

Abstract
Using a repetitive-sequence-based (rep)-PCR (DiversiLab), we have molecularly typed Acinetobacter nosocomial bloodstream isolates (Acinetobacter baumannii [n = 187], Acinetobacter pittii [n = 23], and Acinetobacter nosocomialis [n = 61]) obtained from patients hospitalized in U.S. hospitals over a 10-year period (1995-2004) during a nationwide surveillance study (Surveillance and Control of Pathogens of Epidemiological Importance [SCOPE]). Patterns of A. baumannii rep-PCR were compared to those of previously identified international clonal lineages (ICs) and were further investigated by multilocus sequence typing (MLST) to compare the two typing methods. Forty-seven of the A. baumannii isolates clustered with the previously defined IC 2. ICs 1, 3, 6, and 7 were also detected. The remaining 81 isolates were unrelated to the described ICs. In contrast, A. pittii and A. nosocomialis isolates were more heterogeneous, as determined by rep-PCR. Our MLST results were in good correlation with the rep-PCR clusters. Our study confirms previous data indicating the predominance of a few major clonal A. baumannii lineages in the United States, particularly IC 2. The presence in the United States of A. baumannii ICs 1, 2, and 3 from as early as 1995 suggests that global dissemination of these lineages was an early event.
INTRODUCTION
The Acinetobacter baumannii group comprises three species, Acinetobacter baumannii, Acinetobacter pittii (formerly Acinetobacter genomic species 3), and Acinetobacter nosocomialis (formerly Acinetobacter genomic species [gen. sp.] strain 13TU) (20, 21). These three species are found primarily as nosocomial pathogens, but it is A. baumannii that is of most concern and is associated with multidrug resistance and outbreaks (11, 21).
The impact of hospital-acquired infections is costly both in human and monetary terms (6). For those tasked with tracking these infections, there now exist a plethora of techniques with which to investigate the sources, spread, and eradication of the infectious agents, with pulsed-field gel electrophoresis (PFGE) still considered the gold standard for typing outbreak-related isolates of A. baumannii (25). A less discriminatory method is multilocus sequence typing (MLST), but this method is primarily useful for population genetics studies (2, 9) while the semiautomated repetitive-sequence-based (rep)-PCR-based DiversiLab system has been shown to be almost as discriminatory as PFGE (24). rep-PCR has the advantage of being less labor-intensive than PFGE and MLST while allowing for the investigation of large numbers of isolates (13, 14, 17).
Portability of typing results is important, and despite attempts to standardize PFGE it is still difficult to compare results generated at different sites (25). The reproducibility of rep-PCR fingerprints between laboratories was recently tested, and clustering was found to be conserved (15). However, the most portable method is MLST, as it relies on DNA sequences. The MLST scheme as proposed by Bartual et al. has been used to identify several clonal complexes (CCs) that correlate with the previously identified European clones (1, 2, 19). An alternative MLST scheme gave similar results (9).
The few studies carried out to investigate the epidemiology of A. pittii and A. nosocomialis have shown these populations to be more heterogeneous than those of A. baumannii, although the typing methods used may have had an impact on the interpretation of the typing results (26, 28, 30). For example, MLST of A. nosocomialis isolated from different hospitals revealed that there was no relatedness between isolates from different outbreaks, i.e., outbreaks were caused by local strains that had not spread to other locations, whereas randomly amplified polymorphic DNA (RAPD) performed by Spence et al. revealed a cluster of A. nosocomialis isolates obtained from patients from 15 different hospitals (28, 34). These results contrast with findings for A. baumannii, for which Dijkshoorn and coworkers identified three clonal lineages that were later termed European clones I, II, and III (10, 32). More recently, with the use of the DiversiLab system, eight worldwide or international carbapenem-resistant clonal lineages (ICs 1 through 8) were identified, three of which corresponded to European clones I through III (9, 14).
Comparisons between rep-PCR and MLST have been performed with several bacterial species, such as Campylobacter spp., Staphylococcus aureus, and Salmonella enterica. These comparisons have shown these methods to be in fairly good agreement, although it was noted that the DNA extraction method affected the rep-PCR results (3, 4, 29). A recent study of 30 A. baumannii isolates from a hospital in west China showed good correlation between the DiversiLab system and MLST (12).
In the present study, we used the Diversilab system to investigate the molecular epidemiology of A. baumannii, A. pittii, and A. nosocomialis bloodstream isolates that were collected in the United States between 1995 and 2004. The presences of the acquired blaOXA carbapenem resistance determinants and the ISAba1 insertion sequences adjacent to blaOXA-51 were also investigated. Additionally, with the A. baumannii isolates, we compared the clustering generated by rep-PCR with that generated by MLST.
(Part of this work was presented at the 8th International Symposium on the Biology of Acinetobacter, Rome, Italy, 1 to 3 September 2010.)
MATERIALS AND METHODS
Bacterial isolates.
Acinetobacter isolates (n = 271) were prospectively collected between 1995 and 2004 from patients with nosocomial bloodstream infections from 52 hospitals in the United States participating in the Surveillance and Control of Pathogens of Epidemiologic Importance (SCOPE) project (33). Identification to the species level (A. baumannii [n = 187], A. pittii [n = 23], and A. nosocomialis [n = 61]) was performed by amplified rRNA gene restriction analysis (ARDRA) and by gyrB multiplex PCR as described previously (35).
Imipenem susceptibility.
Imipenem MICs were investigated by Etest (bioMérieux, Nürtingen, Germany). Isolates with imipenem MICs of ≤4 mg/liter and ≥16 mg/liter were interpreted as susceptible and resistant, respectively (5). MICs of other antimicrobials have been reported previously (35).
Detection of carbapenemases.
Isolates with an imipenem MIC of ≥2 mg/liter were investigated for the presence of OXA-type carbapenemases as described previously (16, 36). This threshold was chosen because the highest imipenem MIC of the wild-type A. baumannii population is 1 mg/liter (see http://www.eucast.org/mic_distributions/). The presence of the insertion element ISAba1 upstream of blaOXA-51-like was investigated by PCR (31).
Molecular typing.
Epidemiological typing of all isolates was performed by rep-PCR (DiversiLab System, bioMérieux, Nürtingen, Germany) as described previously (15). Results were analyzed with the DiversiLab software using the modified Kullback-Leibler statistical method. Isolates that clustered at >95% similarity were considered related and defined rep-PCR clusters (14, 17). Representative isolates belonging to previously identified A. baumannii worldwide clonal clusters 1 through 8 were included as controls (14). A. baumannii isolates (n = 140) were further investigated by MLST. The remaining 47 A. baumannii isolates were part of a large clonal outbreak at a single hospital, as shown by rep-PCR, and therefore were deemed unnecessary to be included in the MLST analysis. MLST was performed as described previously by Bartual et al. (2) with minor modifications. The following modified primers were used for amplification and sequencing: 5′-ACCCGTGAAGGTGAAATCAG-3′ (rpoD_F), 5′-TTCAGCTGGAGCTTTAGCAAT-3′ (rpoD_R), 5′-TGAAGGCGGCTTATCTGAGT-3′ (gyrB_F), 5′-GCTGGGTCTTTTTCCTGACA-3′ (gyrB_R), 5′-GAAATTTCCGGAGCTCACAA-3′ (gpi_F), and 5′-TCAGGAGCAATACCCCACTC-3′ (gpi_R). MLST sequences were uploaded to the A. baumannii MLST sequence type database (http://pubmlst.org/abaumannii/) to determine the alleles and sequence types (ST). Clonal complexes (CCs) were assigned using eBURST V3 (http://eburst.mlst.net/v3/) and were defined as single-locus variants (SLVs) and double-locus variants (DLVs).
RESULTS AND DISCUSSION
In this study we investigated the imipenem susceptibility and molecular epidemiology of Acinetobacter bloodstream isolates from the United States. The majority of Acinetobacter isolates were imipenem susceptible. Forty-nine A. baumannii isolates had imipenem MICs of ≥2 mg/liter, of which 18 (10%) were carbapenem intermediate or carbapenem resistant. More recently, imipenem nonsusceptibility rates of 34 to 75% were reported and confirm the increasing carbapenem resistance seen in this species (1, 8, 12).
The 49 isolates were investigated by PCR for the presence of blaOXA carbapenem resistance determinants. No acquired blaOXA resistance determinants were detected; however, 13 carbapenem-nonsusceptible and 19 carbapenem-susceptible A. baumannii strains had ISAbaI upstream of the blaOXA-51-like gene. Some of these strains were isolated as early as 1996. What was later termed ISAbaI was first described in 2003 and was associated with overexpression of the intrinsic blaampC gene (7). Subsequent to this, ISAbaI was described as adjacent to blaOXA-58 and blaOXA-51-like and is often associated with carbapenem resistance (22, 31). It is interesting that over half of the isolates in our study that had ISAba1 associated with blaOXA-51 remained carbapenem susceptible but had imipenem MICs above the wild-type MIC distribution, suggesting that ISAbaI adjacent to blaOXA-51 is not in itself a good predictor of carbapenem resistance. Therefore, the acquisition of ISAba1 by A. baumannii has occurred earlier than previously described and may have represented the first step in the evolution of resistance against the carbapenems. Several recent studies investigating carbapenem-resistant A. baumannii from the United States have shown ISAba1 to be associated with blaOXA-51 in 46 to 65% of isolates (1, 8).
All 187 A. baumannii isolates were typed by rep-PCR. The comparison between rep-PCR and MLST typing of A. baumannii is summarized in Table 1. Using ≥95% similarity to define a cluster, we used our in-house library to detect 24 clusters, the largest of which contained 94 isolates that clustered with IC 2 (14). Evidence of clonal spread came from the IC 2 isolates that originated from 11 centers, and the IC 1 isolates, although less numerous, were recovered from 4 different centers. Overall, half the isolates were either considered singletons (i.e., not related to another ST) or were small clusters containing 2 to 6 identical isolates from the same center, suggesting small local outbreaks. These singletons and small clusters A through S did not cluster with isolates from ICs 1 through 8 (Table 1).
Table 1.
Correlation between DiversiLab and MLST clustering and year of isolationa
| No. of isolates | DiversiLab cluster | MLSTb |
Allele |
Year of isolation | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ST | CC | gltA | gyrB | gdhB | recA | cpn60 | gpi | rpoD | |||
| 4 | IC 1 | 109 | 109 | 10 | 12 | 4 | 11 | 4 | 9 | 5 | 1996, 1998, 2002 |
| 1 | IC 1 | 95 | 109 | 10 | 12 | 4 | 11 | 1 | 9 | 5 | 1998 |
| 9 | IC 2 | 98 | 92 | 1 | 12 | 3 | 2 | 2 | 3 | 3 | 1995, 1997–99, 2001–2002 |
| 29 | IC 2 | 223 | 92 | 1 | 17 | 3 | 2 | 2 | 56 | 3 | 1995–2004 |
| 1 | IC 2 | 270 | 92 | 1 | 12 | 3 | 2 | 46 | 3 | 3 | 2001 |
| 3 | IC 2 | 281 | 92 | 1 | 17 | 3 | 2 | 2 | 99 | 3 | 1996–1997, 2001 |
| 3 | IC 2 | 282 | 92 | 1 | 17 | 3 | 2 | 2 | 116 | 3 | 1996–1997 |
| 1 | IC 2 | 290 | 92 | 1 | 38 | 3 | 2 | 2 | 56 | 3 | 1997 |
| 1 | IC 2 | 298 | 92 | 1 | 72 | 3 | 2 | 2 | 56 | 3 | 1997 |
| 2 | IC 3 | 266 | 187 | 1 | 1 | 12 | 32 | 1 | 9 | 6 | 1997–1998 |
| 3 | IC 6 | 196 | n.a. | 2 | 21 | 12 | 32 | 26 | 63 | 5 | 1995–1996, 1998 |
| 1 | IC 7 | 276 | 276 | 1 | 15 | 2 | 28 | 1 | 104 | 32 | 1995 |
| 1 | IC 7 | 110 | 276 | 1 | 15 | 2 | 28 | 1 | 52 | 32 | 1995 |
| 1 | A | 267 | n.a. | 1 | 1 | 66 | 12 | 33 | 117 | 41 | 2002 |
| 1 | A | 268 | n.a. | 1 | 1 | 66 | 12 | 33 | 129 | 41 | 2003 |
| 2 | B | 273 | 272 | 1 | 12 | 65 | 45 | 4 | 122 | 26 | 1996, 2002 |
| 1 | B | 272 | 272 | 1 | 12 | 65 | 45 | 4 | 4 | 26 | 1997 |
| 2 | B | 285 | 272 | 1 | 17 | 65 | 45 | 4 | 55 | 26 | 2002 |
| 4 | C | 277 | s | 1 | 15 | 3 | 31 | 4 | 1 | 4 | 2000–2101 |
| 2 | D | 280 | s | 1 | 17 | 2 | 12 | 1 | 138 | 69 | 1996 |
| 2 | E | 283 | n.a. | 1 | 17 | 6 | 1 | 4 | 56 | 6 | 1995–1996 |
| 1 | E | 284 | n.a. | 1 | 17 | 6 | 1 | 47 | 56 | 6 | 1995 |
| 3 | E | 299 | n.a. | 1 | 74 | 6 | 1 | 4 | 125 | 6 | 2002–2003 |
| 2 | F | 291 | n.a. | 1 | 46 | 96 | 54 | 4 | 18 | 59 | 1997, 2001 |
| 3 | G | 292 | s | 1 | 46 | 100 | 12 | 36 | 71 | 6 | 2001 |
| 2 | H | 294 | s | 1 | 52 | 94 | 11 | 1 | 117 | 26 | 1997, 2001 |
| 3 | I | 302 | n.a. | 1 | 81 | 11 | 48 | 18 | 24 | 43 | 1999 |
| 1 | J | 316 | s | 15 | 17 | 2 | 12 | 4 | 64 | 6 | 1997 |
| 2 | J | 317 | s | 15 | 72 | 2 | 12 | 4 | 64 | 6 | 1997 |
| 2 | K | 305 | s | 2 | 1 | 12 | 1 | 4 | 67 | 6 | 2002 |
| 2 | L | 323 | s | 24 | 58 | 107 | 2 | 4 | 54 | 6 | 2001 |
| 1 | M | 332 | n.a. | 28 | 38 | 45 | 1 | 16 | 40 | 2 | 1996 |
| 1 | M | 330 | n.a. | 28 | 17 | 45 | 1 | 16 | 4 | 2 | 1997 |
| 2 | M | 331 | n.a. | 28 | 38 | 45 | 1 | 16 | 4 | 2 | 1996, 2002 |
| 2 | N | 335 | n.a. | 33 | 17 | 12 | 49 | 1 | 54 | 4 | 2002–2003 |
| 4 | O | 339 | s | 44 | 73 | 4 | 11 | 44 | 121 | 4 | 1999, 2001 |
| 2 | P | 340 | n.a. | 45 | 77 | 59 | 2 | 4 | 56 | 64 | 2000 |
| 2 | Q | 341 | s | 46 | 12 | 110 | 1 | 16 | 133 | 50 | 2002–2003 |
| 2 | R | 307 | s | 3 | 77 | 42 | 11 | 16 | 9 | 5 | 2000, 2003 |
| 1 | S | 279 | n.a. | 1 | 15 | 62 | 31 | 4 | 72 | 45 | 1996 |
| 1 | S | 295 | n.a. | 1 | 54 | 62 | 31 | 4 | 50 | 45 | 1996 |
| 1 | Unc | 271 | 271 | 1 | 12 | 40 | 26 | 22 | 117 | 5 | 2000 |
| 1 | Unc | 288 | 271 | 1 | 34 | 40 | 26 | 22 | 1 | 5 | 1996 |
| 1 | Unc | 293 | s | 1 | 52 | 67 | 46 | 1 | 18 | 30 | 2001 |
| 1 | Unc | 318 | s | 18 | 12 | 80 | 12 | 1 | 123 | 3 | 2002 |
| 1 | Unc | 20 | 20 | 1 | 15 | 13 | 12 | 4 | 12 | 2 | 1997 |
| 1 | Unc | 269 | s | 1 | 1 | 116 | 60 | 1 | 132 | 50 | 1995 |
| 1 | Unc | 274 | s | 1 | 12 | 103 | 6 | 29 | 126 | 32 | 2002 |
| 1 | Unc | 17 | 275 | 1 | 12 | 12 | 11 | 4 | 10 | 3 | 1996 |
| 1 | Unc | 278 | 119 | 1 | 15 | 12 | 6 | 28 | 59 | 63 | 1999 |
| 1 | Unc | 287 | s | 1 | 17 | 117 | 11 | 28 | 11 | 32 | 1995 |
| 1 | Unc | 289 | s | 1 | 35 | 64 | 37 | 4 | 132 | 30 | 2001 |
| 1 | Unc | 81 | s | 1 | 54 | 59 | 11 | 4 | 69 | 45 | 2003 |
| 1 | Unc | 297 | s | 1 | 64 | 109 | 1 | 23 | 12 | 26 | 1999 |
| 1 | Unc | 300 | s | 1 | 76 | 106 | 46 | 1 | 131 | 5 | 2000 |
| 1 | Unc | 301 | s | 1 | 78 | 59 | 43 | 1 | 71 | 6 | 2001 |
| 1 | Unc | 303 | s | 1 | 82 | 112 | 1 | 1 | 135 | 66 | 1996 |
| 1 | Unc | 304 | s | 1 | 83 | 114 | 1 | 1 | 136 | 67 | 1995 |
| 1 | Unc | 306 | s | 2 | 79 | 73 | 59 | 1 | 9 | 6 | 2001 |
| 1 | Unc | 319 | s | 21 | 49 | 104 | 11 | 1 | 128 | 30 | 2003 |
| 1 | Unc | 320 | s | 21 | 84 | 64 | 11 | 4 | 9 | 5 | 1995 |
| 1 | Unc | 321 | s | 21 | 85 | 12 | 12 | 26 | 56 | 68 | 1996 |
| 1 | Unc | 322 | n.a. | 23 | 35 | 3 | 27 | 23 | 55 | 7 | 2000 |
| 1 | Unc | 333 | n.a. | 29 | 41 | 48 | 11 | 1 | 137 | 6 | 1995 |
| 1 | Unc | 334 | s | 33 | 12 | 59 | 11 | 32 | 11 | 5 | 2000 |
| 1 | Unc | 336 | s | 35 | 63 | 113 | 1 | 1 | 71 | 6 | 1995 |
| 1 | Unc | 337 | s | 37 | 53 | 67 | 6 | 42 | 64 | 7 | 1997 |
| 1 | Unc | 342 | s | 48 | 48 | 58 | 42 | 36 | 54 | 41 | 1996 |
Only those isolates that were typed using both methods are shown.
ST, sequence type; CC, clonal complex; s, singleton; n.a., clonal complex not assigned, but these STs are DLVs of other published STs.
A possible explanation for the successful spread of IC 2 isolates in the hospital may be that they have a selective advantage over other unrelated strains. Antimicrobial resistance is one such advantage, and we found that 34 of 37 A. baumannii isolates with an imipenem MIC of ≥4 mg/liter were IC 2 strains originating from 7 centers (4 mg/liter [17 isolates]; 8 mg/liter [9 isolates]; 16 mg/liter [5 isolates]; 32 mg/liter [2 isolates]; and >32 mg/liter [2 isolates]). Similarly, a previously reported analysis of antimicrobial susceptibility (35) revealed that CC92 isolates were more resistant to levofloxacin, doxycycline, and ampicillin-sulbactam (96%, 21%, and 77% of isolates, respectively) than all other STs combined (23%, 1%, and 4% of isolates, respectively) (data not shown).
These data indicate that IC 2 isolates had spread to many U.S. centers as early as 1996. Similarly, we found IC 1 isolates from the United States as early as 1995. It is worth noting that although IC I (EUI) was described first in 1996, the earliest isolate dates from 1977, and therefore it is not inconceivable that it had spread to the United States before 1995 (10, 18). We did not detect IC 5 isolates, which was surprising because in a previous study IC 5 was the second-most-prevalent cluster and was almost entirely composed of isolates from North and South America (14). However, there were differences in strain composition between the former and latter studies; in the former study, which comprised more recent isolates, IC 5 isolates were carbapenem nonsusceptible, while in this study the majority of isolates were carbapenem susceptible.
With MLST we further typed 140 A. baumannii isolates (Table 1), and we detected 68 sequence types (STs). Using BURST software (http://pubmlst.org/analysis/), we found that the STs formed 24 clusters comprising SLVs/DLVs (the same number of clusters as determined by rep-PCR clustering). Clustering by MLST correlated with rep-PCR clusters, with the exception of ST271 and ST288, which are DLVs but show <90% similarity with rep-PCR. Each rep-PCR cluster had a corresponding sequence type(s) that in some instances included SLV/DLVs (Table 1). We analyzed these data with all published sequence types available on the MLST website to identify clonal complexes (Table 1, see CC column). The IC 2 cluster correlated with CC92, which has been shown to correlate with EUII isolates, while CC109 correlated with IC 1 (1, 23). Using rep-PCR, we split the IC 2 cluster into two subgroups, which also corresponded with the sequence types (Fig. 1). Subgroup IC 2a comprised isolates with ST223 or single-locus variants, while IC 2b comprised ST98 and a single ST270 isolate. ST223 and ST98 are double-locus variants (Fig. 2). A clear delineation between rep-PCR clustering and sequence types is shown in Fig. 2.
Fig 1.

rep-PCR analysis. Dendrogram and computer-generated images of rep-PCR-banding patterns, including MLST sequence types (STs) and allelic profiles of A. baumannii IC 2 isolates. Subgroup IC 2a consists of STs 223, 281, 282, 290, and 298. Subgroup IC 2b consists of STs 98 and 270.
Fig 2.

Dendrogram and computer-generated images of rep-PCR-banding patterns, including MLST sequence types (STs) and allelic profiles of the A. baumannii rep-PCR clusters M, B, IC 7, A, E, and IC 1.
As previously demonstrated in other studies, the gpi allele showed the most variation within a CC, while gltA, recA, gdhB, and rpoD were the most conserved (1, 23, 34). gpi encodes glucose-6-phosphate isomerase, but why this allele has so many variants is not understood. In a recent study using comparative genomics, extensive recombination was found in EUI and EUII strains, but this was linked most often with genes encoding cell-surface-associated proteins (27). However, we have analyzed the published A. baumannii genomes and found that the location of gpi is not adjacent to any such gene.
Thirty-one STs were considered singletons, as they did not cluster with any other published STs. Sixteen STs were not assigned clonal clusters but were SLVs/DLVs of published STs of which a “founder” has not been identified (Table 1). For example, rep-PCR cluster E was represented by three STs (283, 284, and 299) (Table 1). Cluster analysis using all published STs from the MLST website revealed that these three STs also cluster with previously assigned ST40 and ST120, but no CC has been assigned.
Typing of A. pittii by rep-PCR revealed that the majority of isolates were singletons. Of the 23 isolates, 14 did not cluster with any other strain. The remaining 9 isolates formed 4 clusters, with isolates showing a similarity of >97% (Fig. 3). Two of these clusters involved strains isolated from separate hospitals (clusters Ap-3 and Ap-4), suggesting horizontal spread.
Fig 3.

rep-PCR dendrogram of Acinetobacter pittii isolates showing clusters Ap-1 to Ap-4.
A. nosocomialis isolates were more clonal; 28 singletons with the remaining 33 isolates formed 13 clusters with ≥95% similarity. While some of these clusters included strains from the same center, we also found evidence of horizontal spread, with clusters containing strains isolated from multiple centers with rep-PCR fingerprints showing >95 to 98.5% similarity (Fig. 4).
Fig 4.
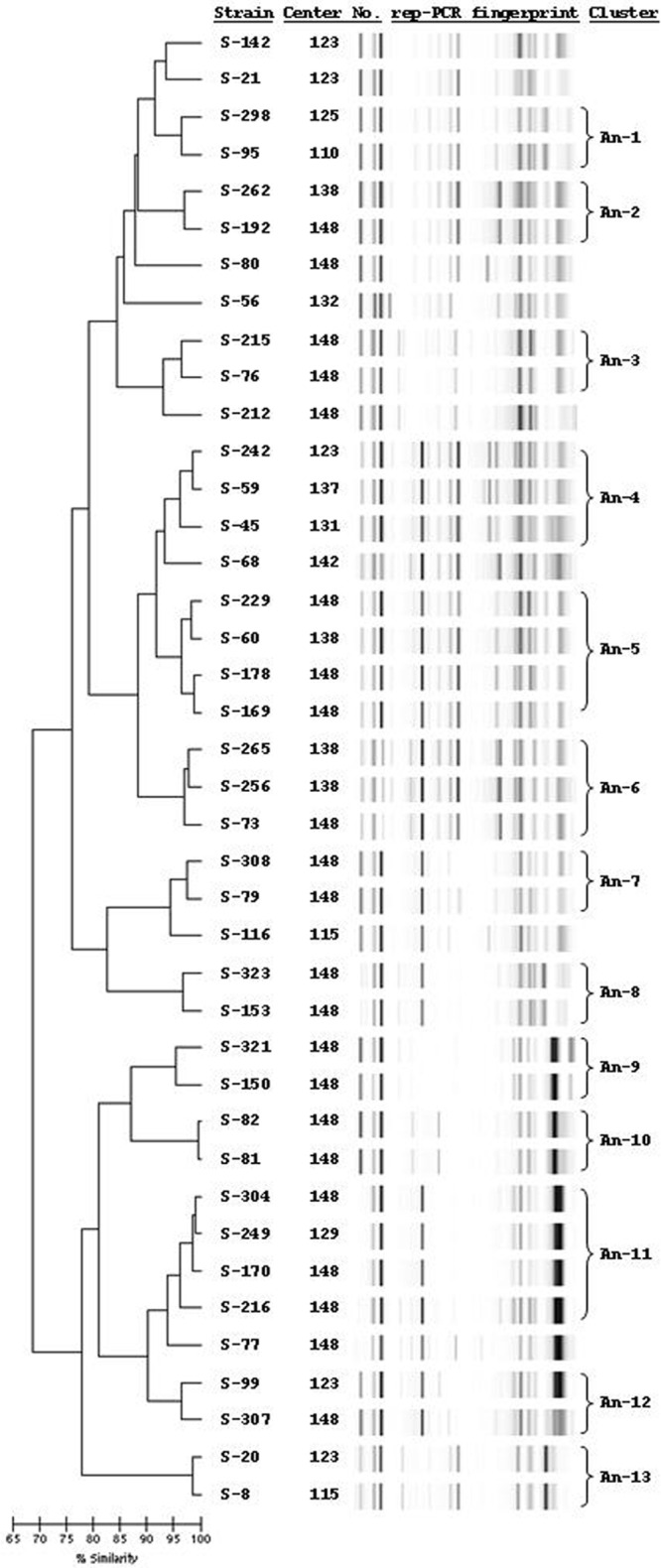
rep-PCR dendrogram of selected Acinetobacter nosocomialis isolates showing singletons and clusters An-1 to An-13.
In summary, these data illustrate several key facts about the epidemiology of the A. baumannii group that causes bloodstream infections. The predominant species is A. baumannii. Global dissemination of the major A. baumannii clones seems to have been an early event, since we found isolates in the United States that clustered with the international (previously European) clones as early as 1995. We also found the ISAbaI element adjacent to blaOXA-51, even when it was not associated with imipenem resistance, in isolates from 1996. MLST correlates very well with DiversiLab rep-PCR typing. A. pittii and A. nosocomialis, in contrast, do not appear to be as widespread and are less clonal in nature than A. baumannii.
ACKNOWLEDGMENTS
The contributions of P.G.H. and H.S. were supported by a grant from the Bundesministerium für Bildung und Forschung (BMBF), Germany, Klinische Forschergruppe Infektiologie (grant 01KI0771).
We thank Danuta Stefanik for technical support and Michael B. Edmond and Richard P. Wenzel for access to the strains used in this study.
Footnotes
Published ahead of print 15 August 2012
REFERENCES
- 1. Adams-Haduch JM, et al. 2011. Molecular epidemiology of carbapenem-nonsusceptible Acinetobacter baumannii in the United States. J. Clin. Microbiol. 49:3849–3854 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Bartual SG, et al. 2005. Development of a multilocus sequence typing scheme for characterization of clinical isolates of Acinetobacter baumannii. J. Clin. Microbiol. 43:4382–4390 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Behringer M, Miller WG, Oyarzabal OA. 2011. Typing of Campylobacter jejuni and Campylobacter coli isolated from live broilers and retail broiler meat by flaA-RFLP, MLST, PFGE and REP-PCR. J. Microbiol. Methods 84:194–201 [DOI] [PubMed] [Google Scholar]
- 4. Ben-Darif E, et al. 2010. Comparison of a semi-automated rep-PCR system and multilocus sequence typing for differentiation of Salmonella enterica isolates. J. Microbiol. Methods 81:11–16 [DOI] [PubMed] [Google Scholar]
- 5. Clinical and Laboratory Standards Institute 2011. Performance standards for antimicrobial susceptibility testing; Twenty-first informational supplement. CLSI Document M100-S21 Clinical and Laboratory Standards Institute, Wayne, PA [Google Scholar]
- 6. Cofsky R, et al. 2002. The cost of antibiotic resistance: effect of resistance among Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, and Pseudomonas aeruginosa on length of hospital stay. Infect. Control Hosp. Epidemiol. 23:106–108 [DOI] [PubMed] [Google Scholar]
- 7. Corvec S, et al. 2003. AmpC cephalosporinase hyperproduction in Acinetobacter baumannii clinical strains. J. Antimicrob. Chemother. 52:629–635 [DOI] [PubMed] [Google Scholar]
- 8. Davies TA, et al. 2011. Longitudinal survey of carbapenem resistance and resistance mechanisms in Enterobacteriaceae and non-fermenters from the U. S. A. in 2007-2009. J. Antimicrob. Chemother. 66:2298–2307 [DOI] [PubMed] [Google Scholar]
- 9. Diancourt L, Passet V, Nemec A, Dijkshoorn L, Brisse S. 2010. The population structure of Acinetobacter baumannii: expanding multiresistant clones from an ancestral susceptible genetic pool. PLoS One 5:e10034 doi:10.1371/journal.pone.0010034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Dijkshoorn L, et al. 1996. Comparison of outbreak and nonoutbreak Acinetobacter baumannii strains by genotypic and phenotypic methods. J. Clin. Microbiol. 34:1519–1525 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Dijkshoorn L, Nemec A, Seifert H. 2007. An increasing threat in hospitals: multidrug-resistant Acinetobacter baumannii. Nat. Rev. Microbiol. 5:939–951 [DOI] [PubMed] [Google Scholar]
- 12. He C, et al. 2011. Increasing imipenem resistance and dissemination of the ISAba1-associated bla(OXA-23) gene among Acinetobacter baumannii isolates in an intensive care unit. J. Med. Microbiol. 60:337–341 [DOI] [PubMed] [Google Scholar]
- 13. Healy M, et al. 2005. Microbial DNA typing by automated repetitive-sequence-based PCR. J. Clin. Microbiol. 43:199–207 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Higgins PG, Dammhayn C, Hackel M, Seifert H. 2010. Global spread of carbapenem-resistant Acinetobacter baumannii. J. Antimicrob. Chemother. 65:233–238 [DOI] [PubMed] [Google Scholar]
- 15. Higgins PG, Hujer AM, Hujer KM, Bonomo RA, Seifert H. 2012. Interlaboratory reproducibility of DiversiLab rep-PCR typing and clustering of Acinetobacter baumannii isolates. J. Med. Microbiol. 61:137–141 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Higgins PG, Lehmann M, Seifert H. 2010. Inclusion of OXA-143 primers in a multiplex polymerase chain reaction (PCR) for genes encoding prevalent OXA carbapenemases in Acinetobacter spp. Int. J. Antimicrob. Agents 35:305. [DOI] [PubMed] [Google Scholar]
- 17. Kohlenberg A, et al. 2009. Outbreak of carbapenem-resistant Acinetobacter baumannii carrying the carbapenemase OXA-23 in a German university medical centre. J. Med. Microbiol. 58:1499–1507 [DOI] [PubMed] [Google Scholar]
- 18. Krizova L, Nemec A. 2010. A 63 kb genomic resistance island found in a multidrug-resistant Acinetobacter baumannii isolate of European clone I from 1977. J. Antimicrob. Chemother. 65:1915–1918 [DOI] [PubMed] [Google Scholar]
- 19. Lee Y, et al. 2011. Carbapenem-non-susceptible Acinetobacter baumannii of sequence type 92 or its single-locus variants with a G428T substitution in zone 2 of the rpoB gene. J. Antimicrob. Chemother. 66:66–72 [DOI] [PubMed] [Google Scholar]
- 20. Nemec A, et al. 2011. Genotypic and phenotypic characterization of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex with the proposal of Acinetobacter pittii sp. nov. (formerly Acinetobacter genomic species 3) and Acinetobacter nosocomialis sp. nov. (formerly Acinetobacter genomic species 13TU). Res. Microbiol. 162:393–404 [DOI] [PubMed] [Google Scholar]
- 21. Peleg AY, Seifert H, Paterson DL. 2008. Acinetobacter baumannii: emergence of a successful pathogen. Clin. Microbiol. Rev. 21:538–582 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Poirel L, Nordmann P. 2006. Genetic structures at the origin of acquisition and expression of the carbapenem-hydrolyzing oxacillinase gene bla(OXA-58) in Acinetobacter baumannii. Antimicrob. Agents Chemother. 50:1442–1448 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Runnegar N, et al. 2010. Molecular epidemiology of multidrug-resistant Acinetobacter baumannii in a single institution over a 10-year period. J. Clin. Microbiol. 48:4051–4056 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Saeed S, Fakih MG, Riederer K, Shah AR, Khatib R. 2006. Interinstitutional and intrainstitutional transmission of a strain of Acinetobacter baumannii detected by molecular analysis: comparison of pulsed-field gel electrophoresis and repetitive sequence-based polymerase chain reaction. Infect. Control Hosp. Epidemiol. 27:981–983 [DOI] [PubMed] [Google Scholar]
- 25. Seifert H, et al. 2005. Standardization and interlaboratory reproducibility assessment of pulsed-field gel electrophoresis-generated fingerprints of Acinetobacter baumannii. J. Clin. Microbiol. 43:4328–4335 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Seifert H, Schulze A, Baginski R, Pulverer G. 1994. Plasmid DNA-fingerprinting of Acinetobacter species other than Acinetobacter baumannii. J. Clin. Microbiol. 32:82–86 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Snitkin ES, et al. 2011. Genome-wide recombination drives diversification of epidemic strains of Acinetobacter baumannii. Proc. Natl. Acad. Sci. U. S. A. 108:13758–13763 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Spence RP, et al. 2002. Population structure and antibiotic resistance of Acinetobacter DNA group 2 and 13TU isolates from hospitals in the UK. J. Med. Microbiol. 51:1107–1112 [DOI] [PubMed] [Google Scholar]
- 29. Te Witt R, Kanhai V, van Leeuwen WB. 2009. Comparison of the DiversiLab (TM) system, pulsed-field gel electrophoresis and multi-locus sequence typing for the characterization of epidemic reference MRSA strains. J. Microbiol. Methods 77:130–133 [DOI] [PubMed] [Google Scholar]
- 30. Traub WH, Bauer D. 2000. Surveillance of nosocomial cross-infections due to three Acinetobacter genospecies (Acinetobacter baumannii, genospecies 3 and genospecies 13) during a 10-year observation period: serotyping, macrorestriction analysis of genomic DNA and antibiotic susceptibilities. Chemotherapy 46:282–292 [DOI] [PubMed] [Google Scholar]
- 31. Turton JF, et al. 2006. The role of ISAba1 in expression of OXA carbapenemase genes in Acinetobacter baumannii. FEMS Microbiol. Lett. 258:72–77 [DOI] [PubMed] [Google Scholar]
- 32. van Dessel H, et al. 2004. Identification of a new geographically widespread multiresistant Acinetobacter baumannii clone from European hospitals. Res. Microbiol. 155:105–112 [DOI] [PubMed] [Google Scholar]
- 33. Wisplinghoff H, et al. 2004. Nosocomial bloodstream infections in US hospitals: analysis of 24,179 cases from a prospective nationwide surveillance study. Clin. Infect. Dis. 39:309–317 [DOI] [PubMed] [Google Scholar]
- 34. Wisplinghoff H, et al. 2008. Molecular epidemiology of clinical Acinetobacter baumannii and Acinetobacter genomic species 13TU isolates using a multilocus sequencing typing scheme. Clin. Microbiol. Infect. 14:708–715 [DOI] [PubMed] [Google Scholar]
- 35. Wisplinghoff H, et al. 2012. Nosocomial bloodstream infections due to Acinetobacter baumannii, Acinetobacter pittii and Acinetobacter nosocomialis in the United States. J. Infect. 64:282–290 [DOI] [PubMed] [Google Scholar]
- 36. Woodford N, et al. 2006. Multiplex PCR for genes encoding prevalent OXA carbapenemases in Acinetobacter spp. Int. J. Antimicrob. Agents 27:351–353 [DOI] [PubMed] [Google Scholar]