

Abstract
Objectives
The Helicobacter pylori virulence-associated genes in hepatobiliary patients, including vacA, iceA, babA2, cagA and cagE, have not been reported. The aim of this study was to investigate these genes and the association of those and the clinical outcomes in hepatobiliary diseases.
Methods
Eighty H. pylori-PCR-positive cases were obtained from hepatobiliary patients, representing both cholangiocarcinoma (CCA) (n = 58) and cholelithiasis (n = 22). The diversity of virulence genes was examined by polymerase chain reaction and DNA sequencing. Phylogenetic analysis of cagA was determined using the maximum parsimony method.
Results
The vacAs1a + c/m1, iceA1 and babA2 genes were the most predominant genotypes in both CCA and cholelithiasis patients. The cagA and cagE genes were found significantly more frequently in patients with CCA than those with cholelithiasis (P < 0.05). The cagA positive samples were the Western-type cagA and showed that almost all of the detected sequences in Thai hepatobiliary and Thai gastric cancer patients were classified in the same cluster but separated from the cluster of Japan and other countries.
Conclusions
The cagA and cagE genes may be associated in the pathogenesis of hepatobiliary diseases, especially of CCA. Besides the bacterial variation, other host factors may be involved in the pathogenesis of hepatobiliary cancer.
Introduction
Helicobacter pylori is well known as a causative agent of gastric adenocarcinoma.1Helicobacter pylori is genetically diverse with a spectrum of virulence factors so infection with different H. pylori strains will lead to diverse clinical outcomes.2 The virulence factors of H. pylori include vacA, iceA, babA2, cagA and cagE, which play various and important roles in pathogenesis.3–5
Cholangiocarcinoma (CCA) is an adenocarcinoma of the bile duct epithelium and this primary liver cancer has a high incidence in Northeast Thailand.6 The presence of H. pylori in patients with hepatobiliary diseases has been investigated and the virulence-associated genes, vacA and cagA, detected.7,8 Previous research confirmed an association of H. pylori and CCA: the former was observed to accelerate biliary inflammation and proliferation.9 However, the association between H. pylori virulence-associated genes (vacA, iceA, babA2, cagA and cagE) and clinical outcomes of hepatobiliary diseases has not been reported, particularly vis-à-vis CCA.
The vacuolating cytotoxin A gene (vacA) is present in all H. pylori strains10 and is associated with the severity of gastrointestinal diseases.11 The vacA gene comprises diverse signalling parts (s1a, s1b, s1c and s2) and a middle region (m1 and m2). The activity of this toxin is higher in the s1/m1 than the s1/m2 subtype and low or absent in the s2/m2 subtype.12
The induction by contact with the epithelium gene (iceA) has been identified as the up-regulator after adhering to the epithelial cells.13 The iceA gene has two major allelic sequence variants, designated iceA1 and iceA2.14Helicobacter pylori possessing the iceA1 gene are significantly more associated with peptic ulcers and related to the severity of gastric inflammation.3,13
The blood group antigen-binding adhesion gene (babA) encodes membrane-bound adhesin that binds the blood-group antigen Lewisb, present on gastric epithelial cells.15 There are three bab alleles (babA1, babA2 and babB) of H. pylori, but only the babA2 gene shows Lewisb-binding activity.16 The presence of the babA2 gene is associated with duodenal ulceration and gastric cancer, whereas strains negative for babA2 are associated with gastritis.16
The cytotoxin-associated gene A (cagA) is located at the end of the cagA pathogenicity islands (cag PAI) and is considered an important marker of H. pylori.17Helicobacter pylori cagA-positive strains have been observed to be more virulent than the H. pylori cagA-negative strains.18 The CagA protein is associated with mucosal inflammation by increasing interleukin (IL)-8 secretion and stimulation of cell proliferation.18–20 The EPIYA motifs (Glu-Pro-Ile-Tyr-Ala) are located at the 3′ region of the CagA protein and these motifs are subdivided to EPIYA A, B, C and D; based on different sequences in front of, and behind, the motif.21 EPIYA A and B are mostly present in all CagA proteins, whereas EPIYA C and D are present in the CagA Western and East Asian type, respectively.17 Infection with East Asian positive strains is associated with the severity of inflammation among patients with gastritis than those with Western positive strains.22 Moreover, the East Asian CagA showed a more significant effect on growth of the gastric mucosa than the Western CagA.23
Inside the cag PAI, cagE is a marker and essential for translocation and phosphorylation of CagA.24 Previous research showed that H. pylori cagE positive strains are associated with the severity of clinical outcomes of gastrointestinal diseases.25
In spite of research into the prevalence of H. pylori in hepatobiliary diseases, an association between H. pylori virulence-associated genes and clinical outcomes in hepatobiliary diseases has not been previously reported. We, therefore, investigated the genotyping of vacA, iceA, babA2, cagA and cagE in H. pylori-infected hepatobiliary patients (specifically those with CCA and cholelithiasis) and the association of those genes and clinical outcomes. In order to clarify the association between the diversity of cagA and the frequency of gastro-hepatobiliary diseases in various countries, the diversity of cagA in H. pylori strains from gastroduodenal diseases of a selection of countries was compared with that of hepatobiliary diseases using phylogenetic analysis.
Methods
Patients and clinical samples
Bile samples were obtained from 140 patients operated on at Srinagarind Hospital, Faculty of Medicine, Khon Kaen University. There were 80 hepatobiliary patients considered as H. pylori-positive by PCR assay of whom 58 had CCA and 22 had cholelithiasis. The 16 cases in the control group were obtained from autopsies, which indicated that four cases were H. pylori PCR positive. Patients with gastrointestinal diseases and hepatitis virus infection were excluded from the study. This study was approved by the Ethics Committee of Khon Kaen University (HE 450525) and performed in accordance with the Helsinki Declaration. All of the patients provided informed consent before participating in the study.
DNA extraction
Bile samples were diluted with 1 volume of sterile phosphate-buffered saline (PBS) and centrifuged at 13 000 g before collecting the bile pellet, which was re-suspended with lysis buffer [containing 4 µl of proteinase K (20 µg/ml)] and incubated at 55°C for 3 h. The mixture was subsequently incubated with 4 µl RNase (25 mg/ml) at 37°C for 1 h. Then, 300 µl protein precipitation solution was added and centrifuged at 13 000 g for 5 min. Genomic DNA in a supernatant was precipitated by 2 volumes of absolute ethanol. The precipitated DNA in the supernatant was purified in a DNA-purified column and washed two times with washing buffer. Finally, the DNA was eluted using a buffer and stored at −20°C until used.
PCR amplification of ureA, vacA, iceA, babA2, cagA and cagE
The primer sequences and PCR condition for ureA (used for detection and confirmation of H. pylori), the virulence genes and genotyping of vacA, iceA, babA2, cagA and cagE were designed based on published research3,26–34 with a modification to the PCR mixture constituents and conditions (Table 1). The PCR was performed in a total volume of 50 µl containing 1× PCR buffer (RBC Bioscience, Taipei, Taiwan), 0.2 mM dNTP, 0.2 µM of each primer (0.5 µM for babA2 and cagA sequencing), 1.25 units of Taq DNA polymerase (RBC Bioscience) and 500 ng of genomic DNA in each sample. The PCR assay was performed using a GeneAmp PCR system 9600 (Perkin Elmer, Waltham, MA, USA). The amplified PCR products were determined by electrophoresis on 1.5–2% agarose gel. After staining with ethidium bromide, bands were visualized under a UV illuminator.
Table 1.
Primer sequences and PCR conditions
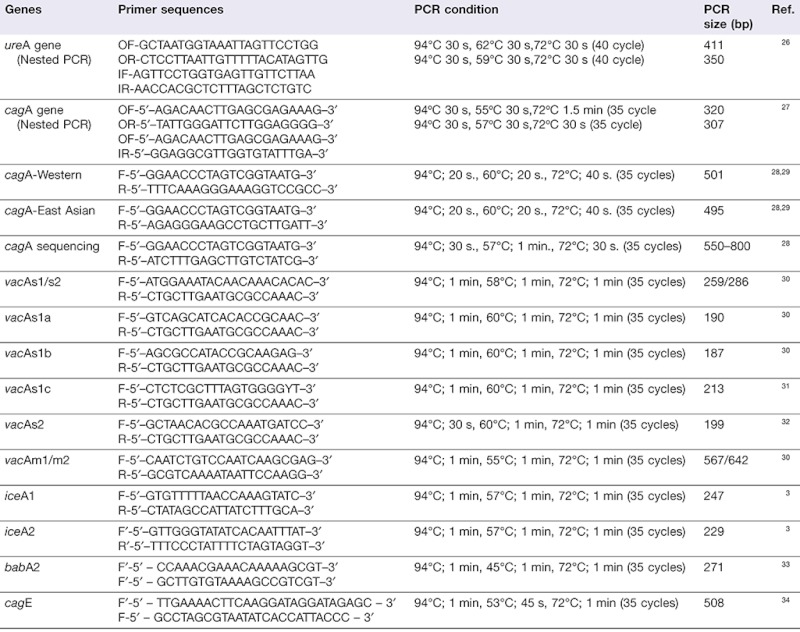 |
DNA sequencing and EPIYA motifs analysis
The amplicons of partial cagA were purified and sequenced using a DYEnamic ET Dye Terminator Cycle Sequencing kit from the MegaBace 1000 DNA Analysis System (GE Healthcare Life Sciences, Little Chalfont, UK). The cagA sequences were subsequently analysed for EPIYA motifs by translating the amino acid sequence using a BMC Search Launcher program (Baylor College of Medicine).
Phylogenetic analysis of cagA
In order to clarify the phylogenetic relationship of cagA, H. pylori cagA+ strains from Thai patients with hepatobiliary diseases and H. pylori cagA+ strains from those with gastroduodenal diseases, reports from Thailand (northeastern), Japan, China, Vietnam, Iran, USA and the Philippines were reviewed. Partial sequences of cagA were aligned using the clustalw program. Of the Thai patients with hepatobiliary disease, 17 cases of CCA and 1 case of cholelithiasis with cagA positive strains were included. Additionally, we simultaneously analysed the DNA sequences of Western type cagA from ethnic northeast Thai patients with gastric cancer (GC196 and GC145). Of the Japanese patients, three strains of Western type cagA were included (AB190942, AB190948, and AB190950). Other strains included a Chinese (AF247651), a Vietnamese (FJ798969), an Iranian (FJ849781), two USA American mixed race grouping (AB015414 and AB015415) and two Filipino (GU173858 and GU173862) were also analysed. A phylogenetic tree was constructed using the maximum parsimony method. Reliability of the tree was confirmed using bootstrap re-sampling and reconstruction with 1000 replicates (using MEGA software version 4.0; Center for Evolutionary Medicine and Informatics, Tempe, AZ, USA)).
Statistical analysis
Chi-square (χ2) or Fisher's exact test were used to analyse the categorical data in the CCA, cholelithiasis and the control group. A P-value less than 0.05 were considered as statistically significant.
Results
Helicobacter pylori-infected patients were evaluated for the relationship of age and gender (Table 2). CCA had a correlation with age (41–60 years) and male gender whereas cholelithiasis was correlated with female gender (P < 0.05).
Table 2.
Distribution of 87 hepatobiliary patients, according to age and gender
| Hepatobiliary diseases | Total | ||
|---|---|---|---|
| CCA | Cholelithiasis | n = 80(%) | |
| n = 58(%) | n = 22(%) | ||
| Age (years) | |||
| <20 | 0 | 2 (9.1) | 2 (2.5) |
| 20–40 | 0 | 1 (4.5) | 1 (1.3) |
| 41–60 | 39 (67.2)* | 7 (31.8) | 46 (57.5) |
| >60 | 19 (32.8) | 12 (54.5) | 31 (38.8) |
| Gender | |||
| Male (M) | 35 (60.3)* | 6 (27.3) | 41 (51.3) |
| Female (F) | 23 (39.7) | 16 (72.7)* | 39 (48.7) |
Significant difference between CCA and cholelithiasis (P < 0.05).
Prevalence of vacA genotypes
The prevalence of vacA, iceA, babA2, cagA and cagE was examined in 80 of the H. pylori-infected patients with hepatobiliary diseases (CCA and cholelithiasis). All of the vacA genes detected in the CCA, cholelithiasis and control groups were the s1 type (100%). Among the vacA s1 subtypes, vacA s1a+c was more prevalent than the other subtypes in both the CCA and cholelithiasis patients (Table 3). Among those in the control group, vacA s1a was encountered more frequently than any other subtype. A comparison among the three groups, revealed no significant relationship vis-à-vis the vacA s1 subtype.
Table 3.
Distribution of Helicobacter pylori vacA, iceA, babA2, cagA and cagE genotypes in hepatobiliary diseases
| Genes | CCA | Cholelithiasis | Control group | Total (CCA and cholelithiasis) |
|---|---|---|---|---|
| n = 58 (%) | n = 22 (%) | n = 4 (%) | n = 80 (%) | |
| vacA s types | ||||
| s1 | 58 (100) | 22 (100) | 4 (100) | 80 (100) |
| s1a | 15 (25.9) | 7 (31.8) | 2 (50) | 22 (27.5) |
| s1c | 10 (17.2) | 3 (13.6) | 1 (25) | 13 (16.3) |
| s1a+c | 33 (56.9) | 12 (54.5) | 1 (25) | 45 (56.2) |
| s2 | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| vacA m types | ||||
| m1 | 53 (91.4) | 19 (86.4) | 4 (100) | 72 (90.0) |
| m2 | 5 (8.6) | 3 (13.6) | 0 (0) | 8 (10.0) |
| iceA | ||||
| iceA1 | 29 (50.0) | 8 (36.4) | 1 (25) | 37 (46.3) |
| iceA2 | 7 (12.1) | 2 (9.1) | 0 (0) | 9 (11.3) |
| iceA1+A2 | 5 (8.6) | 2 (9.1) | 0 (0) | 7 (8.8) |
| iceA - | 17 (29.2) | 10 (45.5) | 3 (75) | 27 (33.8) |
| babA2 | ||||
| babA2+ | 41 (70.7) | 12 (54.5) | 1 (25) | 53 (66.2) |
| babA2- | 17 (29.3) | 10 (45.5) | 3 (75) | 27 (33.8) |
| cagA+ | 21 (36.2)a | 2 (9.1) | 0 (0) | 23 (28.8) |
| Western type | 21 (100) | 2 (100) | ND | 23 (100) |
| East asian type | 0 (0) | 0 (0) | ND | 0 (0) |
| cagA- | 37 (63.8) | 20 (90.9) | 4 (100) | 57 (71.2) |
| cagE+ | 17 (29.3)b | 1 (4.5) | 0 (0) | 18 (22.5) |
| cagE- | 41 (70.7) | 21 (95.5) | 4 (100) | 62 (77.5) |
| cagA+/cagE+ | 17 (29.3)c | 1 (4.5) | 0 (0) | 18 (22.5) |
| cagA+/cagE- | 4 (6.9) | 1 (4.5) | 0 (0) | 5 (6.3) |
| cagA-/cagE+ | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| cagA-/cagE- | 37 (63.8) | 20 (91.0) | 0 (0) | 57 (71.2) |
Significant difference of cagA between CCA and cholelithiasis (P < 0.05).
Significant difference of cagE between CCA and cholelithiasis (P < 0.05).
Significant difference of cagA+/cagE+ between CCA and cholelithiasis (P < 0.05).
The vacA m1 gene was found significantly more frequently than the vacA m2 alleles in patients with either CCA or cholelithiasis. The vacA m1 gene was found only in the control group; however, the difference between these three groups was not significant (Table 3). The combined vacA genotype (viz., vacA s1a + c/m1) was the predominant subtype among patients with either CCA or cholelithiasis and there was no statistically significant association between the combination vacA subtype and clinical outcome (data not shown).
Prevalence of iceA genotypes
The iceA1 allele was the most commonly found among hepatobiliary patients (46.3%; 37/80). Among CCA patients, the respective iceA1 and iceA2 genes were detected in 50.0% (29/58) and 12.1% (7/58). Among cholelithiasis patients, the respective iceA1 and iceA2 genes were detected in 36.4% (8/22) and 9.1% (2/22). The combined iceA1 and iceA2 genes were detected in patients with both CCA and cholelithiasis [8.6% (5/58) and 9.1% (2/22), respectively], but the difference between the two was not significant neither was the clinical outcome (Table 3).
Prevalence of the babA2 gene
The babA2 allele was found in 53 (66.2%) of the 80 hepatobiliary patients: in 70.7% (41/58), 54.5% (12/22) and 25.0% (1/4) of patients in the CCA, cholelithiasis and control group, respectively. There was no significant difference among the patient groups (Table 3).
Prevalence of the cagA genotypes
The cagA gene was detected in 28.8% (23/80) of the H. pylori-infected hepatobiliary patients: in 36.2% (21/58) of those with CCA and 9.1% (2/22) with cholelithiasis. The difference between the groups was statistically significant (P < 0.05). The control group showed cagA negative results (Table 3). Using DNA sequencing and amino acid analysis, we found that 18 of the 23 samples contained EPIYA-ABC, which is the Western type of cagA (Fig. 1). Additionally, 5 of the 23 contained EPIYA-AC (CCA2, CCA7, and CCA15) and EPIYA-BC (CCA3 and GS2). The repeats of each EPIYA motif were not found.
Figure 1.

Examples of amino acid sequences of Western-type CagA showing EPIYA-ABC in CCA (CCA10 and CCA17), EPIYA-AC (CCA2) and EPIYA-BC (CCA3)
Prevalence of the cagE gene
The presence of the cagE gene was found in 22.5% (18/80) of hepatobiliary patients: 29.3% (17/64) of those with CCA and 4.5% (1/22) with cholelithiasis. The difference was statistically significant (P < 0.05) (Table 3). The combination of cagA+ and cagE+ was detected in 29.3% (17/58) of patients with CCA and 4.5% (1/22) of those with cholelithiasis. The difference was statistically significant (P < 0.05) (Table 3).
Combination genotypes of H. pylori
The relationship of the combination genotype in hepatobiliary diseases was analysed (data not shown). The rank of the combinations found were: (i) vacA s1a+c/m1, iceA−, babA2−, cagA−, cagE− (17.5%); (ii) vacA s1a+c/m1, iceA1+, babA2+, cagA−, cagE− (12.5%); (iii) vacA s1a+c/m1, iceA1+, babA2+, cagA+, cagE+ (11.3%); and (iv) vacA s1a/m1, iceA−, babA2−, cagA−, cagE− (8.8%) followed by the other combinations (data not shown). The difference among the the patient groups was not statistically significant.
Phylogenetic analysis of the partial cagA gene
The genetic relationships of the Western type cagA (EPIYA-ABC), among the 18 Thai hepatobiliary patients (CCA = 17 and cholelithiasis = 1), the Thai (northeastern) patients with gastric cancer and the reference strains detected in gastroduodenal diseases in Japan, China, the USA and the Philippines, were determined using maximum parsimony (Fig. 2). The results showed that the partial DNA sequences of the Western type cagA for each region were markedly distinguishable. The sequences of the Western type cagA detected among patients from Japan, China, Iran and the USA were grouped in the same cluster, indicating a genetic similarity. By contrast, the Western type cagA detected among Thai patients with hepatobiliary diseases formed a different cluster: only one strain among the Thai patients (CCA8) was genetically closer to those other groups. Furthermore, we found that the Western type cagA sequences among the Thai patients with hepatobiliary disease were included in the same cluster as the Western type cagA sequences found among Thai patients with gastric cancer (GC45 and GC196) and Filipino patients with gastritis (GU173861 and GU173864) (Fig. 2).
Figure 2.
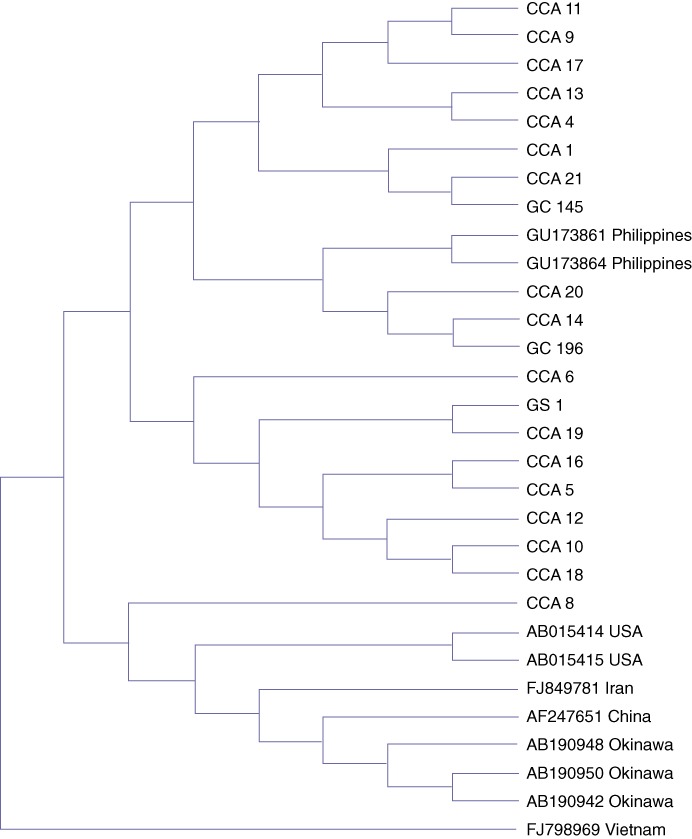
Phylogenetic analysis of partial cagA sequences among ethnic northeast Thai patients with hepatobiliary diseases (cholangiocarcinoma: CCA and cholelithiasis or gallstone: GS) compared with 10 reference strains of Western type cagA in Japanese (AB190942, AB190948, and AB190950), Chinese (AF247651), Iranians (FJ849781), Americans (USA) (AB015414 and AB015415), Vietnamese (FJ798969) and Filipinos (GU173861 and GU173864). The Western CagA in Thai patients with gastric cancer (GC196 and GC145) was included in this analysis. GU = gastric ulcer, GC = gastric cancer
Discussion
Helicobacter pylori was detected in patients with CCA significantly more frequently than in those with cholelithiasis or the control group and, in our previous study, the presence of H. pylori is associated with biliary cell inflammation and proliferation.9 This microorganism has been observed to induce a pathological effect on hepatobiliary cells in animal models.35 These results indicate that there is a likely correlation between H. pylori and hepatobiliary diseases, especially in liver carcinoma. The genetic variations of H. pylori have been previously described and several virulence genes that play a role in the pathogenesis of gastrointestinal tract.2 As there has been no report on genotyping of the H. pylori virulence-associated genes in hepatobiliary diseases, we investigated whether there was any such association including patients with malignant CCA and benign cholelithiasis.
The cagA gene has been reported to be the most important gene related to the severity of gastroduodenal diseases.17 It is present in more than 90% of H. pylori strains isolated from gastroduodenal diseases in Thailand36 compared with 60–70% in Western countries.25 Some researchers have shown that the cagA positive strains, compared with cagA negative strains, significantly increase the risk of developing gastric cancer.18,20 Our previous report demonstrated that cagA positive strains are not significantly different among Thai patients with gastroduodenal diseases.36 In order to prove the role of H. pylori in the hepatobiliary tract, patients with gastroduodenal diseases were excluded from the present study.
A previous report showed that a low prevalence of cagA positive strains (43%) was found among Thai asymptomatic healthy persons as compared with Japanese in whom a high prevalence of cagA positive strains was found (85%).37 It is debated whether the cagA positive gene is significantly more prevalent in CCA (36.2%) than in cholelithiasis patients (9.1%) (P < 0.05).9 Chronic cholestasis is an important factor involved in the development of CCA;38 this condition is presumed to create a condition for H. pylori survival.39,40 It is possible that the H. pylori obstructed by cholestasis in CCA might increase the risk of exposure to H. pylori toxigenic strains, especially cagA positive strains. These data indicate that the cagA gene may be associated with hepatobiliary malignancy as cited by Tiwari et al.8 and Huang et al.,7 who found a correlation between the presence of the cagA gene in hepatobiliary diseases and liver cancer. Additionally, other components of cagPAI and other virulence factors for H. pylori should be emphasized.
The cagPAI encoded proteins for the type 4 secretion apparatus to facilitate production of CagA, peptidoglycan and possibly other bacterial factors in host cells, which are the main cause of inflammation.41 A previous study showed that cagA, cagE, cagM and cagT of cagPAI were used as useful markers for the risk of peptic ulcer disease.42 In this study, besides cagA, cagE was also used as a marker to confirm the presence of cagPAI. The cagE gene is one of the components in cagPAI43 and is involved in the construction of the bacterial transport system44 and induces IL-8 production in gastric epithelial cells.43 As with the presence of the cagA gene observed in this study, it was found that the cagE gene in patients with CCA was significantly higher than in those with cholelithiasis (P < 0.05), which is in accord with a previous report on the cagE gene and gastric cancer.45 The combination of the cagA and cagE positive genes was significantly more prevalent among patients with CCA than in those with cholelithiasis (P < 0.05). The present results show that the presence of the cagE gene is commonly found with the cagA gene, which is a representation of cagPAI. It is, therefore, concluded that the cagPAI may be involved in the progression of hepatobiliary cancer (especially CCA).
Dectecting cagA alone appears to be less important as a virulence marker than determining the type and EPIYA motif.9,28 In this study, the genotypes of the cagA gene were determined using DNA sequencing. This study found that most of the cagA+ samples were Western-type CagA (EPIYA-ABC). The East Asian type cagA (EPIYA-ABD) and the repeated EPIYA motifs were not found in patients with either CCA or cholelithiasis. Our previous study showed that the Western type cagA was detected more commonly than the East Asian type among Northeast Thai dyspeptic patients and was found significantly more frequently in patients with a peptic ulcer but was not significant in gastric cancer.46 In the present study, the cagA gene was found in malignant (CCA) significantly more often than in benign patients (cholelithiasis) but the cagA genotype (Western type) for both was not significantly different between groups. Other factors besides the EPIYA motifs may, therefore, be correlated with the pathogenesis of CCA.
It is well known that the prevalence of gastric cancer in Japan is very high.47 In order to determine whether the cagA of the H. pylori strains found in hepatobiliary and gastroduodenal patients in Thailand and other countries were similar, a phylogenetic analysis of the partial DNA sequences of Western type cagA was determined. The results showed that most of the Western cagA sequences among hepatobiliary patients in Thailand formed a different cluster from that of the Western cagA sequences among Japanese and patients from other countries. In addition, we found that the Western cagA sequences among Thai hepatobiliary patients were classified in the same cluster as the Western cagA sequences of Thai patients with gastric cancer and Filipinos with gastritis. It is therefore possible that the H. pylori detected in the Thais with hepatobiliary and gastroduodenal diseases underwent a similar evolution or that the H. pylori found in the hepatobiliary patients might have first come from gastroduodenal sites. The cluster difference between Thailand and Japan observed in this study is similar to a previous report showing that all Western CagA-positive Filipino strains in gastroduodenal patients were classified in the same Western cluster, but not in the Japan–Western cluster.48 These findings indicate that the variation of bacterial strains might be involved in the development of diseases.
Other virulence genes have been reported to be associated with gastroduodenal diseases.49 VacA affected multiple structural and functional alterations in an epithelial cell line, such as vacuole formation50 and induction of apoptosis. There has, however, been no report on vacA genotypes in hepatobiliary disease. According to our results, the vacA s1a+c/m1 genotypes were frequently detected in patients with either CCA or cholelithiasis. Our findings are similar to previous studies on gastroduodenal diseases, in which the vacA s1/m1 genotypes were found more commonly than the vacA s1/m2 genotype and the vacA s1b genotype was not found at all.36,51,52 This accords with a previous report that found that the vacA s1a+c was the most commonly detected genotype among Northern Thai patients but that it was not associated with clinical outcomes.52 Our results similarly showed no association between the vacA genotypes and clinical outcomes for hepatobiliary diseases.
Helicobacter pylori iceA1 positive strains significantly increased IL-8 production in human gastric epithelial cells (SGC7901) compared with iceA1 mutant strains.53Helicobacter pylori possessing the iceA1 gene are significantly associated with the severity of inflammation and peptic ulcer disease, whereas the iceA2 gene is significantly associated with chronic gastritis and normal cases.54 The present study found that the iceA1 gene was the predominant genotype in hepatobiliary diseases, but that there was no difference among patient groups, as with previous reports on gastric diseases.36,55
The babA2 gene encodes the outer membrane protein that binds to the Leb antigen in gastric cells and these H. pylori babA2 strains are associated with gastric cancer.16 We found a high prevalence of the babA2 gene among hepatobiliary patients but we did not find any significant differences in the frequency of the babA2 genotype between CCA and cholelithiasis patients, as reported after an investigation on the association of this gene and clinical outcomes of gastrointestinal diseases.36 A discrepancy, however, may occur depending on: (i) the other outer membrane protein of H. pylori, which may be involved in colonization of SabA adhesin that binds to the Lex antigen;56 and, (ii) there may be a phase variation of the other outer membrane proteins which may participate in this process.57
We found a high prevalence of vacA s1 (s1a+s1c), vacA m1, iceA1 and babA2 among Thai patients with hepatobiliary diseases, albeit there was no significant difference between the various genes and clinical outcomes, as has been noted with gastroduodenal diseases.36 Whereas the cagA and cagE genes were found to be associated with patients with CCA (P < 0.05), over cholelithiasis, the cagA and cagE genes were found in over 90% of those with H. pylori infection.36 Based on our previous study,9 and other studies in vitro58 and in vivo,49cagA and cagE may play an important role in the development of CCA. However, the role of cagPAI on H. pylori survival in the hepatobiliary tract should be studied further.
Additionally, the answer on why H. pylori could colonize in hepatobiliary system, could might be explained by: (i) bile duct obstruction in cholangitis is generally as a result of gallstone or other factors leading to bacteria ascending from the duodenum into the hepatobiliary system59 or (ii) the bile duct obstruction caused by the effect of CCA38 which might increase a pressure within the biliary system leading to bacterial influx into hepatobiliary tract. However, H. pylori as a main causative factor or co-factor in CCA could not conclude. The explanation of H. pylori may be the main causative factor because we found significantly more H. pylori associated with the inflammation and proliferation surrounding tumor tissues in CCA patients9 whereas the explanation of H. pylori may be the co-factor because the chronic cholestasis had occurred resulting the obstruction of bile duct and then the bacteria was ascended and increased the severity of disease. Therefore, the role of H. pylori as the main causative factor or co-factor should be further investigated.
Besides the variation of H. pylori genotypes, we found that other host factors may be involved in the development of CCA because of the association among age and gender with CCA.
In conclusion, the present research provides new evidence of the prevalence of H. pylori virulence-associated genes in hepatobiliary patients, especially CCA. There was no significant association of the vacA, iceA and babA2 genotypes with the clinical status of hepatobiliary diseases, whereas cagA and cagE genotypes was associated with carcinogenesis of the CCA and might serve as potential markers for prediction of the severity of hepatobiliary diseases. The western-type cagA was the predominant genotype of H. pylori found in hepatobiliary patients but did not associate with the severity of disease. The pathogenesis of H. pylori and its virulence-associated genes in the extra-gastrointestinal tract should, however, be further clarified, particularly in an animal model.
Acknowledgments
The authors would like to thank the Commission on Higher Education, Thailand, for grant support from the Strategic Scholarships for Frontier Research for Joint Thai Ph.D. Programs, the Khon Kaen University for financial support, specimens provided by the Liver Fluke and Cholangiocarcinoma Research Center, the staff of the operating unit for their kind help with specimen collection and Mr Bryan Roderick Hamman and Mrs Janice Loewen-Hamman for assistance with the English-language presentation of the manuscript.
Conflict of interest
The authors declare that they have no competing interests.
References
- 1.Dunn BE, Cohen H, Blaser MJ. Helicobacter pylori. Clin Microbiol Rev. 1997;10:720–741. doi: 10.1128/cmr.10.4.720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Hernandez-Hernandez Ldel C, Lazcano-Ponce EC, Lopez-Vidal Y, Aguilar-Gutierrez GR. Relevance of Helicobacter pylori virulence factors for vaccine development. Salud Publica Mex. 2009;51:S447–S454. doi: 10.1590/s0036-36342009000900010. [DOI] [PubMed] [Google Scholar]
- 3.van Doorn LJ, Figueiredo C, Sanna R, Plaisier A, Schneeberger P, de Boer W, et al. Clinical relevance of the cagA, vacA, and iceA status of Helicobacter pylori. Gastroenterology. 1998;115:58–66. doi: 10.1016/s0016-5085(98)70365-8. [DOI] [PubMed] [Google Scholar]
- 4.Perng CL, Lin HJ, Sun IC, Tseng GY, Facg Helicobacter pylori cagA, iceA and vacA status in Taiwanese patients with peptic ulcer and gastritis. J Gastroenterol Hepatol. 2003;18:1244–1249. doi: 10.1046/j.1440-1746.2003.03214.x. [DOI] [PubMed] [Google Scholar]
- 5.Tan HJ, Rizal AM, Rosmadi MY, Goh KL. Role of Helicobacter pylori virulence factor and genotypes in non-ulcer dyspepsia. J Gastroenterol Hepatol. 2006;21:110–115. doi: 10.1111/j.1440-1746.2005.04063.x. [DOI] [PubMed] [Google Scholar]
- 6.Sripa B, Pairojkul C. Cholangiocarcinoma: lessons from Thailand. Curr Opin Gastroenterol. 2008;24:349–356. doi: 10.1097/MOG.0b013e3282fbf9b3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Huang Y, Fan XG, Wang ZM, Zhou JH, Tian XF, Li N. Identification of helicobacter species in human liver samples from patients with primary hepatocellular carcinoma. J Clin Pathol. 2004;57:1273–1277. doi: 10.1136/jcp.2004.018556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Tiwari SK, Khan AA, Ibrahim M, Habeeb MA, Habibullah CM. Helicobacter pylori and other Helicobacter species DNA in human bile samples from patients with various hepato-biliary diseases. World J Gastroenterol. 2006;12:2181–2186. doi: 10.3748/wjg.v12.i14.2181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Boonyanugomol W, Chomvarin C, Sripa B, Bhudhisawasdi V, Khuntikeo N, Hahnvajanawong C, et al. Helicobacter pylori in Thai patients with cholangiocarcinoma and its association with biliary inflammation and proliferation. HPB (Oxford) 2012;14:177–184. doi: 10.1111/j.1477-2574.2011.00423.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Cover TL, Tummuru MK, Cao P, Thompson SA, Blaser MJ. Divergence of genetic sequences for the vacuolating cytotoxin among Helicobacter pylori strains. J Biol Chem. 1994;269:10566–10573. [PubMed] [Google Scholar]
- 11.Fischer W, Gebert B, Haas R. Novel activities of the Helicobacter pylori vacuolating cytotoxin: from epithelial cells towards the immune system. Int J Med Microbiol. 2004;293:539–547. doi: 10.1078/1438-4221-00300. [DOI] [PubMed] [Google Scholar]
- 12.Atherton JC, Cao P, Peek RM, Jr, Tummuru MK, Blaser MJ, Cover TL. Mosaicism in vacuolating cytotoxin alleles of Helicobacter pylori. Association of specific vacA types with cytotoxin production and peptic ulceration. J Biol Chem. 1995;270:17771–17777. doi: 10.1074/jbc.270.30.17771. [DOI] [PubMed] [Google Scholar]
- 13.Peek RM, Jr, Thompson SA, Donahue JP, Tham KT, Atherton JC, Blaser MJ, et al. Adherence to gastric epithelial cells induces expression of a Helicobacter pylori gene, iceA, that is associated with clinical outcome. Proc Assoc Am Physicians. 1998;110:531–544. [PubMed] [Google Scholar]
- 14.Figueiredo C, Quint WG, Sanna R, Sablon E, Donahue JP, Xu Q, et al. Genetic organization and heterogeneity of the iceA locus of Helicobacter pylori. Gene. 2000;246:59–68. doi: 10.1016/s0378-1119(00)00054-8. [DOI] [PubMed] [Google Scholar]
- 15.Ilver D, Arnqvist A, Ogren J, Frick IM, Kersulyte D, Incecik ET, et al. Helicobacter pylori adhesin binding fucosylated histo-blood group antigens revealed by retagging. Science. 1998;279:373–377. doi: 10.1126/science.279.5349.373. [DOI] [PubMed] [Google Scholar]
- 16.Gerhard M, Lehn N, Neumayer N, Boren T, Rad R, Schepp W, et al. Clinical relevance of the Helicobacter pylori gene for blood-group antigen-binding adhesin. Proc Natl Acad Sci U S A. 1999;96:12778–12783. doi: 10.1073/pnas.96.22.12778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hatakeyama M. The role of Helicobacter pylori CagA in gastric carcinogenesis. Int J Hematol. 2006;84:301–308. doi: 10.1532/IJH97.06166. [DOI] [PubMed] [Google Scholar]
- 18.Blaser MJ, Perez-Perez GI, Kleanthous H, Cover TL, Peek RM, Chyou PH, et al. Infection with Helicobacter pylori strains possessing cagA is associated with an increased risk of developing adenocarcinoma of the stomach. Cancer Res. 1995;55:2111–2115. [PubMed] [Google Scholar]
- 19.Kuipers EJ, Perez-Perez GI, Meuwissen SG, Blaser MJ. Helicobacter pylori and atrophic gastritis: importance of the cagA status. J Natl Cancer Inst. 1995;87:1777–1780. doi: 10.1093/jnci/87.23.1777. [DOI] [PubMed] [Google Scholar]
- 20.Parsonnet J, Friedman GD, Orentreich N, Vogelman H. ) Risk for gastric cancer in people with CagA positive or CagA negative Helicobacter pylori infection. Gut. 1997;40:297–301. doi: 10.1136/gut.40.3.297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Asahi M, Azuma T, Ito S, Ito Y, Suto H, Nagai Y, et al. Helicobacter pylori CagA protein can be tyrosine phosphorylated in gastric epithelial cells. J Exp Med. 2000;191:593–602. doi: 10.1084/jem.191.4.593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Azuma T, Yamazaki S, Yamakawa A, Ohtani M, Muramatsu A, Suto H, et al. Association between diversity in the Src homology 2 domain–containing tyrosine phosphatase binding site of Helicobacter pylori CagA protein and gastric atrophy and cancer. J Infect Dis. 2004;189:820–827. doi: 10.1086/381782. [DOI] [PubMed] [Google Scholar]
- 23.Fu HY, Asahi K, Hayashi Y, Eguchi H, Murata H, Tsujii M, et al. East Asian-type Helicobacter pylori cytotoxin-associated gene A protein has a more significant effect on growth of rat gastric mucosal cells than the Western type. J Gastroenterol Hepatol. 2007;22:355–362. doi: 10.1111/j.1440-1746.2006.04531.x. [DOI] [PubMed] [Google Scholar]
- 24.Odenbreit S, Puls J, Sedlmaier B, Gerland E, Fischer W, Haas R. Translocation of Helicobacter pylori CagA into gastric epithelial cells by type IV secretion. Science. 2000;287:1497–1500. doi: 10.1126/science.287.5457.1497. [DOI] [PubMed] [Google Scholar]
- 25.Allen LA. Intracellular niches for extracellular bacteria: lessons from Helicobacter pylori. J Leukoc Biol. 1999;66:753–756. doi: 10.1002/jlb.66.5.753. [DOI] [PubMed] [Google Scholar]
- 26.Oliveira AG, das Gracas Pimenta Sanna M, Rocha GA, Rocha AM, Santos A, Dani R, et al. Helicobacter species in the intestinal mucosa of patients with ulcerative colitis. J Clin Microbiol. 2004;42:384–386. doi: 10.1128/JCM.42.1.384-386.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ito Y, Azuma T, Ito S, Miyaji H, Hirai M, Yamazaki Y, et al. Analysis and typing of the vacA gene from cagA-positive strains of Helicobacter pylori isolated in Japan. J Clin Microbiol. 1997;35:1710–1714. doi: 10.1128/jcm.35.7.1710-1714.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Argent RH, Zhang Y, Atherton JC. Simple method for determination of the number of Helicobacter pylori CagA variable-region EPIYA tyrosine phosphorylation motifs by PCR. J Clin Microbiol. 2005;43:791–795. doi: 10.1128/JCM.43.2.791-795.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Schmidt HM, Goh KL, Fock KM, Hilmi I, Dhamodaran S, Forman D, et al. Distinct cagA EPIYA motifs are associated with ethnic diversity in Malaysia and Singapore. Helicobacter. 2009;14:256–263. doi: 10.1111/j.1523-5378.2009.00684.x. [DOI] [PubMed] [Google Scholar]
- 30.Qiao W, Hu JL, Xiao B, Wu KC, Peng DR, Atherton JC, et al. cagA and vacA genotype of Helicobacter pylori associated with gastric diseases in Xi'an area. World J Gastroenterol. 2003;9:1762–1766. doi: 10.3748/wjg.v9.i8.1762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Yamazaki S, Yamakawa A, Okuda T, Ohtani M, Suto H, Ito Y, et al. Distinct diversity of vacA, cagA, and cagE genes of Helicobacter pylori associated with peptic ulcer in Japan. J Clin Microbiol. 2005;43:3906–3916. doi: 10.1128/JCM.43.8.3906-3916.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Sillakivi T, Aro H, Ustav M, Peetsalu M, Peetsalu A, Mikelsaar M. Diversity of Helicobacter pylori genotypes among Estonian and Russian patients with perforated peptic ulcer, living in Southern Estonia. FEMS Microbiol Lett. 2001;195:29–33. doi: 10.1111/j.1574-6968.2001.tb10493.x. [DOI] [PubMed] [Google Scholar]
- 33.Sheu BS, Sheu SM, Yang HB, Huang AH, Wu JJ. ) Host gastric Lewis expression determines the bacterial density of Helicobacter pylori in babA2 genopositive infection. Gut. 2003;52:927–932. doi: 10.1136/gut.52.7.927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Tomasini ML, Zanussi S, Sozzi M, Tedeschi R, Basaglia G, De Paoli P. Heterogeneity of cag genotypes in Helicobacter pylori isolates from human biopsy specimens. J Clin Microbiol. 2003;41:976–980. doi: 10.1128/JCM.41.3.976-980.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Goo MJ, Ki MR, Lee HR, Yang HJ, Yuan DW, Hong IH, et al. Helicobacter pylori promotes hepatic fibrosis in the animal model. Lab Invest. 2009;89:1291–1303. doi: 10.1038/labinvest.2009.90. [DOI] [PubMed] [Google Scholar]
- 36.Chomvarin C, Namwat W, Chaicumpar K, Mairiang P, Sangchan A, Sripa B, et al. Prevalence of Helicobacter pylori vacA, cagA, cagE, iceA and babA2 genotypes in Thai dyspeptic patients. Int J Infect Dis. 2008;12:30–36. doi: 10.1016/j.ijid.2007.03.012. [DOI] [PubMed] [Google Scholar]
- 37.Hirai I, Sasaki T, Kimoto A, Yamamoto Y, Azuma T, Mahachai V, et al. Infection of less virulent Helicobacter pylori strains in asymptomatic healthy individuals in Thailand as a potential contributing factor to the Asian enigma. Microbes Infect. 2010;12:227–230. doi: 10.1016/j.micinf.2009.12.007. [DOI] [PubMed] [Google Scholar]
- 38.Yang H, Li TW, Peng J, Tang X, Ko KS, Xia M, et al. A mouse model of cholestasis-associated cholangiocarcinoma and transcription factors involved in progression. Gastroenterology. 2011;141:378–388. doi: 10.1053/j.gastro.2011.03.044. 388 e371-374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Magnuson TH, Lillemoe KD, Zarkin BA, Pitt HA. Patients with uncomplicated cholelithiasis acidify bile normally. Dig Dis Sci. 1992;37:1517–1522. doi: 10.1007/BF01296496. [DOI] [PubMed] [Google Scholar]
- 40.Tompkins DS, West AP. Campylobacter pylori, acid, and bile. J Clin Pathol. 1987;40:1387. doi: 10.1136/jcp.40.11.1387-a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kusters JG, van Vliet AH, Kuipers EJ. Pathogenesis of Helicobacter pylori infection. Clin Microbiol Rev. 2006;19:449–490. doi: 10.1128/CMR.00054-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Mattar R, Marques SB, Monteiro Mdo S, Dos Santos AF, Iriya K, Carrilho FJ. Helicobacter pylori cag pathogenicity island genes: clinical relevance for peptic ulcer disease development in Brazil. J Med Microbiol. 2007;56:9–14. doi: 10.1099/jmm.0.46824-0. [DOI] [PubMed] [Google Scholar]
- 43.Tummuru MK, Sharma SA, Blaser MJ. Helicobacter pylori picB, a homologue of the Bordetella pertussis toxin secretion protein, is required for induction of IL-8 in gastric epithelial cells. Mol Microbiol. 1995;18:867–876. doi: 10.1111/j.1365-2958.1995.18050867.x. [DOI] [PubMed] [Google Scholar]
- 44.Censini S, Lange C, Xiang Z, Crabtree JE, Ghiara P, Borodovsky M, et al. cag, a pathogenicity island of Helicobacter pylori, encodes type I-specific and disease-associated virulence factors. Proc Natl Acad Sci U S A. 1996;93:14648–14653. doi: 10.1073/pnas.93.25.14648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Lima VP, de Lima MA, Ferreira MV, Barros MA, Rabenhorst SH. The relationship between Helicobacter pylori genes cagE and virB11 and gastric cancer. Int J Infect Dis. 2010;14:e613–e617. doi: 10.1016/j.ijid.2009.09.006. [DOI] [PubMed] [Google Scholar]
- 46.Chomvarin C, Sawadpanich K, Mairiang P, Namwat W, Wongkham C, Hahnvajanawong C. Prevalence of cagA EPIYA motifs in Helicobacter pylori among dyspeptic patients in Northeast Thailand. Southeast Asian J Trop Med Public Health. 2012;43:105–115. [PubMed] [Google Scholar]
- 47.Fock KM, Ang TL. Epidemiology of Helicobacter pylori infection and gastric cancer in Asia. J Gastroenterol Hepatol. 2010;25:479–486. doi: 10.1111/j.1440-1746.2009.06188.x. [DOI] [PubMed] [Google Scholar]
- 48.Cortes MC, Yamakawa A, Casingal CR, Fajardo LS, Juan ML, De Guzman BB, et al. Diversity of the cagA gene of Helicobacter pylori strains from patients with gastroduodenal diseases in the Philippines. FEMS Immunol Med Microbiol. 2010;60:90–97. doi: 10.1111/j.1574-695X.2010.00722.x. [DOI] [PubMed] [Google Scholar]
- 49.Huang Y, Tian XF, Fan XG, Fu CY, Zhu C. The pathological effect of Helicobacter pylori infection on liver tissues in mice. Clin Microbiol Infect. 2009;15:843–849. doi: 10.1111/j.1469-0691.2009.02719.x. [DOI] [PubMed] [Google Scholar]
- 50.Leunk RD, Johnson PT, David BC, Kraft WG, Morgan DR. Cytotoxic activity in broth-culture filtrates of Campylobacter pylori. J Med Microbiol. 1988;26:93–99. doi: 10.1099/00222615-26-2-93. [DOI] [PubMed] [Google Scholar]
- 51.Vilaichone RK, Mahachai V, Tumwasorn S, Wu JY, Graham DY, Yamaoka Y. Molecular epidemiology and outcome of Helicobacter pylori infection in Thailand: a cultural cross roads. Helicobacter. 2004;9:453–459. doi: 10.1111/j.1083-4389.2004.00260.x. [DOI] [PubMed] [Google Scholar]
- 52.Linpisarn S, Suwan W, Lertprasertsuk N, Koosirirat C, Steger HF, Prommuangyong K, et al. Helicobacter pylori cagA, vacA and iceA genotypes in northern Thai patients with gastric disease. Southeast Asian J Trop Med Public Health. 2007;38:356–362. [PubMed] [Google Scholar]
- 53.Ma YJ, Duan GC, Zhang RG, Fan QT, Zhang WD. Mutation of iceA in Helicobacter pylori compromised IL-8 induction from human gastric epithelial cells. J Basic Microbiol. 2010;50:S83–S88. doi: 10.1002/jobm.200900410. [DOI] [PubMed] [Google Scholar]
- 54.Caner V, Yilmaz M, Yonetci N, Zencir S, Karagenc N, Kaleli I, et al. H. pylori iceA alleles are disease-specific virulence factors. World J Gastroenterol. 2007;13:2581–2585. doi: 10.3748/wjg.v13.i18.2581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Miciuleviciene J, Calkauskas H, Jonaitis L, Kiudelis G, Tamosiunas V, Praskevicius A, et al. Helicobacter pylori genotypes in Lithuanian patients with chronic gastritis and duodenal ulcer. Medicina (Kaunas, Lithuania) 2008;44:449–454. [PubMed] [Google Scholar]
- 56.Mahdavi J, Sonden B, Hurtig M, Olfat FO, Forsberg L, Roche N, et al. Helicobacter pylori SabA adhesin in persistent infection and chronic inflammation. Science. 2002;297:573–578. doi: 10.1126/science.1069076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Goodwin AC, Weinberger DM, Ford CB, Nelson JC, Snider JD, Hall JD, et al. Expression of the Helicobacter pylori adhesin SabA is controlled via phase variation and the ArsRS signal transduction system. Microbiology. 2008;154:2231–2240. doi: 10.1099/mic.0.2007/016055-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Boonyanugomol W, Chomvarin C, Baik SC, Song JY, Hahnvajanawong C, Kim KM, et al. Role of cagA-positive Helicobacter pylori on cell proliferation, apoptosis, and inflammation in biliary cells. Dig Dis Sci. 2011;56:1682–1692. doi: 10.1007/s10620-010-1512-y. [DOI] [PubMed] [Google Scholar]
- 59.Kinney TP. Management of ascending cholangitis. Gastrointest Endosc Clin N Am. 2007;17:289–306. doi: 10.1016/j.giec.2007.03.006. vi. [DOI] [PubMed] [Google Scholar]