

Abstract
In sexual reproduction, two gamete cells (i.e., egg and sperm) fuse (fertilization) to create a newborn with a genetic identity distinct from those of the parents. In the course of these developmental processes, a variety of signal transduction events occur simultaneously in each of the two gametes, as well as in the fertilized egg/zygote/early embryo. In particular, a growing body of knowledge suggests that the tyrosine kinase Src and/or other protein-tyrosine kinases are important elements that facilitate successful implementation of the aforementioned processes in many animal species. In this paper, we summarize recent findings on the roles of protein-tyrosine phosphorylation in many sperm-related processes (from spermatogenesis to epididymal maturation, capacitation, acrosomal exocytosis, and fertilization).
1. Introduction
Protein-tyrosine kinase (PTK) activity and tyrosine phosphorylation of cellular protein were initially discovered by Hunter and colleagues [1–3]; they analyzed the protein kinase activity associated with the protein complex of polyoma virus middle T antigen and viral Src gene product, a cellular counterpart of which is the cellular Src protein. At that time, phosphorylation events on amino acids other than tyrosine (i.e., serine and threonine residues) were already known as posttranslational modifications of physiological importance. However, the discovery of tyrosine phosphorylation for the first time opened a window to understand the relationship between protein phosphorylation (including serine/threonine phosphorylation) and malignant cell transformation (e.g., development of cancer) [4]. In addition, a growing body of evidence has demonstrated that tyrosine phosphorylation catalyzed by cellular Src and other PTKs expressed in normal cells and tissues regulates a variety of cellular functions such as developmental processes, disorder of normal cell functions, immunological responses, neuronal differentiation and transmission, pathological infection, and senescence. Thus, protein-tyrosine phosphorylation has emerged as a signal transduction mechanism of fundamental importance in all eukaryotic cells and, in some cases, prokaryotic cell behavior [5–7].
In the sexual reproduction system, two different kinds of gamete cell: egg and sperm, interact and fuse with each other to accomplish fertilization that gives rise to a newborn [8]. In this fundamental biological event, both egg and sperm undergo a number of biochemical and cell biological reactions that culminate in successful embryogenesis and early development. Especially in the case of multicellular organisms including humans, egg and sperm are special cells in view of their appearance as a single cell. To become such a specialized type of cell, the ancestor of the gametes, that is, primordial germ cell (PGC), along with sex determination in the host, must undergo meiotic cell division [9]. Moreover, to become fully competent for fertilization, egg and sperm must undergo a series of “differentiation” or “maturation” events [10–12]. During the past several decades, a number of studies have dealt with the cellular and molecular mechanisms of gametogenesis, fertilization, and embryogenesis. Among these are characterizations of protein-tyrosine phosphorylation in these events that involved identification of the responsible PTKs (e.g., Src), their regulators and substrates, and evaluation of their roles for cellular functions [13–19]. In this paper, we will briefly discuss the biology of sperm (gametogenesis, differentiation, maturation, and fertilization), recent achievements in understanding the involvement of PTKs and protein-tyrosine phosphorylation in the biology of sperm, and future directions for this research field (Figure 1).
Figure 1.
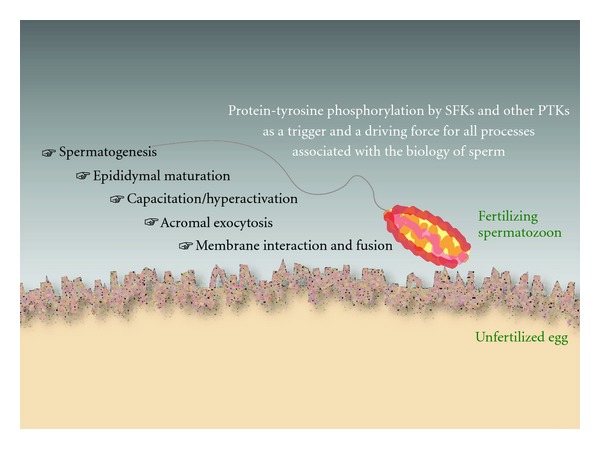
Protein-tyrosine phosphorylation and the biology of sperm. A sequence of events in the sperm must be done to facilitate a successful fertilization. The events include spermatogenesis and epididymal maturation that occur in the male reproductive organs, capacitation/hyperactivation and acrosomal exocytosis (or acrosome reaction, AE) in the female reproductive tract (in the case of species employing internal fertilization: e.g., mammals) or in the extracellular space (in the case of species employing external fertilization: e.g., frogs and fishes), and gamete interaction and fusion at the plasma membranes. In all of these processes, protein-tyrosine phosphorylation catalyzed by SFKs (e.g., Src) and/or other PTKs (e.g., EGFR, Abl) is suggested to play an important role. For details, see text.
2. General View of Sperm Biology
Spermatogenesis is a highly specialized process of cellular differentiation in which diploid progenitor cells of the testis differentiate into haploid spermatozoa [20]. The entire process is divided into three sequential mitotic, meiotic, and postmeiotic stages. In the male meiotic stage, after PGCs migrate into the genital ridges, they become gonocytes and start differentiation into spermatogonia at the basement of seminiferous tubules. Some of them, spermatogonial stem cells (SSCs), also retain the ability for self-renewal [21]. Owing to the role of SSCs, sperm are produced continually (more than 50,000,000 a day in humans) almost throughout the lifetime. Meiosis is the event in which chromosome pairing and genetic recombination occur in the functional tetraploid pachytene spermatocytes [22]. In this process, the genes are shuffled between homologous chromosomes, which results in genetic diversity. This helps the species to survive through natural selection.
Most of the components found in mature spermatozoa are primarily produced at the postmeiotic phase in mammals, and developing spermatids display a variety of morphological and biochemical changes [23]. Many of the organelles in spermatids are transformed into specific structures; the acrosome originates from Golgi body and the main part of the flagellum is composed of spindle-shaped body. The flagellum contains cytoskeletal components and signal transduction mediators. The fibrous sheath, a unique mammalian cytoskeletal structure surrounding the axoneme, serves as a scaffold for constituents of signaling cascades in the regulation of sperm motility [24]. The nucleus is also changed into the tightly compacted shape and size of sperm head by the sequential replacement of the histones with transition proteins and protamines [25].
All stages in spermatogenesis are regulated by the stage-specific expression of a wide variety of genes. Other factors that influence spermatogenesis are the interactions between Sertoli cells and testosterone produced from Leydig cells [26]. In female animals, however, the limited numbers of oogonia differentiate, progress through the first meiotic prophase, and are arrested in the infant ovary. Then, with the onset of adolescence, they mature to second meiotic metaphase and are arrested again. Some of them are released by ovulation and complete meiosis by the entry of a sperm [27]. Similar criteria for oogenesis and oocyte maturation also apply in other kinds of vertebrates, including frog and fish [28, 29]. Testicular sperm look morphologically mature, but they are immotile. Therefore, after sperm leave the testis, they require a further maturation process to acquire the functions for fertilizing an oocyte during transmission through the epididymis [30].
The sperm pass through the caput, corpus, and cauda epididymides sequentially. Then, they are stored at the cauda epididymis until ejaculation. While transiting through the epididymis, they undergo biochemical and physiological modifications, resulting in the acquisition of basal motility and the ability to fertilize an oocyte. These modifications include changes in the glycosylation of acrosomal proteins [31, 32] and in the lipid composition of sperm [33], as well as elevations of cyclic adenosine monophosphate (cAMP) [34, 35] and negative charge on the sperm surface [36]. The other difference between caput and cauda epididymal sperm is the pattern of protein tyrosine phosphorylation [37].
Mammalian sperm need to change their status further to acquire the ability to become competent to bind and fuse with an oocyte after release into the female reproductive tract [38, 39]. This change is termed “capacitation” and confers hyperactivated motility (hyperactivation) and an ability to undergo acrosomal exocytosis (AE) [40] or acrosome reaction to the sperm [8]. To be capacitated, sperm require a period of incubation and interaction in the female reproductive tract. However, this can be induced in vitro in an appropriate experimental medium [40]. Capacitation promotes changes in cholesterol content, plasma membrane fluidity, and intracellular ion concentrations [42]. Another good hallmark for capacitation is an increase in protein tyrosine phosphorylation [43]. Chemotaxis is a phenomenon that guides cells to undergo correct movement toward or away from certain chemicals. This is also known to be important for sperm to interact with an oocyte in the female reproductive tract [44], maybe because sperm are extremely small compared with oocytes. Chemotaxis for sperm guidance was discovered first in marine invertebrate species [45], then in amphibians and mammals [46]. In eutherian mammals, AE releases proteolytic enzymes from the acrosome stored in sperm head [8]. It was believed that these enzymes assist in sperm penetration through the zona pellucida (ZP), the glycoprotein coating on the surface of oocytes, and fusion with an oocyte. However, a recent observation from in vitro fertilization suggests that most sperm undergo AE before contact with the ZP [47], showing the need to reconsider the timing and biological significance of AE during a series of sperm events.
Fertilization-related phenomena include gamete interaction and fusion, egg activation, polyspermy block, and nuclear fusion, all of which culminates in initiation of embryonic development. To date, the sperm membrane protein Izumo1 [48] and the oocyte surface protein CD9 [49–51] are reported to be indispensable for the fusion between sperm and the oocyte plasma membrane in mouse. The gamete fusion triggers repeated increases (e.g., mouse) or a transient elevation (e.g., frog) in intracellular calcium ([Ca2+]i) in oocyte, so-called Ca2+ oscillation or Ca2+ wave, which serves as an initiator of egg activation [17, 52, 53]. It is still debatable how sperm can act as a trigger for egg activation [19]. One possibility is receptor-mediated activator, while another is diffusible activator, “sperm factor.” Recent findings suggest that inositol trisphosphate (IP3) acts as a second messenger for the Ca2+ release reactions and that the egg-associated Src-phospholipase Cγ (PLCγ) (e.g., sea urchin, frog) [14, 16] or the sperm-derived components such as PLCζ (e.g., mouse) [54] and citrate synthase (newt) [55] mediate the gamete interaction/fusion and the activation of IP3-dependent Ca2+ release. After a sperm enters an oocyte, the nucleus has to be decondensed as a pronucleus for nuclear fusion. Then, the fertilized egg starts DNA synthesis for the following early embryogenesis.
Recently, using proteomics approaches, a number of sperm proteins in mouse and rat have been identified as those that are phosphorylated on tyrosine residues during epididymal maturation and capacitation [56–58]. Our group has also reported important roles of Src family PTKs (SFKs) in the sperm-induced egg activation during gamete interaction and fusion by using the African clawed frog, Xenopus laevis. The following sections are an overview of the recent progress to understand the correlations between PTKs and various sperm events.
3. Involvement of PTKs in Spermatogenesis
PTKs play various biological roles in many types of somatic cell, so it is not surprising that they act on spermatogenic cells and their supporting cells, that is, Sertoli cells, in the testis. Actually, it has been demonstrated that several families of PTKs including Src kinase are correlated with most spermatogenic events (Figure 2). In adult mouse testis, the protein expression of Src, Lyn, and Hck were observed, while the expressions of eight members of the SFK were detected by quantitative polymerase chain reaction. For instance, Src protein localizes weakly to the cytoplasm in spermatocytes and strongly in round and elongated spermatids, leading to strong accumulation in acrosome of cauda epididymal sperm with entire flagellum detection [59]. In humans, Src protein was detected strongly around the acrosomal region in round and elongated spermatids [60].
Figure 2.
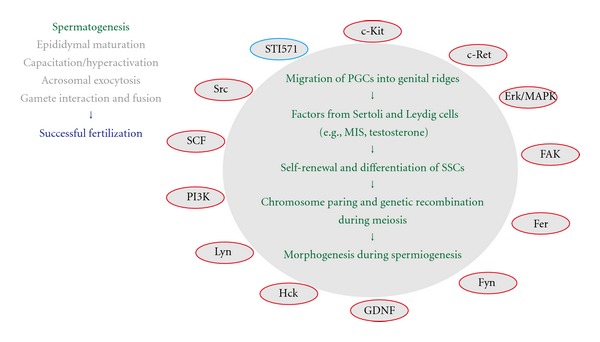
Protein-tyrosine phosphorylation and a sequence of events associated with spermatogenesis. For details of spermatogenesis and its associated signaling molecules (the full spelling of all abbreviations as well), see text. Note that MIS, Mullerian-inhibiting substance, is a testicular-differentiating factor that is produced in Sertoli cells and Leydig cells, whose differentiation is promoted by the actions of Sry and other sex-determining gene products. Also note that MIS acts in concert with testosterone. Note that positive regulators for protein-tyrosine phosphorylation are indicated in red circles
c-Kit is a transmembrane tyrosine kinase receptor that binds to stem cell factor (SCF). SCF induces dimerization of c-Kit that activates the tyrosine kinase residues by autophosphorylation [61], leading to the downstream signaling through phospho-tyrosine-binding adaptor proteins such as PLCγ1. During mouse early development, c-Kit is essential for the migration of PGCs to the genital ridges in the embryo and then functions in the maintenance of PGCs [62]. Src kinase is also involved in this process [63]. In adult mouse testis, the expression of c-Kit is detected in differentiating spermatogonia, whereas the expression of SCF is detected in Sertoli cells; therefore, c-Kit/SCF is important for maintaining differentiating type A spermatogonia [64]. On the other hand, to maintain the property of self-renewal in mouse SSCs, c-Ret tyrosine kinase receptor mediates between glial cell line-derived neurotrophic factor (GDNF) and Src family kinase signaling [65]. Recently, it has also been suggested that c-Kit plays a pivotal role in regulating the ratio between differentiation and self-renewal during maintenance of the SSC population [66].
There are a few reports about tyrosine phosphorylation in meiosis during spermatogenesis (Figure 2). Mouse has Fes-related proteins (Fer) of two different size [67], which correspond to 94 kDa or 51 kDa tyrosine kinase. The latter accumulates in primary spermatocytes where the cell cycle is in the first meiotic prophase, and the role of phosphorylation for the timing of meiosis entry is suggested in mammals as well as in yeast [68]. c-Kit/SCF system is also required for transition when mouse spermatogonia undergo cell division to enter meiosis [69]. This was also examined using a specific inhibitor to c-Kit (STI571), resulting in reduction of the number of mouse meiotic cells under the control of retinoic acid [70].
Interestingly, some of the truncated forms of PTK seem to have roles in the process of sperm morphogenesis: spermiogenesis. At least three examples of nonreceptor tyrosine kinase, Fyn as well as Fer and Hck [71], have been reported. Truncated Fer was detected in the Golgi, acroplaxome, and manchette of rat spermatid [72], while truncated Hck was observed mainly at the acrosome of bovine sperm [73]. They are also suggested to regulate actin assembly via phosphorylation. Therefore, these observations suggest that truncated forms of Fer kinase and Hck may participate in sperm head shaping. Similarly, Golgi membrane in spermatids contains truncated Fyn that is missing the kinase domain, and this protein may be required for acrosome biogenesis [72]. A truncated isoform of c-Kit has also been detected in mouse round spermatids [74]. This protein lacks SCF-binding and dimerization domains, but retains a part of the kinase domain that would facilitate activation of PLCγ1 [75, 76]. It is suggested that truncated c-Kit is related to DNA integrity in human sperm [77]; however, its role is still unclear.
Another correlation of PTKs with spermatogenesis is in the regulation of Sertoli cell tight junction, including at the blood-testis barrier (BTB). Male germ cells need to contact with Sertoli cells during most spermatogenic processes. Spermatogonia differentiate to preleptotene/leptotene spermatocytes in the basal compartment of the seminiferous epithelium. In addition, these spermatocytes have to translocate to the adluminal compartment of the seminiferous epithelium for further differentiation. However, there is the BTB, which acts as the immunological barrier between basal and adluminal compartments. Recently, it has been demonstrated that focal adhesion kinase (FAK), a nonreceptor tyrosine kinase, plays a key role in this process. FAK regulates the opening and/or closing of BTB by modulating the phosphorylation status of integral membrane proteins [78]. Besides, traditionally, FAK has been suggested to be involved in adherens junctions (AJ) between Sertoli and germ cells by the interactions with β1-integrin and other associated proteins including Src [78]. Moreover, Fer kinase has been shown to participate in the regulation of rat AJ [79]. Fyn-functions in the basal ectoplasmic specialization (ES) of actin filaments: at the junction between Sertoli cells as well as apical ES and at the junction between spermatids and Sertoli cells [80]. Apical ES also contains many lipids and protein kinases such as phosphatidylinositol 3-kinase (PI3K) and extracellular signal-regulated kinase/mitogen-activated protein kinase (Erk/MAPK), which are associated with Src [81].
The major problem for research on mammalian spermatogenesis was the lack of a stable in vitro culture system, despite the efforts of many investigators [82, 83]. However, recently, an improved organ culture system using neonatal testis has been established, which can make SSCs differentiate to mature sperm in mouse [84]. This method for in vitro spermatogenesis should greatly facilitate the identification and characterization of more factors and genes correlated with PTKs for self-renewal and differentiation in spermatogonia, meiosis in spermatocytes, and morphogenesis in spermatids.
4. Involvement of PTKs in Epididymal Maturation
Like other cells, sperm need adenosine trisphosphate (ATP) as an energy resource for their functions, for example, motility. The dominant pathway for ATP production in mouse sperm is glycolysis, while spermatocytes and spermatids prefer oxidative phosphorylation [83–85]. It is suggested that this switching to glycolysis occurs during epididymal maturation in rabbit [86]. During epididymal maturation, sperm proteins contain a greater number of disulfide bonds, leading to the stabilization of sperm structures and promotion of tyrosine phosphorylation of sperm proteins (Figure 3) [87–89].
Figure 3.
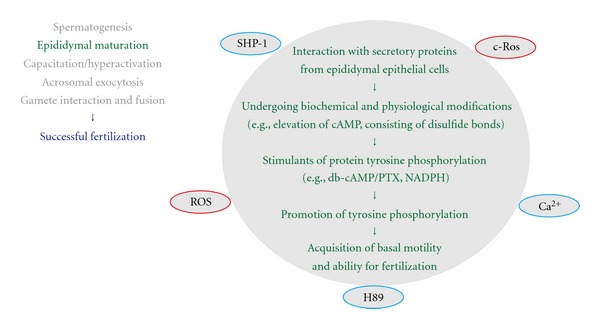
Protein-tyrosine phosphorylation and a sequence of events associated with epididymal maturation of sperm. For details of the epididymal maturation and its associated signaling molecules (the full spelling of all abbreviations as well), see text. Note that positive regulators for protein-tyrosine phosphorylation are indicated in red circles.
To investigate the importance of protein tyrosine phosphorylation during epididymal maturation, most analyses were performed with the antiphosphotyrosine antibody. Using western blotting, the contents of plasma membranes were compared between hamster caput and cauda epididymal sperm, resulting in a differential phosphorylation pattern: the proteins with sizes of 94, 52, and 47 kDa looked more intense in cauda epididymal sperm while the 67 kDa band had more intensity in caput epididymal sperm [90]. However, western blotting detected caput epididymal sperm-specific phosphotyrosine expression in 93, 66, and 45 kDa bands in boar [91]; in addition, rat sperm from caput epididymis tended to show a stronger total band pattern of tyrosine phosphorylation than that of cauda epididymal sperm [37]. Immunofluorescence analyses with the antiphosphotyrosine antibody were performed to visualize the distribution of tyrosine phosphorylation in sperm. After permeabilization with methanol, boar sperm from proximal caput epididymis had strong labeling on the midacrosome as well as a faint signal on the whole tail. After transit through distal caput and corpus epididymides, this signal was detected only as a triangular shape on the posterior region of the midacrosome [91]. In mouse and rat, caput epididymal sperm, permeabilized with Nonidet P-40, resulted in fluorescence over the whole equatorial segment; however, the signal became restricted to a small region in the posterior equatorial segment after spermmoved to the cauda epididymis [92]. It is suggested that the equatorial segment plasma membrane works as a site of fusion with an oocyte membrane during fertilization; therefore, the accumulation of tyrosine phosphorylation may be connected to the later fusion process.
Lewis and Aitken have investigated the tyrosine phosphorylation pattern of sperm proteins after stimulation with cAMP by adding dibutyryl cAMP (db-cAMP) and pentoxifylline (PTX) [37]. By western blotting with the antiphosphotyrosine antibody in rat, the increase of cAMP resulted in more intense tyrosine phosphorylation bands in caput epididymal sperm proteins and much more intensity for cauda epididymal sperm proteins. However, this induction of tyrosine phosphorylation was inhibited by a protein kinase A (PKA)-inhibitor, H89 [37]. Immunofluorescence using the sperm fixed with methanol increased the signal in the tail region after db-cAMP/PTX stimulation [37]. Similar results were observed when the reduced form of NADPH (nicotinamide adenine dinucleotide phosphate) was added instead of db-cAMP, suggesting that this cAMP-dependent tyrosine phosphorylation is regulated by the redox system during epididymal maturation [93]. Furthermore, db-cAMP/PTX stimulation showed drastic change of the phosphotyrosine pattern in mouse sperm permeabilized with Triton X-100 as follows: staining on the acrosome and the principal piece of sperm from the proximal caput epididymis, strong on the midpiece as well as the acrosome and the principal piece of sperm from the distal caput and corpus epididymides, still strong on the midpiece and weak on the principal piece without any signal on the acrosome of sperm from the cauda epididymis [94]. It is also suggested that the signal leading to tyrosine phosphorylation in mouse sperm is negatively regulated by Ca2+ [95]. However, this inhibitory effect did not work when sperm arrived at the cauda epididymis [94]. Even with these observations, the mechanism of activation for this tyrosine phosphorylation has not been elucidated. One explanation of tyrosine phosphorylation in the midpiece is that the generation of reactive oxygen species (ROS) activates tyrosine phosphorylation signaling; however, the role of oxidative phosphorylation in sperm mitochondria is still controversial. At present, the role of tyrosine phosphorylation in the acrosome is unknown.
The progress of proteomic analysis has contributed to the identification of sperm proteins that are important for epididymal maturation, including the protein phosphorylation process. Using two-dimensional fluorescence difference gel electrophoresis, eight rat sperm proteins were identified as candidates that undergo posttranslational modifications during epididymal maturation, and one of them, β-subunit of mitochondrial F1-ATPase, was serine-phosphorylated [96]. Recently, new methods using titanium dioxide have been developed to identify phosphopeptides, suggesting that 77 titanium-dioxide-enriched peptides (corresponding to 53 proteins) showed significant modifications during rat epididymal maturation [97].
Here, if we focus on PTKs in epididymal epithelium, the receptor tyrosine kinase Ros and Src homology-2 (SH2-) domain-containing protein tyrosine phosphatase SHP-1 are expressed there. The mutant mice for Ros or for SHP-1 showed defects in the differentiation of the epididymis [98, 99]. Moreover, the sperm interact with various secretory proteins from epithelial cells of epididymides during epididymal transit, and some of them are proposed to be involved in sperm maturation [100, 101]. Therefore, it will be necessary to study the epididymal luminal environment as well as sperm proteins to obtain a deeper understanding of the role of PTKs in the sperm maturation process.
Note —
During the processes of publishing the preent paper, one paper about Src and epididymal development and sperm functions was published [212]. As highlighted in our paper, Src has been identified as a PTK involved in capacitation-associated tyrosine phosphorylation downstream of PKA pathway. Added to this aspect, in this newly published paper, Visconti and colleagues reported that details about the male reproductive phenotypes of Src (KO) mice and Src localization in epididymis as well as in sperm. Src is not detected in caput epididymal sperm but in the midpiece and the postacrosomal region of cauda epididymal sperm. Src is also detected strongly in clear cells and weakly in principle cells of cauda epididymis and is shown to transfer into cauda epididymal sperm via epididymosomes during epididymal transit. Src KO mice have smaller size of cauda epididymis and reduced sperm motility, leading to unsuccessful in vitro fertilization.
5. Involvement of PTKs in Capacitation
Extratesticular sperm that has completed epididymal maturation must undergo a process called capacitation, a prerequisite for hyperactivated motility and acrosome reaction, in the female reproductive tract. Two researchers discovered this process independently in the 1950s [38, 39]. Later studies have demonstrated that capacitation can be reconstituted in vitro by using cauda (but not caput) epididymal or ejaculated sperm and artificial media supplemented with components that promote changes associated with in vivo capacitation. Capacitation seems to be a phenomenon specific to mammals, and accumulating evidence indicates that it generally involves a burst of protein-tyrosine phosphorylation (Figure 4).
Figure 4.
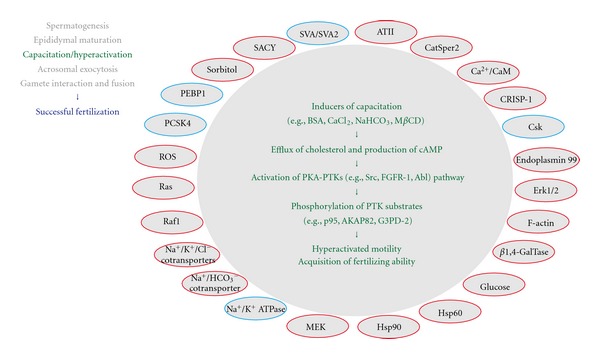
Protein-tyrosine phosphorylation and a sequence of events associated with capacitation and/or hyperactivation of sperm. For details of the capacitation/hyperactivation and its associated signaling molecules (the full spelling of all abbreviations as well), see text. Note that positive regulators for protein-tyrosine phosphorylation are indicated in red circles, whereas the negative regulators are indicated in blue circles.
In mice, treatment of sperm with capacitation-inducing media promotes cAMP-dependent (i.e., PKA-dependent) tyrosine phosphorylation of several sperm proteins with molecular sizes of 116, 105, 95, 86, 76, and 54 kDa [102]. In particular, it is suggested that the 95 kDa phosphotyrosine-containing protein is identical to one that has been identified as a ZP3-dependent PTK substrate, namely, p95/zona receptor kinase (ZRK)/hexokinase (see below). Further studies by Visconti and colleagues have demonstrated that the sperm media should include bovine serum albumin (BSA), CaCl2, and NaHCO3 to induce capacitation and its associated tyrosine phosphorylation (proteins of 40–120 kDa) [103]. Interestingly, caput epididymal sperm, which lack an ability to undergo capacitation in vivo, cannot induce the tyrosine phosphorylation event in response to the treatment with capacitation media, indicating that epididymal maturation is required for the sperm response. In addition, it has also been shown that the requirement for BSA, CaCl2, and NaHCO3 in capacitation and associated PTK signaling is completely overcome by the addition of cAMP or its active analogs and that chemical inhibitors for PKAs (H-89, a substance that blocks ATP binding, and Rp-cAMPS, a nonhydrolysable AMP analog) interfere with the aforementioned processes [104]. These results clearly demonstrate that capacitation involves sequential activation of cAMP production and PKA-PTK pathway in response to the capacitation-inducing substances.
A similar system has also been demonstrated in other species including human [105] and mice of both domestic and wild-field species [106, 107]. Unlike mouse sperm, however, human sperm do not contain the 95 kDa phosphotyrosine-containing protein (p95/ZRK/hexokinase). Instead, the fibrous sheath proteins, AKAP82 (A-kinase/PKA anchoring protein 82: now referred as AKAP4), its precursor pro-AKAP82, and FSP95, a structural homolog of AKAP82, have been identified as prominently tyrosine-phosphorylated proteins in the capacitated sperm [108, 109]. Artificial Ca2+ signals, which promote the occurrence of acrosome reaction, lead to dephosphorylation of a subset of these phosphotyrosine-containing proteins. AKAP82 has also been identified as the major protein of the fibrous sheath of the mouse sperm flagellum, and its possible function to compartmentalize inactive PKA (before capacitation) to the cytoskeleton has been suggested [110]. Immunocytochemical and/or biochemical experiments also demonstrate that the tyrosine-phosphorylated forms of c-Abl tyrosine kinase are present in the capacitated human sperm [111].
It has been shown that the cholesterol-binding heptasaccharides, methyl-β-cyclodextrin (MβCD) and OH-propyl-β-cyclodextrin, primarily promote release of cholesterol from the sperm plasma membrane and induce PTK signaling and capacitation in the absence of BSA [112, 113]. The MβCD's effects, like the BSA's effects, depend on both NaHCO3 and PKA activity, suggesting that they resemble those under physiological capacitation. In fact, BSA has also been shown to promote the release of cholesterol, and the addition of exogenous cholesterol interferes with the BSA-induced PTK signaling and capacitation [112]. These and other results suggest that efflux of cholesterol plays a pivotal role in upregulation of the cAMP-PKA-PTK pathway leading to capacitation [114].
Other important factors that promote or suppress the onset of PTK signaling and/or capacitation include calmodulin, which may act as a positive regulator for the production of cAMP [115, 116]; seminal vesicle autoantigen, which has been shown to block BSA-induced capacitation [117, 118]; fertilization-promoting peptide or adenosine, which stimulates and inhibits PTK signaling in uncapacitated and capacitated sperm, respectively [119, 120]; extracellular glucose, whose shortage has been shown to delay the appearance of protein tyrosine phosphorylation [121]; Na+/HCO3 − cotransporter in the sperm, which provides Na+ ions as a positive regulator of PTK signaling and capacitation [122]; F-actin, whose generation and breakdown is required for capacitation and AE, respectively [123, 124]; endogenous redox activity, which is up-regulated under the control of the actions of HCO3 − [125, 126]; molecular chaperones such as hsp90, endoplasmin 99, and hsp60, which become tyrosine-phosphorylated upon capacitation and some of them may be involved in sperm-ZP recognition (in mouse, but not human) [127–129]; sperm-specific voltage-gated cation channel, CatSper2, whose gene knockout significantly alters sperm production; PTK signaling, which is associated with capacitation and induction of the AE [130]; extracellular Ca2+ ions, which may suppress tyrosine phosphorylation by decreasing the availability of intracellular ATP [131]; β 1,4-galactosyltransferase I, a possible ZP3-interacting protein whose gene knockout leads to precocious capacitation, which may be involved because of spontaneous elevation of cAMP [132]; angiotensin II, which is found in seminal plasma and has been shown to induce PTK signaling and capacitation via stimulation of adenylyl cyclase-dependent accumulation of cAMP [133]; HCO3 −- and Ca2+-responsive soluble adenylyl cyclase (SACY), which has been identified as the dominant source of cAMP production [134, 135]; phosphatidylethanolamine-binding protein 1, a possible decapacitation factor, whose acquisition on the sperm surface during epididymal maturation and release before the onset of capacitation have been identified [136]; Na+/K+ ATPase, whose interaction with ouabain, a specific inhibitor of Na+/K+ ATPase, promotes PTK signaling and capacitation [137]; a 130 kDa CCCTC-binding nuclear factor, which becomes tyrosine-phosphorylated at capacitation and more potently binds to its target DNA sequence [138]; seminal vesicle protein secretion 2, which acts as a decapacitation factor by interacting with ejaculated sperm heads after copulation [139]; the cystic fibrosis transmembrane conductance regulator, a Cl− channel that controls the activity of several transport proteins, including ENaCs (epithelial Na+ channels), and whose pharmacological inhibition leads to the failure of capacitation without affecting PTK signaling [34]; sorbitol, which is present in semen and has been shown to be effective in inducing PTK signaling via the action of sorbitol dehydrogenase [140]; Na+/K+/Cl−cotransporters, which may act as a source of chloride ions necessary for the onset of PKA-dependent PTK signaling at capacitation [141]; glycerol-3-phosphate dehydrogenase 2 (G3PD-2), which is expressed in the acrosome and principal piece and becomes tyrosine-phosphorylated upon capacitation [142, 143]; proprotein convertase subtilisin/kexin type 4 (PCSK4), whose null (thus impaired in fertility) sperm exhibit enhanced tyrosine phosphorylation in response to capacitation [144]; so-called Erk module, including Ras, Raf1, MAPK kinase (MAPKK/MEK), and Erk/MAPK, which is suggested to be involved in the presentation of phosphotyrosine-containing proteins on the sperm surface at capacitation [145].
Recent proteomics analysis has revealed more identities of tyrosine-phosphorylated proteins in response to capacitation: they include voltage-dependent anion channel, tubulin, pyruvate dehydrogenase E1 β chain, glutathione-S-transferase, NADH dehydrogenase (ubiquinone) Fe-S protein 6, acrosin-binding protein precursor (sp32), proteasome subunit αtype 6b, and cytochrome b-c1 complex [42], although their functions remain to be elucidated. A more recent study has shown that Toll-like receptors 2 and 4 present on cumulus cells were activated by coculture with sperm in a hyaluronan fragment-dependent manner and that chemokines secreted from cumulus-oocyte complexes induced sperm PTK signaling and capacitation [146], providing evidence for the association of chemotaxis with capacitation. In addition, study of the knockout mouse suggests that an epididymal secretory protein CRISP-1 contributes to PTK signaling during capacitation [147].
Candidate PTKs related to capacitation include Src, whose interaction with PKA and enzymatic activation are seen in capacitated sperm [148, 149]; C-terminal Src kinase (Csk), whose negative regulatory function toward Src is canceled by serine phosphorylation (maybe by PKA) at capacitation [148, 150]; fibroblast growth factor receptor-1, whose dominant-negative mutant leads to the failure of PTK signaling and capacitation [151]; Abl tyrosine kinase, which is activated in response to capacitation in a PKA-dependent manner [152].
6. Involvement of PTKs in Acrosomal Exocytosis
Early reports by Saling and colleagues have demonstrated that a 95 kDa mouse sperm protein, termed p95/ZRK (for zona receptor kinase)/hexokinase, is a tyrosine kinase substrate, whose phosphorylation level is elevated in response to sperm binding to zona pellucida glycoprotein ZP3 [153, 154]. It has long been believed that the physiological trigger for sperm AE is the binding of sperm to the ZP structures, namely, ZP3; so the aforementioned study has opened a window to analyze the roles of PTKs for AE. Subsequent studies have examined not only the physiological importance of the ZP3-induced tyrosine phosphorylation of the 95 kDa protein [155, 156] and other sperm proteins (e.g., 51 and 14–18 kDa proteins), but also the effect of several kinds of AE inducers other than ZP3 and/or various PTK or protein kinase inhibitors that could affect the tyrosine phosphorylation events associated with AE (Figure 5) [157–160]. A PTK inhibitor, tyrphostin, blocks the ZP-induced activation of PLC [161], suggesting that the γ isoform of PLC is involved in the ZP-induced PTK signaling leading to AE. A more recent study has shown, however, that δ4-isoform of PLC is essential for ZP3- or progesterone (PG)-induced Ca2+ release during mouse AE [162, 163]. AE induced by mannose-bovine serum albumin and an antibody against p95/ZRK, but not that induced by Ca2+ ionophore, can be inhibited by wortmannin, a specific inhibitor of PI3K, without inhibiting PTK signaling, implying a role of PI3K downstream of PTK signaling [164, 165]. AE induced by PG or platelet-activating factor has also been shown to involve an increase in protein-tyrosine phosphorylation of 75 and 97 kDa proteins, and PTK inhibitors (erbstatin, genistein) interfere with the induction of AE [166]. The PG-induced tyrosine phosphorylation is involved in the generation of the plateau phase of Ca2+ influx [167] and modulation of sperm GABAA-like receptor/chloride channel (chloride efflux) [168]. Study of domestic cat sperm has shown that ZP-induced AE, but not Ca2+ ionophore- or spontaneously induced AE, is inhibited by PTK inhibitors (genistein, tyrphostin), indicating that PTK signaling acts upstream of the Ca2+ increase during AE [169]. The involvement of PTK signaling mediated by Src has also been suggested for the promotion of capacitative Ca2+ entry, as reconstituted by thapsigargin treatment of sperm, during AE [170].
Figure 5.
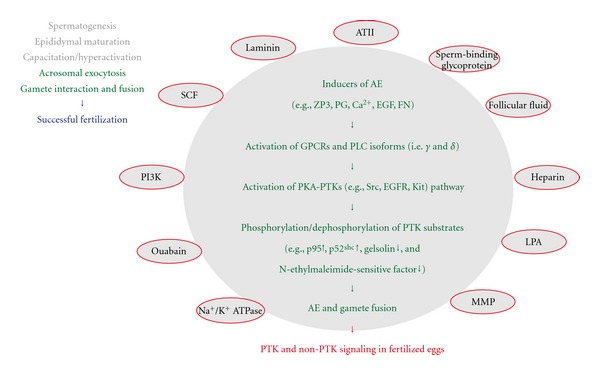
Protein-tyrosine phosphorylation and a sequence of events associated with acrosomal exocytosis. For details of the acrosomal exocytosis and its associated signaling molecules (the full spelling of all abbreviations as well), see text.
Tyrosine-phosphorylated proteins during AE also include p52shc, an isoform of the Shc adaptor proteins [171], a 107 kDa protein, whose phospholevel correlates well with the extent of AE (induced by Ca2+ ionophore) [172], and a heparin-binding sperm membrane protein (during AE induced by heparin) [173]. On the other hand, recent studies suggest the importance of proteintyrosine dephosphorylation in AE [174]. In support of this, tyrosine dephosphorylation of N-ethylmaleimide-sensitive factor, which undergoes SNARE complex disassembly, by protein-tyrosine phosphatase 1B has been shown to be required for the Ca2+ ionophore-induced AE [175], and gelsolin, an actin-severing protein that becomes tyrosinephosphorylated and inactivated during capacitation, has been shown to be dephosphorylated during AE, allowing its activation leading to actin depolymerization [176].
Another line of evidence demonstrates the identity of PTKs working during AE. An early report by Lax et al. showed that epidermal growth factor (EGF) can induce AE in bovine sperm [177], suggesting that EGF receptor (EGFR)/kinase is involved in this process. Further studies using this species have demonstrated that AE primarily involving activation of G protein-coupled receptors by lysophosphatidic acid or angiotensin II or AE induced by ouabain-Na+/K+ ATPase system promotes transactivation of EGFR/kinase via PKA-Src-matrixmetalloproteinase (MMP) or PKA-Src pathway [178, 179]. In the former system, G-protein-mediated production of AMP promotes PKA activation, PKA up-regulates Src (as seen in capacitated sperm), Src activates the secretion of heparin-binding EGF-like growth factor via MMP activation, thereby activating EGFR/kinase, and Src also affects the activity of EGFR/kinase through direct phosphorylation on tyrosine 845 [178], an Src-dependent phosphorylation site, whose phosphorylation has been implicated in some types of cancer cells [180–182]. Src activation and its importance for AE have also been demonstrated in humans [183, 184]. Another line of evidence suggests that SCF is involved in the promotion of mouse sperm AE through the activation of its cognate receptor/PTK c-Kit, PLCγ1, and phosphatidylinositol 3-kinase (PI3K) [185].
While many studies using mammals have shown the importance of PTK signaling in sperm AE, only limited findings have been described on the same subject in nonmammalian species. One potentially interesting finding reported recently demonstrates that egg components are capable of promoting protein-tyrosine phosphorylation and capacitation-like changes in sperm of the amphibian Bufo arenarum [186], implying its subsequent functions in AE.
A recent report by Hirohashi and colleagues has shown that most fertilizing mouse sperm have undergone AE before contact with ZP during in vitro fertilization [46]. Furthermore, it has been shown that sperm binding to the zona pellucida is not sufficient to induce AE and that some mechanical process is important for physiological AE [187]. These results lead us to reconsider where and how sperm AE is initiated under physiological conditions and when and how PTK signaling contributes to the “real” AE. As described above, not only ZP3, but also other reagents or experimental conditions (e.g., PG) are reportedly inducible for AE in vitro. Additionally, possible oviductal substances such as sperm-binding glycoprotein [188], laminin [184], fibronectin [189], and follicular fluid [190] have been shown to induce AE accompanied by PTK signaling. Taking these findings together, further analysis focusing on the roles of the sperm microenvironment during capacitation and AE (before reaching the egg plasma membrane) in vivo should enable greater understanding of the physiological impact of PTK signaling.
7. Involvement of PTKs in Gamete Interaction Fusion
Compared with the aforementioned categories of sperm biology, the relationship between sperm's PTK signaling and gamete interaction, especially at the level of plasma membranes (i.e., adhesion and fusion of gametes), has not yet been fully investigated. Immunocytochemical study demonstrates that sperm tail displays a time-dependent increase in tyrosine phosphorylation in response to ZP-free oocyte-sperm interactions [121], although its physiological importance and molecular detail have not yet been described. This seems to be mainly due to a technical problem in analyzing sperm functions at this point. In physiological conditions, a fertilizing spermatozoon closely interacts with or fuses with the plasma membrane of an egg, which has a protein content several hundredfold or more than that of a single sperm, so that the biochemical and cell biological experiments for evaluating not only protein tyrosine phosphorylation but also other molecular events associated with gamete interaction tend to fall into the analysis of those of the “fertilized egg (mixture of egg and sperm)” or egg itself, but not sperm itself. Under these circumstances, eggs of some animal species have been analyzed for the sperm or sperm-mimetic-induced PTK signaling. Accumulating evidence demonstrates that egg-associated Src and/or some other SFKs (i.e., Fyn and Yes) may play a crucial role for some events at fertilization: they include transient increase(s) in [Ca2+]i concentrations (sea urchin, starfish, ascidian, fish, and frog) [191–194], MII spindle structures and functions (mammals) [195], and cleavage furrow ingression during mitosis (mammals) [196]. Roles played by the SFKs vary among species; however, it is worth noting that a wide range of animal species (from sea invertebrates to mammals) employ egg-associated SFKs as a sperm-induced trigger for activation of development. In this connection, it has been demonstrated that the sperm acrosomal or perinuclear theca-associated proteins may act as a trigger of signal transduction for initiation of development inside fertilized egg: so-called “sperm factors” (see “General View of Sperm Biology”). Among these are truncated c-Kit protein [197–199] and a WW domain-binding protein PAWP [200], both of which are specific proteins that may contribute to the modulation of the PTK signaling in eggs.
Egg analysis often involves parthenogenetic experiments, in which one or more of sperm's function-mimetic substances (e.g., Ca2+ ionophore) are used to reconstitute signaling events of fertilization, allowing easier functional evaluation of the egg-associated proteins. On the other hand, an absence of substitutes for sperm analysis has remained a problem. Ideally, some egg- or egg plasma membranemimetic substances, if applicable, would be helpful for solving this technical problem. In this regard, we suggest that egg membrane microdomains (MDs) could serve as excellent model materials of physiological value. As mentioned earlier, MDs or alternatively lipid/membrane “rafts” have been generally recognized as cholesterol-dependent micron- or nanometer-scaled membrane structures of cells, where a specific subset of glycosphingolipids, membrane-spanning and cytoplasmic proteins, and some other membrane components are assembled [201, 202]. Detailed analysis of egg MDs and fertilization signaling was first reported in sea urchin [203] and frog [204, 205], and thereafter, eggs or early embryos of mouse have also been documented to some extent [206, 207].
In Xenopus laevis, the egg MDs are suggested to serve as a platform for sperm-induced Src PTK signaling. Namely, Src has been shown to be concentrated in the MDs of unfertilized eggs, and it is activated upon fertilization. MβCD treatment impairs the ability of eggs to undergo sperm induced initiation of development [204]. An MD-associated, transmembrane protein, uroplakin III, has been identified as a target of sperm protease, whose activity is required for Xenopus egg fertilization [208, 209], and as an intracellular substrate of Src [210]. In addition, we have found that sperm and some other sperm mimetics are capable of activating Src in MD fractions isolated from unfertilized Xenopus eggs, in vitro [211]. These results demonstrate that egg MDs would be useful materials for reconstitution of sperm-induced PTK signaling in the fertilized egg. If so, an opposite idea might also be valid, that is, egg MDs would be useful for reconstitution of egg (plasma membrane)-induced PTK signaling (or any other signaling event if it occurs) in the fertilizing sperm. To develop these ideas, we are now in the process of evaluating sperm functions before and after interaction with isolated egg MDs. It seems that this kind of reconstitution experiment can also be carried out in other animal species where isolation of egg MDs is possible, and thus its validity and physiological importance will soon be evaluated.
8. Conclusion and Perspectives
Among all the cells constituting multicellular organisms, egg and sperm are unique in terms of their history of production (i.e., gametogenesis, maturation, and/or differentiation), final structures, and physiological functions. In spite of enormous research efforts in recent years, many questions remain about how egg and sperm are produced and how they acquire their gamete-specific functions; in addition, new questions are continuously arising. Recent studies using pluripotent stem cells (e.g., embryonic or induced pluripotent stem cells) and/or molecular genetic approaches (e.g., gene knockout/KO and transgenic animals) have begun to disclose the genetic as well as cell biological background of gametogenesis, fertilization, and subsequent early embryogenesis. Moreover, study on the gametogenesis and fertilization in nonanimal species (e.g., plants, algae), which is not highlighted in this paper, and that in animal species have begun to merge, enabling researchers to learn more about the general scheme of sexual reproduction. Taking this background into account, it is certain that study on the signal transduction system involving protein-tyrosine phosphorylation in egg, sperm, and fertilized egg/zygote/early embryo will continue to be at the cutting edge of this research field.
Acknowledgments
The authors apologize to those whose work was not cited or insufficiently cited. This work is supported by a grant for the collaboration research from the Asahi Kasei Corporation, a Grant-in-Aid on Innovative Areas (22112522, 24112714), and a grant for Private University Strategic Research Foundation Support Program (S0801060) from the Ministry of Education, Culture, Sports, Science and Technology, Japan to K.-i. Sato.
Abbreviations
- PTK:
Protein-tyrosine kinase
- PGC:
Primordial germ cell
- SSC:
Spermatogonial stem cell
- cAMP:
Cyclic adenosine monophosphate
- AE:
Acrosomal exocytosis
- ZP:
Zona pellucida
- [Ca2+]i:
Intracellular calcium
- IP3:
Inositol trisphosphate
- PLC:
Phospholipase C
- SFK:
Src family protein-tyrosine kinase
- SCF:
Stem cell factor
- GDNF:
Glial cell-derived neurotropic factor
- BTB:
Blood-testis barrier
- FAK:
Focal adhesion kinase
- AJ:
Adherence junction
- ES:
Ectoplasmic specialization
- PI3K:
Phosphatidylinositol 3-kinase
- Erk:
Extracellular signal-regulated kinase
- MAPK:
Mitogen-activated protein kinase
- ATP:
Adenosine trisphosphate
- db-cAMP:
Dibutyryl cAMP
- PTX:
Pentoxifylline
- PKA:
Protein kinase A
- NADPH:
Nicotinamide adenine dinucleotide phosphate
- ROS:
Reactive oxygen species
- SH2:
Src homology 2
- SHP:
SH2 domain-containing protein-tyrosine phosphatase
- ZRK:
Zona receptor kinase
- BSA:
Bovine serum albumin
- AKAP:
A-kinase/PKA-anchoring protein
- MβCD:
Methyl-β-cyclodextrin
- SACY:
Soluble adenylyl cyclase
- ENaC:
Epithelial Na+ channel
- G3PD-2:
Glycerol-3-phosphate dehydrogenase 2
- PCSK4:
Pro-protein convertase subtilisin/kexin type 4
- MAPKK:
MAPK kinase
- Csk:
C-terminal Src kinase
- PG:
Progesterone
- EGF:
Epidermal growth factor
- MMP:
Matrix metalloproteinase
- MD:
Membrane microdomain.
References
- 1.Hunter T. Tyrosine phosphorylation: thirty years and counting. Current Opinion in Cell Biology. 2009;21(2):140–146. doi: 10.1016/j.ceb.2009.01.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Hunter T, Sefton BM. Transforming gene product of Rous sarcoma virus phosphorylates tyrosine. Proceedings of the National Academy of Sciences of the United States of America. 1980;77(3 I):1311–1315. doi: 10.1073/pnas.77.3.1311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Eckhart W, Hutchinson MA, Hunter T. An activity phosphorylating tyrosine in polyoma T antigen immunoprecipitates. Cell. 1979;18(4):925–933. doi: 10.1016/0092-8674(79)90205-8. [DOI] [PubMed] [Google Scholar]
- 4.Bishop JM. Molecular themes in oncogenesis. Cell. 1991;64(2):235–248. doi: 10.1016/0092-8674(91)90636-d. [DOI] [PubMed] [Google Scholar]
- 5.Grangeasse C, Cozzone AJ, Deutscher J, Mijakovic I. Tyrosine phosphorylation: an emerging regulatory device of bacterial physiology. Trends in Biochemical Sciences. 2007;32(2):86–94. doi: 10.1016/j.tibs.2006.12.004. [DOI] [PubMed] [Google Scholar]
- 6.Pincus D, Letunic I, Bork P, Lim WA. Evolution of the phospho-tyrosine signaling machinery in premetazoan lineages. Proceedings of the National Academy of Sciences of the United States of America. 2008;105(28):9680–9684. doi: 10.1073/pnas.0803161105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Thomas SM, Brugge JS. Cellular functions regulated by SRC family kinases. Annual Review of Cell and Developmental Biology. 1997;13:513–609. doi: 10.1146/annurev.cellbio.13.1.513. [DOI] [PubMed] [Google Scholar]
- 8.Yanagimachi R. Mammalian fertilization. In: Knobil E, Neil JD, editors. The Physiology of Reproduction. New York, NY, USA: Raven Press; 1994. pp. 189–317. [Google Scholar]
- 9.Wei G, Mahowald AP. The germline: familiar and newly uncovered properties. Annual Review of Genetics. 1994;28:309–324. doi: 10.1146/annurev.ge.28.120194.001521. [DOI] [PubMed] [Google Scholar]
- 10.Darszon A, Nishigaki T, Beltran C, Treviño CL. Calcium channels in the development, maturation, and function of spermatozoa. Physiological Reviews. 2011;91(4):1305–1355. doi: 10.1152/physrev.00028.2010. [DOI] [PubMed] [Google Scholar]
- 11.Toshimori K. Dynamics of the mammalian sperm head: modifications and maturation events from spermatogenesis to egg activation. Advances in Anatomy, Embryology, and Cell Biology. 2009;204:5–94. [PubMed] [Google Scholar]
- 12.Voronina E, Wessel GM. The regulation of oocyte maturation. Current Topics in Developmental Biology. 2003;58:53–110. doi: 10.1016/s0070-2153(03)58003-6. [DOI] [PubMed] [Google Scholar]
- 13.Ciapa B, Chiri S. Egg activation: upstream of the fertilization calcium signal. Biology of the Cell. 2000;92(3-4):215–233. doi: 10.1016/s0248-4900(00)01065-0. [DOI] [PubMed] [Google Scholar]
- 14.Hasan AM, Fukami Y, Sato KI. Gamete membrane microdomains and their associated molecules in fertilization signaling. Molecular Reproduction and Development. 2011;78(10-11):814–830. doi: 10.1002/mrd.21336. [DOI] [PubMed] [Google Scholar]
- 15.Kinsey WH. Tyrosine kinase signaling at fertilization. Biochemical and Biophysical Research Communications. 1997;240(3):519–522. doi: 10.1006/bbrc.1997.7586. [DOI] [PubMed] [Google Scholar]
- 16.Mcginnis LK, Carroll DJ, Kinsey WH. Protein tyrosine kinase signaling during oocyte maturation and fertilization. Molecular Reproduction and Development. 2011;78(10-11):831–845. doi: 10.1002/mrd.21326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Runft LL, Jaffe LA, Mehlmann LM. Egg activation at fertilization: where it all begins. Developmental Biology. 2002;245(2):237–254. doi: 10.1006/dbio.2002.0600. [DOI] [PubMed] [Google Scholar]
- 18.Sato KI, Iwasaki T, Hirahara S, Nishihira Y, Fukami Y. Molecular dissection of egg fertilization signaling with the aid of tyrosine kinase-specific inhibitor and activator strategies. Biochimica et Biophysica Acta. 2004;1697(1-2):103–121. doi: 10.1016/j.bbapap.2003.11.017. [DOI] [PubMed] [Google Scholar]
- 19.Whitaker M. Calcium at fertilization and in early development. Physiological Reviews. 2006;86(1):25–88. doi: 10.1152/physrev.00023.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Handel MA. Genetic control of spermatogenesis in mice. Results and Problems in Cell Differentiation. 1987;15:1–62. doi: 10.1007/978-3-540-47184-4_1. [DOI] [PubMed] [Google Scholar]
- 21.de Rooij DG. Stem cells in the testis. International Journal of Experimental Pathology. 1998;79(2):67–80. doi: 10.1046/j.1365-2613.1998.t01-1-00057.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Roeder GS. Meiotic chromosomes: it takes two to tango. Genes and Development. 1997;11(20):2600–2621. doi: 10.1101/gad.11.20.2600. [DOI] [PubMed] [Google Scholar]
- 23.Toshimori K. Biology of spermatozoa maturation: an overview with an introduction to this issue. Microscopy Research and Technique. 2003;61(1):1–6. doi: 10.1002/jemt.10311. [DOI] [PubMed] [Google Scholar]
- 24.Eddy EM, Toshimori K, O’Brien DA. Fibrous sheath of mammalian spermatozoa. Microscopy Research and Technique. 2003;61(1):103–115. doi: 10.1002/jemt.10320. [DOI] [PubMed] [Google Scholar]
- 25.Steger K. Transcriptional and translational regulation of gene expression in haploid spermatids. Anatomy and Embryology. 1999;199(6):471–487. doi: 10.1007/s004290050245. [DOI] [PubMed] [Google Scholar]
- 26.Eddy EM. Male germ cell gene expression. Recent Progress in Hormone Research. 2002;57:103–128. doi: 10.1210/rp.57.1.103. [DOI] [PubMed] [Google Scholar]
- 27.Senthilkumaran B. Recent advances in meiotic maturation and ovulation: comparing mammals and pisces. Frontiers in Bioscience. 2011;16(5):1898–1914. doi: 10.2741/3829. [DOI] [PubMed] [Google Scholar]
- 28.Yamashita M. Molecular mechanisms of meiotic maturation and arrest in fish and amphibian oocytes. Seminars in Cell and Developmental Biology. 1998;9(5):569–579. doi: 10.1006/scdb.1998.0251. [DOI] [PubMed] [Google Scholar]
- 29.Deng J, Carbajal L, Evaul K, Rasar M, Jamnongjit M, Hammes SR. Nongenomic steroid-triggered oocyte maturation: of mice and frogs. Steroids. 2009;74(7):595–601. doi: 10.1016/j.steroids.2008.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Cornwall GA. New insights into epididymal biology and function. Human Reproduction Update. 2009;15(2):213–227. doi: 10.1093/humupd/dmn055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Deng X, Czymmek K, Martin-DeLeon PA. Biochemical maturation of Spam1 (PH-20) during epididymal transit of mouse sperm involves modifications of N-linked oligosaccharides. Molecular Reproduction and Development. 1999;52(2):196–206. doi: 10.1002/(SICI)1098-2795(199902)52:2<196::AID-MRD11>3.0.CO;2-A. [DOI] [PubMed] [Google Scholar]
- 32.Morin G, Lalancette C, Sullivan R, Leclerc P. Identification of the bull sperm p80 protein as a PH-20 ortholog and its modification during the epididymal transit. Molecular Reproduction and Development. 2005;71(4):523–534. doi: 10.1002/mrd.20308. [DOI] [PubMed] [Google Scholar]
- 33.Nikolopoulou M, Soucek DA, Vary JC. Changes in the lipid content of boar sperm plasma membranes during epididymal maturation. Biochimica et Biophysica Acta. 1985;815(3):486–498. doi: 10.1016/0005-2736(85)90377-3. [DOI] [PubMed] [Google Scholar]
- 34.Hernández-González EO, Treviño CL, Castellano LE, et al. Involvement of cystic fibrosis transmembrane conductance regulator in mouse sperm capacitation. Journal of Biological Chemistry. 2007;282(33):24397–24406. doi: 10.1074/jbc.M701603200. [DOI] [PubMed] [Google Scholar]
- 35.White DR, Aitken RJ. Influence of epididymal maturation on cyclic AMP levels in hamster spermatozoa. International Journal of Andrology. 1989;12(1):29–43. doi: 10.1111/j.1365-2605.1989.tb01283.x. [DOI] [PubMed] [Google Scholar]
- 36.Yanagimachi R, Noda YD, Fujimoto M, Nicolson GL. The distribution of negative surface charges on mammalian spermatozoa. American Journal of Anatomy. 1972;135(4):497–519. doi: 10.1002/aja.1001350405. [DOI] [PubMed] [Google Scholar]
- 37.Lewis B, Aitken RJ. Impact of epididymal maturation on the tyrosine phosphorylation patterns exhibited by rat spermatozoa. Biology of Reproduction. 64(5):1545–1556. doi: 10.1095/biolreprod64.5.1545. [DOI] [PubMed] [Google Scholar]
- 38.Austin CR. Observations on the penetration of the sperm in the mammalian egg. Australian Journal of Scientific Research B. 1951;4(4):581–596. doi: 10.1071/bi9510581. [DOI] [PubMed] [Google Scholar]
- 39.Chang MC. Fertilizing capacity of spermatozoa deposited into the fallopian tubes. Nature. 1951;168(4277):697–698. doi: 10.1038/168697b0. [DOI] [PubMed] [Google Scholar]
- 40.Yanagimachi R, Chang MC. Fertilization of hamster eggs in vitro. Nature. 1963;200(4903):281–282. doi: 10.1038/200281b0. [DOI] [PubMed] [Google Scholar]
- 41.Hernández-González EO, Sosnik J, Edwards J, et al. Sodium and epithelial sodium channels participate in the regulation of the capacitation-associated hyperpolarization in mouse sperm. Journal of Biological Chemistry. 2006;281(9):5623–5633. doi: 10.1074/jbc.M508172200. [DOI] [PubMed] [Google Scholar]
- 42.Arcelay E, Salicioni AM, Wertheimer E, Visconti PE. Identification of proteins undergoing tyrosine phosphorylation during mouse sperm capacitation. International Journal of Developmental Biology. 2008;52(5-6):463–472. doi: 10.1387/ijdb.072555ea. [DOI] [PubMed] [Google Scholar]
- 43.Eisenbach M, Giojalas LC. Sperm guidance in mammals—an unpaved road to the egg. Nature Reviews Molecular Cell Biology. 2006;7(4):276–285. doi: 10.1038/nrm1893. [DOI] [PubMed] [Google Scholar]
- 44.Yoshida M, Kawano N, Yoshida K. Control of sperm motility and fertility: diverse factors and common mechanisms. Cellular and Molecular Life Sciences. 2008;65(21):3446–3457. doi: 10.1007/s00018-008-8230-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Burnett LA, Xiang X, Bieber AL, Chandler DE. Crisp proteins and sperm chemotaxis: discovery in amphibians and explorations in mammals. International Journal of Developmental Biology. 2008;52(5-6):489–501. doi: 10.1387/ijdb.072545lb. [DOI] [PubMed] [Google Scholar]
- 46.Jin M, Fujiwara E, Kakiuchi Y, et al. Most fertilizing mouse spermatozoa begin their acrosome reaction before contact with the zona pellucida during in vitro fertilization. Proceedings of the National Academy of Sciences of the United States of America. 2011;108(12):4892–4896. doi: 10.1073/pnas.1018202108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Inoue N, Ikawa M, Isotani A, Okabe M. The immunoglobulin superfamily protein Izumo is required for sperm to fuse with eggs. Nature. 2005;434(7030):234–238. doi: 10.1038/nature03362. [DOI] [PubMed] [Google Scholar]
- 48.Kaji K, Oda S, Shikano T, et al. The gamete fusion process is defective in eggs of Cd9-deficient mice. Nature Genetics. 2000;24(3):279–282. doi: 10.1038/73502. [DOI] [PubMed] [Google Scholar]
- 49.Boucheix C. Severely reduced female fertility in CD9-deficient mice. Science. 2000;287(5451):319–321. doi: 10.1126/science.287.5451.319. [DOI] [PubMed] [Google Scholar]
- 50.Miyado K, Yamada G, Yamada S, et al. Requirement of CD9 on the egg plasma membrane for fertilization. Science. 2000;287(5451):321–324. doi: 10.1126/science.287.5451.321. [DOI] [PubMed] [Google Scholar]
- 51.Ducibella T, Fissore R. The roles of Ca2+, downstream protein kinases, and oscillatory signaling in regulating fertilization and the activation of development. Developmental Biology. 2008;315(2):257–279. doi: 10.1016/j.ydbio.2007.12.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Stricker SA. Comparative biology of calcium signaling during fertilization and egg activation in animals. Developmental Biology. 1999;211(2):157–176. doi: 10.1006/dbio.1999.9340. [DOI] [PubMed] [Google Scholar]
- 53.Nomikos M, Swann K, Lai FA. Starting a new life: sperm PLC-zeta mobilizes the Ca 2+ signal that induces egg activation and embryo development: an essential phospholipase C with implications for male infertility. BioEssays. 2012;34(2):126–134. doi: 10.1002/bies.201100127. [DOI] [PubMed] [Google Scholar]
- 54.Harada Y, Matsumoto T, Hirahara S, et al. Characterization of a sperm factor for egg activation at fertilization of the newt Cynops pyrrhogaster . Developmental Biology. 2007;306(2):797–808. doi: 10.1016/j.ydbio.2007.04.019. [DOI] [PubMed] [Google Scholar]
- 55.Baker MA, Smith ND, Hetherington L, et al. Label-free quantitation of phosphopeptide changes during rat sperm capacitation. Journal of Proteome Research. 2010;9(2):718–729. doi: 10.1021/pr900513d. [DOI] [PubMed] [Google Scholar]
- 56.Baker MA, Reeves G, Hetherington L, Aitken RJ. Analysis of proteomic changes associated with sperm capacitation through the combined use of IPG-strip prefractionation followed by RP chromatography LC-MS/ MS analysis. Proteomics. 2010;10(3):482–495. doi: 10.1002/pmic.200900574. [DOI] [PubMed] [Google Scholar]
- 57.Piatt MD, Salicioni AM, Hunt DF, Visconti PE. Use of differential isotopic labeling and mass spectrometry to analyze capacitation-associated changes in the phosphorylation status of mouse sperm proteins. Journal of Proteome Research. 2009;8(3):1431–1440. doi: 10.1021/pr800796j. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Goupil S, La Salle S, Trasler JM, Bordeleau LJ, Leclerc P. Developmental expression of Src-related tyrosine kinases in the mouse testis. Journal of Andrology. 2011;32(1):95–110. doi: 10.2164/jandrol.110.010462. [DOI] [PubMed] [Google Scholar]
- 59.Lawson C, Goupil S, Leclerc P. Increased activity of the human sperm tyrosine kinase SRC by the cAMP-dependent pathway in the presence of calcium. Biology of Reproduction. 2008;79(4):657–666. doi: 10.1095/biolreprod.108.070367. [DOI] [PubMed] [Google Scholar]
- 60.Lev S, Yarden Y, Givol D. Dimerization and activation of the kit receptor by monovalent and bivalent binding of the stem cell factor. Journal of Biological Chemistry. 1992;267(22):15970–15977. [PubMed] [Google Scholar]
- 61.Godin I, Deed R, Cooke J, Zsebo K, Dexter M, Wylie CC. Effects of the steel gene product on mouse primordial germ cells in culture. Nature. 1991;352(6338):807–809. doi: 10.1038/352807a0. [DOI] [PubMed] [Google Scholar]
- 62.Farini D, La Sala G, Tedesco M, De Felici M. Chemoattractant action and molecular signaling pathways of Kit ligand on mouse primordial germ cells. Developmental Biology. 2007;306(2):572–583. doi: 10.1016/j.ydbio.2007.03.031. [DOI] [PubMed] [Google Scholar]
- 63.Yoshinaga K, Nishikawa S, Ogawa M, et al. Role of c-kit in mouse spermatogenesis: identification of spermatogonia as a specific site of c-kit expression and function. Development. 1991;113(2):689–699. doi: 10.1242/dev.113.2.689. [DOI] [PubMed] [Google Scholar]
- 64.Oatley JM, Avarbock MR, Brinster RL. Glial cell line-derived neurotrophic factor regulation of genes essential for self-renewal of mouse spermatogonial stem cells is dependent on Src family kinase signaling. Journal of Biological Chemistry. 2007;282(35):25842–25851. doi: 10.1074/jbc.M703474200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Morimoto H, Kanatsu-Shinohara M, Takashima S, et al. Phenotypic plasticity of mouse spermatogonial stem cells. PLoS ONE. 2009;4(11) doi: 10.1371/journal.pone.0007909.e7909 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Navon A, Schwarz Y, Hazan B, Kassir Y, Nir U. Meiosis-dependent tyrosine phosphorylation of a yeast protein related to the mouse p51(ferT) Molecular and General Genetics. 1994;244(2):160–167. doi: 10.1007/BF00283517. [DOI] [PubMed] [Google Scholar]
- 67.Vincent S, Segretain D, Nishikawa S, et al. Stage-specific expression of the Kit receptor and its ligand (KL) during male gametogenesis in the mouse: a Kit-KL interaction critical for meiosis. Development. 1998;125(22):4585–4593. doi: 10.1242/dev.125.22.4585. [DOI] [PubMed] [Google Scholar]
- 68.Pellegrini M, Filipponi D, Gori M, et al. ATRA and KL promote differentiation toward the meiotic program of male germ cells. Cell Cycle. 2008;7(24):3878–3888. doi: 10.4161/cc.7.24.7262. [DOI] [PubMed] [Google Scholar]
- 69.Lalancette C, Bordeleau LJ, Faure RL, Leclerc P. Bull testicular haploid germ cells express a messenger encoding for a truncated form of the protein tyrosine kinase HCK. Molecular Reproduction and Development. 2006;73(4):520–530. doi: 10.1002/mrd.20422. [DOI] [PubMed] [Google Scholar]
- 70.Kierszenbaum AL, Rivkin E, Talmor-Cohen A, Shalgi R, Tres LL. Expression of full-length and truncated fyn tyrosine kinase transcripts and encoded proteins during spermatogenesis and localization during acrosome biogenesis and fertilization. Molecular Reproduction and Development. 2009;76(9):832–843. doi: 10.1002/mrd.21049. [DOI] [PubMed] [Google Scholar]
- 71.Bordeleau LJ, Leclerc P. Expression of hck-tr, a truncated form of the src-related tyrosine kinase hck, in bovine spermatozoa and testis. Molecular Reproduction and Development. 2008;75(5):828–837. doi: 10.1002/mrd.20814. [DOI] [PubMed] [Google Scholar]
- 72.Albanesi C, Geremia R, Giorgio M, Dolci S, Sette C, Rossi P. A cell- and developmental stage-specific promoter drives the expression of a truncated c-kit protein during mouse spermatid elongation. Development. 1996;122(4):1291–1302. doi: 10.1242/dev.122.4.1291. [DOI] [PubMed] [Google Scholar]
- 73.Rossi P, Marziali G, Albanesi C, Charlesworth A, Geremia R, Sorrentino V. A novel c-kit transcript, potentially encoding a truncated receptor, originates within a kit gene intron in mouse spermatids. Developmental Biology. 1992;152(1):203–207. doi: 10.1016/0012-1606(92)90172-d. [DOI] [PubMed] [Google Scholar]
- 74.Paronetto MP, Venables JP, Elliott DJ, Geremia R, Rossi P, Sette C. Tr-kit promotes the formation of a multimolecular complex composed by Fyn, PLCγ1 and Sam68. Oncogene. 2003;22(54):8707–8715. doi: 10.1038/sj.onc.1207016. [DOI] [PubMed] [Google Scholar]
- 75.Muciaccia B, Sette C, Paronetto MP, et al. Expression of a truncated form of KIT tyrosine kinase in human spermatozoa correlates with sperm DNA integrity. Human Reproduction. 2010;25(9):2188–2202. doi: 10.1093/humrep/deq168. [DOI] [PubMed] [Google Scholar]
- 76.Siu MKY, Mruk DD, Lee WM, Cheng CY. Adhering junction dynamics in the testis are regulated by an interplay of β1-integrin and focal adhesion complex-associated proteins. Endocrinology. 2003;144(5):2141–2163. doi: 10.1210/en.2002-221035. [DOI] [PubMed] [Google Scholar]
- 77.Chen YM, Lee NPY, Mruk DD, Lee WM, Cheng CY. Fer kinase/FerT and adherens junction dynamics in the testis: an in vitro and in vivo study. Biology of Reproduction. 2003;69(2):656–672. doi: 10.1095/biolreprod.103.016881. [DOI] [PubMed] [Google Scholar]
- 78.Maekawa M, Toyama Y, Yasuda M, Yagi T, Yuasa S. Fyn tyrosine kinase in sertoli cells is involved in mouse spermatogenesis. Biology of Reproduction. 2002;66(1):211–221. doi: 10.1095/biolreprod66.1.211. [DOI] [PubMed] [Google Scholar]
- 79.Siu MKY, Wong CH, Lee WM, Cheng CY. Sertoli-germ cell anchoring junction dynamics in the testis are regulated by an interplay of lipid and protein kinases. Journal of Biological Chemistry. 2005;280(26):25029–25047. doi: 10.1074/jbc.M501049200. [DOI] [PubMed] [Google Scholar]
- 80.Rassoulzadegan M, Paquis-Flucklinger V, Bertino B, et al. Transmeiotic differentiation of male germ cells in culture. Cell. 1993;75(5):997–1006. doi: 10.1016/0092-8674(93)90543-y. [DOI] [PubMed] [Google Scholar]
- 81.Hofmann MC, Hess RA, Goldberg E, Millán JL. Immortalized germ cells undergo meiosis in vitro. Proceedings of the National Academy of Sciences of the United States of America. 1994;91(12):5533–5537. doi: 10.1073/pnas.91.12.5533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Sato T, Katagiri K, Gohbara A, et al. In vitro production of functional sperm in cultured neonatal mouse testes. Nature. 2011;471(7339):504–507. doi: 10.1038/nature09850. [DOI] [PubMed] [Google Scholar]
- 83.Miki K, Qu W, Goulding EH, et al. Glyceraldehyde 3-phosphate dehydrogenase-S, a sperm-specific glycolytic enzyme, is required for sperm motility and male fertility. Proceedings of the National Academy of Sciences of the United States of America. 2004;101(47):16501–16506. doi: 10.1073/pnas.0407708101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Nakamura M, Okinaga S, Arai K. Metabolism of round spermatids: evidence that lactate is preferred substrate. The American journal of physiology. 1984;247(2):E234–E242. doi: 10.1152/ajpendo.1984.247.2.E234. [DOI] [PubMed] [Google Scholar]
- 85.Nakamura M, Okinaga S, Arai K. Metabolism of pachytene primary spermatocytes from rat testes: pyruvate maintenance of adenosine triphosphate level. Biology of Reproduction. 1984;30(5):1187–1197. doi: 10.1095/biolreprod30.5.1187. [DOI] [PubMed] [Google Scholar]
- 86.Storey BT, Kayne FJ. Energy metabolism of spermatozoa. V. The Embden Myerhof pathway of glycolysis: activities of pathway enzymes in hypotonically treated rabbit epididymal spermatozoa. Fertility and Sterility. 1975;26(12):1257–1265. [PubMed] [Google Scholar]
- 87.Calvin HI, Bedford JM. Formation of disulphide bonds in the nucleus and accessory structures of mammalian spermatozoa during maturation in the epididymis. Journal of reproduction and fertility. Supplement. 1971;13(supplement 13):65–75. [PubMed] [Google Scholar]
- 88.Cornwall GA, Vindivich D, Tillman S, Chang TS. The effect of sulfhydryl oxidation on the morphology of immature hamster epididymal spermatozoa induced to acquire motility in vitro. Biology of Reproduction. 1988;39(1):141–155. doi: 10.1095/biolreprod39.1.141. [DOI] [PubMed] [Google Scholar]
- 89.Seligman J, Zipser Y, Kosower NS. Tyrosine phosphorylation, thiol status, and protein tyrosine phosphatase in rat epididymal spermatozoa. Biology of Reproduction. 2004;71(3):1009–1015. doi: 10.1095/biolreprod.104.028035. [DOI] [PubMed] [Google Scholar]
- 90.Devi KU, Ahmad MB, Shivaji S. A maturation-related differential phosphorylation of the plasma membrane proteins of the epididymal spermatozoa of the hamster by endogenous protein kinases. Molecular Reproduction and Development. 1997;47(3):341–350. doi: 10.1002/(SICI)1098-2795(199707)47:3<341::AID-MRD13>3.0.CO;2-0. [DOI] [PubMed] [Google Scholar]
- 91.Fàbrega A, Puigmulé M, Yeste M, Casas I, Bonet S, Pinart E. Impact of epididymal maturation, ejaculation and in vitro capacitation on tyrosine phosphorylation patterns exhibited of boar (Sus domesticus) spermatozoa. Theriogenology. 2011;76(7):1356–1366. doi: 10.1016/j.theriogenology.2011.06.007. [DOI] [PubMed] [Google Scholar]
- 92.Jones R, James PS, Oxley D, Coadwell J, Suzuki-Toyota F, Howes EA. The equatorial subsegment in mammalian spermatozoa is enriched in tyrosine phosphorylated proteins. Biology of Reproduction. 2008;79(3):421–431. doi: 10.1095/biolreprod.107.067314. [DOI] [PubMed] [Google Scholar]
- 93.Lewis B, Aitken RJ. A redox-regulated tyrosine phosphorylation cascade in rat spermatozoa. Journal of Andrology. 2001;22(4):611–622. [PubMed] [Google Scholar]
- 94.Lin M, Yun HL, Xu W, Baker MA, Aitken RJ. Ontogeny of tyrosine phosphorylation-signaling pathways during spermatogenesis and epididymal maturation in the mouse. Biology of Reproduction. 2006;75(4):588–597. doi: 10.1095/biolreprod.106.052712. [DOI] [PubMed] [Google Scholar]
- 95.Ecroyd H, Asquith KL, Jones RC, Aitken RJ. The development of signal transduction pathways during epididymal maturation is calcium dependent. Developmental Biology. 2004;268(1):53–63. doi: 10.1016/j.ydbio.2003.12.015. [DOI] [PubMed] [Google Scholar]
- 96.Baker MA, Witherdin R, Hetherington L, Cunningham-Smith K, Aitken RJ. Identification of post-translational modifications that occur during sperm maturation using difference in two-dimensional gel electrophoresis. Proteomics. 2005;5(4):1003–1012. doi: 10.1002/pmic.200401100. [DOI] [PubMed] [Google Scholar]
- 97.Baker MA, Smith ND, Hetherington L, Pelzing M, Condina MR, Aitken RJ. Use of titanium dioxide to find phosphopeptide and total protein changes during epididymal sperm maturation. Journal of Proteome Research. 2011;10(3):1004–1017. doi: 10.1021/pr1007224. [DOI] [PubMed] [Google Scholar]
- 98.Sonnenberg-Riethmacher E, Walter B, Riethmacher D, Gödecke S, Birchmeier C. The c-ros tyrosine kinase receptor controls regionalization and differentiation of epithelial cells in the epididymis. Genes and Development. 1996;10(10):1184–1193. doi: 10.1101/gad.10.10.1184. [DOI] [PubMed] [Google Scholar]
- 99.Keilhack H, Müller M, Böhmer SA, et al. Negative regulation of Ros receptor tyrosine kinase signaling: an epithelial function of the SH2 domain protein tyrosine phosphatase SHP-1. Journal of Cell Biology. 2001;152(2):325–334. doi: 10.1083/jcb.152.2.325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Syntin P, Dacheux F, Druart X, Gatti JL, Okamura N, Dacheux JL. Characterization and identification of proteins secreted in the various regions of the adult boar epididymis. Biology of Reproduction. 1996;55(5):956–974. doi: 10.1095/biolreprod55.5.956. [DOI] [PubMed] [Google Scholar]
- 101.Dacheux JL, Belghazi M, Lanson Y, Dacheux F. Human epididymal secretome and proteome. Molecular and Cellular Endocrinology. 2006;250(1-2):36–42. doi: 10.1016/j.mce.2005.12.022. [DOI] [PubMed] [Google Scholar]
- 102.Duncan AE, Fraser LR. Cyclic AMP-dependent phosphorylation of epididymal mouse sperm proteins during capacitation in vitro: identification of an M(r) 95, 000 phosphotyrosine-containing protein. Journal of Reproduction and Fertility. 1993;97(1):287–299. doi: 10.1530/jrf.0.0970287. [DOI] [PubMed] [Google Scholar]
- 103.Visconti PE, Bailey JL, Moore GD, Pan D, Olds-Clarke P, Kopf GS. Capacitation of mouse spermatozoa. 1. Correlation between the capacitation state and protein tyrosine phosphorylation. Development. 1995;121(4):1129–1137. doi: 10.1242/dev.121.4.1129. [DOI] [PubMed] [Google Scholar]
- 104.Visconti PE, Moore GD, Bailey JL, et al. Capacitation of mouse spermatozoa. II. Protein tyrosine phosphorylation and capacitation are regulated by a cAMP-dependent pathway. Development. 1995;121(4):1139–1150. doi: 10.1242/dev.121.4.1139. [DOI] [PubMed] [Google Scholar]
- 105.Baldi E, Luconi M, Bonaccorsi L, Krausz C, Forti G. Human sperm activation during capacitation and acrosome reaction: role of calcium, protein phosphorylation and lipid remodelling pathways. Frontiers in Bioscience. 1996;1:d189–d205. doi: 10.2741/a125. [DOI] [PubMed] [Google Scholar]
- 106.Pukazhenthi BS, Wildt DE, Ottinger MA, Howard J. Compromised sperm protein phosphorylation after capacitation, swim-up, and zona pellucida exposure in teratospermic domestic cats. Journal of Andrology. 1996;17(4):409–419. [PubMed] [Google Scholar]
- 107.Pukazhenthi BS, Long JA, Wildt DE, Ottinger MA, Armstrong DL, Howard J. Regulation of sperm function by protein tyrosine phosphorylation in diverse wild felid species. Journal of Andrology. 1998;19(6):675–685. [PubMed] [Google Scholar]
- 108.Carrera A, Moos J, Ning XP, et al. Regulation of protein tyrosine phosphorylation in human sperm by a calcium/calmodulin-dependent mechanism: identification of A kinase anchor proteins as major substrates for tyrosine phosphorylation. Developmental Biology. 1996;180(1):284–296. doi: 10.1006/dbio.1996.0301. [DOI] [PubMed] [Google Scholar]
- 109.Mandal A, Naaby-Hansen S, Wolkowicz MJ, et al. FSP95, a testis-specific 95-kilodalton fibrous sheath antigen that undergoes tyrosine phosphorylation in capacitated human spermatozoa. Biology of Reproduction. 1999;61(5):1184–1197. doi: 10.1095/biolreprod61.5.1184. [DOI] [PubMed] [Google Scholar]
- 110.Visconti PE, Johnson LR, Oyaski M, et al. Regulation, localization, and anchoring of protein kinase a subunits during mouse sperm capacitation. Developmental Biology. 1997;192(2):351–363. doi: 10.1006/dbio.1997.8768. [DOI] [PubMed] [Google Scholar]
- 111.Naz RK. c-abl proto-oncoprotein is expressed and tyrosine phosphorylated in human sperm cell. Molecular Reproduction and Development. 1998;51(2):210–217. doi: 10.1002/(SICI)1098-2795(199810)51:2<210::AID-MRD11>3.0.CO;2-8. [DOI] [PubMed] [Google Scholar]
- 112.Visconti PE, Ning X, Fornés MW, et al. Cholesterol efflux-mediated signal transduction in mammalian sperm: cholesterol release signals an increase in protein tyrosine phosphorylation during mouse sperm capacitation. Developmental Biology. 1999;214(2):429–443. doi: 10.1006/dbio.1999.9428. [DOI] [PubMed] [Google Scholar]
- 113.Visconti PE, Galantino-Homer H, Ning X, et al. Cholesterol efflux-mediated signal transduction in mammalian sperm: β-cyclodextrins initiate transmembrane signaling leading to an increase in protein tyrosine phosphorylation and capacitation. Journal of Biological Chemistry. 1999;274(5):3235–3242. doi: 10.1074/jbc.274.5.3235. [DOI] [PubMed] [Google Scholar]
- 114.Osheroff JE, Visconti PE, Valenzuela JP, Travis AJ, Alvarez J, Kopf GS. Regulation of human sperm capacitation by a cholesterol efflux-stimulated signal transduction pathway leading to protein kinase A-mediated up-regulation of protein tyrosine phosphorylation. Molecular Human Reproduction. 1999;5(11):1017–1026. doi: 10.1093/molehr/5.11.1017. [DOI] [PubMed] [Google Scholar]
- 115.Si Y, Olds-Clarke P. Evidence for the involvement of calmodulin in mouse sperm capacitation. Biology of Reproduction. 2000;62(5):1231–1239. doi: 10.1095/biolreprod62.5.1231. [DOI] [PubMed] [Google Scholar]
- 116.Zeng HT, Tulsiani DRP. Calmodulin antagonists differentially affect capacitation-associated protein tyrosine phosphorylation of mouse sperm components. Journal of Cell Science. 2003;116(10):1981–1989. doi: 10.1242/jcs.00396. [DOI] [PubMed] [Google Scholar]
- 117.Huang YH, Chu ST, Chen YH. A seminal vesicle autoantigen of mouse is able to suppress sperm capacitation-related events stimulated by serum albumin. Biology of Reproduction. 2000;63(5):1562–1566. doi: 10.1095/biolreprod63.5.1562. [DOI] [PubMed] [Google Scholar]
- 118.Huang YH, Kuo SP, Lin MH, et al. Signals of seminal vesicle autoantigen suppresses bovine serum albumin-induced capacitation in mouse sperm. Biochemical and Biophysical Research Communications. 2005;338(3):1564–1571. doi: 10.1016/j.bbrc.2005.10.120. [DOI] [PubMed] [Google Scholar]
- 119.Adeoya-Osiguwa SA, Fraser LR. Fertilization promoting peptide and adenosine, acting as first messengers, regulate cAMP production and consequent protein tyrosine phosphorylation in a capacitation-dependent manner. Molecular Reproduction and Development. 2000;57(4):384–392. doi: 10.1002/1098-2795(200012)57:4<384::AID-MRD11>3.0.CO;2-U. [DOI] [PubMed] [Google Scholar]
- 120.Funahashi H, Asano A, Fujiwara T, Nagai T, Niwa K, Fraser LR. Both fertilization promoting peptide and adenosine stimulate capacitation but inhibit spontaneous acrosome loss in ejaculated boar spermatozoa in vitro. Molecular Reproduction and Development. 2000;55(1):117–124. doi: 10.1002/(SICI)1098-2795(200001)55:1<117::AID-MRD16>3.0.CO;2-7. [DOI] [PubMed] [Google Scholar]
- 121.Urner F, Leppens-Luisier G, Sakkas D. Protein tyrosine phosphorylation in sperm during gamete interaction in the mouse: the influence of glucose. Biology of Reproduction. 2001;64(5):1350–1357. doi: 10.1095/biolreprod64.5.1350. [DOI] [PubMed] [Google Scholar]
- 122.Demarco IA, Espinosa F, Edwards J, et al. Involvement of a Na+/HCO− 3 cotransporter in mouse sperm capacitation. Journal of Biological Chemistry. 2003;278(9):7001–7009. doi: 10.1074/jbc.M206284200. [DOI] [PubMed] [Google Scholar]
- 123.Brener E, Rubinstein S, Cohen G, Shternall K, Rivlin J, Breitbart H. Remodeling of the actin cytoskeleton during mammalian sperm capacitation and acrosome reaction. Biology of Reproduction. 2003;68(3):837–845. doi: 10.1095/biolreprod.102.009233. [DOI] [PubMed] [Google Scholar]
- 124.Buffone MG, Ijiri TW, Cao W, Merdiushev T, Aghajanian HK, Gerton GL. Heads or tails? Structural events and molecular mechanisms that promote mammalian sperm acrosomal exocytosis and motility. Molecular Reproduction and Development. 2012;79(1):4–18. doi: 10.1002/mrd.21393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Ecroyd HW, Jones RC, Aitken RJ. Endogenous redox activity in mouse spermatozoa and its role in regulating the tyrosine phosphorylation events associated with sperm capacitation. Biology of Reproduction. 2003;69(1):347–354. doi: 10.1095/biolreprod.102.012716. [DOI] [PubMed] [Google Scholar]
- 126.Aitken RJ, Baker MA. Oxidative stress and male reproductive biology. Reproduction, Fertility and Development. 2004;16(5):581–588. doi: 10.10371/RD03089. [DOI] [PubMed] [Google Scholar]
- 127.Ecroyd H, Jones RC, Aitken RJ. Tyrosine phosphorylation of HSP-90 during mammalian sperm capacitation. Biology of Reproduction. 2003;69(6):1801–1807. doi: 10.1095/biolreprod.103.017350. [DOI] [PubMed] [Google Scholar]
- 128.Asquith KL, Baleato RM, McLaughlin EA, Nixon B, Aitken RJ. Tyrosine phosphorylation activates surface chaperones facilitating sperm-zona recognition. Journal of Cell Science. 2004;117:3645–3657. doi: 10.1242/jcs.01214. [DOI] [PubMed] [Google Scholar]
- 129.Mitchell LA, Nixon B, Aitken RJ. Analysis of chaperone proteins associated with human spermatozoa during capacitation. Molecular Human Reproduction. 2007;13(9):605–613. doi: 10.1093/molehr/gam043. [DOI] [PubMed] [Google Scholar]
- 130.Quill TA, Sugden SA, Rossi KL, Doolittle LK, Hammer RE, Garbers DL. Hyperactivated sperm motility driven by CatSper2 is required for fertilization. Proceedings of the National Academy of Sciences of the United States of America. 2003;100(25):14869–14874. doi: 10.1073/pnas.2136654100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Baker MA, Hetherington L, Ecroyd H, Roman SD, Aitken RJ. Analysis of the mechanism by which calcium negatively regulates the tyrosine phosphorylation cascade associated with sperm capacitation. Journal of Cell Science. 2004;117:211–222. doi: 10.1242/jcs.00842. [DOI] [PubMed] [Google Scholar]
- 132.Rodeheffer C, Shur BD. Sperm from β1,4-galactosyltransferase l-null mice exhibit precocious capacitation. Development. 2004;131(3):491–501. doi: 10.1242/dev.00885. [DOI] [PubMed] [Google Scholar]
- 133.Mededovic S, Fraser LR. Angiotensin II stimulates cAMP production and protein tyrosine phosphorylation in mouse spermatozoa. Reproduction. 2004;127(5):601–612. doi: 10.1530/rep.1.00062. [DOI] [PubMed] [Google Scholar]
- 134.Hess KC, Jones BH, Marquez B, et al. The “soluble” adenylyl cyclase in sperm mediates multiple signaling events required for fertilization. Developmental Cell. 2005;9(2):249–259. doi: 10.1016/j.devcel.2005.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Xie F, Garcia MA, Carlson AE, et al. Soluble adenylyl cyclase (sAC) is indispensable for sperm function and fertilization. Developmental Biology. 2006;296(2):353–362. doi: 10.1016/j.ydbio.2006.05.038. [DOI] [PubMed] [Google Scholar]
- 136.Nixon B, MacIntyre DA, Mitchell LA, Gibbs GM, O’Bryan M, Aitken RJ. The identification of mouse sperm-surface-associated proteins and characterization of their ability to act as decapacitation factors. Biology of Reproduction. 2006;74(2):275–287. doi: 10.1095/biolreprod.105.044644. [DOI] [PubMed] [Google Scholar]
- 137.Thundathil JC, Anzar M, Buhr MM. Na+/K+ ATPase as a signaling molecule during bovine sperm capacitation. Biology of Reproduction. 2006;75(3):308–317. doi: 10.1095/biolreprod.105.047852. [DOI] [PubMed] [Google Scholar]
- 138.Tang JB, Chen YH. Identification of a tyrosine-phosphorylated CCCTC-binding nuclear factor in capacitated mouse spermatozoa. Proteomics. 2006;6(17):4800–4807. doi: 10.1002/pmic.200600256. [DOI] [PubMed] [Google Scholar]
- 139.Kawano N, Yoshida M. Semen-coagulating protein, SVS2, in mouse seminal plasma controls sperm fertility. Biology of Reproduction. 2007;76(3):353–361. doi: 10.1095/biolreprod.106.056887. [DOI] [PubMed] [Google Scholar]
- 140.Cao W, Aghajanian HK, Haig-Ladewig LA, Gerton GL. Sorbitol can fuel mouse sperm motility and protein tyrosine phosphorylation via sorbitol dehydrogenase. Biology of Reproduction. 2009;80(1):124–133. doi: 10.1095/biolreprod.108.068882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Wertheimer EV, Salicioni AM, Liu W, et al. Chloride is essential for capacitation and for the capacitation-associated increase in tyrosine phosphorylation. Journal of Biological Chemistry. 2008;283(51):35539–35550. doi: 10.1074/jbc.M804586200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Kota V, Dhople VM, Shivaji S. Tyrosine phosphoproteome of hamster spermatozoa: role of glycerol-3-phosphate dehydrogenase 2 in sperm capacitation. Proteomics. 2009;9(7):1809–1826. doi: 10.1002/pmic.200800519. [DOI] [PubMed] [Google Scholar]
- 143.Kota V, Rai P, Weitzel JM, Middendorff R, Bhande SS, Shivaji S. Role of glycerol-3-phosphate dehydrogenase 2 in mouse sperm capacitation. Molecular Reproduction and Development. 2010;77(9):773–783. doi: 10.1002/mrd.21218. [DOI] [PubMed] [Google Scholar]
- 144.Gyamera-Acheampong C, Vasilescu J, Figeys D, Mbikay M. PCSK4-null sperm display enhanced protein tyrosine phosphorylation and ADAM2 proteolytic processing during in vitro capacitation. Fertility and Sterility. 2010;93(4):1112–1123. doi: 10.1016/j.fertnstert.2008.12.013. [DOI] [PubMed] [Google Scholar]
- 145.Nixon B, Bielanowicz A, Anderson AL, et al. Elucidation of the signaling pathways that underpin capacitation-associated surface phosphotyrosine expression in mouse spermatozoa. Journal of Cellular Physiology. 2010;224(1):71–83. doi: 10.1002/jcp.22090. [DOI] [PubMed] [Google Scholar]
- 146.Shimada M, Yanai Y, Okazaki T, et al. Hyaluronan fragments generated by sperm-secreted hyaluronidase stimulate cytokine/chemokine production via the TLR2 and TLR4 pathway in cumulus cells of ovulated COCs, which may enhance fertilization. Development. 2008;135(11):2001–2011. doi: 10.1242/dev.020461. [DOI] [PubMed] [Google Scholar]
- 147.Da Ros VG, Maldera JA, Willis WD, et al. Impaired sperm fertilizing ability in mice lacking Cysteine-RIch Secretory Protein 1 (CRISP1) Developmental Biology. 2008;320(1):12–18. doi: 10.1016/j.ydbio.2008.03.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Baker MA, Hetherington L, Aitken RJ. Identification of SRC as a key PKA-stimulated tyrosine kinase involved in the capacitation-associated hyperactivation of murine spermatozoa. Journal of Cell Science. 2006;119:3182–3192. doi: 10.1242/jcs.03055. [DOI] [PubMed] [Google Scholar]
- 149.Mitchel LA, Nixon B, Baker MA, Aitken RJ. Investigation of the role of SRC in capacitation-associated tyrosine phosphorylation of human spermatozoa. Molecular Human Reproduction. 2008;14(4):235–243. doi: 10.1093/molehr/gan007. [DOI] [PubMed] [Google Scholar]
- 150.Krapf D, Arcelay E, Wertheimer EV, et al. Inhibition of Ser/Thr phosphatases induces capacitation-associated signaling in the presence of Src kinase inhibitors. Journal of Biological Chemistry. 2010;285(11):7977–7985. doi: 10.1074/jbc.M109.085845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Cotton L, Gibbs GM, Sanchez-Partida LG, Morrison JR, de Kretser DM, O’Bryan MK. Erratum: FGFR-1 signaling is involved in spermiogenesis and sperm capacitation. Journal of Cell Science. 2006;119:75–84. doi: 10.1242/jcs.02704. [DOI] [PubMed] [Google Scholar]
- 152.Baker MA, Hetherington L, Curry B, Aitken RJ. Phosphorylation and consequent stimulation of the tyrosine kinase c-Abl by PKA in mouse spermatozoa; its implications during capacitation. Developmental Biology. 2009;333(1):57–66. doi: 10.1016/j.ydbio.2009.06.022. [DOI] [PubMed] [Google Scholar]
- 153.Leyton L, Saling P. 95 kd sperm protein binds ZP3 and serve as tyrosine kinase substrates in response to zona binding. Cell. 1989;57(7):1123–1130. doi: 10.1016/0092-8674(89)90049-4. [DOI] [PubMed] [Google Scholar]
- 154.Leyton L, LeGuen P, Bunch D, Saling PM. Regulation of mouse gamete interaction by a sperm tyrosine kinase. Proceedings of the National Academy of Sciences of the United States of America. 1992;89(24):11692–11695. doi: 10.1073/pnas.89.24.11692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Naz RK, Morte C, Ahmad K, Martinez P. Hexokinase present in human sperm is not tyrosine phosphorylated but its antibodies affect fertilizing capacity. Journal of Andrology. 1996;17(2):143–150. [PubMed] [Google Scholar]
- 156.Travis AJ, Foster JA, Rosenbaum NA, et al. Targeting of a germ cell-specific type 1 hexokinase lacking a porin- binding domain to the mitochondria as well as to the head and fibrous sheath of murine spermatozoa. Molecular Biology of the Cell. 1998;9(2):263–276. doi: 10.1091/mbc.9.2.263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Benoff S. Modelling human sperm-egg interactions in vitro: signal transduction pathways regulating the acrosome reaction. Molecular Human Reproduction. 1998;4(5):453–471. doi: 10.1093/molehr/4.5.453. [DOI] [PubMed] [Google Scholar]
- 158.Berruti G, Martegani E. Identification of proteins cross-reactive to phosphotyrosine antibodies and of a tyrosine kinase activity in boar spermatozoa. Journal of Cell Science. 1989;93:667–674. doi: 10.1242/jcs.93.4.667. [DOI] [PubMed] [Google Scholar]
- 159.Naz RK. Protein tyrosine phosphorylation and signal transduction during capacitation-acrosome reaction and zona pellucida binding in human sperm. Systems Biology in Reproductive Medicine. 1996;37(1):47–55. doi: 10.3109/01485019608988501. [DOI] [PubMed] [Google Scholar]
- 160.Naz RK, Rajesh PB. Role of tyrosine phosphorylation in sperm capacitation/acrosome reaction. Reproductive Biology and Endocrinology. 2004;2, article 75 doi: 10.1186/1477-7827-2-75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Tomes CN, Mcmaster CR, Saling PM. Activation of mouse sperm phosphatidylinositol-4,5 bisphosphate-phospholipase C by zona pellucida is modulated by tyrosine phosphorylation. Molecular Reproduction and Development. 1996;43(2):196–204. doi: 10.1002/(SICI)1098-2795(199602)43:2<196::AID-MRD9>3.0.CO;2-M. [DOI] [PubMed] [Google Scholar]
- 162.Fukami K, Yoshida M, Inoue T, et al. Phospholipase Cδ4 is required for Ca2+ mobilization essential for acrosome reaction in sperm. Journal of Cell Biology. 2003;161(1):79–88. doi: 10.1083/jcb.200210057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Fukami K, Nakao K, Inoue T, et al. Requirement of phospholipase Cδ4 for the zona pellucida-induced acrosome reaction. Science. 2001;292(5518):920–923. doi: 10.1126/science.1059042. [DOI] [PubMed] [Google Scholar]
- 164.Fisher HM, Brewis IA, Barratt CLR, Cooke ID, Moore HDM. Phosphoinositide 3-kinase is involved in the induction of the human sperm acrosome reaction downstream of tyrosine phosphorylation. Molecular Human Reproduction. 1998;4(9):849–855. doi: 10.1093/molehr/4.9.849. [DOI] [PubMed] [Google Scholar]
- 165.Breitbart H, Rotman T, Rubinstein S, Etkovitz N. Role and regulation of PI3K in sperm capacitation and the acrosome reaction. Molecular and Cellular Endocrinology. 2010;314(2):234–238. doi: 10.1016/j.mce.2009.06.009. [DOI] [PubMed] [Google Scholar]
- 166.Luconi M, Bonaccorsi L, Krausz C, Gervasi G, Forti G, Baldi E. Stimulation of protein tyrosine phosphorylation by platelet-activating factor and progesterone in human spermatozoa. Molecular and Cellular Endocrinology. 1995;108(1-2):35–42. doi: 10.1016/0303-7207(95)92576-a. [DOI] [PubMed] [Google Scholar]
- 167.Bonaccorsi L, Luconi M, Forti G, Baldi E. Tyrosine kinase inhibition reduces the plateau phase of the calcium increase in response to progesterone in human sperm. FEBS Letters. 1995;364(1):83–86. doi: 10.1016/0014-5793(95)00369-k. [DOI] [PubMed] [Google Scholar]
- 168.Meizel S, Turner KO. Chloride efflux during the progesterone-initiated human sperm acrosome reaction is inhibited by lavendustin A, a tyrosine kinase inhibitor. Journal of Andrology. 1996;17(4):327–330. [PubMed] [Google Scholar]
- 169.Pukazhenthi BS, Wildt DE, Ottinger MA, Howard J. Inhibition of domestic cat spermatozoa acrosome reaction and zona pellucida penetration by tyrosine kinase inhibitors. Molecular Reproduction and Development. 1998;49(1):48–57. doi: 10.1002/(SICI)1098-2795(199801)49:1<48::AID-MRD6>3.0.CO;2-O. [DOI] [PubMed] [Google Scholar]
- 170.Dorval V, Dufour M, Leclerc P. Role of protein tyrosine phosphorylation in the thapsigargin-induced intracellular Ca2+ store depletion during human sperm acrosome reaction. Molecular Human Reproduction. 2003;9(3):125–131. doi: 10.1093/molehr/gag017. [DOI] [PubMed] [Google Scholar]
- 171.Morte C, Iborra A, Martínez P. Phosphorylation of Shc proteins in human sperm in response to capacitation and progesterone treatment. Molecular Reproduction and Development. 1998;50(1):113–120. doi: 10.1002/(SICI)1098-2795(199805)50:1<113::AID-MRD14>3.0.CO;2-9. [DOI] [PubMed] [Google Scholar]
- 172.Picherit-Marchenay C, Bréchard S, Boucher D, Grizard G. Correlation between tyrosine phosphorylation intensity of a 107 kDa protein band and A23187-induced acrosome reaction in human spermatozoa. Andrologia. 2004;36(6):370–377. doi: 10.1111/j.1439-0272.2004.00634.x. [DOI] [PubMed] [Google Scholar]
- 173.Mor V, Das T, Bhattacharjee M, Chatterjee T. Protein tyrosine phosphorylation of a heparin-binding sperm membrane mitogen (HBSM) is associated with capacitation and acrosome reaction. Biochemical and Biophysical Research Communications. 2007;352(2):404–409. doi: 10.1016/j.bbrc.2006.11.026. [DOI] [PubMed] [Google Scholar]
- 174.Tomes CN, Roggero CM, De Blas G, Saling PM, Mayorga LS. Requirement of protein tyrosine kinase and phosphatase activities for human sperm exocytosis. Developmental Biology. 2004;265(2):399–415. doi: 10.1016/j.ydbio.2003.09.032. [DOI] [PubMed] [Google Scholar]
- 175.Zarelli VEP, Ruete MC, Roggero CM, Mayorga LS, Tomes CN. PTP1B dephosphorylates N-ethylmaleimide-sensitive factor and elicits SNARE complex disassembly during human sperm exocytosis. Journal of Biological Chemistry. 2009;284(16):10491–10503. doi: 10.1074/jbc.M807614200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Finkelstein M, Etkovitz N, Breitbart H. Role and regulation of sperm gelsolin prior to fertilization. Journal of Biological Chemistry. 2010;285(51):39702–39709. doi: 10.1074/jbc.M110.170951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Lax Y, Rubinstein S, Breitbart H. Epidermal growth factor induces acrosomal exocytosis in bovine sperm. FEBS Letters. 1994;339(3):234–238. doi: 10.1016/0014-5793(94)80422-2. [DOI] [PubMed] [Google Scholar]
- 178.Etkovitz N, Tirosh Y, Chazan R, et al. Bovine sperm acrosome reaction induced by G protein-coupled receptor agonists is mediated by epidermal growth factor receptor transactivation. Developmental Biology. 2009;334(2):447–457. doi: 10.1016/j.ydbio.2009.08.002. [DOI] [PubMed] [Google Scholar]
- 179.Daniel L, Etkovitz N, Weiss SR, Rubinstein S, Ickowicz D, Breitbart H. Regulation of the sperm EGF receptor by ouabain leads to initiation of the acrosome reaction. Developmental Biology. 2010;344(2):650–657. doi: 10.1016/j.ydbio.2010.05.490. [DOI] [PubMed] [Google Scholar]
- 180.Sato KI, Sato A, Aoto M, Fukami Y. C-SRC phosphorylates epidermal growth factor receptor on tyrosine 845. Biochemical and Biophysical Research Communications. 1995;215(3):1078–1087. doi: 10.1006/bbrc.1995.2574. [DOI] [PubMed] [Google Scholar]
- 181.Tice DA, Biscardi JS, Nickles AL, Parsons SJ. Mechanism of biological synergy between cellular Src and epidermal growth factor receptor. Proceedings of the National Academy of Sciences of the United States of America. 1999;96(4):1415–1420. doi: 10.1073/pnas.96.4.1415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Biscardi JS, Maa MC, Tice DA, Cox ME, Leu TH, Parsons SJ. C-Src-mediated phosphorylation of the epidermal growth factor receptor on Tyr845 and Tyr1101 is associated with modulation of receptor function. Journal of Biological Chemistry. 1999;274(12):8335–8343. doi: 10.1074/jbc.274.12.8335. [DOI] [PubMed] [Google Scholar]
- 183.Varano G, Lombardi A, Cantini G, Forti G, Baldi E, Luconi M. Src activation triggers capacitation and acrosome reaction but not motility in human spermatozoa. Human Reproduction. 2008;23(12):2652–2662. doi: 10.1093/humrep/den314. [DOI] [PubMed] [Google Scholar]
- 184.Tapia S, Rojas M, Morales P, Ramirez MA, Diaz ES. The laminin-induced acrosome reaction in human sperm is mediated by src kinases and the proteasome. Biology of Reproduction. 2011;85(2):357–366. doi: 10.1095/biolreprod.111.092254. [DOI] [PubMed] [Google Scholar]
- 185.Feng H, Sandlow JI, Sandra A. The c-kit receptor and its possible signaling transduction pathway in mouse spermatozoa. Molecular Reproduction and Development. 1998;49(3):317–326. doi: 10.1002/(SICI)1098-2795(199803)49:3<317::AID-MRD12>3.0.CO;2-T. [DOI] [PubMed] [Google Scholar]
- 186.Krapf D, Visconti PE, Arranz SE, Cabada MO. Egg water from the amphibian Bufo arenarum induces capacitation-like changes in homologous spermatozoa. Developmental Biology. 2007;306(2):516–524. doi: 10.1016/j.ydbio.2007.03.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Baibakov B, Gauthier L, Talbot P, Rankin TL, Dean J. Sperm binding to the zona pellucida is not sufficient to induce acrosome exocytosis. Development. 2007;134(5):933–943. doi: 10.1242/dev.02752. [DOI] [PubMed] [Google Scholar]
- 188.Teijeiro JM, Cabada MO, Marini PE. Sperm binding glycoprotein (SBG) produces calcium and bicarbonate dependent alteration of acrosome morphology and protein tyrosine phosphorylation on boar sperm. Journal of Cellular Biochemistry. 2008;103(5):1413–1423. doi: 10.1002/jcb.21524. [DOI] [PubMed] [Google Scholar]
- 189.Diaz ES, Kong M, Morales P. Effect of fibronectin on proteasome activity, acrosome reaction, tyrosine phosphorylation and intracellular calcium concentrations of human sperm. Human Reproduction. 2007;22(5):1420–1430. doi: 10.1093/humrep/dem023. [DOI] [PubMed] [Google Scholar]
- 190.Gye MC. Changes in sperm phosphotyrosine proteins by human follicular fluid in mice. Archives of Andrology. 2003;49(6):417–422. [PubMed] [Google Scholar]
- 191.Townley IK, Schuyler E, Parker-Gür M, Foltz KR. Expression of multiple Src family kinases in sea urchin eggs and their function in Ca2+ release at fertilization. Developmental Biology. 2009;327(2):465–477. doi: 10.1016/j.ydbio.2008.12.032. [DOI] [PubMed] [Google Scholar]
- 192.Giusti AF, Carroll DJ, Abassi YA, Terasaki M, Foltz KR, Jaffef LA. Requirement of a Src family kinase for initiating calcium release at fertilization in starfish eggs. Journal of Biological Chemistry. 1999;274(41):29318–29322. doi: 10.1074/jbc.274.41.29318. [DOI] [PubMed] [Google Scholar]
- 193.Runft LL, Jaffe LA. Sperm extract injection into ascidian eggs signals Ca2+ release by the same pathway as fertilization. Development. 2000;127(15):3227–3236. doi: 10.1242/dev.127.15.3227. [DOI] [PubMed] [Google Scholar]
- 194.Sato KI, Tokmakov AA, Iwasaki T, Fukami Y. Tyrosine kinase-dependent activation of phospholipase Cγ is required for calcium transient in Xenopus egg fertilization. Developmental Biology. 2000;224(2):453–469. doi: 10.1006/dbio.2000.9782. [DOI] [PubMed] [Google Scholar]
- 195.McGinnis LK, Albertini DF, Kinsey WH. Localized activation of Src-family protein kinases in the mouse egg. Developmental Biology. 2007;306(1):241–254. doi: 10.1016/j.ydbio.2007.03.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Levi M, Maro B, Shalgi R. Fyn kinase is involved in cleavage furrow ingression during meiosis and mitosis. Reproduction. 2010;140(6):827–834. doi: 10.1530/REP-10-0312. [DOI] [PubMed] [Google Scholar]
- 197.Sette C, Bevilacqua A, Bianchini A, Mangia F, Geremia R, Rossi P. Parthenogenetic activation of mouse eggs by microinjection of a truncated c-kit tyrosine kinase present in spermatozoa. Development. 1997;124(11):2267–2274. doi: 10.1242/dev.124.11.2267. [DOI] [PubMed] [Google Scholar]
- 198.Sette C, Bevilacqua A, Geremia R, Rossi P. Involvement of phospholipase Cγ1 in mouse egg activation induced by a truncated form of the c-kit tyrosine kinase present in spermatozoa. Journal of Cell Biology. 1998;142(4):1063–1074. doi: 10.1083/jcb.142.4.1063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Sette C, Paronetto MP, Barchi M, Bevilacqua A, Geremia R, Rossi P. Tr-kit-induced resumption of the cell cycle in mouse eggs requires activation of a Src-like kinase. The EMBO Journal. 2002;21(20):5386–5395. doi: 10.1093/emboj/cdf553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Wu ATH, Sutovsky P, Manandhar G, et al. PAWP, a sperm-specific WW domain-binding protein, promotes meiotic resumption and pronuclear development during fertilization. Journal of Biological Chemistry. 2007;282(16):12164–12175. doi: 10.1074/jbc.M609132200. [DOI] [PubMed] [Google Scholar]
- 201.Simons K, Toomre D. Lipid rafts and signal transduction. Nature Reviews Molecular Cell Biology. 2000;1(1):31–39. doi: 10.1038/35036052. [DOI] [PubMed] [Google Scholar]
- 202.Pike LJ. Rafts defined: a report on the Keystone symposium on lipid rafts and cell function. Journal of Lipid Research. 2006;47(7):1597–1598. doi: 10.1194/jlr.E600002-JLR200. [DOI] [PubMed] [Google Scholar]
- 203.Belton RJ, Jr., Adams NL, Foltz KR. Isolation and characterization of sea urchin egg lipid rafts and their possible function during fertilization. Molecular Reproduction and Development. 2001;59(3):294–305. doi: 10.1002/mrd.1034. [DOI] [PubMed] [Google Scholar]
- 204.Sato KI, Iwasaki T, Ogawa K, Konishi M, Tokmakov AA, Fukami Y. Low density detergent-insoluble membrane of Xenopus eggs: subcellular microdomain for tyrosine kinase signaling in fertilization. Development. 2002;129(4):885–896. doi: 10.1242/dev.129.4.885. [DOI] [PubMed] [Google Scholar]
- 205.Luria A, Vegelyte-Avery V, Stith B, et al. Detergent-free domain isolated from Xenopus egg plasma membrane with properties similar to those of detergent-resistant membranes. Biochemistry. 2002;41(44):13189–13197. doi: 10.1021/bi026107b. [DOI] [PubMed] [Google Scholar]
- 206.Comiskey M, Warner CM. Spatio-temporal localization of membrane lipid rafts in mouse oocytes and cleaving preimplantation embryos. Developmental Biology. 2007;303(2):727–739. doi: 10.1016/j.ydbio.2006.12.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Sato B, Katagiri YU, Miyado K, et al. Lipid rafts enriched in monosialylGb5Cer carrying the stage-specific embryonic antigen-4 epitope are involved in development of mouse preimplantation embryos at cleavage stage. BMC Developmental Biology. 2011;11, article 22 doi: 10.1186/1471-213X-11-22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Hasan AKM, Ou Z, Sakakibara K, et al. Characterization of Xenopus egg membrane microdomains containing uroplakin Ib/III complex: roles of their molecular interactions for subcellular localization and signal transduction. Genes to Cells. 2007;12(2):251–267. doi: 10.1111/j.1365-2443.2007.01048.x. [DOI] [PubMed] [Google Scholar]
- 209.Hasan AKM, Sato KI, Sakakibara K, et al. Uroplakin III, a novel Src substrate in Xenopus egg rafts, is a target for sperm protease essential for fertilization. Developmental Biology. 2005;286(2):483–492. doi: 10.1016/j.ydbio.2005.08.020. [DOI] [PubMed] [Google Scholar]
- 210.Sakakibara K, Sato KI, Yoshino KI, et al. Molecular identification and characterization of Xenopus egg uroplakin III, an egg raft-associated transmembrane protein that is tyrosine-phosphorylated upon fertilization. Journal of Biological Chemistry. 2005;280(15):15029–15037. doi: 10.1074/jbc.M410538200. [DOI] [PubMed] [Google Scholar]
- 211.Sato KI, Tokmakov AA, He CL, et al. Reconstitution of Src-dependent phospholipase Cγ phosphorylation and transient calcium release by using membrane rafts and cell-free extracts from Xenopus eggs. Journal of Biological Chemistry. 2003;278(40):38413–38420. doi: 10.1074/jbc.M302617200. [DOI] [PubMed] [Google Scholar]
- 212.Krapf D, Chun Ruan Y, Wertheimer EV, et al. cSrc is necessary for epididymal development and is incorporated into sperm during epididymal transit. Developmental Biology. 2012;369(1):43–53. doi: 10.1016/j.ydbio.2012.06.017. [DOI] [PMC free article] [PubMed] [Google Scholar]