

Summary
Self-renewing, multipotential retinal stem cells (RSCs) reside in the pigmented ciliary epithelium of the peripheral retina in adult mammals. RSCs can give rise to rhodopsin positive-cells, which can integrate into early postnatal retina, and represent a potentially useful option for cellular therapy. The ability to purify a stem cell population and direct the differentiation toward a particular cell lineage is a challenge facing the application of stem cells in regenerative medicine. Here we use cell sorting to prospectively enrich mouse RSCs based on size, granularity and low expression of P-cadherin and demonstrate that only rare cells with defined properties proliferate to form colonies. We show that clonally-derived mouse and human RSC progeny are multipotent and can differentiate into mature rhodopsin-positive cells with high efficiency using combinations of exogenous culture additives known to influence neural retinal development, including taurine and retinoic acid. This directed RSC differentiation follows the temporal sequence of photoreceptor differentiation in vivo, and the cells exhibit morphology, protein and gene expression consistent with primary cultures of rods in vitro. These results demonstrate that the RSC, an adult stem cell, can be enriched and directed to produce photoreceptors as a first step toward a targeted cell replacement strategy to treat retinal degenerative disease.
Keywords: retina, stem cells, photoreceptor differentiation
Introduction
Adult stem cells are characterized by two cardinal properties: self-renewal and the ability to generate all mature cells of a specific tissue. We previously showed that adult mammalian retinal stem cells (RSCs) can be isolated from the pigmented ciliary epithelium (CE) of the mouse (Tropepe et al., 2000) and human eye (Coles et al., 2004). Self-renewal of mammalian RSCs was demonstrated by clonal single sphere passaging, and the progeny showed multipotency by differentiation to mature retinal cell types including: photoreceptors, bipolar cells, retinal ganglion cells, Müller glia and retinal pigmented epithelium (RPE) (Tropepe et al., 2000). Transplantation assays into neonatal mice demonstrated the ability of clonal RSC progeny to integrate into the various neural layers of developing host retina and differentiate in vivo (Coles et al., 2004). These progeny included a number of mature retinal cell types including ROM1-positive rod photoreceptors. Inoue et al. demonstrated that a small number of RSC-derived photoreceptors could integrate and exhibit functional photoreceptor replacement (Inoue et al., 2010).
Recent reports suggest optimum integration of transplanted photoreceptors into host retina depends on the stage of donor photoreceptor differentiation (MacLaren et al., 2006). The use of stem cells for photoreceptor replacement strategies in humans would require differentiation protocols that can generate highly pure, fate-specified, populations of photoreceptor precursors. However, pan-retinal differentiation of adult RSCs in vitro using serum yields a minority of rhodopsin-positive cells relative to other cell types (Tropepe et al., 2000; Coles et al., 2004; Inoue et al., 2010).
To demonstrate the feasibility of their use in adult stem cell therapy we show that RSCs can be specifically isolated from the CE, and that their progeny can be clonally expanded in vitro and induced to differentiate into rod photoreceptors efficiently with exogenous factors. Recent reports have challenged the stem cell nature of the RSC (Cicero et al., 2009; Gualdoni et al., 2010): the present results strongly support a stem cell hypothesis. These results also give us further insight into the rigorous definition of the stem cell character of RSCs, as compared to other adult stem cell populations. RSCs represent an attractive source of highly purified photoreceptors for use in cell transplantation and avoid ethical issues and tumorigenic concerns that arise from the use embryonic stem cells.
Results
Retinal stem cells can be isolated based on cell size and pigmentation
RSCs are a rare population of pigmented cells (Tropepe et al., 2000) present in the CE intermingled with mature pigmented epithelium cells. There are currently no specific cell surface markers to differentiate between the two cell types. We sought to determine properties unique to RSCs and enrich this population. We confirmed that RSC colonies were derived only from pigmented cells by single cell sorting: 1.8% of single pigmented cells gave rise to clonal colonies and none arose from the non-pigmented cell fraction (n = 1691 cells, 2 experiments). We observed that cells from primary CE dissections present with a large variation in size and pigmentation (Fig. 1A–C). We tested whether RSCs could be reliably enriched based on these properties using cell sorting. When analyzed by flow cytometry, we found that CE samples contained a population of cells with high side scatter (SSC), which was not present in control neural retina samples (Fig. 1D). The high SSC population was not present in sorts from albino animals, consistent with SSC indicating degree of pigmentation/granularity. We verified that this population consisted of pigmented cells by visual inspection of freshly sorted cells. Pigmented cells were subdivided into quadrants based on forward scatter (FSC, indicating cell size) and SSC (Fig. 1E). We found that colonies arose only from the FSC-high/SSC-high population, indicating that RSCs are large, pigmented cells: 0.018±0.005% of cells in this fraction gave rise to a colony, compared with 0.005±0.005% of un-gated (non-sorted) controls (N = 3), representing an approximate 3.5-fold enrichment. This double high population was then further subdivided into an additional 4 quadrants (Fig. 1F) to further characterize the enrichment: 0.19±0.08% of cells in the region with the heaviest pigmentation but smaller size formed colonies compared with 0.019±0.01% of un-gated controls (N = 5, P<0.05), a 10-fold enrichment. The absolute frequency of colony formation was relatively low, perhaps due to our finding that CE cells were exceptionally sensitive to the process of cell sorting. The recovery rate of cells was less than 10% of the starting population, so RSC frequency is likely underestimated. In single cell per well experiments above where live cells were counted 12 hours after sorting, the higher frequency of colony formation can be accounted for by the exclusion of apoptotic cells shortly after being plated. These data confirm that only a subset of pigmented cells from the CE with defined characteristics (medium-large in size and the most heavily pigmented) are capable of proliferating to form clonal RSC colonies.
Fig. 1. RSCs can be isolated from ciliary epithelial (CE) cells using forward and side scatter and express the surface adhesion molecule P-cadherin.

CE dissection yields a variety of cell types: single non-pigmented cells are present (A) along with (B) lightly pigmented, and (C) heavily pigmented cells. Scale bars represent 25 µm. (D) Pigmented cells from dissociated CE make up a population with high side scatter that is not present in dissociated neural retina (NR). (E) This pigmented cell population was divided into 4 quadrants, of which only one gave rise to RSC colonies. (F) The population of large, heavily pigmented cells (P7) was subdivided into 4 new quadrants and RSCs colonies formed only from the smaller, most heavily pigmented cells. Cells were plated at clonal density (10 cells/µL) for colony forming assays. Error bars represent s.e.m., N = 3 for each.. Dissociated CE cells were immunostained for P-cadherin. Cells were divided into negative (P7), low (P5), and high (P6) P-cadherin level (G), FITC intensity along horizontal axis), which was not present in the secondary-only controls. (H) A histogram of cell counts for sorting of dissociated CE cells, while sorting dissociated NR (I) confirms that gate P7 is sorting for P-cadherin-negative cells, as NR cells do not express P-cadherin. All RSC colonies are derived from P-cadherin-low cells, N = 2. Freshly isolated CE expresses high levels of mRNA for P-cadherin, expressed in the pigmented CE and N-cadherin, expressed in the non-pigmented layer. (J) RSC colonies are derived from pigmented CE cells, but also express high levels of both cadherins as spheres are a mix of pigmented RSCs and non-pigmented progenitors. Data shown are deltaCt compared to the endogenous control rplp0 ± s.e.m., N = 2 or 3.
Retinal stem cells express P-cadherin
We also sought to determine whether specific surface markers exist which could identify RSCs. P-cadherin is an adhesion molecule expressed only in the outer pigmented layer of the CE where RSCs reside; inner non-pigmented CE cells express only N-cadherin (Xu et al., 2002; Inagaki et al., 2005). We expected RSCs to express P-cadherin but not N-cadherin. Isolated CE cells were immunostained for P-cadherin and analyzed by flow cytometry (Fig. 1G). We used albino mice in order to avoid masking fluorescence by pigment granules. P-cadherin positive cells comprised 34.5±4.5% of cells collected and all colonies arose from P-cadherin expressing cells. The frequency of sphere formation was 0.16±0.03%, (N = 3) similar to results obtained with size and pigmentation-based sorts. When P-cadherin levels were subdivided into low and high (Fig. 1G), all colonies came from the P-cadherin-low fraction (sphere formation frequency 0.22±0.15%, N = 3), with an 8.8-fold enrichment in P-cadherin-low over ungated cells (Fig. 1H). Samples from neural retina represented negative controls (Fig. 1I). No colonies arose from the N-cadherin positive cell population. In agreement with pigment-based sorts, we also found that only large sized P-cadherin-expressing cells gave rise to colonies. mRNA of P-cadherin and N-cadherin is high in CE-derived colonies because these colonies represent a mixture of retinal stem and progenitor cells (Fig. 1J). These results demonstrate that low P-cadherin level is a reliable surface marker to specifically isolate RSCs from the CE and also confirm the localization of RSCs to the pigmented epithelium of the CE.
Exogenous factors can direct undifferentiated RSC progeny to produce rod photoreceptors in vitro
When adult RSCs were transplanted into adult mouse retina in a hydrogel matrix, the majority of undifferentiated RSC progeny integrated into the RPE layer (Ballios et al., 2010). However, in order to overcome most primary vision loss, the photoreceptor must be replaced. In order to approach a cellular population ideal for transplantation, a screen of factors shown to be important in rod fate specification during retinal development was performed. These include combinations of taurine (Altshuler et al., 1993), retinoic acid (RA) (Kelley et al., 1994; Hyatt et al., 1996), sodium butyrate and sonic hedgehog (Guillemot and Cepko, 1992; Levine et al., 1997). Strong expression of mature rod photoreceptor phenotypes following application of taurine/RA for 40 days in vitro was described recently (G. C. Demontis, C. Aruta, A. De Marzo, V. Marigo, personal communication).
Rhodopsin expression was used to evaluate the efficiency of rod photoreceptor differentiation. Immunocytochemistry demonstrated stark differences in rod differentiation potential, ranging from 10% in 1% FBS to 95% in the best conditions (supplementary material Fig. S1). The screening results showed a partial increase in photoreceptor fate in the presence of basic fibroblast growth factor (FGF2) alone (40%). FGF2 is known to be an important factor in neural retinal differentiation (Pittack et al., 1997) and rod fate specification during retinogenesis (Guillemot and Cepko, 1992; Merhi-Soussi et al., 2006). Other individual factors induced an intermediate level of rhodopsin expression, approximately 50% in RA, taurine, or SHH alone. The combinations yielding the highest enrichment for rods (90–95%) consistently required a combination of taurine and RA in addition to the other factors (Fig. 2A,B).
Fig. 2. Exogenous factors can induce directed-differentiation of mouse RSC progeny to a rod photoreceptor fate in a pattern consistent with developmental rod production.

(A,B) Representative images of rhodopsin-positive cells (red, cytoplasmic) following 40 days of culture in (A) 1% FBS + F/H, (B) T+RA+F/H. Nuclei are shown with Hoechst staining (blue). Scale bars represent 50 µm. (C) Q-PCR analysis comparing rhodopsin expression between 1% FBS + F/H and T+RA+F/H culture conditions, compared to primary cultures of dissociated adult neural retina and undissociated (fresh) adult neural retina. Data are reported relative to expression in primary undifferentiated RSC spheres. Error bars represent s.e.m. (1% FBS+F/H, N = 4; T+RA+F/H, N = 6; NR culture, N = 2; fresh NR, N = 3). (D–E) Representative electron microscopic images of cells differentiated at 40 days in (D) T+RA+F/H or (E) 1%FBS+F/H. Cells differentiated in T+RA+F/H show a morphology lacking in ciliation and pigmentation (D and inset, black arrowhead), compared to the extensive ciliation and retention of pigment granules seen with cells differentiated in serum. Light grey intracellular regions represent lysosomes and are not pigment granules. Coloured arrowheads have been used to indicate the difference between pigment granules (red arrowheads) and cellular organelles (blue arrowheads) – which include lysosomes and nucleoli. Scale bars represent 10 µm (2 µm inset) (D) or 2 µm (E). (F) RSC-derived rods staining positive for rhodopsin (red) at 28 days differentiation in taurine/retinoic acid show a neuronal morphology (arrows) with long process extending from a central cell body (arrowheads), similar to cultured primary rod photoreceptors from adult mice for 4 days in T+RA. Scale bar represents 50 μm. (G,H) Primary adult rods are positive for (G) rhodopsin (red cytoplasmic) and (H) Crx (purple nuclear). Where these cells form aggregates when plated, the morphology looks similar to aggregates of RSC-derived rod photoreceptors (see Fig. 4). Nuclei are shown with Hoechst staining (blue). Scale bars represent 50 µm. (I) Schematic representation of the differentiation protocol developed for directed-differentiation of rod photoreceptors from primary RSCs isolated from Nrl.gfp animals. (J) Expression profiles of Nrl (green) and rhodopsin (red) over 40 days of differentiation in 1% FBS+F/H, T+RA+F/H, 1% FBS+T+RA+F/H, and the [1% FBS+F/H]/[T+RA+F/H] sequential protocol. Nrl expression precedes rhodopsin expression by approximately 1 week in T+RA, and is consistent regardless of a two week expansion in 1% FBS+F/H. Error bars represent s.e.m. (N>3). (K) Q-PCR analysis comparing genes for multiple retinal cell types between 1% FBS+F/H (40 days), T+RA+F/H (21 days), and T+RA+F/H (40 days). Data are reported relative to expression in primary undifferentiated RSC spheres. Error bars represent the s.e.m. of independent experiments (N = 4–6).
For subsequent experiments, FGF2 was included in control (1% FBS) and taurine/RA conditions as RSC progeny were found to adhere and migrate better on laminin substrate when FGF2 was present. Q-PCR analysis revealed increased expression of rhodopsin in taurine/RA cultures compared to 1% FBS differentiation, similar to levels of rhodopsin in rods from dissociated cultures of neural retinal cells (Fig. 2C).
A recent study used electron microscopy analysis to show that RSC progeny grown in pan-retinal differentiation conditions retain expression of CE morphology, and included the observation that every RSC sphere-derived cell subjected to differentiation in FBS contained at least one pigment granule (Cicero et al., 2009). Electron microscopy was performed on 44 day differentiated RSC progeny in 1%FBS and taurine/retinoic acid. A significant number of cells (66±21%) cultured in taurine/retinoic acid differentiation conditions completely lose pigmentation and adopt morphology inconsistent with retinal epithelial cells in vitro, including loss of ciliation (Fig. 2D,E). Many of the remaining cells included at most one pigment granule. In contrast, 80±20% of cells cultured in 1%FBS retained at least one pigment granule when cultured for 44 days. In low-density culture, RSC-derived rhodopsin-positive cells show a neuronal morphology indistinguishable from primary cultures of adult rod photoreceptors (Fig. 2F–H). The loss of normal photoreceptor morphology in rod photoreceptors isolated from neural retina, dissociated and cultured in vitro is well known (Sparrow et al., 1990) and is hypothesized to be due to the absence of supporting RPE. The lack of clear outer segment morphology in vitro is consistent with our finding that primary cultures of rods express lower levels of rhodopsin, similar to levels observed in RSC-derived rods.
RSC-derived rhodopsin-positive cells develop Nrl and rhodopsin expression in a pattern similar to normal retinal development
Given the marked effect on photoreceptor differentiation efficiency, we sought to understand the mechanism of the taurine/RA effect. Critically, the effect could be one of selective cell survival of a rhodopsin-positive population or a fate change with enhanced proliferation and differentiation of a photoreceptor population.
A time-course of the differentiation was used to address the mechanism (Fig. 2I). Nrl is expressed before rhodopsin in early post-mitotic rods and remains active in mature rods, but is not present in the mature CE or RPE. The Nrl.gfp transgene is expressed specifically in rod photoreceptors but is not confined to the nucleus (Akimoto et al., 2006). RSCs were derived from Nrl.gfp reporter mice and the observation of up-regulated Nrl and rhodopsin expression allowed us to determine exactly when the differentiating RSCs transition to immature and mature rod phenotypes, respectively (Fig. 2J). Q-PCR analysis was performed to investigate the influence of the exogenous factors on pan-retinal gene expression in the differentiating cultures, with expression levels for each transcript compared to its expression in the starting RSC sphere population (Fig. 2K). The presumptive eye field transcription factors Pax6 and Rx are higher at 2–3 weeks but lower in differentiated cultures at 44 days, consistent with an early proliferation of multipotent retinal progenitors. Chx10, a marker of neural retinal progenitors, is decreased in differentiated cultures.
Expression curves were constructed from the immunostaining: these included both % rhodopsin/% Nrl expression (Fig. 2J) and absolute numbers of cells (Fig. 3). Cells always proliferated to a greater extent in the presence of FBS; however, the absolute number of rods was higher in taurine/RA cultures (∼11,000±2,000/well) compared to 1% FBS (∼6,000±1,000/well) despite a significant decrease in total cell number in the taurine/RA cultures (P<0.05, Fig. 3). By using a differentiation protocol in which cells are exposed to FBS for the first 2 weeks and defined exogenous factors for the subsequent 4 weeks (Fig. 2I), the absolute number of rhodopsin-positive cells was increased significantly (∼82,000±18,000/well) (Fig. 3). Whether RSCs begin differentiation in taurine/RA or whether they are treated sequentially with taurine/RA after 2 weeks in 1% FBS, the temporal sequence of Nrl and rhodopsin expression is similar relative to first exposure to taurine/RA (Fig. 2J). Nrl expression precedes rhodopsin expression by approximately 7 days, similar to that observed during retinogenesis (Akimoto et al., 2006).
Fig. 3. Absolute numbers of cells/well over 40 days of RSC differentiation in various culture conditions.

Red bars represent the number of rhodopsin-positive cells present in each condition at each time point. Significantly greater proliferation is observed in media supplemented with 1% FBS than taurine and retinoic acid (* P<0.05); however, the absolute number of rhodopsin-positive cells is greater at 40 days. Error bars represent s.e.m.
Consistent with immunocytochemistry, rhodopsin transcript levels are higher in the cultures differentiated in taurine/RA (Fig. 2K). The specificity of the neural differentiation is supported by the decrease in RPE65 and vimentin expression, markers of mature RPE and Müller glia. It was observed that rhodospin expression appeared in a stochastic, patch-like pattern (Fig. 4), and was one reason that absolute number of rhodopsin-expressing cells in an entire well was quantified for cell counts.
Fig. 4. Nrl and rhodopsin protein expression profiles in differentiating RSCs.

(A) Representative images of differentiating cultures in 1% FBS+F/H and T+RA+F/H conditions at 4, 12, 21, and 40 days. In T+RA conditions, 12 days corresponds to high Nrl and lower rhodopsin expression, while at 21 days both Nrl and rhodopsin expression are high. By 40 days, cytoplasmic spreading results from further differentiation and the GFP is more widespread in the cultures. The images show overlap of Nrl (gfp, green) and rhodopsin (red) expression in the cell cytoplasm, including a number of Nrl+/rho+ and Nrl+/rho− cells. Nuclei are shown with Hoechst staining (blue). Scale bars represent 100 µm (4d) or 50 µm (12d, 21d, 40d). (B,C) Similar morphology is exhibited at the single cell level in low-density culture, between (B) a mature RSC-derived rhodopsin-positive cell (28 days) and (C) a cultured primary adult rod photoreceptor (3 days in vitro). The cytoplasmic morphology is visualized with ACTN.GFP (B) and Nrl.gfp (C). Scale bars represent 50 µm.
The up-regulation of mature cell type markers as assessed by mRNA levels was lower than would be expected when compared with the differences in protein expression assayed by immunocytochemistry. This is likely due to our finding that, before any differentiation period, primary sphere colonies expressed mRNA for most differentiated cell markers. It is known that stem cells can express lineage-specific markers prior to becoming committed to a particular cell fate (Hu et al., 1997). RSC spheres are a mixture of stem and more mature progenitors which are likely already expressing some neural and photoreceptor mRNAs, similar to what has been observed in neural stem cell sphere colonies (Kukekov et al., 1999; Sievertzon et al., 2005).
Taurine/retinoic acid induces a fate change and not a selective survival effect on RSC-derived rhodopsin-positive cells
The presence of taurine/RA in addition to 1% FBS (1% FBS+T+RA+F/H) does not effect the growth rate compared to 1% FBS alone (Fig. 3). However, an early progenitor expansion using 2 weeks in 1% FBS followed by 4 weeks in taurine/RA ([1% FBS+F/H]/[T+RA+F/H]) leads to higher absolute numbers of rhodopsin-positive cells at 40 days (P<0.05, Fig. 3). This sequential treatment shows a similar proliferative rate to 1% FBS alone demonstrating that FBS has no impact on progenitor proliferation beyond the first 2 weeks in culture (Fig. 3). With no impact on cell growth or the relative profiles of Nrl and rhodopsin expression, a survival effect on post-mitotic rods is unlikely. The steadily increasing expression of mature photoreceptor markers supports a fate change hypothesis for the effect of taurine/RA on RSC differentiation (see Discussion). The loss of proliferation in these differentiating cultures after the progenitor expansion phase is supported by a drop in cells detectable in cycle (ki-67 staining).
Rod photoreceptor differentiation from adult human RSC progeny
Employing a similar differentiation strategy, adult human RSC progeny could be directed to produce rhodopsin-positive cells with high yield, over a protracted differentiation time course relative to mouse RSC progeny (Fig. 5A). These human RSC-derived rods expressed rhodopsin and adopted a neuronal morphology consistent with rod photoreceptors in vitro (Fig. 5B,C). Specifically, they expressed protein markers characteristic of mature photoreceptors (CRX) and not markers of other retinal lineages, such as mature RPE cells (RPE65), Müller glia (CRALBP) or retinal interneurons (calbindin) (data not shown).
Fig. 5. Exogenous factors can induce directed-differentiation of human RSC progeny to a rod photoreceptor fate.

(A) Expression profiles of rhodopsin (red) in human RSC progeny over 150 days of differentiation in T+RA+F/H, 1% FBS+F/H. Error bars represent s.e.m. (N = 2–3). (B,C) Representative images of rod photoreceptor progeny arising in differentiating cultures of human RSC progeny at 105 and 123 days of differentiation in T+RA+F/H. Note the mature polarized neuronal morphology characteristic of mature rods adopted by isolated RSC-derived rods (arrowhead). Nuclei are shown with Hoechst staining (blue). Scale bars represent 100 µm.
RSC-derived rhodopsin-positive cells express additional mature photoreceptor markers, and do not express markers of other retinal lineages
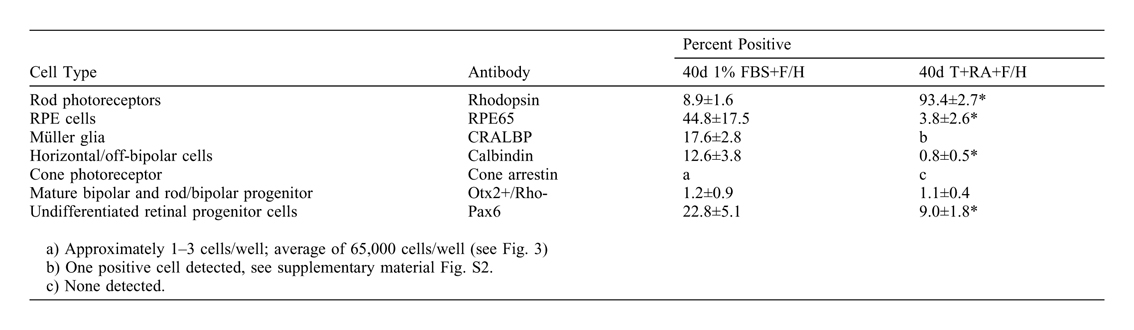
In order to unambiguously confirm the identity of the differentiated cells as post-mitotic rod photoreceptors, staining was performed for other markers of the retinal lineage at 6 weeks of differentiation (supplementary material Fig. S2). Markers of mature RPE cells (RPE65+), Müller glia (CRALBP+), mature bipolar cells or immature rod/bipolar progenitors (Otx2+Rho-) (Nishida et al., 2003; Fossat et al., 2007), and horizontal cells (calbindin+) were present in prominent levels in 1% FBS cultures (supplementary material Table S1), consistent with previous reports of the differentiation potential of RSCs (Tropepe et al., 2000; Coles et al., 2004). The expression of these markers was reduced significantly in taurine/RA cultures at 40 days. The undifferentiated RSC marker Pax6 (Xu et al., 2007) showed significantly higher expression at 21 days (48.9±6.9%, P<0.05), consistent with an expansion of progenitor cells prior to fate specification. This level dropped significantly by 40 days in taurine/RA culture. The predominant remaining differentiated cells at 40 days in the taurine/RA culture (i.e. rhodopsin-negative cells) were RPE65-positive cells (3.8±2.6%) (supplementary material Table S1, Fig. S2G). Notably, while cone-rod homeobox (Crx) transcript levels were not significantly different at 21 and 40 days of differentiation (Fig. 2K), the specificity of the rhodopsin immunostaining for detecting mature post-mitotic rod photoreceptors was confirmed by Crx/rhodospin double-labeling (supplementary material Fig. S2E,K). Crx exhibits both nuclear and peri-nuclear staining consistent with previous reports of staining patterns in mature retinal cells in vitro (Meyer et al., 2009), and characteristic of other photoreceptor markers that display a strong nuclear signal in the developing cell and become progressively diffuse and peri-nuclear with differentiation, such as Otx2 or Nr2e3 (Baas et al., 2000; Peng et al., 2005). Cone markers, such as cone arrestin (supplementary material Fig. S2J), blue- and red/green-opsin could not be found by immunocytochemistry in 40 day taurine/RA culture. These data indicate that the progeny of adult RSCs, clonally isolated from the CE can undergo rod photoreceptor differentiation.
Discussion
The RSC was identified as a rare pigmented cell in the CE of adult mouse (Tropepe et al., 2000) and human (Coles et al., 2004) eye, with the ability to self-renew through long-term passaging and the capacity to differentiate into all retinal cell types. We find that it is only a defined subset of medium-large, heavily pigmented cells that exhibits stem cell properties, in contrast to a model in which colony formation is a general property of cultured CE or adjacent RPE cells (Cicero et al., 2009). P-cadherin is expressed early in eye development as the outer optic vesicle becomes specified towards RPE and continues to be expressed in adult pigmented CE and RPE (Xu et al., 2002; Bharti et al., 2006). We show that P-cadherin is also expressed at low levels in adult RSCs and that RSCs have at least two properties in common with nonproliferative RPE cells: the presence of at least some level of pigmentation and P-cadherin expression. This resemblance of a tissue-specific stem cell to a fully differentiated cell is similar to the case in the mammalian brain, where adult neural stem cells closely resemble mature astroglial cells (Doetsch et al., 1999; Seri et al., 2001).
Analogy has been made between the pigmented CE and the ciliary marginal zone of lower vertebrates. In cold-blooded vertebrates (e.g. teleosts/zebrafish) the retina continues to grow throughout adult life and in response to injury by the proliferative addition of new neurons from a germinal zone at the ciliary margin (Otteson and Hitchcock, 2003). A small population of proliferating retinal progenitors in the marginal zone in post-natal chickens also has been identified (Fischer and Reh, 2000).
Recent reports have suggested alternate hypotheses to the observation of RSC stem cell character in terms of colony isolation, expansion and differentiation in vitro. These hypotheses include: the equal ability of all CE cells to undergo in vitro clonal expansion and transdifferentiation to produce mature retinal cell types; and unconvincing photoreceptor differentiation due to an epithelial character (Cicero et al., 2009; Gualdoni et al., 2010). Transdifferentiation is defined as the conversion of one differentiated, non-stem, cell type directly to another differentiated cell type and has been demonstrated in a number of amphibians as well as embryonic chick and rat (Guillemot and Cepko, 1992; Park and Hollenberg, 1993; Opas and Dziak, 1994; Pittack et al., 1997). As reported above, enrichment of a clonally proliferative and multipotent stem cell based on the criteria of size and pigmentation suggests that transdifferentiation is an unlikely mechanism to describe these results. Additionally, in Chx10-null and Mitf-null mice with reduced neural retinal and RPE progenitor populations, respectively, a 3–8 fold increase of RSCs was observed (Coles et al., 2006). This is the opposite of what would be predicted by a CE-transdifferentiation model: in the Mitf-mutant, fewer pigmented epithelial cells should be available for transdifferentiation into RSCs.
Inefficient photoreceptor differentiation was reported in studies that exposed cultures to retinal differentiation conditions for 2–3 weeks (Cicero et al., 2009; Gualdoni et al., 2010). Our own studies corroborate that similar differentiation in 1% FBS results in low expression of rod photoreceptor markers (this study and Tropepe et al., 2000), but our data suggests that longer-term retinal differentiation is required to acquire phenotypic maturity of retinal cell types, and that the first 14 days of culture consist primarily of progenitor expansion. In addition, it is possible that a monolayer culturing method with multiple passages into serum could promote the selective growth of Müller cells, as expression of Müller glial markers Cx43 and vimentin (Kihara et al., 2006; Zahs and Ceelen, 2006) were up-regulated and we find that Müller glial cells are one of the most common cell types to differentiate when RSCs are exposed to serum (supplementary material Table S1). Other markers used in the studies by Gualdoni et al. and Cicero et al. to mark an epithelial identity include palmdelphin – expressed in the mature neural retinal (Gualdoni et al., 2010) – and ZO-1, which persists in the apical cell membranes of undifferentiated neural tube neuroepithelium (Aaku-Saraste et al., 1996). The presence of ZO-1 indicates that the cells are neuroepithelial-like and undifferentiated, and supports a model of RSCs as a multipotent cell type.
Our results for Nrl and rhodopsin expression profiles in vitro (Fig. 2J) using Nrl.gfp reporter mice, suggest that the ontogenic stages of rod photoreceptor gene expression in vivo (Akimoto et al., 2006) can be recapitulated in differentiating RSCs. The conservation of the identical temporal sequence of Nrl/rhodopsin expression regardless of the presence of a two-week expansion of retinal precursors from RSCs in 1%FBS+F/H ([1%FBS+F/H]/[T+RA+F/H] sequence, Fig. 2I,J) supports a fate change hypothesis, but does not in itself rule out a selective photoreceptor survival effect. Our data show steadily increasing rhodopsin-positive cell counts over 44 days of culture (Fig. 3). Combined with the similar proliferative rates between [1%FBS+F/H]/[T+RA+F/H] and 1%FBS alone, this strongly argues for a fate change effect of taurine/RA. A two-week progenitor expansion phase is sufficient to expand the population without limiting photoreceptor differentiation potential. The ability to expand the RSC-derived progenitor pool prior to differentiation is advantageous in increasing the number of cells available for transplantation to clinically relevant levels (Daley and Scadden, 2008).
Taurine/RA culture was found to have a similar effect on adult human RSC differentiation, though the time course was considerably protracted. This is consistent with observations of photoreceptor differentiation in hES cell-derived retinal progenitors. An onset of opsin expression at day 70 of differentiation has been reported (Meyer et al., 2009), and a second group detected no opsin expression after 130 days of differentiation and observed that less than 10% of cells were rhodopsin-positive at day 200 (Osakada et al., 2008). Our new differentiation protocol produces more than 90% rhodospin-positive cells after 120 days.
We cannot rule out the possibility of taurine and RA encouraging the survival of an early photoreceptor specific progenitor (i.e. one which does not express Nrl or rhodospin). While the existence of a proliferative photoreceptor specific progenitor has been denied (Turner and Cepko, 1987; Turner et al., 1990), there remains controversy in the literature (Williams and Goldowitz, 1992; Cayouette et al., 2006), and no specific marker exists for such a cell. Clonal lineage analysis of the differentiating progenitors is needed to clarify the exact mechanisms governing the directed-differentiation.
Based on our knowledge of photoreceptor development, we can postulate a mechanism of action of the taurine/RA factors in photoreceptor fate specification. Otx2/Crx/NeuroD1-expressing photoreceptors develop as rods or cones depending on the expression of Nrl (Swaroop et al., 1992). Nrl binds and directly activates the rhodopsin promoter and is required for rod photoreceptor genesis (Mears et al., 2001): deletion of Nrl results in rods developing as cones (Daniele et al., 2005). RA promotes Nrl expression directly through a RA response element (RARE) in the Nrl promoter (Khanna et al., 2006). Taurine acts through the glycine receptor (Young and Cepko, 2004) on post-mitotic cells, altering opsin expression (Wallace and Jensen, 1999). The differentiation of retinal progenitors requires both exogenous signals, as well as cell-intrinsic restriction of potency (Adler and Hatlee, 1989; Reh and Kijavin, 1989; Cepko et al., 1996). Our results confirm the importance of exogenous (cell non-autonomous) signals in directing the fate of retinal progenitors, as well as supporting phenotypic maturation.
The directed-differentiation of an adult stem cell population toward a therapeutically relevant cell type effectively (high purity) and efficiently (high yield) is a major barrier to regenerative medicine. The adult RSC shows distinct advantages as a donor cell population. The use of exogenous factors to drive photoreceptor differentiation in RSC progeny avoids genomic manipulation by viral vectors, which eliminates the risk of insertional mutagenesis (Halme and Kessler, 2006). Employing fetal retinal progenitors for transplantation (Klassen et al., 2004) requires isolation of these cells from the developing neural retina and this raises significant ethical issues. A number of groups have developed protocols to differentiate retinal cells types from human ES cells (Banin et al., 2006; Lamba et al., 2006; Osakada et al., 2008; Meyer et al., 2009) and iPS cells (Lamba et al., 2010). Without going through a definitive RSC stage, these methods cannot exclude non-retinal cell types.
Our study clarifies the characteristics of the adult RSC as a rare pigmented cell in the CE that can be enriched based on size, pigmentation, and low P-cadherin expression. Directing photoreceptor differentiation from the adult RSC is in agreement with current models of retinogenesis, where the ability to express markers of a mature rod phenotype are dependent on exogenous factors (Altshuler et al., 1993; Kelley et al., 1994). These conclusions elucidate novel tissue-specific stem cell characteristics of the adult RSC, and the ability to enrich for rod specific lineages from human RSCs may prove useful for replacement of photoreceptors damaged by retinal disease.
Materials and Methods
Mouse Strains
C57BL/6, Actin.gfp, and Nrl.gfp (Akimoto et al., 2006) transgenic strains were used in this study (of either sex). The Nrl.gfp animal expresses GFP under the Nrl promoter, resulting in cytoplasmic fluorescence in Nrl-positive cells. Experimental procedures were performed in accordance with the Guide to the Care and Use of Experimental Animals and approved by the Animal Care Committee at the University of Toronto.
Primary Cell Isolation and Culture
RSCs were derived from the CE of adult mice (minimum 6 weeks old) (Coles et al., 2006) and from adult human donor tissue (of either sex) (Coles et al., 2004) as described previously. Human eyes were procured from the Eye Bank of Canada (Toronto, ON) within 24 h post-mortem. Cells were plated in serum free media (SFM) on non-adherent tissue culture plates (Nunc; Thermo Fisher Scientific) at a density of 20 cells/µL.
Differentiation of RSC Spheres
Individual RSC spheres were selected after 7 days primary culture. Spheres were derived from Actin.gfp mice to confirm appropriate sub-cellular localization of protein products on immunofluorescence (cytoplasmic v. nuclear), or from Nrl.gfp mice. Spheres (2/well) were plated on laminin (50 ng/mL)-coated 24-well plates (Nunc). Various time courses of differentiation are illustrated in Fig. 2I. Following four days of culture in SFM plus FGF2 (10 ng/mL, human recombinant; Sigma) and heparin (2 ng/mL; Sigma) to encourage sphere adhesion and spreading, the media is replaced with the following (refreshed every four days):
Standard differentiation media: SFM plus 1% FBS (Invitrogen).
Retinal differentiation media: SFM plus combinations of taurine (100 µM; Sigma), RA (500 nM; Sigma), FGF2 (10 ng/mL).
Flow Cytometry and Cell Labeling
Dissociated primary CE dissections cells were sorted using a FacsAria (BD Biosciences). For pigmentation sorts, cells were sorted based on forward and side scatter without the use of surface markers. For detection of P-cadherin, a FITC conjugated primary antibody (N-19, 1:200, Santa Cruz Biotechnology) or rat anti P-cadherin (1:200, Zymed Laboratories) in combination with Alexa anti-rat 488 (1:400) was used. Cells were counterstained with propidium iodide (0.9 µg/µL, Invitrogen) to assess viability. Analysis was performed using BD FacsDiva Software V6.1.2.
When defining a pigmented population from the CE for sorting, a population of cells with high side scatter (SSC) was chosen by comparison to control neural retina samples (which do not contain pigmented cells). This sort was displayed by plotting against a second, empty channel (FITC) as is standard practice. Cells exhibit some autofluorescence in the FITC spectrum, but were not specifically stained with any fluorescent-tagged antibodies, or sorted based on this. For sorting cells from the CE based on forward and side scatter, four approximately equal sized gates were drawn guided by clusters within the population.
High and low gates for P-cadherin were defined by cell clusters observed in fluorescence values in stained cells, as is standard procedure. Gate widths were kept constant between experiments. The lower border for “low” P-cadherin expression was defined by placing the gate just outside the peak fluorescence value for negative control (P-cadherin stained neural retinal cells) in each sort. Gate width (fluorescence intensity) for low P-cadherin expression was smaller, as the density of cell was higher for low fluorescence intensity values. Low P-cadherin expressing cells composed 20–40% of the total cells. High P-cadherin expressing cells composed the top 4–10% of total cells.
Quantitative RT Polymerase Chain Reaction (Q-PCR)
Real time RT-PCR experiments were performed as previously described (Xu et al., 2007). All Q-PCR is reported relative to transcript levels of a particular gene in undifferentiated RSC progeny (i.e. RSC spheres) prior to differentiation. RNA was isolated using a Qiagen RNeasy extraction kit with DNase to remove genomic DNA contamination and a specified amount of cDNA was reverse transcribed using SuperscriptIII (Invitrogen). Q-PCR was performed using Taqman Gene Expression Assays in a 7900HT Fast Real-Time PCR System (both from Applied Biosystems). The following Taqman assays (Applied Biosystems) were used: rax 01258704_m1, pax6 00443081_m1, chx10 00432549_m1, rho 00520345_m1, crx 00483994_m1, vim 01333430_m1, calbindin 00801461_m1, opsin1mw 00433560, rpe65 00504133_m1, cdh3 (pcdh) 01249215_m1, cdh2 (ncdh) 00483213_m1, rplp0 00725448_s1. Quantification was performed using the delta Ct method with rplp0 ribosomal protein as endogenous control.
Immunostaining
Immunostaining was performed as previously described (Coles et al., 2004). Immunofluorescence was reviewed by epifluorescence (Zeiss AxioObserver.D1), as well as confocal image analysis (Olympus Fluoview FV1000) to confirm co-localization of indicated protein products. Cell counts were performed using ImageJ image analysis software. The following antibodies were used: mouse anti rhodopsin (RetP1, 1:250; Millipore), mouse anti RPE65 (1:250; Millipore), mouse anti ki-67 (1:10; BD Pharmigen), rabbit anti blue opsin (1:200; Millipore), rabbit anti red/green opsin (1:200; Millipore), mouse anti Pax6 (1:400; Developmental Studies Hybridoma Bank, Iowa City, IA), mouse anti Cralbp (1:500; Abcam, Cambridge, MA), mouse anti calbindin (1:500; Sigma), rabbit anti GFP (1:500; Molecular Probes, Invitrogen), rabbit anti cone arrestin (1:10,000; a gift of Cheryl Craft, University of California, Los Angeles) (Zhu et al., 2003), rabbit anti Otx2/Crx (1:1000; Cheryl Craft) (Zhu and Craft, 2000). Alexa-tagged secondary antibodies (Molecular Probes, Invitrogen) were used at 1:400. All antibodies were tested on sections of neural retina/RPE as positive controls.
Statistics
All cell counts and pooled data are presented as averages with standard error of the mean (s.e.m.). Statistics were performed using Prism 4. Significance is noted using Student's t test to compare two groups or ANOVA when comparing three or more groups. Dunnett's post-hoc test was used to compare multiple groups to a single control group, and Bonferroni's post-hoc test to compare multiple groups to each other.
Supplementary Material
Acknowledgments
We thank Vince Tropepe and members of the Shoichet and van der Kooy groups for helpful discussion. Anand Swaroop kindly provided the Nrl.gfp mouse strain. We thank Pier-Andree Penttila for her assistance with cell sorting. We thank Cheryl Craft at the Mary D. Allen Laboratory for Vision Research, Doheny Eye Institute for sharing antibodies. The anti Pax6 antibody developed by A. Kawakami was obtained from the Developmental Studies Hybridoma Bank developed under the auspices of the NICHD and maintained by The University of Iowa, Department of Biology, Iowa City, IA 52242. BGB and LC are supported by a Canadian Institutes of Health Research (CIHR) MD/PhD studentship. BGB also holds a McLaughlin Centre Graduate Fellowship, a University of Toronto Open Fellowship and a McLaughlin Centre for Molecular Medicine award. This work was supported by the CIHR, NIH (R01 EY015716) (DvdK) and the Foundation Fighting Blindness Canada. BGB, LC and BLKC were involved with collection and assembly of data. All authors were involved with conception and design, and manuscript writing.
Footnotes
Competing interests: None to declare.
References
- Aaku-Saraste E., Hellwig A., Huttner W. B. (1996). Loss of occludin and functional tight junctions, but not ZO-1, during neural tube closure-Remodeling of the neuroepithelium prior to neurogenesis. Dev. Biol. 180, 664–679 10.1006/dbio.1996.0336 [DOI] [PubMed] [Google Scholar]
- Adler R., Hatlee M. (1989). Plasticity and differentiation of embryonic retinal cells after terminal mitosis. Science 243, 391–393 10.1126/science.2911751 [DOI] [PubMed] [Google Scholar]
- Akimoto M., Cheng H., Zhu D., Brzezinski J. A., Khanna R., Filippova E., Oh E. C. T., Jing Y., Linares J.-L., Brooks M. et al. (2006). Targeting of GFP to newborn rods by Nrl promoter and temporal expression profiling of flow-sorted photoreceptors. Proc. Natl. Acad. Sci. USA 103, 3890–3895 10.1073/pnas.0508214103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Altshuler D., Lo, Turco J. J., Rush J., Cepko C. (1993). Taurine promotes the differentiation of a vertebrate retinal cell type in vitro. Development 119, 1317–1328 [DOI] [PubMed] [Google Scholar]
- Baas D., Bumsted K. M., Martinez J. A., Vaccarino F. M., Wikler K. C., Barnstable C. J. (2000). The subcellular localization of Otx2 is cell-type specific and developmentally regulated in the mouse retina. Brain Res. Mol. Brain Res. 78, 26–37 10.1016/S0169-328X(00)00060-7 [DOI] [PubMed] [Google Scholar]
- Ballios B. G., Cooke M. J., van der Kooy D., Shoichet M. S. (2010). A hydrogel-based stem cell delivery system to treat retinal degenerative diseases. Biomaterials 31, 2555–2564 10.1016/j.biomaterials.2009.12.004 [DOI] [PubMed] [Google Scholar]
- Banin E., Obolensky A., Idelson M., Hemo I., Reinhardtz E., Pikarsky E., Ben-Hur T., Reubinoff B. (2006). Retinal incorporation and differentiation of neural precursors derived from human embryonic stem cells. Stem Cells 24, 246–257 10.1634/stemcells.2005-0009 [DOI] [PubMed] [Google Scholar]
- Bharti K., Nguyen M. T., Skuntz S., Bertuzzi S., Arnheiter H. (2006). The other pigment cell: specification and development of the pigmented epithelium of the vertebrate eye. Pigment Cell Res. 19, 380–394 10.1111/j.1600-0749.2006.00318.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cayouette M., Poggi L., Harris W. A. (2006). Lineage in the vertebrate retina. Trends Neurosci. 29, 563–570 10.1016/j.tins.2006.08.003 [DOI] [PubMed] [Google Scholar]
- Cepko C. L., Austin C. P., Yang X., Alexiades M., Ezzeddine D. (1996). Cell fate determination in the vertebrate retina. Proc. Natl. Acad. Sci. USA 93, 589–595 10.1073/pnas.93.2.589 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cicero S. A., Johnson D., Reyntjens S., Frase S., Connell S., Chow L. M. L., Baker S. J., Sorrentino B. P., Dyer M. A. (2009). Cells previously identified as retinal stem cells are pigmented ciliary epithelial cells. Proc. Natl. Acad. Sci. USA 106, 6685–6690 10.1073/pnas.0901596106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coles B. L., Horsford D. J., McInnes R. R., van der Kooy D. (2006). Loss of retinal progenitor cells leads to an increase in the retinal stem cell population in vivo. Eur. J. Neurosci. 23, 75–82 10.1111/j.1460-9568.2005.04537.x [DOI] [PubMed] [Google Scholar]
- Coles B. L. K., Angenieux B., Inoue T., Del R.-T. K., Spence J. R., McInnes R. R., Arsenijevic Y., van der Kooy D. (2004). Facile isolation and the characterization of human retinal stem cells. Proc. Natl. Acad. Sci. USA 101, 15772–15777 10.1073/pnas.0401596101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daley G. Q., Scadden D. T. (2008). Prospects for stem cell-based therapy. Cell 132, 544–548 10.1016/j.cell.2008.02.009 [DOI] [PubMed] [Google Scholar]
- Daniele L. L., Lillo C., Lyubarsky A. L., Nikonov S. S., Philp N., Mears A. J., Swaroop A., Williams D. S., Pugh E. N. J. (2005). Cone-like morphological, molecular, and electrophysiological features of the photoreceptors of the Nrl knockout mouse. Invest. Ophthalmol. Vis. Sci. 46, 2156–2167 10.1167/iovs.04-1427 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doetsch F., Caille I., Lim D. A., Garcia-Verdugo J. M., Alvarez-Buylla A. (1999). Subventricular zone astrocytes are neural stem cells in the adult mammalian brain. Cell 97, 703–716 10.1016/S0092-8674(00)80783-7 [DOI] [PubMed] [Google Scholar]
- Fischer A. J., Reh T. A. (2000). Identification of a proliferating marginal zone of retinal progenitors in postnatal chickens. Dev. Biol. 220, 197–210 10.1006/dbio.2000.9640 [DOI] [PubMed] [Google Scholar]
- Fossat N., Le Greneur C., Beby F., Vincent S., Godement P., Chatelain G., Lamonerie T. (2007). A new GFP-tagged line reveals unexpected Otx2 protein localization in retinal photoreceptors. BMC Dev. Biol. 7, 122 10.1186/1471-213X-7-122 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gualdoni S., Baron M., Lakowski J., Decembrini S., Smith A. J., Pearson R. A., Ali R. R., Sowden J. C. (2010). Adult ciliary epithelial cells, previously identified as retinal stem cells with potential for retinal repair, fail to differentiate into new rod photoreceptors. Stem Cells 28, 1048–1059 10.1002/stem.423 [DOI] [PubMed] [Google Scholar]
- Guillemot F., Cepko C. L. (1992). Retinal fate and ganglion cell differentiation are potentiated by acidic FGF in an in vitro assay of early retinal development. Development 114, 743–754 [DOI] [PubMed] [Google Scholar]
- Halme D. G., Kessler D. A. (2006). FDA regulation of stem-cell-based therapies. N. Engl. J. Med. 355, 1730–1735 10.1056/NEJMhpr063086 [DOI] [PubMed] [Google Scholar]
- Hu M., Krause D., Greaves M., Sharkis S., Dexter M., Heyworth C., Enver T. (1997). Multilineage gene expression precedes commitment in the hemopoietic system. Genes Dev. 11, 774–785 10.1101/gad.11.6.774 [DOI] [PubMed] [Google Scholar]
- Hyatt G. A., Schmitt E. A., Fadool J. M., Dowling J. E. (1996). Retinoic acid alters photoreceptor development in vivo. Proc. Natl. Acad. Sci. USA 93, 13298–13303 10.1073/pnas.93.23.13298 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inagaki M., Irie K., Ishizaki H., Tanaka-Okamoto M., Morimoto K., Inoue E., Ohtsuka T., Miyoshi J., Takai Y. (2005). Roles of cell-adhesion molecules nectin 1 and nectin 3 in ciliary body development. Development 132, 1525–1537 10.1242/dev.01697 [DOI] [PubMed] [Google Scholar]
- Inoue T., Coles B. L., Dorval K., Bremner R., Bessho Y., Kageyama R., Hino S., Matsuoka M., Craft C. M., McInnes R. R. et al. (2010). Maximizing functional photoreceptor differentiation from adult human retinal stem cells. Stem Cells 28, 489–500 10.1002/stem.279 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelley M. W., Turner J. K., Reh T. A. (1994). Retinoic acid promotes differentiation of photoreceptors in vitro. Development 120, 2091–2102 [DOI] [PubMed] [Google Scholar]
- Khanna H., Akimoto M., Siffroi-Fernandez S., Friedman J. S., Hicks D., Swaroop A. (2006). Retinoic acid regulates the expression of photoreceptor transcription factor NRL. J. Biol. Chem. 281, 27327–27334 10.1074/jbc.M605500200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kihara A. H., Mantovani D. C. L., Belmonte M. A., Yan C. Y. I., Moriscot A. S., Hamassaki D. E. (2006). Expression of connexins 36, 43, and 45 during postnatal development of the mouse retina. J. Neurobiol. 66, 1397–1410 10.1002/neu.20299 [DOI] [PubMed] [Google Scholar]
- Klassen H. J., Ng T. F., Kurimoto Y., Kirov I., Shatos M., Coffey P., Young M. J. (2004). Multipotent retinal progenitors express developmental markers, differentiate into retinal neurons, and preserve light-mediated behavior. Invest. Ophthalmol. Vis. Sci. 45, 4167–4173 10.1167/iovs.04-0511 [DOI] [PubMed] [Google Scholar]
- Kukekov V. G., Laywell E. D., Suslov O., Davies K., Scheffler B., Thomas L. B., O'Brien T. F., Kusakabe M., Steindler D. A. (1999). Multipotent stem/progenitor cells with similar properties arise from two neurogenic regions of adult human brain. Exp. Neurol. 156, 333–344 10.1006/exnr.1999.7028 [DOI] [PubMed] [Google Scholar]
- Lamba D. A., Karl M. O., Ware C. B., Reh T. A. (2006). Efficient generation of retinal progenitor cells from human embryonic stem cells. Proc. Natl. Acad. Sci. USA 103, 12769–12774 10.1073/pnas.0601990103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lamba D. A., McUsic A., Hirata R. K., Wang P. R., Russell D., Reh T. A. (2010). Generation, purification and transplantation of photoreceptors derived from human induced pluripotent stem cells. PLoS ONE 5, e8763 10.1371/journal.pone.0008763 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levine E. M., Roelink H., Turner J., Reh T. A. (1997). Sonic hedgehog promotes rod photoreceptor differentiation in mammalian retinal cells in vitro. J. Neurosci. 17, 6277–6288 [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacLaren R. E., Pearson R. A., MacNeil A., Douglas R. H., Salt T. E., Akimoto M., Swaroop A., Sowden J. C., Ali R. R. (2006). Retinal repair by transplantation of photoreceptor precursors. Nature 444, 203–207 10.1038/nature05161 [DOI] [PubMed] [Google Scholar]
- Mears A. J., Kondo M., Swain P. K., Takada Y., Bush R. A., Saunders T. L., Sieving P. A., Swaroop A. (2001). Nrl is required for rod photoreceptor development. Nat. Genet. 29, 447–452 10.1038/ng774 [DOI] [PubMed] [Google Scholar]
- Merhi-Soussi F., Angenieux B., Canola K., Kostic C., Tekaya M., Hornfeld D., Arsenijevic Y. (2006). High yield of cells committed to the photoreceptor fate from expanded mouse retinal stem cells. Stem Cells 24, 2060–2070 10.1634/stemcells.2005-0311 [DOI] [PubMed] [Google Scholar]
- Meyer J. S., Shearer R. L., Capowski E. E., Wright L. S., Wallace K. A., McMillan E. L., Zhang S. C., Gamm D. M. (2009). Modeling early retinal development with human embryonic and induced pluripotent stem cells. Proc. Natl. Acad. Sci. USA 106, 16,698–16,703 10.1073/pnas.0905245106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishida A., Furukawa A., Koike C., Tano Y., Aizawa S., Matsuo I., Furukawa T. (2003). Otx2 homeobox gene controls retinal photoreceptor cell fate and pineal gland development. Nat. Neurosci. 6, 1255–1263 10.1038/nn1155 [DOI] [PubMed] [Google Scholar]
- Opas M., Dziak E. (1994). bFGF-induced transdifferentiation of RPE to neuronal progenitors is regulated by the mechanical properties of the substratum. Dev. Biol. 161, 440–454 10.1006/dbio.1994.1043 [DOI] [PubMed] [Google Scholar]
- Osakada F., Ikeda H., Mandai M., Wataya T., Watanabe K., Yoshimura N., Akaike A., Sasai Y., Takahashi M. (2008). Toward the generation of rod and cone photoreceptors from mouse, monkey and human embryonic stem cells. Nat. Biotechnol. 26, 215–224 10.1038/nbt1384 [DOI] [PubMed] [Google Scholar]
- Otteson D. C., Hitchcock P. F. (2003). Stem cells in the teleost retina: persistent neurogenesis and injury-induced regeneration. Vision Res. 43, 927–936 10.1016/S0042-6989(02)00400-5 [DOI] [PubMed] [Google Scholar]
- Park C. M., Hollenberg M. J. (1993). Growth factor-induced retinal regeneration in vivo. Int. Rev. Cytol. 146, 49–74 10.1016/S0074-7696(08)60379-4 [DOI] [PubMed] [Google Scholar]
- Peng G. H., Ahmad O., Ahmad F., Liu J., Chen S. (2005). The photoreceptor-specific nuclear receptor Nr2e3 interacts with Crx and exerts opposing effects on the transcription of rod versus cone genes. Hum. Mol. Genet. 14, 747–764 10.1093/hmg/ddi070 [DOI] [PubMed] [Google Scholar]
- Pittack C., Grunwald G. B., Reh T. A. (1997). Fibroblast growth factors are necessary for neural retina but not pigmented epithelium differentiation in chick embryos. Development 124, 805–816 [DOI] [PubMed] [Google Scholar]
- Reh T. A., Kijavin I. J. (1989). Age of differentiation determines rat retinal germinal cell phenotype: induction of differentiation by dissociation. J. Neurosci. 9, 4179–4189 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seri B., Garcia-Verdugo J. M., McEwen B. S., Alvarez-Buylla A. (2001). Astrocytes give rise to new neurons in the adult mammalian hippocampus. J. Neurosci. 21, 7153–7160 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sievertzon M., Wirta V., Mercer A., Meletis K., Erlandsson R., Wikstrom L., Frisen J., Lundeberg J. (2005). Transcriptome analysis in primary neural stem cells using a tag cDNA amplification method. BMC Neurosci. 6, 28 10.1186/1471-2202-6-28 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sparrow J. R., Hicks D., Barnstable C. J. (1990). Cell commitment and differentiation in explants of embryonic rat neural retina. Comparison with the developmental potential of dissociated retina. Brain Res. Dev. Brain Res. 51, 69–84 10.1016/0165-3806(90)90259-2 [DOI] [PubMed] [Google Scholar]
- Swaroop A., Xu J. Z., Pawar H., Jackson A., Skolnick C., Agarwal N. (1992). A conserved retina-specific gene encodes a basic motif/leucine zipper domain. Proc. Natl. Acad. Sci. USA 89, 266–270 10.1073/pnas.89.1.266 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tropepe V., Coles B. L. K., Chiasson B. J., Horsford D. J., Elia A. J., McInnes R. R., van der Kooy D. (2000). Retinal stem cells in the adult mammalian eye. Science 287, 2032–2036 10.1126/science.287.5460.2032 [DOI] [PubMed] [Google Scholar]
- Turner D. L., Cepko C. L. (1987). A common progenitor for neurons and glia persists in rat retina late in development. Nature 328, 131–136 10.1038/328131a0 [DOI] [PubMed] [Google Scholar]
- Turner D. L., Snyder E. Y., Cepko C. L. (1990). Lineage-independent determination of cell type in the embryonic mouse retina. Neuron 4, 833–845 10.1016/0896-6273(90)90136-4 [DOI] [PubMed] [Google Scholar]
- Wallace V. A., Jensen A. M. (1999). IBMX, taurine and 9-cis retinoic acid all act to accelerate rhodopsin expression in postmitotic cells. Exp. Eye Res. 69, 617–627 10.1006/exer.1999.0741 [DOI] [PubMed] [Google Scholar]
- Williams R. W., Goldowitz D. (1992). Lineage versus environment in embryonic retina: a revisionist perspective. Trends Neurosci. 15, 368–373 10.1016/0166-2236(92)90181-7 [DOI] [PubMed] [Google Scholar]
- Xu L., Overbeek P. A., Reneker L. W. (2002). Systematic analysis of E-, N- and P-cadherin expression in mouse eye development. Exp. Eye Res. 74, 753–760 10.1006/exer.2002.1175 [DOI] [PubMed] [Google Scholar]
- Xu S., Sunderland M. E., Coles B. L., Kam A., Holowacz T., Ashery-Padan R., Marquardt T., McInnes R. R., van der Kooy D. (2007). The proliferation and expansion of retinal stem cells require functional Pax6. Dev. Biol. 304, 713–721 10.1016/j.ydbio.2007.01.021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Young T. L., Cepko C. L. (2004). A role for ligand-gated ion channels in rod photoreceptor development. Neuron 41, 867–879 10.1016/S0896-6273(04)00141-2 [DOI] [PubMed] [Google Scholar]
- Zahs K. R., Ceelen P. W. (2006). Gap junctional coupling and connexin immunoreactivity in rabbit retinal glia. Vis. Neurosci. 23, 1–10 10.1017/S0952523806231018 [DOI] [PubMed] [Google Scholar]
- Zhu X., Craft C. M. (2000). Modulation of CRX transactivation activity by phosducin isoforms. Mol. Cell. Biol. 20, 5216–5226 10.1128/MCB.20.14.5216-5226.2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu X., Brown B., Li A., Mears A. J., Swaroop A., Craft C. M. (2003). GRK1-dependent phosphorylation of S and M opsins and their binding to cone arrestin during cone phototransduction in the mouse retina. J. Neurosci. 23, 6152–6160 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}
{kind=link}