high affinity for the coreceptor that is necessary for viral entry into the cell (Fig. 4). These are intriguing and potentially important hypotheses and questions that should be addressed in the future studies.
Figure 4.

A hypothetic mechanism for how CXCR4 exploits the flexible ligand binding surface of CXCR4 to evade the recognition of a neutralizing antibody while retaining the ability to bind CXCR4 for cellular entry. The flexibility of the CXCR4–ligand interface may allow sequence and conformational variations of V3 loop of the HIV-1 gp120. If the resulting changes in the gp120 V3 loop are accommodated by the flexible binding surface of CXCR4 but not by that of an antibody, the virus can then evade neutralizing antibodies while maintaining a high affinity for the coreceptor that is necessary for viral entry into the cell.
DEVELOPMENT AND CLINICAL APPLICATIONS OF NOVEL CXCR4-TARGETED MOLECULES DESIGNED BASED ON THE NATURAL CHEMOKINES
CXCR4 inhibitors that have entered clinical trials so far were originated from chemical screening and medicinal chemistry efforts, as exemplified by the much studied AMD3100 and its derivatives. As a complementary approach, chemically synthesized and engineered molecules designed based on the natural chemokine ligands of CXCR4, such as agonist SDF-1α and antagonist vMIP-II, are also important therapeutic candidates for drug development and clinical applications. For instance, SMM-chemokines are a class of such molecules with enhanced biological properties compared to natural chemokines, and they were developed by using concepts and techniques of synthetic biology and protein medicinal chemistry. Being engineered from the natural chemokines to target only the selective receptor and, perhaps in the future, a particular receptor-mediated disease pathway, these natural protein and peptide derived agents have been useful as not only tools to study the functions of the native ligands, but also promising leads to be examined in the future translational research to test their clinical values. As the techniques of nanotechnology and synthetic biology (which have already been applied to generate 9 and DV1 as shown in Table 1) can be used to enhance the enzymatic stability and in vivo half-lives of these peptide and protein-based therapeutics, and they do not need to cross the cell membrane for their therapeutic targets (which are the two major road blocks for peptide and protein drugs), it can be anticipated that synthetic modified analogs of the natural chemokines of CXCR4 will have significant values in the clinical settings.
Finally, whether drug discovery research strategies and results from the above areas of future research topics on CXCR4 are applicable to other chemokine receptors or GPCRs in general will need to be investigated. The mechanisms underlying CXCR4 ligand binding, signaling, and dimerization/oligomerization, and the design and discovery of new drugs based on the understanding of these mechanisms and the high-resolution crystal structures of CXCR4 may offer new insights or ideas for drug discovery research that targets other chemokine receptors and GPCRs. If so, CXCR4 will then become a useful model system for obtaining a general mechanistic understanding of drug discovery strategies for GPCRs, which currently represent the largest class of therapeutic drug targets.
ACKNOWLEDGMENT
We apologize to our colleagues if we have missed or failed to cite their work and papers in this review due to the specific focus and space constraint of this review and the diverse research topics and rapid development of this field. We thank all former and current members of our laboratories and collaborators at other institutions who contributed to the work cited in this review. Our studies were supported by grants from the National Institutes of Health, the Carol M. Baldwin Breast Cancer Research Fund, the Connolly Endowment/Hendricks Fund, and the LUNGevity Foundation.
ABBREVIATIONS
- CXCR4
CXC chemokine receptor 4
- GPCR
G-protein-coupled receptor
- HIV
human immunodeficiency virus
- AIDS
acquired immune deficiency syndrome
- SMM
synthetically and modularly modified
- SDF-1α
stromal cell-derived factor-1α
- vMIP-II
viral macrophage inflammatory protein-II
- MIP-1β
macrophage inflammatory protein-1β
- RANTES
regulated on activation, normal T-cell expressed and secreted
- TM
transmembrane
- IC
intracellular loop
- ECL
extracellular loop
- PI3K
PI3 kinase
- Akt
serine/threonine protein kinase
- BAD
Bcl-2-associated death promoter
- MEK
MAP kinase kinase
- JAK
Janus kinase
- STAT
signal transducer and activator of transcription
- cAMP
cyclic adenosine monophosphate
- MAb
monoclonal antibody
- BM
bone marrow
- HSC
hematopoietic stem cell
- HPC
hematopoietic progenitor cell
- G-CSF
granulocyte colony-stimulating factor
- PMN
polymorphonuclear neutrophil
- NHL
non-Hodgkin’s lymphoma
- CLL
chronic lymphocytic leukemia
- AML
acute myelogenous leukemia
- ALL
acute lymphoblastic leukemia
- NSC
neural stem cell
- CNS
central nervous system
- hNSC
human neural stem cell
- RA
rheumatoid arthritis
- IL
interleukin
- IFN
interferon
- TNF
tumor necrosis factor
- Th1
T helper 1
- AHR
airway hyper-responsiveness
- HAD
HIV-associated dementia
- WHIM
warts, hypogammaglobulinemia, infections and myelokathexis
BIOGRAPHIES
Won-Tak Choi earned his M.D. and Ph.D. (in Biochemistry) from the University of Illinois at Urbana-Champaign in 2009. He worked with Professor Ziwei Huang on the biochemical characterization of chemokine receptor CXCR4 interactions with HIV-1, natural ligands, and de novo designed inhibitors. Following his graduation in 2009, Dr. Choi completed his medical internship in the Department of Obstetrics and Gynecology at the University of California at San Francisco. He also undertook postdoctoral research working with Professor Donald E. Mosier at the Scripps Research Institute before joining the Department of Pathology at the University of Washington School of Medicine as a Resident Physician in 2011. His research interests include structure–function studies of receptor–ligand interactions implicated in human diseases and development of new therapeutics.
Srinivas Duggineni received his Ph.D. in Organic Chemistry under the supervision of Professor Bijoy Kundu at the Central Drug Research Institute Lucknow, India, in 2005. Upon graduation, he joined Aurigene Drug Discovery Ltd. at Bangalore from 2005 to 2007. Dr. Duggineni carried out postdoctoral research with Professor Ziwei Huang at the Sanford-Burnham Medical Research Institute, La Jolla, California, from 2007 to 2009. He is currently continuing his postdoctoral research with Professor Ziwei Huang at the Cancer Research Institute, Department of Pharmacology, State University of New York at Syracuse, New York. His main research interests include synthesis of peptides, peptidomimetics, and small molecules for anti-cancer and anti-HIV therapeutics.
Yan Xu graduated in medicinal chemistry at Jilin University in 2009 and received a M.S. degree under the guidance of Professors Xiaohong Yang and Song Li. In 2010, she received financial support from China Scholarship Council and worked with Professor Ziwei Huang as an exchange Ph.D. student at the State University of New York at Syracuse, New York. Her main research interests include medicinal chemistry, peptide chemistry, and nanocarriers for cancer therapy.
Ziwei Huang received a Ph.D. in chemistry in 1993 at the University of California at San Diego (UCSD) and then conducted postdoctoral research at the University of California at San Francisco. From 1995-2009, he held various faculty positions at the Sanford-Burnham Medical Research Institute, UCSD, the University of Illinois at Urbana-Champaign, and Kimmel Cancer Center of Jefferson Medical College. Since 2009, Dr. Huang is a Professor and Chairman of Department of Pharmacology, State University of New York (SUNY) at Syracuse, New York. In addition, he is the Director of SUNY Upstate Cancer Research Institute. His main research interests are the discovery and development of new synthetic probes and therapeutics targeting protein–protein interactions implicated in human diseases such as cancer, HIV infection, and neurodegeneration.
Jing An received her M.D. degree from China Medical University in 1984 and a Ph.D. degree from McGill University in 1997. She is currently an Associate Professor in the Department of Pharmacology, State University of New York at Syracuse, New York. She has worked for more than 10 years on the GPCR family proteins, CXCR4 and CCR5, and their signal regulation with a focus on discovery of peptides and small molecules that target CXCR4 and CCR5 as anti-HIV-1 entry agents. Over the past several years, she has participated in the development and characterization of several novel targeted therapeutic agents (e.g., Bcl-2, beta-catenin, CXCR4, CCR5, CD4, and CD8) using a combination of structural, chemical, molecular and cellular techniques, and animal models.
REFERENCES
- 1.Premack BA, Schall TJ. Chemokine receptors: gateways to inflammation and infection. Nature Medicine. 1996;2:1174–1178. doi: 10.1038/nm1196-1174. [DOI] [PubMed] [Google Scholar]
- 2.Murphy PM. The molecular biology of leukocyte chemoattractant receptors. Annu Rev Immunol. 1994;12:593–633. doi: 10.1146/annurev.iy.12.040194.003113. [DOI] [PubMed] [Google Scholar]
- 3.Berger EA, Murphy PM, Farber JM. Chemokine receptors as HIV-1 coreceptors: roles in viral entry, tropism, and disease. Annu Rev Immunol. 1999;17:657–700. doi: 10.1146/annurev.immunol.17.1.657. [DOI] [PubMed] [Google Scholar]
- 4.Kobilka B. Adrenergic receptors as models for G Protein-coupled receptors. Annu Rev Neurosci. 1992;15:87–114. doi: 10.1146/annurev.ne.15.030192.000511. [DOI] [PubMed] [Google Scholar]
- 5.Strader CD, Fong TM, Tota MR, Underwood D. Structure and function of G Protein-coupled Receptors. Annu Rev Biochem. 1994;63:101–132. doi: 10.1146/annurev.bi.63.070194.000533. [DOI] [PubMed] [Google Scholar]
- 6.Busillo JM, Benovic JL. Regulation of CXCR4 signaling. Biochim Biophys Acta. 2007;1768:952–963. doi: 10.1016/j.bbamem.2006.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hesselgesser J, Halks-Miller M, DelVecchio V, Peiper SC, Hoxie J, Kolson DL, Taub D, Horuk R. CD4-independent association between HIV-1 gp120 and CXCR4: functional chemokine receptors are expressed in human neurons. Curr Biol. 1997;7:112–121. doi: 10.1016/s0960-9822(06)00055-8. [DOI] [PubMed] [Google Scholar]
- 8.Hesselgesser J, Taub D, Baskar P, Greenberg M, Hoxie J, Kolson DL, Horuk R. Neuronal apoptosis induced by HIV-1 gp120 and the chemokine SDF-1 alpha is mediated by the chemokine receptor CXCR4. Curr Biol. 1998;8:595–598. doi: 10.1016/s0960-9822(98)70230-1. [DOI] [PubMed] [Google Scholar]
- 9.Sabroe IC, Gerard DM, Li NP, Collins Y, Post PD, Jose TW, Williams PJ, Gerard TJ, Ponath CJ, D. P. Cloning and characterization of the guinea pig eosinophil eotaxin receptor, C-C chemokine receptor-3: blockade using a monoclonal antibody in vivo. J Immunol. 1998;161:6139–6147. [PubMed] [Google Scholar]
- 10.Teicher BA, Fricker SP. CXCL12 (SDF-1)/CXCR4 pathway in cancer. Clin Cancer Res. 2010;16:2927–2931. doi: 10.1158/1078-0432.CCR-09-2329. [DOI] [PubMed] [Google Scholar]
- 11.Sodhi A, Montaner S, Gutkind JS. Viral hijacking of G-protein-coupled-receptor signalling networks. Nat Rev Mol Cell Biol. 2004;5:998–1012. doi: 10.1038/nrm1529. [DOI] [PubMed] [Google Scholar]
- 12.Mocchetti I, Bachis A, Masliah E. Chemokine receptors and neurotrophic factors: potential therapy against aids dementia? J Neurosci Res. 2008;86:243–255. doi: 10.1002/jnr.21492. [DOI] [PubMed] [Google Scholar]
- 13.Roggero R, Robert-Hebmann V, Harrington S, Roland J, Vergne L, Jaleco S, Devaux C, Biard-Piechaczyk M. Binding of human immunodeficiency virus type 1 gp120 to CXCR4 induces mitochondrial transmembrane depolarization and cytochrome c-mediated apoptosis independently of Fas signaling. J Virol. 2001;75:7637–7650. doi: 10.1128/JVI.75.16.7637-7650.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Bachis A, Biggio F, Major EO, Mocchetti I. M- and T-tropic HIVs promote apoptosis in rat neurons. J Neuroimmune Pharmacol. 2009;4:150–160. doi: 10.1007/s11481-008-9141-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Bardi G, Sengupta R, Khan MZ, Patel JP, Meucci O. Human immunodeficiency virus gp120-induced apoptosis of human neuroblastoma cells in the absence of CXCR4 internalization. J Neurovirol. 2006;12:211–218. doi: 10.1080/13550280600848373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Vila-Coro AJ, Rodríguez-Frade JM, De Ana A. Martín, Moreno-Ortíz MC, Martínez-A C, Mellado M. The chemokine SDF-1alpha triggers CXCR4 receptor dimerization and activates the JAK/STAT pathway. FASEB J. 1999;13:1699–1710. [PubMed] [Google Scholar]
- 17.Crump MP, Gong JH, Loetscher P, Rajarathnam K, Amara A, Arenzana-Seisdedos F, Virelizier JL, Baggiolini M, Sykes BD, Clark-Lewis I. Solution structure and basis for functional activity of stromal cell-derived factor-1; dissociation of CXCR4 activation from binding and inhibition of HIV-1. EMBO J. 1997;16:6996–7007. doi: 10.1093/emboj/16.23.6996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Dealwis C, Fernandez EJ, Thompson DA, Simon RJ, Siani MA, Lolis E. Crystal structure of chemically synthesized [N33A] stromal cell-derived factor 1alpha, a potent ligand for the HIV-1 “fusin” coreceptor. Proc Natl Acad Sci USA. 1998;95:6941–6946. doi: 10.1073/pnas.95.12.6941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Fernandez EJ, Wilken J, Thompson DA, Peiper SC, Lolis E. Comparison of the structure of vMIP-II with eotaxin-1, RANTES, and MCP-3 suggests a unique mechanism for CCR3 activation. Biochemistry. 2000;39:12837–12844. doi: 10.1021/bi001166f. [DOI] [PubMed] [Google Scholar]
- 20.Liwang AC, Wang ZX, Sun Y, Peiper SC, Liwang PJ. The solution structure of the anti-HIV chemokine vMIP-II. Protein Sci. 1999;8:2270–2280. doi: 10.1110/ps.8.11.2270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Lodi PJ, Garrett DS, Kuszewski J, Tsang ML, Weatherbee JA, Leonard WJ, Gronenborn AM, Clore GM. High-resolution solution structure of the beta chemokine hMIP-1 beta by multidimensional NMR. Science. 1994;263:1762–1767. doi: 10.1126/science.8134838. [DOI] [PubMed] [Google Scholar]
- 22.Chung CW, Cooke RM, Proudfoot AE, Wells TN. The three-dimensional solution structure of RANTES. Biochemistry. 1995;34:9307–9314. doi: 10.1021/bi00029a005. [DOI] [PubMed] [Google Scholar]
- 23.Proudfoot A. Chemokine receptors: multifaceted therapeutic targets. Nature Reviews/Immunology. 2002;2:106–115. doi: 10.1038/nri722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Fernandez EJ, Lolis E. Structure, function, and inhibition of chemokines. Annu Rev Pharmacol Toxicol. 2002;42:469–499. doi: 10.1146/annurev.pharmtox.42.091901.115838. [DOI] [PubMed] [Google Scholar]
- 25.Feng Y, Broder CC, Kennedy PE, Berger EA. HIV-1 entry cofactor: functional cDNA cloning of a seven-transmembrane, G protein-coupled receptor. Science. 1996;272:872–877. doi: 10.1126/science.272.5263.872. [DOI] [PubMed] [Google Scholar]
- 26.Deng H, Liu R, Ellmeier W, Choe S, Unutmaz D, Burkhart M, Marzio PD, Marmon S, Sutton RE, Hill CM, Davis CB, Peiper SC, Schall TJ, Littman DR, Landau NR. Identification of a major co-receptor for primary isolates of HIV-1. Nature. 1996;381:661–666. doi: 10.1038/381661a0. [DOI] [PubMed] [Google Scholar]
- 27.Dragic T, Litwin V, Allaway GP, Martin SR, Huang Y, Nagashima KA, Cayanan C, Maddon PJ, Koup RA, Moore JP, Paxton WA. HIV-1 entry into CD4+ cells is mediated by the chemokine receptor CC-CKR-5. Nature. 1996;381:667–673. doi: 10.1038/381667a0. [DOI] [PubMed] [Google Scholar]
- 28.Alkhatib G, Combadiere C, Broder CC, Feng Y, Kennedy PE, Murphy PM. CC CKR5: a RANTES, MIP-1alpha, MIP-1beta receptor as a fusion cofactor for macrophage-tropic HIV-1. Science. 1996;272:1955–1958. doi: 10.1126/science.272.5270.1955. [DOI] [PubMed] [Google Scholar]
- 29.Gershoni JM, Denisova G, Raviv D, Smorodinsky NI, Buyaner D. HIV binding to its receptor creates specific epitopes for the CD4/gp120 complex. FASEB J. 1993;7:1185–1187. doi: 10.1096/fasebj.7.12.7690724. [DOI] [PubMed] [Google Scholar]
- 30.Clements GJ, Proce-Jones MJ, Stephens PE, Sutton C, Schulz TF, Clapham PR, McKeating JA, McClure MO, Thomson S, Marsh M, Kay J, Weiss RA, Moore HP. The V3 loops of the HIV-1 and HIV-2 surface glycoproteins contain proteolytic cleavage sites: a possible function in viral fusion? AIDS Research & Human Retroviruses. 1991;7:3–16. doi: 10.1089/aid.1991.7.3. [DOI] [PubMed] [Google Scholar]
- 31.Sattentau QJ, Moore JP, Vignaux F, Traincard F, Poignard P. Conformational changes induced in the envelope glycoproteins of the human and simian immunodeficiency viruses by soluble receptor binding. J Virol. 1993;67:7383–7393. doi: 10.1128/jvi.67.12.7383-7393.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Lapham CK, Ouyang J, Chandrasekhar B, Nguyen NY, Dimitrov DS, Golding H. Evidence for cell-surface association between fusin and the CD4-gp120. Science. 1996;274:602–605. doi: 10.1126/science.274.5287.602. [DOI] [PubMed] [Google Scholar]
- 33.Wu L, Gerard NP, Wyatt R, Choe H, Parolin C, Ruffing N, Borsetti A, Cardoso AA, Desjardin E, Newman W, Gerard C, Sodroski J. CD4-induced interaction of primary HIV-1 gp120 glycoproteins with the chemokine receptor CCR-5. Nature. 1996;6605:179–183. doi: 10.1038/384179a0. [DOI] [PubMed] [Google Scholar]
- 34.Trkola A, Dragic T, Arthos J, Binley JM, Olson WC, Allaway GP, Cheng-Mayer C, Robinson J, Maddon PJ, Moore JP. CD4-dependent, antibody-sensitive interactions between HIV-1 and its co-receptor CCR-5. Nature. 1996;384:184–187. doi: 10.1038/384184a0. [DOI] [PubMed] [Google Scholar]
- 35.Cheng-Mayer C, Seto D, Tateno M, Levy JA. Biologic features of HIV-1 that correlate with virulence in the host. Science. 1988;240:80–82. doi: 10.1126/science.2832945. [DOI] [PubMed] [Google Scholar]
- 36.Tersmette M, Lange JM, de Goede RE, de Wolf F, Eeftink-Schattenkerk JK, Schellekens PT, Coutinho RA, Huisman JG, Goudsmit J, Miedema F. Association between biological properties of human immunodeficiency virus variants and risk for AIDS and AIDS mortality. Lancet. 1989;1:983–985. doi: 10.1016/s0140-6736(89)92628-7. [DOI] [PubMed] [Google Scholar]
- 37.Schellekens PT, Tersmette M, Roos MT, Keet RP, de Wolf F, Coutinho RA, Miedema F. Biphasic rate of CD4+ cell count decline during progression to AIDS correlates with HIV-1 phenotype. AIDS. 1992;6:665–669. doi: 10.1097/00002030-199207000-00008. [DOI] [PubMed] [Google Scholar]
- 38.Bleul CC, Farzan M, Choe H, Parolin C, Clark-Lewis I, Sodroski J. The lymphocyte chemoattractant SDF-1 is a ligand for LESTR/fusin and blocks HIV-1 entry. Nature. 1996;382:829–833. doi: 10.1038/382829a0. [DOI] [PubMed] [Google Scholar]
- 39.Oberlin E, Amara A, Bachelerie F, Bessia C, Virelizier JL, Arenzana-Seisdedos F, Schwartz O, Heard JM, Clark-Lewis I, Legler DF, Loetscher M, Baggiolini M, Moser B. The CXC chemokine SDF-1 is the ligand for LESTR/fusin and prevents infection by T-cell-line-adapted HIV-1. Nature. 1996;382:833–835. doi: 10.1038/382833a0. [DOI] [PubMed] [Google Scholar]
- 40.Amara A, Gall SL, Schwartz O, Salamero J, Montes M, Loetscher P, Baggiolini M, Virelizier J-L, Arenzana-Seisdedos F. HIV coreceptor downregulation as antiviral principle: SDF-1α-dependent internalization of the chemokine receptor CXCR4 contributes to inhibition of HIV replication. J Exp Med. 1997;186:139–146. doi: 10.1084/jem.186.1.139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Förster R, Kremmer E, Schubel A, Breitfeld D, Kleinschmidt A, Nerl C, Bernhardt G, Lipp M. Intracellular and surface expression of the HIV-1 coreceptor CXCR4/fusin on various leukocyte subsets: rapid internalization and recycling upon activation. J Immunol. 1998;160:1522–1531. [PubMed] [Google Scholar]
- 42.Moncunill G, Armand-Ugón M, Pauls E, Clotet B, Esté JA. HIV-1 escape to CCR5 coreceptor antagonism through selection of CXCR4-using variants in vitro. AIDS. 2008;22:23–31. doi: 10.1097/QAD.0b013e3282f303e6. [DOI] [PubMed] [Google Scholar]
- 43.Arastéh K, Stocker H. Tropism switch in patients infected with HIV-1 and its clinical implications for the treatment with CCR5-receptor inhibitors. Eur J Med Res. 2007;12:397–402. [PubMed] [Google Scholar]
- 44.Westby M, Lewis M, Whitcomb J, Youle M, Pozniak AL, James IT, Jenkins TM, Perros M, van der Ryst E. Emergence of CXCR4-using human immunodeficiency virus type 1 (HIV-1) variants in a minority of HIV-1-infected patients following treatment with the CCR5 antagonist maraviroc is from a pretreatment CXCR4-using virus reservoir. J Virol. 2006;80:4909–4920. doi: 10.1128/JVI.80.10.4909-4920.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Westby M, Smith-Burchnell C, Mori J, Lewis M, Mosley M, Stockdale M, Dorr P, Ciaramella G, Perros M. Reduced maximal inhibition in phenotypic susceptibility assays indicates that viral strains resistant to the CCR5 antagonist maraviroc utilize inhibitor-bound receptor for entry. J Virol. 2007;81:2359–2371. doi: 10.1128/JVI.02006-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Nedellec R, Coetzer M, Lederman MM, Offord RE, Hartley O, Mosier DE. Resistance to the CCR5 inhibitor 5P12-RANTES requires a difficult evolution from CCR5 to CXCR4 coreceptor use. PLoS One. 2011;6:e22020. doi: 10.1371/journal.pone.0022020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Rucker J, Samson M, Doranz BJ, Libert F, Berson JF, Yi Y, Smyth RJ, Collman RG, Broder CC, Vassart G, Doms RW, Parmentier M. Regions in beta-chemokine receptors CCR5 and CCR2b that determine HIV-1 cofactor specificity. Cell. 1996;87:437–446. doi: 10.1016/s0092-8674(00)81364-1. [DOI] [PubMed] [Google Scholar]
- 48.Atchison RE, Gosling J, Monteclaro FS, Franci C, Digilio L, Charo IF, Goldsmith MA. Multiple extracellular elements of CCR5 and HIV-1 entry: dissociation from response to chemokines. Science. 1996;274:1924–1926. doi: 10.1126/science.274.5294.1924. [DOI] [PubMed] [Google Scholar]
- 49.Brelot A, Heveker N, Pleskoff O, Sol N, Alizon M. Role of the first and third extracellular domains of CXCR-4 in human immunodeficiency virus coreceptor activity. J Virol. 1997;71:4744–4751. doi: 10.1128/jvi.71.6.4744-4751.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Lu Z, Berson JF, Chen Y, Turner JD, Zhang T, Sharron M, Jenks MH, Wang Z, Kim J, Rucker J, Hoxie JA, Peiper SC, Doms RW. Evolution of HIV-1 coreceptor usage through interactions with distinct CCR5 and CXCR4 domains. Proc Natl Acad Sci U S A. 1997;94:6426–6431. doi: 10.1073/pnas.94.12.6426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Bieniasz PD, Fridell RA, Aramori I, Ferguson SS, Caron MG, Cullen BR. HIV-1-induced cell fusion is mediated by multiple regions within both the viral envelope and the CCR-5 co-receptor. EMBO J. 1997;16:2599–2609. doi: 10.1093/emboj/16.10.2599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Alkhatib G, Ahuja SS, Light D, Mummidi S, Berger EA, Ahuja SK. CC chemokine receptor 5-mediated signaling and HIV-1 co-receptor activity share common structure determinants. J Biol Chem. 1997;272:19771–19776. doi: 10.1074/jbc.272.32.19771. [DOI] [PubMed] [Google Scholar]
- 53.Doranz BJ, Grovit-Ferbas K, Sharron MP, Mao SH, Goetz MB, Daar ES, Doms RW, O’Brien WA. A small-molecule inhibitor directed against the chemokine receptor CXCR4 prevents its use as an HIV-1 coreceptor. J Exp Med. 1997;186:1395–1400. doi: 10.1084/jem.186.8.1395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Chabot DJ, Broder CC. Substitutions in a homologous region of extracellular loop 2 of CXCR4 and CCR5 alter coreceptor activities for HIV-1 membrane fusion and virus entry. J Biol Chem. 2000;275:23774–23782. doi: 10.1074/jbc.M003438200. [DOI] [PubMed] [Google Scholar]
- 55.Tian S, Choi WT, Liu D, Pesavento J, Wang Y, An J, Sodroski JG, Huang Z. Distinct functional sites for human immunodeficiency virus type 1 and stromal cell-derived factor 1a on CXCR4 transmembrane helical domains. J Virol. 2005;79:12667–12673. doi: 10.1128/JVI.79.20.12667-12673.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Wu B, Chien EY, Mol CD, Fenalti G, Liu W, Katritch V, Abagyan R, Brooun A, Wells P, Bi FC, Hamel DJ, Kuhn P, Handel TM, Cherezov V, Stevens RC. Structures of the CXCR4 chemokine GPCR with small-molecule and cyclic peptide antagonists. Science. 2010;330:1066–1071. doi: 10.1126/science.1194396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Schols D, Struyf S, van Damme J, Este JA, Henson G, De Clercq E. Inhibition of T-tropic HIV strains by selective antagonization of the chemokine receptor CXCR4. J Exp Med. 1997;186:1383–1388. doi: 10.1084/jem.186.8.1383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Murakami T, Nakajima T, Koyanagi Y, Tachibana K, Fujii N, Tamamura H, Yoshida N, Waki M, Matsumoto A, Yoshie O, Kishimoto T, Yamamoto N, Nagasawa T. A small molecule CXCR4 inhibitor that blocks T cell line-tropic HIV-1 infection. J Exp Med. 1997;186:1389–1393. doi: 10.1084/jem.186.8.1389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Luo Z, Zhou N, Luo J, Hall JW, Huang Z. The role of positively charged residues in CXCR4 recognition probed with synthetic peptides. Biochem Biophy Res Comm. 1999;263:691–695. doi: 10.1006/bbrc.1999.1441. [DOI] [PubMed] [Google Scholar]
- 60.Luo J, Luo Z, Zhou N, Hall JW, Huang Z. Attachment of C-terminus of SDF-1 enhances the biological activity of its N-terminal peptide. Biochem Biophys Res Commun. 1999;264:42–47. doi: 10.1006/bbrc.1999.1476. [DOI] [PubMed] [Google Scholar]
- 61.Luo Z, Fan X, Zhou N, Hiraoka M, Luo J, Kaji H, Huang Z. Structure-function study and anti-HIV activity of synthetic peptide analogues derived from viral chemokine vMIP-II. Biochemistry. 2000;39:13545–13550. doi: 10.1021/bi000633q. [DOI] [PubMed] [Google Scholar]
- 62.Huang Z. Structural chemistry and therapeutic intervention of protein-protein interaction in immune response, HIV entry and apoptosis. Pharmacology & Therapeutics. 2000;86:201–215. doi: 10.1016/s0163-7258(00)00052-8. [DOI] [PubMed] [Google Scholar]
- 63.Zhou N, Luo Z, Luo J, Hall JW, Huang Z. A novel peptide antagonist of CXCR4 derived from the the N-terminus of the viral chemokine vMIP-II. Biochemistry. 2000;39:3782–3787. doi: 10.1021/bi992750v. [DOI] [PubMed] [Google Scholar]
- 64.Zhou N, Luo Z, Luo J, Fan X, Cayabyab M, Hiraoka M, Liu D, Han X, Pesavento J, Dong CZ, Wang Y, An J, Kaji H, Sodroski JG, Huang Z. Exploring the stereochemistry of CXCR4-peptide recognition and inhibiting HIV-1 entry with D-peptides derived from chemokines. J Biol Chem. 2002;277:17476–17485. doi: 10.1074/jbc.M202063200. [DOI] [PubMed] [Google Scholar]
- 65.Donzella GA, Schols D, Lin SW, Este JA, Nagashima KA, Maddon PJ, Allaway GP, Sakmar TP, Henson G, De Clercq E, Moore JP. AMD3100, a small molecule inhibitor of HIV-1 entry via the CXCR4 co-receptor. Nature Medicine. 1998;4:72–77. doi: 10.1038/nm0198-072. [DOI] [PubMed] [Google Scholar]
- 66.Fenard D, Lambeau G, Maurin T, Lefebvre JC, Doglio A. A peptide derived from bee venom-secreted phospholipase A2 inhibits replication of T-cell tropic HIV-1 strains via interaction with the CXCR4 chemokine receptor. Mol Pharmacol. 2001;60:341–347. doi: 10.1124/mol.60.2.341. [DOI] [PubMed] [Google Scholar]
- 67.Heveker N, Montes M, Germeroth L, Amara A, Trautmann A, Alizon M, Schneider-Mergener J. Dissociation of the signalling and antiviral properties of SDF-1-derived small peptides. Current Biology. 1998;8:369–376. doi: 10.1016/s0960-9822(98)70155-1. [DOI] [PubMed] [Google Scholar]
- 68.Sachpatzidis A, Benton BK, Manfredi JP, Wang H, Hamilton A, Dohlman HG, Lolis E. Identification of allosteric peptide agonists of CXCR4. J Biol Chem. 2003;278:896–907. doi: 10.1074/jbc.M204667200. [DOI] [PubMed] [Google Scholar]
- 69.Loetscher P, Gong JH, Dewald B, Baggiolini M, Clark-Lewis I. N-terminal peptides of stromal cell-derived factor-1 with CXC chemokine receptor 4 agonist and antagonist activities. J Biol Chem. 1998;273:22279–22283. doi: 10.1074/jbc.273.35.22279. [DOI] [PubMed] [Google Scholar]
- 70.Samson M, Libert F, Doranz BJ, Rucker J, Liesnard C, Farber CM, Saragosti S, Lapoumeroulie C, Cognaux J, Forceille C, Muyldermans G, Verhofstede C, Burtonboy G, Georges M, Imai T, Rana S, Yi Y, Smyth RJ, Collman RG, Doms RW, Vassart G, Parmentier M. Resistance to HIV-1 infection in caucasian individuals bearing mutant alleles of the CCR-5 chemokine receptor gene. Nature. 1996;382:722–725. doi: 10.1038/382722a0. [DOI] [PubMed] [Google Scholar]
- 71.Lim JK, McDermott DH, Lisco A, Foster GA, Krysztof D, Follmann D, Stramer SL, Murphy PM. CCR5 deficiency is a risk factor for early clinical manifestations of West Nile virus infection but not for viral transmission. J Infect Dis. 2010;201:178–185. doi: 10.1086/649426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.O’Brien WA, Sumner-Smith M, Mao SH, Sadeghi S, Zhao JQ, Chen IS. Anti-human immunodeficiency virus type 1 activity of an oligocationic compound mediated via gp120 V3 interactions. J Virol. 1996;70:2825–2831. doi: 10.1128/jvi.70.5.2825-2831.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Sumner-Smith M, Zheng Y, Zhang YP, Twist EM, Climie SC. Antiherpetic activities of N-alpha-acetyl-nona-D-arginine amide acetate. Drugs Exp Clin Res. 1995;21:1–6. [PubMed] [Google Scholar]
- 74.Gerlach LO, Skerlj RT, Bridger GJ, Schwartz TW. Molecular interactions of cyclam and bicyclam non-peptide antagonists with the CXCR4 chemokine receptor. J Biol Chem. 2001;276:14153–14160. doi: 10.1074/jbc.M010429200. [DOI] [PubMed] [Google Scholar]
- 75.Rosenkilde MM, Gerlach LO, Jakobsen JS, Skerlj RT, Bridger GJ, Schwartz TW. Molecular mechanism of AMD3100 antagonism in the CXCR4 receptor: transfer of binding site to the CXCR3 receptor. J Biol Chem. 2004;279:3033–3041. doi: 10.1074/jbc.M309546200. [DOI] [PubMed] [Google Scholar]
- 76.Kawatkar SP, Yan M, Gevariya H, Y. M, L, Eisold S, Zhu X, Huang Z, An J. Computational analysis of the structural mechanism of inhibition of chemokine receptor CXCR4 by small molecule antagonists. Exp Biol Med. 2011;236:844–850. doi: 10.1258/ebm.2011.010345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Zhan W, Liang Z, Zhu A, Kurtkaya S, Shim H, Snyder JP, Liotta DC. Discovery of small molecule CXCR4 antagonists. J Med Chem. 2007;50:5655–5664. doi: 10.1021/jm070679i. [DOI] [PubMed] [Google Scholar]
- 78.Hatse S, Princen K, De Clercq E, Rosenkilde MM, Schwartz TW, Hernandez-Abad PE, Skerlj RT, Bridger GJ, Schols D. AMD3465, a monomacrocyclic CXCR4 antagonist and potent HIV entry inhibitor. Biochem Pharmacol. 2005;70:752–761. doi: 10.1016/j.bcp.2005.05.035. [DOI] [PubMed] [Google Scholar]
- 79.Bodart V, Anastassov V, Darkes MC, Idzan SR, Labrecque J, Lau G, Mosi RM, Neff KS, Nelson KL, Ruzek MC, Patel K, Santucci Z, Scarborough R, Wong RS, Bridger GJ, Macfarland RT, Fricker SP. Pharmacology of AMD3465: a small molecule antagonist of the chemokine receptor CXCR4. Biochem Pharmacol. 2009;78:993–1000. doi: 10.1016/j.bcp.2009.06.010. [DOI] [PubMed] [Google Scholar]
- 80.Stone ND, Dunaway SB, Flexner C, Tierney C, Calandra GB, Becker S, Cao YJ, Wiggins IP, Conley J, MacFarland RT, Park JG, Lalama C, Snyder S, Kallungal B, Klingman KL, Hendrix CW. Multiple-dose escalation study of the safety, pharmacokinetics, and biologic activity of oral AMD070, a selective CXCR4 receptor inhibitor, in human subjects. Antimicrob Agents Chemother. 2007;51:2351–2358. doi: 10.1128/AAC.00013-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Skerlj RT, Bridger GJ, Kaller A, McEachern EJ, Crawford JB, Zhou Y, Atsma B, Langille J, Nan S, Veale D, Wilson T, Harwig C, Hatse S, Princen K, De Clercq E, Schols D. Discovery of novel small molecule orally bioavailable C-X-C chemokine receptor 4 antagonists that are potent inhibitors of T-tropic (X4) HIV-1 replication. J Med Chem. 2010;53:3376–3388. doi: 10.1021/jm100073m. [DOI] [PubMed] [Google Scholar]
- 82.Skerlj R, Bridger G, McEachern E, Harwig C, Smith C, Wilson T, Veale D, Yee H, Crawford J, Skupinska K, Wauthy R, Yang W, Zhu Y, Bogucki D, Di Fluri M, Langille J, Huskens D, De Clercq E, Schols D. Synthesis and SAR of novel CXCR4 antagonists that are potent inhibitors of T tropic (X4) HIV-1 replication. Bioorg Med Chem Lett. 2011;21:262–266. doi: 10.1016/j.bmcl.2010.11.023. [DOI] [PubMed] [Google Scholar]
- 83.Skerlj R, Bridger G, McEachern E, Harwig C, Smith C, Kaller A, Veale D, Yee H, Skupinska K, Wauthy R, Wang L, Baird I, Zhu Y, Burrage K, Yang W, Sartori M, Huskens D, De Clercq E, Schols D. Design of novel CXCR4 antagonists that are potent inhibitors of T-tropic (X4) HIV-1 replication. Bioorg Med Chem Lett. 2011;21:1414–1418. doi: 10.1016/j.bmcl.2011.01.021. [DOI] [PubMed] [Google Scholar]
- 84.Ichiyama K, Yokoyama-Kumakura S, Tanaka Y, Tanaka R, Hirose K, Bannai K, Edamatsu T, Yanaka M, Niitani Y, Miyano-Kurosaki N, Takaku H, Koyanagi Y, Yamamoto N. A duodenally absorbable CXC chemokine receptor 4 antagonist, KRH-1636, exhibits a potent and selective anti-HIV-1 activity. Proc Natl Acad Sci U S A. 2003;100:4185–4190. doi: 10.1073/pnas.0630420100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Murakami T, Kumakura S, Yamazaki T, Tanaka R, Hamatake M, Okuma K, Huang W, Toma J, Komano J, Yanaka M, Tanaka Y, Yamamoto N. The novel CXCR4 antagonist KRH-3955 is an orally bioavailable and extremely potent inhibitor of human immunodeficiency virus type 1 infection: comparative studies with AMD3100. Antimicrob Agents Chemother. 2009;53:2940–2948. doi: 10.1128/AAC.01727-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Iwasaki Y, Akari H, Murakami T, Kumakura S, Dewan MZ, Yanaka M, Yamamoto N. Efficient inhibition of SDF-1alpha-mediated chemotaxis and HIV-1 infection by novel CXCR4 antagonists. Cancer Sci. 2009;100:778–781. doi: 10.1111/j.1349-7006.2009.01104.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Jenkinson S, Thomson M, McCoy D, Edelstein M, Danehower S, Lawrence W, Wheelan P, Spaltenstein A, Gudmundsson K. Blockade of X4-tropic HIV-1 cellular entry by GSK812397, a potent noncompetitive CXCR4 receptor antagonist. Antimicrob Agents Chemother. 2010;54:817–824. doi: 10.1128/AAC.01293-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Tamamura H, Omagari A, Hiramatsu K, Gotoh K, Kanamoto T, Xu Y, Kodama E, Matsuoka M, Hattori T, Yamamoto N, Nakashima H, Otaka A, Fujii N. Development of specific CXCR4 inhibitors possessing high selectivity indexes as well as complete stability in serum based on an anti-HIV peptide T140. Bioorg Med Chem Lett. 2001;11:1897–1902. doi: 10.1016/s0960-894x(01)00323-7. [DOI] [PubMed] [Google Scholar]
- 89.Fujii NO, Hiramatsu S, Araki K, Ueda T, Tamamura S, Otaka H, Kusano A, Terakubo S, Nakashima S, Broach H, Trent JA, Wang JO, Peiper ZX, C. S. Molecular-size reduction of a potent CXCR4-chemokine antagonist using orthogonal combination of conformation- and sequence-based libraries. Angew Chem Int Ed Engl. 2003;42:3251–3253. doi: 10.1002/anie.200351024. [DOI] [PubMed] [Google Scholar]
- 90.Narumi T, Hayashi R, Tomita K, Kobayashi K, Tanahara N, Ohno H, Naito T, Kodama E, Matsuoka M, Oishi S, Fujii N. Synthesis and biological evaluation of selective CXCR4 antagonists containing alkene dipeptide isosteres. Org Biomol Chem. 2010;8:616–621. doi: 10.1039/b917236j. [DOI] [PubMed] [Google Scholar]
- 91.Ueda S, Oishi S, Wang ZX, Araki T, Tamamura H, Cluzeau J, Ohno H, Kusano S, Nakashima H, Trent JO, Peiper SC, Fujii N. Structure-activity relationships of cyclic peptide-based chemokine receptor CXCR4 antagonists: disclosing the importance of side-chain and backbone functionalities. J Med Chem. 2007;50:192–198. doi: 10.1021/jm0607350. [DOI] [PubMed] [Google Scholar]
- 92.Ueda S, Kato M, Inuki S, Ohno H, Evans B, Wang ZX, Peiper SC, Izumi K, Kodama E, Matsuoka M, Nagasawa H, Oishi S, Fujii N. Identification of novel non-peptide CXCR4 antagonists by ligand-based design approach. Bioorg Med Chem Lett. 2008;18:4124–4129. doi: 10.1016/j.bmcl.2008.05.092. [DOI] [PubMed] [Google Scholar]
- 93.Hamy F, Felder ER, Heizmann G, Lazdins J, Aboulela F, Varani G, Karn J, Klimkait T. An inhibitor of the Tat/TAR RNA interaction that effectively suppresses HIV-1 replication. Proc Nat Acad Sci USA. 1997;94:3548–3553. doi: 10.1073/pnas.94.8.3548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Daelemans D, Schols D, Witvrouw M, Pannecouque C, Hatse S, van Dooren S, Hamy F, Klimkait T, de Clercq E, VanDamme AM. A second target for the peptoid Tat/transactivation response element inhibitor CGP64222: inhibition of human immunodeficiency virus replication by blocking CXC-chemokine receptor 4-mediated virus entry. Mol Pharmacol. 2000;57:116–124. [PubMed] [Google Scholar]
- 95.LaBarge MA, Blau HM. Biological progression from adult bone marrow to mononucleate muscle stem cell to multinucleate muscle fiber in response to injury. Cell. 2002;111:589–601. doi: 10.1016/s0092-8674(02)01078-4. [DOI] [PubMed] [Google Scholar]
- 96.Kledal TN, Rosenkilde MM, Coulin F, Simmons G, Johnsen AH, Alouani S, Power CA, Luttichau HR, Gerstoft J, Clapham PR, Clark-Lewis I, Wells TNC, Schwartz TW. A broad-spectrum chemokine antagonist encoded by Kaposi’s sarcoma-associated herpesvirus. Science. 1997;277:1656–1659. doi: 10.1126/science.277.5332.1656. [DOI] [PubMed] [Google Scholar]
- 97.Boshoff C, Endo Y, Collins PD, Takeuchi Y, Reeves JD, Schweickart VL, Siani MA, Sasaki T, Williams TJ, Gray PW, Moore PS, Chang Y, Weiss RA. Angiogenic and HIV-inhibitory functions of KSHV-encoded chemokines. Science. 1997;278:290–294. doi: 10.1126/science.278.5336.290. [DOI] [PubMed] [Google Scholar]
- 98.Kumar S, Choi WT, Dong CZ, Madani N, Tian S, Liu D, Wang Y, Pesavento J, Wang J, Fan X, Yuan J, Fritzsche WR, An J, Sodroski JG, Richman DD, Huang Z. SMM-Chemokines: a class of unnatural synthetic molecules as chemical probes of chemical receptor biology and leads for therapeutic development. Chemistry & Biology. 2006;13:69–79. doi: 10.1016/j.chembiol.2005.10.012. [DOI] [PubMed] [Google Scholar]
- 99.Choi WT, Tian S, Dong CZ, Kumar S, Liu D, Madani N, An J, Sodroski JG, Huang Z. Unique ligand binding sites on CXCR4 probed by a chemical biology approach: implications for the design of selective human immunodeficiency virus type 1 inhibitors. J Virol. 2005;79:15398–15404. doi: 10.1128/JVI.79.24.15398-15404.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Zou Y, Kottmann A, Kuroda M, Taniuchi I, Littman D. Function of the chemokine receptor CXCR4 in haematopoiesis and in cerebellar development. Nature. 1998;393:595–599. doi: 10.1038/31269. [DOI] [PubMed] [Google Scholar]
- 101.Tachibana K, Hirota S, Iizasa H, Yoshida H, Kawabata K, Kataoka Y, Kitamura Y, Matsushima K, Yoshida N, Nishikawa S, Kishimoto T, Nagasawa T. The chemokine receptor CXCR4 is essential for vascularization of the gastrointestinal tract. Nature. 1998;393:591–594. doi: 10.1038/31261. [DOI] [PubMed] [Google Scholar]
- 102.Nagasawa T, Hirota S, Tachibana K, Takakura N, Nishikawa S, Kitamura Y, Yoshida N, Kikutani H, Kishimoto T. Defects of B-cell lymphopoiesis and bone-marrow myelopoiesis in mice lacking the CXC chemokine PBSF/SDF-1. Nature. 1996;382:635–638. doi: 10.1038/382635a0. [DOI] [PubMed] [Google Scholar]
- 103.Doranz BJ, Filion LG, Diaz-Mitoma F, Sitar DS, Sahai J, Baribaud F, Orsinis MJ, Benovic JL, Cameron W, Doms RW. Safe use of the CXCR4 inhibitor ALX40-4C in humans. AIDS Res Hum Retroviruses. 2001;17:475–486. doi: 10.1089/08892220151126508. [DOI] [PubMed] [Google Scholar]
- 104.Pelus LM. Peripheral blood stem cell mobilization: new regimens, new cells, where do we stand. Curr Opin Hematol. 2008;15:285–292. doi: 10.1097/MOH.0b013e328302f43a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Robinson WA, Kurnick JE, Pike BL. Colony growth of human leukemic peripheral blood cells in vitro. Blood. 1971;38:500–508. [PubMed] [Google Scholar]
- 106.McCredie KB, Hersh EM, Freireich EJ. Cells capable of colony formation in the peripheral blood of man. Science. 1971;171:293–294. doi: 10.1126/science.171.3968.293. [DOI] [PubMed] [Google Scholar]
- 107.Roberts AW, DeLuca E, Begley CG, Basser R, Grigg AP, Metcalf D. road inter-individual variations in circulating progenitor cell numbers induced by granulocyte colony-stimulating factor therapy. Stem Cells. 1995;13:512–516. doi: 10.1002/stem.5530130508. [DOI] [PubMed] [Google Scholar]
- 108.Lapidot T, Kollet O. The essential roles of the chemokine SDF-1 and its receptor CXCR4 in human stem cell homing and repopulation of transplanted immune-deficient NOD/SCID and NOD/SCID/B2m(null) mice. Leukemia. 2002;16:1992–2003. doi: 10.1038/sj.leu.2402684. [DOI] [PubMed] [Google Scholar]
- 109.Kollet O, Petit I, Kahn J, Samira S, Dar A, Peled A, Deutsch V, Gunetti M, Piacibello W, Nagler A, Lapidot T. Human CD34(+)CXCR4(-) sorted cells harbor intracellular CXCR4, which can be functionally expressed and provide NOD/SCID repopulation. Blood. 2002;100:2778–2786. doi: 10.1182/blood-2002-02-0564. [DOI] [PubMed] [Google Scholar]
- 110.Kim CH, Broxmeyer HE. In vitro behavior of hematopoietic progenitor cells under the influence of chemoattractants: stromal cell-derived factor-1, steel factor, and the bone marrow environment. Blood. 1998;91:100–110. [PubMed] [Google Scholar]
- 111.Broxmeyer HE, Orschell CM, Clapp DW, Hangoc G, Cooper S, Plett PA, Liles WC, Li X, Graham-Evans B, Campbell TB, Calandra G, Bridger G, Dale DC, Srour EF. Rapid mobilization of murine and human hematopoietic stem and progenitor cells with AMD3100, a CXCR4 antagonist. J Exp Med. 2005;201:1307–1318. doi: 10.1084/jem.20041385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Abraham M, Biyder K, Begin M, Wald H, Weiss ID, Galun E, Nagler A, Peled A. Enhanced unique pattern of hematopoietic cell mobilization induced by the CXCR4 antagonist 4F-benzoyl-TN14003. Stem Cells. 2007;25:2158–2166. doi: 10.1634/stemcells.2007-0161. [DOI] [PubMed] [Google Scholar]
- 113.Pusic I, DiPersio JF. Update on clinical experience with AMD3100, an SDF-1/CXCL12-CXCR4 inhibitor, in mobilization of hematopoietic stem and progenitor cells. Curr Opin Hematol. 2010;17:319–326. doi: 10.1097/MOH.0b013e328338b7d5. [DOI] [PubMed] [Google Scholar]
- 114.Choi HY, Yong CS, Yoo BK. Plerixafor for stem cell mobilization in patients with non-Hodgkin’s lymphoma and multiple myeloma. Ann Pharmacother. 2010;44:117–126. doi: 10.1345/aph.1M380. [DOI] [PubMed] [Google Scholar]
- 115.Calandra G, Bridger G, Fricker S. CXCR4 in clinical hematology. Curr Top Microbiol Immunol. 2010;341:173–191. doi: 10.1007/82_2010_26. [DOI] [PubMed] [Google Scholar]
- 116.Tchernychev B, Ren Y, Sachdev P, Janz JM, Haggis L, O’Shea A, McBride E, Looby R, Deng Q, McMurry T, Kazmi MA, Sakmar TP, Hunt S. r., Carlson KE. Discovery of a CXCR4 agonist pepducin that mobilizes bone marrow hematopoietic cells. Proc Nat Acad Sci USA. 2010;107:22255–22259. doi: 10.1073/pnas.1009633108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Jähnichen S, Blanchetot C, Maussang D, Gonzalez-Pajuelo M, Chow KY, Bosch L, De Vrieze S, Serruys B, Ulrichts H, Vandevelde W, Saunders M, De Haard HJ, Schols D, Leurs R, Vanlandschoot P, Verrips T, Smit MJ. CXCR4 nanobodies (VHH-based single variable domains) potently inhibit chemotaxis and HIV-1 replication and mobilize stem cells. Proc Nat Acad Sci USA. 2010;107:20565–20570. doi: 10.1073/pnas.1012865107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Chen Y, Zeng Z, Shi Y, Jacamo R, Ludin C, Dembowsky K, Frenette P, Konopleva M, Andreeff M. Targeting CXCR4, SDF1 and Beta-Adrenergic Receptors In the AML Microenvironment by Novel Antagonist POL6326, G-CSF and Isoproterenol. 53rd ASH Annual meeting and Exposition Abstract.2011. p. 2179. [Google Scholar]
- 119.Schmitt S, Weinhold N, Dembowsky K, Neben K, Witzens-Harig M, Braun M, Klemmer J, Wuchter P, Ludin C, Ho AD, Goldschmidt H. First Results of a Phase-II Study with the New CXCR4 Antagonist POL6326 to Mobilize Hematopoietic Stem Cells (HSC) In Multiple Myeloma (MM). 53rd ASH Annual meeting and Exposition Abstract.2011. p. 824. [Google Scholar]
- 120.Burger JA, Tsukada N, Burger M, Zvaifler NJ, Dell’Aquila M, Kipps TJ. Blood-derived nurse-like cells protect chronic lymphocytic leukemia B cells from spontaneous apoptosis through stromal cell-derived factor-1. Blood. 2000;96:2655–2663. [PubMed] [Google Scholar]
- 121.Burger JA, Burger M, Kipps TJ. Chronic lymphocytic leukemia B cells express functional CXCR4 chemokine receptors that mediate spontaneous migration beneath bone marrow stromal cells. Blood. 1999;94:3658–3667. [PubMed] [Google Scholar]
- 122.Konopleva M, Konoplev S, Hu W, Zaritskey AY, Afanasiev BV, Andreeff M. Stromal cells prevent apoptosis of AML cells by up-regulation of anti-apoptotic proteins. Leukemia. 2002;16:1713–1724. doi: 10.1038/sj.leu.2402608. [DOI] [PubMed] [Google Scholar]
- 123.Juarez J, Dela Pena A, Baraz R, Hewson J, Khoo M, Cisterne A, Fricker S, Fujii N, Bradstock KF, Bendall LJ. CXCR4 antagonists mobilize childhood acute lymphoblastic leukemia cells into the peripheral blood and inhibit engraftment. Leukemia. 2007;21:1249–1257. doi: 10.1038/sj.leu.2404684. [DOI] [PubMed] [Google Scholar]
- 124.Zeng Z, Samudio IJ, Munsell M, An J, Huang Z, Estey E, Andreeff M, Konopleva M. Inhibition of CXCR4 with the novel RCP168 peptide overcomes stroma-mediated chemoresistance in chronic and acute leukemias. Mol Cancer Ther. 2006;5:3113–3121. doi: 10.1158/1535-7163.MCT-06-0228. [DOI] [PubMed] [Google Scholar]
- 125.Nervi B, Ramirez P, Rettig MP, Uy GL, Holt MS, Ritchey JK, Prior JL, Piwnica-Worms D, Bridger G, Ley TJ, DiPersio JF. Chemosensitization of acute myeloid leukemia (AML) following mobilization by the CXCR4 antagonist AMD3100. Blood. 2009;113:6206–6214. doi: 10.1182/blood-2008-06-162123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Müller A, Homey B, Soto H, Ge N, Catron D, Buchanan ME, McClanahan T, Murphy E, Yuan W, Wagner SN, Barrera JL, Mohar A, Verástegui E, Zlotnik A. Involvement of chemokine receptors in breast cancer metastasis. Nature. 2001;410:50–56. doi: 10.1038/35065016. [DOI] [PubMed] [Google Scholar]
- 127.Hinton CV, Avraham S, Avraham HK. Role of the CXCR4/CXCL12 signaling axis in breast cancer metastasis to the brain. Clin Exp Metastasis. 2010;27:97–105. doi: 10.1007/s10585-008-9210-2. [DOI] [PubMed] [Google Scholar]
- 128.Huang EH, Singh B, Cristofanilli M, Gelovani J, Wei C, Vincent L, Cook KR, Lucci A. A CXCR4 antagonist CTCE-9908 inhibits primary tumor growth and metastasis of breast cancer. J Surg Res. 2009;155:231–236. doi: 10.1016/j.jss.2008.06.044. [DOI] [PubMed] [Google Scholar]
- 129.Richert MM, Vaidya KS, Mills CN, Wong D, Korz W, Hurst DR, Welch DR. Inhibition of CXCR4 by CTCE-9908 inhibits breast cancer metastasis to lung and bone. Oncol Rep. 2009;21:761–767. [PubMed] [Google Scholar]
- 130.Kwong J, Kulbe H, Wong D, Chakravarty P, Balkwill F. An antagonist of the chemokine receptor CXCR4 induces mitotic catastrophe in ovarian cancer cells. Mol Cancer Ther. 2009;8:1893–1905. doi: 10.1158/1535-7163.MCT-08-0966. [DOI] [PubMed] [Google Scholar]
- 131.Imitola J, Snyder EY, Khoury SJ. Genetic programs and responses of neural stem/progenitor cells during demyelination: potential insights into repair mechanisms in multiple sclerosis. Physiol Genomics. 2003;14:171–197. doi: 10.1152/physiolgenomics.00021.2002. [DOI] [PubMed] [Google Scholar]
- 132.Imitola J, Park KI, Teng YD, Nisim S, Lachyankar M, Ourednik J, Mueller FJ, Yiou R, Atala A, Sidman RL, Tuszynski M, Khoury SJ, Snyder EY. Stem cells: cross-talk and developmental programs. Philos Trans R Soc Lond B Biol Sci. 2004;359:823–837. doi: 10.1098/rstb.2004.1474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Park KI, Ourednik J, Ourednik V, Taylor RM, Aboody KS, Auguste KI, Lachyankar MB, Redmond DE, Snyder EY. Global gene and cell replacement strategies via stem cells. Gene Ther. 2002;9:613–624. doi: 10.1038/sj.gt.3301721. [DOI] [PubMed] [Google Scholar]
- 134.Park KI, Teng YD, Snyder EY. The injured brain interacts reciprocally with neural stem cells supported by scaffolds to reconstitute lost tissue. Nat Biotechnol. 2002;20:1111–1117. doi: 10.1038/nbt751. [DOI] [PubMed] [Google Scholar]
- 135.Snyder EY, Yoon C, Flax JD, Macklis JD. Multipotent neural precursors can differentiate toward replacement of neurons undergoing targeted apoptotic degeneration in adult mouse neocortex. Proc Natl Acad Sci U S A. 1997;94:11663–11668. doi: 10.1073/pnas.94.21.11663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Sharma GD, He J, Bazan HE. p38 and ERK1/2 coordinate cellular migration and proliferation in epithelial wound healing: evidence of cross-talk activation between MAP kinase cascades. J Biol Chem. 2003;278:21989–21997. doi: 10.1074/jbc.M302650200. [DOI] [PubMed] [Google Scholar]
- 137.Dechert MA, Holder JM, Gerthoffer WT. p21-activated kinase 1 participates in tracheal smooth muscle cell migration by signaling to p38 Mapk. Am J Physiol Cell Physiol. 2001;281:C123–132. doi: 10.1152/ajpcell.2001.281.1.C123. [DOI] [PubMed] [Google Scholar]
- 138.Cara DC, Kaur J, Forster M, McCafferty DM, Kubes P. Role of p38 mitogen-activated protein kinase in chemokine-induced emigration and chemotaxis in vivo. J Immunol. 2001;167:6552–6558. doi: 10.4049/jimmunol.167.11.6552. [DOI] [PubMed] [Google Scholar]
- 139.Suzuki Y, Rahman M, Mitsuya H. Diverse transcriptional response of CD4(+) T cells to stromal cell-derived factor (SDF)-1: cell survival promotion and priming effects of SDF-1 on CD4(+) T cells. J Immunol. 2001;167:3064–3073. doi: 10.4049/jimmunol.167.6.3064. [DOI] [PubMed] [Google Scholar]
- 140.Wong EV, Schaefer AW, Landreth G, Lemmon V. Involvement of p90rsk in neurite outgrowth mediated by the cell adhesion molecule L1. J Biol Chem. 1996;271:18217–18223. doi: 10.1074/jbc.271.30.18217. [DOI] [PubMed] [Google Scholar]
- 141.Li G, Gustafson-Brown C, Hanks SK, Nason K, Arbeit JM, Pogliano K, Wisdom RM, Johnson RS. c-Jun is essential for organization of the epidermal leading edge. Dev Cell. 2003;4:865–877. doi: 10.1016/s1534-5807(03)00159-x. [DOI] [PubMed] [Google Scholar]
- 142.Huang C, Rajfur Z, Borchers C, Schaller MD, Jacobson K. JNK phosphorylates paxillin and regulates cell migration. Nature. 2003;424:219–223. doi: 10.1038/nature01745. [DOI] [PubMed] [Google Scholar]
- 143.Abbott JD, Huang Y, Liu D, Hickey R, Krause DS, Giordano FJ. Stromal cell-derived factor-1alpha plays a critical role in stem cell recruitment to the heart after myocardial infarction but is not sufficient to induce homing in the absence of injury. Circulation. 2004;110:3300–3305. doi: 10.1161/01.CIR.0000147780.30124.CF. [DOI] [PubMed] [Google Scholar]
- 144.Askari AT, Unzek S, Popovic ZB, Goldman CK, Forudi F, Kiedrowski M, Rovner A, Ellis SG, Thomas JD, DiCorleto PE, Topol EJ, Penn MS. Effect of stromal-cell-derived factor 1 on stem-cell homing and tissue regeneration in ischaemic cardiomyopathy. Lancet. 2003;362:697–703. doi: 10.1016/S0140-6736(03)14232-8. [DOI] [PubMed] [Google Scholar]
- 145.Misao Y, Takemura G, Arai M, Ohno T, Onogi H, Takahashi T, Minatoguchi S, Fujiwara T, Fujiwara H. Importance of recruitment of bone marrow-derived CXCR4+ cells in post-infarct cardiac repair mediated by G-CSF. Cardiovasc Res. 2006;71:455–465. doi: 10.1016/j.cardiores.2006.05.002. [DOI] [PubMed] [Google Scholar]
- 146.Broxmeyer HE, Orschell CM, Clapp DW, Hangoc G, Cooper S, Plett PA, Liles WC, Li X, Graham-Evans B, Campbell TB, Calandra G, Bridger G, Dale DC, Srour EF. Rapid mobilization of murine and human hematopoietic stem and progenitor cells with AMD3100, a CXCR4 antagonist. J Exp Med. 2005;201:1307–1318. doi: 10.1084/jem.20041385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Capoccia BJ, Shepherd RM, Link DC. G-CSF and AMD3100 mobilize monocytes into the blood that stimulate angiogenesis in vivo through a paracrine mechanism. Blood. 2006;108:2438–2445. doi: 10.1182/blood-2006-04-013755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Morimoto CR, Fox PL, Anderson DA, DiMaggio P, Levine M, Schlossman H, F. S. Abnormalities in CD4+ T-lymphocyte subsets in inflammatory rheumatic diseases. Am J Med. 1988;84:817–825. doi: 10.1016/0002-9343(88)90058-7. [DOI] [PubMed] [Google Scholar]
- 149.Kohem CLB, Wisbey RI, Tortorella H, Lipsky C, Oppenheimer-Marks PE, N Enrichment of differentiated CD45RBdim,CD27-memory T cells in the peripheral blood, synovial fluid, and synovial tissue of patients with rheumatoid arthritis. Arthritis Rheum. 1996;39:844–854. doi: 10.1002/art.1780390518. [DOI] [PubMed] [Google Scholar]
- 150.Morita YY, Kawashima M, Harada M, Tsuji S, Shibuya K, Maruyama K, Makino K, H Flow cytometric single-cell analysis of cytokine production by CD4+ T cells in synovial tissue and peripheral blood from patients with rheumatoid arthritis. Arthritis Rheum. 1998;41:1669–1676. doi: 10.1002/1529-0131(199809)41:9<1669::AID-ART19>3.0.CO;2-G. [DOI] [PubMed] [Google Scholar]
- 151.Nanki T. a. L., E. P. Cytokine, activation marker, and chemokine receptor expression by individual CD4(+) memory T cells in rheumatoid arthritis synovium. Arthritis Res. 2000;2:415–423. doi: 10.1186/ar120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Nanki TH, El-Gabalawy K, Suson HS, Shi S, Girschick K, Yavuz HJ, Lipsky S, E. P. Stromal cell-derived factor-1-CXC chemokine receptor 4 interactions play a central role in CD4+ T cell accumulation in rheumatoid arthritis synovium. J Immunol. 2000;165:6590–6598. doi: 10.4049/jimmunol.165.11.6590. [DOI] [PubMed] [Google Scholar]
- 153.Dolhain R. J. t. H., Hoefakker NT, Tak S, de Ley PP, Claassen M, Breedveld E, Miltenburg FC, M. A. Increased expression of interferon (IFN)-gamma together with IFN-gamma receptor in the rheumatoid synovial membrane compared with synovium of patients with osteoarthritis. Br J Rheumatol. 1996;35:24–32. doi: 10.1093/rheumatology/35.1.24. [DOI] [PubMed] [Google Scholar]
- 154.Cohen SBK, Chu PD, Thomssen CQ, Webb H, Maini LM, Londei RN, Feldmann M, M. High level of interleukin-10 production by the activated T cell population within the rheumatoid synovial membrane. Arthritis Rheum. 1995;38:946–952. doi: 10.1002/art.1780380710. [DOI] [PubMed] [Google Scholar]
- 155.Smeets TJD, Miltenburg RJEM, de Kuiper AM, Breedveld R, Tak FC, P. P. Poor expression of T cell-derived cytokines and activation and proliferation markers in early rheumatoid synovial tissue. Clin Immunol Immunopathol. 1998;88:84–90. doi: 10.1006/clin.1998.4525. [DOI] [PubMed] [Google Scholar]
- 156.Grom AAM, Luyrink KJ, Emery L, Passo H, Glass MH, Bowlin DN, Edwards T, C. Patterns of expression of tumor necrosis factor alpha, tumor necrosis factor beta, and their receptors in synovia of patients with juvenile rheumatoid arthritis and juvenile spondylarthropathy. Arthritis Rheum. 1996;39:1703–1710. doi: 10.1002/art.1780391013. [DOI] [PubMed] [Google Scholar]
- 157.MacDonald KPN, Lipsky Y, Thomas PE, R. Functional CD40 ligand is expressed by T cells in rheumatoid arthritis. J Clin Invest. 1997;100:2404–2414. doi: 10.1172/JCI119781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Buckley CD, Amft N, Bradfield PF, Pilling D, Ross E, Arenzana-Seisdedos F, Amara A, Curnow SJ, Lord JM, Scheel-Toellner D, Salmon M. Persistent induction of the chemokine receptor CXCR4 by TGF-beta 1 on synovial T cells contributes to their accumulation within the rheumatoid synovium. J Immunol. 2000;165:3423–3429. doi: 10.4049/jimmunol.165.6.3423. [DOI] [PubMed] [Google Scholar]
- 159.Bradfield PF, Amft N, Vernon-Wilson E, Exley AE, Parsonage G, Rainger GE, Nash GB, Thomas AM, Simmons DL, Salmon M, Buckley CD. Rheumatoid fibroblast-like synoviocytes overexpress the chemokine stromal cell-derived factor 1 (CXCL12), which supports distinct patterns and rates of CD4+ and CD8+ T cell migration within synovial tissue. Arthritis Rheum. 2003;48:2472–2482. doi: 10.1002/art.11219. [DOI] [PubMed] [Google Scholar]
- 160.Matthys P, Hatse S, Vermeire K, Wuyts A, Bridger G, Henson GW, De Clercq E, Billiau A, Schols D. AMD3100, a potent and specific antagonist of the stromal cell-derived factor-1 chemokine receptor CXCR4, inhibits autoimmune joint inflammation in IFN-gamma receptor-deficient mice. J Immunol. 2001;167:4686–4692. doi: 10.4049/jimmunol.167.8.4686. [DOI] [PubMed] [Google Scholar]
- 161.De Klerck B, Geboes L, Hatse S, Kelchtermans H, Meyvis Y, Vermeire K, Bridger G, Billiau A, Schols D, Matthys P. Pro-inflammatory properties of stromal cell-derived factor-1 (CXCL12) in collagen-induced arthritis. Arthritis Res Ther. 2005;7:R1208–1220. doi: 10.1186/ar1806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Tamamura H, Fujii N. The therapeutic potential of CXCR4 antagonists in the treatment of HIV infection, cancer metastasis and rheumatoid arthritis. Expert Opin Ther Targets. 2005;9:1267–1282. doi: 10.1517/14728222.9.6.1267. [DOI] [PubMed] [Google Scholar]
- 163.Tamamura H, Tsutsumi H, Masuno H, Fujii N. Development of low molecular weight CXCR4 antagonists by exploratory structural tuning of cyclic tetra- and pentapeptide-scaffolds towards the treatment of HIV infection, cancer metastasis and rheumatoid arthritis. Curr Med Chem. 2007;14:93–102. doi: 10.2174/092986707779313499. [DOI] [PubMed] [Google Scholar]
- 164.Busse WWL, F. R. Advances in immunology: asthma. N Engl J Med. 2001;344:350–362. doi: 10.1056/NEJM200102013440507. [DOI] [PubMed] [Google Scholar]
- 165.O’Byrne PI, Parameswaran M, K. The trials and tribulations of IL-5, eosinophils, and allergic asthma. J Allergy Clin Immunol. 2001;108:503–508. doi: 10.1067/mai.2001.119149. [DOI] [PubMed] [Google Scholar]
- 166.Leckie M. J. t. B., Khan A, Diamant J, O’Connor Z, Walls BJ, Mathur CM, Cowley AK, Chung HC, Djukanovic KF, Hansel R, Holgate TT, Sterk ST, Barnes PJ, J. P. Effects of an interleukin-5 blocking monoclonal antibody on eosinophils, airway hyper-responsiveness, and the late asthmatic response. Lancet. 2000;356:2144–2148. doi: 10.1016/s0140-6736(00)03496-6. [DOI] [PubMed] [Google Scholar]
- 167.Gonzalo JA, Lloyd CM, Peled A, Delaney T, Coyle AJ, Gutierrez-Ramos JC. Critical involvement of the chemotactic axis CXCR4/stromal cell-derived factor-1 alpha in the inflammatory component of allergic airway disease. J Immunol. 2000;165:499–508. doi: 10.4049/jimmunol.165.1.499. [DOI] [PubMed] [Google Scholar]
- 168.Jourdan P, Vendrell JP, Huguet MF, Segondy M, Bousquet J, Pène J, Yssel H. Cytokines and cell surface molecules independently induce CXCR4 expression on CD4+ CCR7+ human memory T cells. J Immunol. 2000;165:716–724. doi: 10.4049/jimmunol.165.2.716. [DOI] [PubMed] [Google Scholar]
- 169.Williams CM, Galli SJ. Mast cells can amplify airway reactivity and features of chronic inflammation in an asthma model in mice. J Exp Med. 2000;192:455–462. doi: 10.1084/jem.192.3.455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Lukacs NW, Berlin A, Schols D, Skerlj RT, Bridger GJ. AMD3100, a CxCR4 antagonist, attenuates allergic lung inflammation and airway hyperreactivity. Am J Pathol. 2002;160:1353–1360. doi: 10.1016/S0002-9440(10)62562-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Hu JS, Freeman CM, Stolberg VR, Chiu BC, Bridger GJ, Fricker SP, Lukacs NW, Chensue SW. AMD3465, a novel CXCR4 receptor antagonist, abrogates schistosomal antigen-elicited (type-2) pulmonary granuloma formation. Am J Pathol. 2006;169:424–432. doi: 10.2353/ajpath.2006.051234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Ghosh S, Nie A, An J, Huang Z. Structure-based virtual screening of chemical libraries for drug discovery. Curr Opin Chem Biol. 2006;10:194–202. doi: 10.1016/j.cbpa.2006.04.002. [DOI] [PubMed] [Google Scholar]
- 173.Zhou NL, Luo Z, Fan J, Cayabyab X, Hiraoka M, Liu M, Han D, Pesavento X, Dong J, Wang CZ, An Y, Kaji J, Sodroski H, G. J. Exploring the stereochemistry of CXCR4-peptide recognition and inhibiting HIV-1 entry with D-peptides derived from chemokines. J Biol Chem. 2002;277:17476–17485. doi: 10.1074/jbc.M202063200. [DOI] [PubMed] [Google Scholar]
- 174.Dong CZ, Kumar S, Choi WT, Madani N, Tian S, An J, Sodroski JG, Huang Z. Different stereochemical requirements for CXCR4 binding and signaling functions as revealed by an anti-HIV, D-amino acid-containing SMM-chemokine ligand. J Med Chem. 2005;48:7923–7924. doi: 10.1021/jm050829u. [DOI] [PubMed] [Google Scholar]
- 175.Wang J, Liu D, Zhang Z, Shan S, Han X, Srinivasula SM, Croce CM, Alnemri ES, Huang Z. Structure-based discovery of a novel organic compound that binds Bcl-2 protein and induces apoptosis of tumor cells. Proc Natl Acad Sci USA. 2000;97:7124–7129. doi: 10.1073/pnas.97.13.7124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Mizutani MY, Itai A. Efficient method for high-throughput virtual screening based on flexible docking: discovery of novel acetylcholinesterase inhibitors. J Med Chem. 2004;47:4818–4828. doi: 10.1021/jm030605g. [DOI] [PubMed] [Google Scholar]
- 177.Irwin JJ, Raushel FM, Shoichet BK. Virtual screening against metalloenzymes for inhibitors and substrates. Biochem. 2005;44:12316–12328. doi: 10.1021/bi050801k. [DOI] [PubMed] [Google Scholar]
- 178.Becker OM, Marantz Y, Shacham S, Inbal B, Heifetz A, Kalid O, Bar-Haim S, Warshaviak D, Fichman M, Noiman S. G protein-coupled receptors: in silico drug discovery in 3D. Proc Natl Acad Sci U S A. 2004;101:11304–11309. doi: 10.1073/pnas.0401862101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Ma Q, Jones D, Borghesani PR, Segal RA, Nagasawa T, Kishimoto T, Bronsoni RT, Springer TA. Impaired B-lymphopoiesis, myelopoiesis, and derailed cerebellar neuron migration in CXCR4- and SDF-1-deficient mice. Proc Natl Acad Sci USA. 1998;95:9448–9453. doi: 10.1073/pnas.95.16.9448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Imitola J, Raddassi K, Park KI, Mueller FJ, Nieto M, Teng YD, Frenkel D, Li J, Sidman RL, Walsh CA, Snyder EY, Khoury SJ. Directed migration of neural stem cells to sites of CNS injury by the stromal cell-derived factor 1alpha/CXC chemokine receptor 4 pathway. Proc Natl Acad Sci U S A. 2004;101:18117–18122. doi: 10.1073/pnas.0408258102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Choi WT, Kaul M, Kumar S, Wang J, Kumar IM, Dong CZ, An J, Lipton SA, Huang Z. Neuronal apoptotic signaling pathways probed and intervened by synthetically and modularly modified (SMM) chemokines. J Biol Chem. 2007;282:7154–7163. doi: 10.1074/jbc.M611599200. [DOI] [PubMed] [Google Scholar]
- 182.Pelus LM, Bian H, Fukuda S, Wong D, Merzouk A, Salari H. The CXCR4 agonist peptide, CTCE-0021, rapidly mobilizes polymorphonuclear neutrophils and hematopoietic progenitor cells into peripheral blood and synergizes with granulocyte colony-stimulating factor. Exp Hematol. 2005;33:295–307. doi: 10.1016/j.exphem.2004.11.008. [DOI] [PubMed] [Google Scholar]
- 183.Tang J, Wang J, Yang J, Kong X. Adenovirus-mediated stromal cell-derived-factor-1alpha gene transfer induces cardiac preservation after infarction via angiogenesis of CD133+ stem cells and anti-apoptosis. Interact Cardiovasc Thorac Surg. 2008;7:767–770. doi: 10.1510/icvts.2007.169896. [DOI] [PubMed] [Google Scholar]
- 184.Tang J, Wang J, Song H, Huang Y, Yang J, Kong X, Guo L, Zheng F, Zhang L. Adenovirus-mediated stromal cell-derived factor-1 alpha gene transfer improves cardiac structure and function after experimental myocardial infarction through angiogenic and antifibrotic actions. Mol Biol Rep. 2010;37:1957–1969. doi: 10.1007/s11033-009-9642-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Lefrançois M, Lefebvre MR, Saint-Onge G, Boulais PE, Lamothe S, Leduc R, Lavigne P, Heveker N, Escher E. Agonists for the chemokine receptor CXCR4. ACS Med Chem Lett. 2011;2:597–602. doi: 10.1021/ml200084n. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Siciliano SJ, Rollins TE, DeMartino J, Konteatis Z, Malkowitz L, Riper GV, Bondy S, Rosen H, Springer MS. Two-site binding of C5a by its receptor: An alternative binding paradigm for G protein-coupled receptors. Proc Natl Acad Sci USA. 1994;91:1214–1218. doi: 10.1073/pnas.91.4.1214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Gupta SK, Pillarisetti K, Thomas RA, Aiyar N. Pharmacological evidence for complex and multiple site interaction of CXCR4 with SDF-1alpha: implications for development of selective CXCR4 antagonists. Immunol Lett. 2001;78:29–34. doi: 10.1016/s0165-2478(01)00228-0. [DOI] [PubMed] [Google Scholar]
- 188.Liu D, Madani N, Li Y, Cao R, Choi WT, Kumar S, Dong C, Russell JD, Lefebure CR, An J, Wilson S, Gao YG, Pallansch LA, Sodroski JG, Huang Z. Crystal structure and structural mechanism of a novel anti-human immunodeficiency virus and D-amino acid-containing chemokine. J Virol. 2007;81:11489–11498. doi: 10.1128/JVI.02845-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Babcock GJ, Farzan M, Sodroski J. Ligand-independent dimerization of CXCR4, a principal HIV-1 coreceptor. J Biol Chem. 2003;278:3378–3385. doi: 10.1074/jbc.M210140200. [DOI] [PubMed] [Google Scholar]
- 190.Percherancier Y, Berchiche YA, Slight I, Volkmer-Engert R, Tamamura H, Fujii N, Bouvier M, Heveker N. Bioluminescence resonance energy transfer reveals ligand-induced conformational changes in CXCR4 homo- and heterodimers. J Biol Chem. 2005;280:9895–9903. doi: 10.1074/jbc.M411151200. [DOI] [PubMed] [Google Scholar]
- 191.Hernandez PA, Gorlin RJ, Lukens JN, Taniuchi S, Bohinjec J, Francois F, Klotman ME, Diaz GA. Mutations in the chemokine receptor gene CXCR4 are associated with WHIM syndrome, a combined immunodeficiency disease. Nat Genet. 2003;34:70–74. doi: 10.1038/ng1149. [DOI] [PubMed] [Google Scholar]
- 192.Hamatake M, Aoki T, Futahashi Y, Urano E, Yamamoto N, Komano J. Ligand-independent higher-order multimerization of CXCR4, a G-protein-coupled chemokine receptor involved in targeted metastasis. Cancer Sci. 2009;100:95–102. doi: 10.1111/j.1349-7006.2008.00997.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Daniels DJ, Lenard NR, Etienne CL, Law PY, Roerig SC, Portoghese PS. Opioid-induced tolerance and dependence in mice is modulated by the distance between pharmacophores in a bivalent ligand series. Proc Nat Acad Sci USA. 2005;102:19208–19213. doi: 10.1073/pnas.0506627102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.George SR, O’Dowd BF, Lee SP. G-protein-coupled receptor oligomerization and its potential for drug discovery. Nat Rev Drug Discov. 2002;1:808–820. doi: 10.1038/nrd913. [DOI] [PubMed] [Google Scholar]
- 195.Messer WSJ. Bivalent ligands for G protein-coupled receptors. Curr Pharm Des. 2004;10:2015–2020. doi: 10.2174/1381612043384213. [DOI] [PubMed] [Google Scholar]
- 196.Berque-Bestel I, Lezoualc’h F, Jockers R. Bivalent ligands as specific pharmacological tools for G protein-coupled receptor dimers. Curr Drug Discov Technol. 2008;5:312–318. doi: 10.2174/157016308786733591. [DOI] [PubMed] [Google Scholar]
- 197.Tanaka T, Nomura W, Narumi T, Masuda A, Tamamura H. Bivalent ligands of CXCR4 with rigid linkers for elucidation of the dimerization state in cells. J Am Chem Soc. 2010;132:15899–15901. doi: 10.1021/ja107447w. [DOI] [PubMed] [Google Scholar]
- 198.Sohy D, Yano H, de Nadai P, Urizar E, Guillabert A, Javitch JA, Parmentier M, Springael JY. Hetero-oligomerization of CCR2, CCR5, and CXCR4 and the protean effects of "selective" antagonists. J Biol Chem. 2009;284:31270–31279. doi: 10.1074/jbc.M109.054809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Sierro F, Biben C, Martínez-Muñoz L, Mellado M, Ransohoff RM, Li M, Woehl B, Leung H, Groom J, Batten M, Harvey RP, Martínez A, C., Mackay CR, Mackay F. Disrupted cardiac development but normal hematopoiesis in mice deficient in the second CXCL12/SDF-1 receptor, CXCR7. Proc Nat Acad Sci USA. 2007;104:14759–14764. doi: 10.1073/pnas.0702229104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.De Clercq EY, Pauwels N, Baba R, Schols M, Nakashima D, Balzarini H, Debyser J, Murrer Z, Schwartz BA, Thornton D, Bridger D, Fricker G, Henson S, Abrams G, M., Picker D. Potent and selective inhibition of human immunodeficiency virus (HIV)-1 and HIV-2 replication by a class of bicyclams interacting with a viral uncoating event. Proc Natl Acad Sci U S A. 1992;89:5286–5290. doi: 10.1073/pnas.89.12.5286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Tamamura H, Xu Y, Hattori T, Zhang X, Arakaki R, Kanbara K, Omagari A, Otaka A, Ibuka T, Yamamoto N, Nakashima H, Fujii N. A low-molecular-weight inhibitor against the chemokine receptor CXCR4: a strong anti-HIV peptide T140. Biochem Biophys Res Comm. 1998;253:877–882. doi: 10.1006/bbrc.1998.9871. [DOI] [PubMed] [Google Scholar]
- 202.Demmer O, Dijkgraaf I, Schumacher U, Marinelli L, Cosconati S, Wester HJ, Kessler H. Design, synthesis and functionalization of dimeric peptides targetting chemokine receptor CXCR4. J Med Chem. 2011 doi: 10.1021/jm2009716. In press. [DOI] [PubMed] [Google Scholar]
- 203.Cabrera C, Gutiérrez A, Blanco J, Barretina J, Litovchick A, Lapidot A, Evdokimov AG, Clotet B, Esté JA. Anti-human immunodeficiency virus activity of novel aminoglycoside-arginine conjugates at early stages of infection. AIDS Res Hum Retroviruses. 2000;16:627–34. doi: 10.1089/088922200308855. [DOI] [PubMed] [Google Scholar]