

Abstract
Streptococcus suis serotype 2 (SS2), a major swine pathogen and an emerging zoonotic agent, has greatly challenged global public health. The encoding proteins with unknown functions the bacterium encodes are an obstruction to studies of the pathogenesis. A novel surface protective antigen HP0197 is one of these proteins which have no sequence homology to any known protein. In the present study, the protein was determined to be involved in bacterial virulence through an evaluation of the isogenic mutant (Δhp0197) in both mice and pigs. The experimental infection also indicated that Δhp0197 could be cleared easily during infection, which could be attributed to the reduced thickness of the capsular polysaccharides (CPS) and the significantly reduced phagocytotic resistance. Microarrays-based comparative transcriptome analysis suggested that the suppressed expression of the operon responsible for CPS synthesis might be reversed by CcpA activity, which controlled global regulation of carbon catabolite through the binding of the CcpA and HPr-Ser-46-P to the catabolite-responsive elements (cre) of the target operons. The hypothesis was approved by the fact that the purified FLAG-tagged HPr from WT stain exhibited a higher binding activity to cre with CcpA compared to the Δhp0197 by the Electrophoretic Mobility Shift Assay, suggesting lower level of phosphorylation of the phosphocarrier protein HPr at residue Ser-46 (HPr-Ser-46P) in Δhp0197. These indicated that HP0197 could enhance CcpA activity to control the expression of genes involved in carbohydrate utilization and CPS synthesis, thus contributing to the virulence of S. suis.
Introduction
Streptococcus suis (S. suis) is a major swine pathogen responsible for severe economic losses in the porcine industry. It is also a severe threat to human health, especially to people who are in close contact with swine or pork by-products. S. suis serotype 2 (SS2) is considered the most pathogenic and the most prevalent capsular type among the thirty-three serotypes (types 1 to 31, 33, and 1/2) [1], [2], [3]. The infection caused by S. suis has been reported in more than 20 countries, in which more than 700 people have been infected since the first reported case of S. suis-caused human meningitis in Denmark in 1968 [4]. Two recent large-scale outbreaks of human SS2 epidemics in China (the first had 25 cases with 14 deaths in Jiangsu in 1998, and the second had 204 cases with 38 deaths in Sichuan in 2005) have raised people’s awareness of the public health threat because the cases presented clinically with a streptococcal toxic shock-like syndrome, which indicates that new virulent bacterial variants are currently emerging in Asia [5], [6], [7], [8], [9], [10], [11]. Besides, the bacteria have also caused sporadic human illness in other countries [12], including Thailand, the United Kingdom, Portugal, Italy, Japan, Australia, Netherlands and the United States. S. suis has also been identified as the third most common cause of community-acquired bacterial meningitis in Hong Kong and as the leading cause of adult meningitis in Vietnam [7], [13], [14]. The infected cases have occasionally been documented in North America, and some investigators believe that more cases could have happened than what have been reported, and the relatively low number of reported cases of human S. suis infections is hypothesized to be a result of misdiagnosis rather than the true absence of the disease [15], [16].
Herein, S. suis infection has attracted a great deal of attention from the scientific community and the popular press [17]. However, the current understanding of the S. suis pathogenesis is still limited. The polysaccharide capsule has generally been considered to be essential for bacterial virulence [18], [19], [20]. Suilysin, the extracellular protein factor, along with a muramidase-released protein have also been shown to be linked to, but not essential for, the full virulence of S. suis [21]. GapdH [22], Enolase [23], [24], Fibronectin/Fibrinogen-binding protein [25], HAM1 [26] and Adhesion [27], [28], [29], [30], [31], [32] have been shown to be involved in S. suis adherence and virulence. Serum opacity-like factor [33], D-Alanylation of Lipoteichoic Acid [34], Peptidoglycan GlcNAc deacetylase [35], IgA protease [36], [37], TroA [38], SodA [39], SsFHB [40] and Subtilisin-like serine protease [41], [42], [43] are also considered to be related to S. suis virulence. In addition, studies have reported that SalK/SalR [44] and CovR [45] affected the virulence of Chinese isolates of S. suis. Although series virulence-associated factors have been discovered, which helped people understand S. suis pathogenesis, the infection process related to the disease remains unknown.
Genomics and proteomics studies have revealed that there are several hypothetical proteins on the surface of the bacterium [9], [46], [47], [48], [49], [50]. Among these, HP0197 (SSU98_0197 in strain 98HAH33 or SSU05_0196 in strain 05ZYH33) has an YSIRK-type signal peptide presented at the N terminus and a typical C-terminal sorting signal of “LPATG” motif and is located on the bacterial surface [48], [50]. Our previous study indicated that HP0197 presented in all tested clinical SS2 isolates and was identified as a surface protective antigen which could confer significant protection against challenge with lethal dose of SS2 in mice and pigs [51]. All of these information indicated that the protein may be involved in the pathogenesis of S. suis. However, this hypothetical protein has no similar domain architectures except for a G5 domain (pfam07501) which has been identified in several proteins involved in the metabolism of the bacterial cell walls [52]. Therefore, it is not easy to predict the function of the protein by bioinformatics analysis because only 76 aa (418–493 aa) out of 562 aa are found to be similar to the G5 domain; the rest of this protein is not similar to any known protein domain. Thus, evaluation on the contribution of HP0197 to the virulence would strengthen the understanding of this protein, and undoubtedly be conductive to the understanding of the pathogenicity of the S. suis.
Results
Construction of Δhp0197
To study the role of HP0197 in S.suis virulence, an isogenic hp0197 mutant was derived from strain 05ZY ( Fig. 1A ). In Δhp0197 mutant, hp0197 gene (1.7 kb) was replaced with egfp gene (0.75 kb). So PCR with primer pairs of HP0197-P1/HP0197-P2 which targeted to the 176 bp and 252 bp from the upstream and downstream sequence of hp0197 gene respectively could be used to confirm the deletion of hp0197 gene. A 2.1 kb or 1.2 kb DNA band was obtained respectively when the genome DNA of WT or Δhp0197 strain served as a template. Besides, an egfp gene could not be amplified from the genome of WT strain but from the constructed Δhp0197. These tests indicated that hp0197 gene was successfully replaced by egfp gene ( Fig. 1B and 1C ). HP0197 antibodies could react with proteins extracted from WT and cΔhp0197 (complemented strain of Δhp0197) rather than from Δhp0197 in the immunoblotting assay, which indicated no expression of HP0197 in Δhp0197 and the successful complementation for cΔhp0197 strain ( Fig. 1D ).
Figure 1. Construction and confirmation of Δhp0197 and a complementary strain, cΔhp0197.

(A) Construction strategy for Δhp0197, which was derived from the SS2 strain 05ZY. Approximately 600 bp of sequence flanking hp0197 was constructed into the temperature-sensitive S. suis-E. coli shuttle vector pSET4s, hp0197 was replaced with egfp. (B) The egfp gene could not be amplified from the WT strain (lane 1 and 2) but was amplified from the Δhp0197 (lane 5 and 6) and cΔhp0197 strains (lane 3 and 4). (C) A PCR targeted to the 176 bp and 252 bp from the upstream and downstream sequence of hp0197 sequence respectively with primer pairs of HP0197-P1/HP0197-P2 was performed to confirm the successful construction. And a 2.1-kb band could be amplified from DNA of the WT strain (lanes 1 and 2), and a 1.2-kb DNA fragment could be amplified from the Δhp0197 mutant (lanes 5 and 6). Two bands were amplified from the cΔhp0197 strain (lanes 3 and 4). Lane 7 in A and B were PCR negative controls. (D) HP0197 antibodies did not bind proteins extracted from the Δhp0197 mutant strain (lane 2) but was able to bind proteins extracted from the WT strain (lane 1) and the cΔhp0197 strain (lane 3) in the immunoblot assay. (E) The growth rates of S. suis strains in THB. (F) qRT-PCR amplification of hp0197 (ssu98_0197) flanking genes (ssu98_0195 to ssu98_0199) yielded identical results in WT and Δhp0197 while hp0197 could only be amplified from WT.
Δhp0197 showed slower growth rate than WT ( Fig. 1E ), which provoked us to illustrate whether this phenomenon was resulted from the polar effect when hp0197 gene was deleted. The expression of the hp0197 flanking genes (SSU98_0196 and SSU98_0198) was not changed significantly in WT and Δhp0197 by qRT-PCR, which further indicated that the deletion of hp0197 could not cause obvious polar effect ( Fig. 1F ). These results suggested that the deletion of hp0197 may retard the growth rate of S. suis at given time course.
HP0197 Contributes Significantly to S. suis Virulence in Mice
HP0197 has been identified as an important vaccine candidate that exists in almost all tested pathogenic clinical SS2 isolates [51]. Therefore, it is worth evaluating its role in S. suis virulence. Independent experimental infections with the wild-type (WT), Δhp0197 and cΔhp0197 were performed in mice ( Fig. 2 ). While all mice manifested depression, weakness and prostration during the first day post-inoculation (pi), the mice in the WT and cΔhp0197 groups showed more severe clinical signs. Eighty percent of the mice in the WT group and 90% of the mice in the cΔhp0197 group died within the first 2 days pi. After 2 days, the remaining mice recovered from serious clinical symptoms. In contrast, only 2 mice in the Δhp0197 group died on day 1; the rest of the mice survived during the trial ( Fig. 2A ). These indicated that the deletion of hp0197 decreased the lethality of SS2 to mice (P<0.05). The loss of body weight usually reflects upon the health of the mice, and the body weight of mice in Δhp0197 group recovered more rapidly than mice in WT group since day 3 p.i. (p<0.05) ( Fig. 2B ), which further indicated that the deletion of hp0197 decreased the pathogenicity of SS2 to mice.
Figure 2. HP0197 contributes significantly to S. suis virulence in mice and pigs.

(A) The survival of experimental mice inoculated with WT, Δhp0197 and cΔhp0197 strains. Ten mice in each group were inoculated by intraperitoneal injection with 109 CFU of WT, Δhp0197 or cΔhp0197 respectively and the mice were monitored for clinical signs of infection three times a day during 7 days. “*” represents P value <0.05 of WT versus Δhp0197 group or Δhp0197 versus cΔhp0197 group respectively. (B) The loss of body weight of mice inoculated with the WT, Δhp0197 and cΔhp0197 strains, which usually reflects the health status of mice. “*” represents P value <0.05 of WT versus Δhp0197 group. (C) The kinetics of bacterial clearance from the blood of mice inoculated with the WT, Δhp0197 and cΔhp0197 strains. “*” represents P<0.05 of WT versus Δhp0197 group at given time course. (D) The survival of experimental pigs inoculated with WT and Δhp0197 strains. Eight pigs in each group were inoculated intranasally with 2×107 CFU of WT and Δhp0197 strains respectively. Four pigs were inoculated with PBS and served as a control. “*” represents P<0.05 of WT versus Δhp0197 group. (E) Clinical scores (daily means and standard deviations) of pigs after inoculation. “*” represents P value <0.05 of WT versus Δhp0197 group at given time course.
The resistance to clearance from circulation could reflect the evasion of pathogen from host defense system. The level of recovered bacterial in each group did not show significant difference on day 1 p.i., and the bacterial reached the highest level on day 2 p.i. ( Fig. 2C ). However, the recovered SS2 in Δhp0197 group decreased more rapidly on day 3 p.i. than WT group (p<0.05). Furthermore, no bacterium could be isolated from circulation of Δhp0197 group since day 4 p.i. while bacteria still sustained in the blood of mice in WT group or in cΔhp0197 group. These indicated that Δhp0197 was easily cleared by the host ( Fig. 2C ).
This trial revealed that compared to the WT strain, Δhp0197 was significantly attenuated and that the reintroduction of the hp0197 gene into Δhp0197 (cΔhp0197) restored high levels of pathogenicity to the bacterial infection. The experimental infection in the mice strongly suggested that HP0197 played an important role in the pathogenesis of SS2 to mice.
Infection with the Δhp0197 is Significantly Attenuated in Pigs
To further delineate the role of hp0197 in S. suis virulence, we conducted a trial in pigs, which are the natural hosts of infection. Four pigs in the sham-inoculated group did not manifest any clinical signs of infection during the test. Conversely, all of the pigs in the WT group (n = 8) died within 3 days ( Fig. 2D ), and they all showed severe clinical signs of disease, including severe respiratory symptoms, lameness associated with swollen joints and central nervous system signs that were characterized by ataxia, recumbency and opisthotonus ( Fig. 2E ). Macroscopic lesions typical of S. suis infection were identified in all animals infected with the WT strain. The predominant gross lesions observed in the WT-infected group were meningitis, lung lesions which were characterized by well-demarcated red-purple consolidation involving 10% to 20% of the lung, fibrin deposits in the abdominal cavity, and increased fluid in the joints. Obvious histopathological lesions were also observed (Fig. S1). In comparison, animals inoculated with the Δhp0197 strain showed no clinical signs of infection, and all of the pigs survived for the duration of the test ( Fig. 2D ). Similarly, no obvious macroscopic lesions or histopathological lesions associated with S. suis were observed in any animals in this group (Fig. S1). S. suis was detected in the blood of the pigs in the WT group (range from 106 to 108 CFU ml−1), but no bacteria could be isolated from the blood of any of the pigs in the Δhp0197 group at any given point during the test. The experimental infection on pigs also indicated that hp0197 contributed significantly to the virulence of SS2.
Contribution of hp0197 to the Resistance to Phagocytotic Killing and to CPS Synthesis
The experimental infection demonstrated that Δhp0197 could be cleared easily during infection ( Fig. 2 ), which suggested that Δhp0197 was not as efficient in avoiding destruction by phagocytotic processes as its parent strain. To confirm this hypothesis, in vitro killing of the WT strain and Δhp0197 by phagocytes was compared in the presence of normal complete serum without opsonizing antibodies. Compared to the WT strain (only 5.0±4.2% of bacteria were killed), which could resist the bactericidal effect of phagocytes [18], [53], the isogenic cps mutant derived from strain 05ZYS (Δcps) was readily destroyed by phagocytes as described before [18]. Interestingly, a notable number of the Δhp0197 bacteria (28.0±4.6%) were killed and the reintroduction of hp0197 into Δhp0197 could recover the resistance to the phagocytic processes to some extent (14.1±9.1%) ( Fig. 3A ). Furthermore, these bacteria showed no significant difference without murine phagocytes. Therefore, the data suggested that hp0197 directly or indirectly contributes to the resistance of S. suis to phagocytic killing.
Figure 3. Contribution of hp0197 to the resistance to phagocytotic killing and to CPS synthesis.

(A) Percent of bacteria killed after 90 min incubation with murine phagocytes. The experiment was repeated five times. Error bars indicate standard deviations. “*” represents P value <0.05 of WT versus Δcps group or WT versus Δhp0197 group respectively. (B) Morphology of the WT (left), the Δhp0197 (middle) and cΔhp0197 (right) strains. Transmission electron microscopy revealed that the WT strain was surrounded by a thick polysaccharide capsule (52.8±10.4 nm); the thickness of the Δhp0197 capsule was markedly reduced (1.1±0.3 nm), and cΔhp0197 could if not completely, partially restore this phenotype (21.1±6.6 nm).
CPS is thought to be a critical antiphagocytic factor that can down-regulate signaling pathways involved in macrophage phagocytosis, and could protect S. suis against killing and phagocytosis [54], [55]. The decreased capacity of Δhp0197 to resist phagocytotic killing encouraged us to test the effect of hp0197 on CPS thickness. Electron microscopical characterization of the WT strain revealed a thick CPS, (52.8±10.4 nm). In contrast, Δhp0197 exhibited a markedly reduced thickness of the capsule (1.1±0.3 nm), corresponding to 2.1% of WT, and cΔhp0197 could restore the phenotype to some extent (21.1±6.6 nm) ( Fig. 3B ). This suggested that hp0197 could contribute to CPS thickness, which in turn affect the resistance of the bacteria to phagocytotic killing.
Microarrays-based Comparative Transcriptomics Analysis of Δhp0197 and WT Suggests the Correlation between hp0197 and CcpA Activity
To gain insight into the role of hp0197 in CPS synthesis and virulence, the transcriptome profile of Δhp0197 was compared to that of WT using a SS2 genomic microarray. A total of 193 genes were differentially expressed (change ratio> = 2, p-value <0.05), among which 150 genes were up-regulated and 43 genes were down-regulated ( Fig. 4A and Table S1, S2). The different expression patterns identified by the microarrays analysis were validated by qRT-PCR based on the data obtained from a total of 12 genes ( Fig. 4B ), and the comparative proteomics based on the 2-DE/MS (Fig. S2 and Table S3) further partially confirmed the differential expression.
Figure 4. Microarray-based comparative transcriptomic analysis of Δhp0197 and WT strains.

(A) Microarray-based comparative transcriptomic analysis of Δhp0197 and WT strains. A hierarchical cluster of 193 transcripts in the Δhp0197 compared to the WT strain. The increased and decreased transcript expression levels were indicated by red and green, respectively. (B) Different expression profiles identified by microarray analysis were validated by qRT-PCR based on the data obtained from a total of 12 genes. From left to right: ssu05_1372, ssu05_1933, ssu05_0167, ssu05_2076, ssu05_2137, ssu05_1039, ssu05_0360, ssu05_1401, ssu05_0812, ssu05_0573 (cps 2J), ssu05_0265 and ssu05_0469. (C) Functional classification of the differentially expressed genes. J: Translation, ribosomal structure and biogenesis; K: Transcription; L: Replication, recombination and repair; D: Cell cycle control and cell division; V: Defense mechanisms; T: Signal transduction mechanisms; M: Cell membrane and envelope biogenesis; U: Intracellular trafficking, secretion and vesicular transport; O: Posttranslational modification, protein turnover, chaperones; C: Energy production and conversion; G: Carbohydrate transport and metabolism; E: Amino acid transport and metabolism; F: Nucleotide transport and metabolism; H: Coenzyme transport and metabolism; I: Lipid transport and metabolism; P: Inorganic ion transport and metabolism; Q: Secondary metabolites biosynthesis, transport and catabolism; R: General function prediction only; S: Function unknown; Not: Not in COGs.
The functional classifications of these differentially expressed genes provided clear indications of the changes that occurred in Δhp0197 compared to WT ( Fig. 4C ). It was intriguing that 48% (72 of 150) of the up-regulated genes were predicted to encode proteins involved in carbohydrate transport and metabolism (COG: G) when hp0197 was absent, including ATP-binding cassette (ABC) transporters and PTS operons responsible for the transport of glucose, lactose, maltose/maltodextrin, mannose, fructose, and cellobiose (Table S2).The differential expressions of these genes have also been documented in a study on Group A Streptococcus when ccpa (catabolite control protein A), a global regulator of sugar metabolism, was absent [56]. Among the up-regulated genes, 57% (86 of 150) were documented in the Δccpa originated from SS2 [57]. Not surprisingly, 8/19 genes loci which had a putative CcpA-binding sequence “cre” were significantly up-regulated. Interestingly, 39% (17 of 43) of the down-regulated genes showed lower expression levels in the isogenic ccpa mutant [57], including the entire operon locus responsible for CPS synthesis (COG: M), which was confirmed by qRT-PCR for ssu05_0573 (cps 2J) (Fig. 4B). Additionally, lower expression of hp0197 (ssu98_0197 in strain 98HAH33 or ssu05_0196 in strain 05ZYH33) was also observed in the Δccpa [57]. Therefore, a correlation must exist between hp0197 and CcpA activity.
hp0197 Controls the CcpA Activity via HPr-Ser-46P
CcpA is a critical global regulator of carbohydrate metabolism and controls the expression of genes involved in complex carbohydrate utilization and virulence, such as an operon for CPS synthesis. A lower CcpA activity could contribute to the reduced thickness of the CPS and the decreased virulence in group A Streptococcus and S. suis [56], [57]. However, CcpA expression was not down-regulated as anticipation but up-regulated in Δhp0197. Therefore, we attempted to determine why CcpA was up-regulated while the cre-binding activity was decreased in Δhp0197 mutant.
CcpA activity is controlled by a complex interaction with HPr [58]. When glucose or other readily metabolized carbohydrates are present at sufficiently high concentrations, phosphorylation of the phosphocarrier protein HPr at residue Ser-46 (HPr-Ser-46P) occurs. This protein is a co-effector for CcpA that could mediate CcpA binding to cre sites [59]. To determine whether hp0197 controls CcpA activity through HPr, we purified FLAG-tagged HPr from both the WT and Δhp0197 bacterial strains. Purified HPr from log-phase WT bacteria had good binding activity to cre in associated with purified recombinant His-tagged CcpA, but HPr from Δhp0197 had a weaker binding effect ( Fig. 5 ), indicating that lower levels of HPr-Ser-46-P were produced in Δhp0197. Lower activity could also be observed when HPr was isolated from stable-phase WT bacteria, which had very low level of HPr-Ser-46 due to insufficient energy. These data supported the hypothesis that hp0197 controlled CcpA activity by HPr-Ser-46P binding.
Figure 5. hp0197 controls CcpA activity by HPr-Ser-46P.

The purified FLAG-tagged HPr from the log-phase WT or Δhp0197 bacteria were compared to evaluate the binding of CcpA to its target site (cre) by EMSA. The HPr from the stable-phase WT strain served as a control. Significant more DNA (indicated by *) with CcpA and HPr from Log-phase WT migrated slower than DNA with CcpA and HPr from Δhp0197 or from the stable-phase WT strain, indicating that HPr from log-phase WT could enhance the binding of CcpA to cre and the HPr from log-phase Δhp0197could not enhance the ability significantly.
Discussion
The 2005 Sichuan outbreak of S. suis provoked strong interest and extensive research; some progress has been achieved through genomic and proteomic studies [17], [60]. However, many aspects of the pathogenesis of the bacteria remain uncertain. For example, the biological properties of a few hypothetical proteins encoded by the bacteria need to be characterized.
Several hypothetical proteins are expressed on the surface of the S. suis, and several studies have suggested their potential roles in bacterial pathogenesis [47], [48], [50], [51], [61], [62]. HP0197 is one such hypothetical protein that does not have significant sequence homology to any other known protein and does not have similar domain architecture besides the G5 domain, which accounts for only 14% of the total amino acid length. These findings indicate that HP0197 is a novel protein. Furthermore, a previous study [51] demonstrated that HP0197 was an important vaccine candidate and was present in almost all tested pathogenic clinical SS2 isolates. Therefore, it was believed that clarifying its biological function would contribute to an increased understanding of its gene structure and its biological functions.
To clarify the pathogenic role of hp0197 in S. suis virulence, mice and pigs were experimentally infected with the bacteria. WT strain cause high mortality in mice (80%) and pigs (100%), and mortality was significantly reduced when hp0197 was absent; only 2 mice (n = 10) and no pig (n = 8) died during the experimental infection ( Fig. 2 ). Additionally, obvious clinical signs, significant pathological lesions, and high levels of bacteria in the blood were observed in the WT-infected animals. However, only slight or no significant clinical signs and pathological lesions were observed in mice or pigs inoculated with Δhp0197, respectively (Fig. S1). In contrast to pathogenic SS2, Δhp0197 infection did not induce high levels of cytokine expression [63], which could be provoked by highly pathogenic SS2 [55], [63], [64]. These data indicated that hp0197 significantly contributed to S. suis virulence.
During the experimental infection, no bacteria could be isolated from the Δhp0197-infected pigs, and the number of strains recovered from the blood in Δhp0197-infected mice was much lower and was cleared quickly compared to WT ( Fig. 2C ). These results suggested that Δhp0197 could not resist phagocytotic killing. The in vitro assay confirmed that hp0197 played an essential role in the resistance of S. suis to phagocytic killing ( Fig. 3A ); this was attributed to a reduced thickness of the CPS in Δhp0197 mutant ( Fig. 3B ).
How could a surface protein control CPS thickness and also influence S. suis virulence? To answer this question, we compared the transcriptome profiles of Δhp0197 and WT, and determined that 53% (103 of 193) of the differentially expressed genes in Δhp0197 showed expression profiles that were similar to the Δccpa [57]. CcpA is a critical global regulator of carbohydrate metabolism and controls the expression of genes involved in complex carbohydrate utilization and virulence, such as an operon for CPS synthesis [56], [57]. Therefore, a correlation should exist between hp0197 and CcpA activity. However, CcpA expression was not down-regulated but up-regulated in Δhp0197 (Table S2). In fact, CcpA activity is controlled by a complex interaction with HPr [58]. When glucose or other readily metabolized carbohydrates are present at sufficiently high concentrations, HPr is phosphorylated at residue Ser-46 (HPr-Ser-46P), which subsequently served as a co-effector for CcpA that could mediates CcpA binding to cre sites [59]. Purified HPr from Δhp0197 had a decreased binding activity to CcpA compared to WT ( Fig. 5 ), suggesting that lower levels of HPr-Ser-46-P were produced in Δhp0197. This result suggests that HP0197 contributed to the utilization of sugars. However, none of the genes responsible for glycolysis or for the phosphoenolpyruvate-dependent carbohydrate:phosphotransferase system (PTS) of glucose were observed to be down-regulated (Table S1) and some of them were even up-regulated (Table S2) in Δhp0197. This suggested that HP0197 could not contribute to transport and glycolysis of glucose and might facilitates the uptake of saccharides.
In summary, the deletion of hp0197 would decrease the activity of CcpA, and would further alleviate the carbon catabolite repression and down-regulate genes for CPS synthesis. This would decrease the bacterial resistance to phagocytotic killing and would thus decrease the virulence of S. suis ( Fig. 6 ). In conclusion, HP0197 contributes to CPS synthesis and the virulence of Streptococcus suis via CcpA.
Figure 6. Schematic representation how HP0197 contributes the CPS sysnthesis and virulence of Streptococcus suis via CcpA activity.

HP0197 could promote the level of phosphorylation of the phosphocarrier protein HPr at residue Ser-46 (HPr-Ser-46P), which could enhance the binding of CcpA to the cre of the target operons, subsequently control the expression of genes involved in carbohydrate utilization and CPS synthesis. A thin CPS could allow the pathogenic bacteria to be killed more easily by phagocytes and cleared from the host. Thus, HP0197 ultimately contributes to CPS synthesis and the virulence of S. suis via CcpA activity.
Materials and Methods
Bacterial Strains, Plasmids and Growth Conditions
S. suis serotype 2 strain 05ZY was chose as the WT strain. This strain was isolated from the brain of a diseased piglet and expresses muramidase-released protein, extracellular protein factor and suilysin was chosen as wild type (WT) strain [5]. A temperature-sensitive S. suis-E. coli shuttle vector (pSET4s), that carries spectinomycin resistance gene (spcr), was used to construct the Δhp0197 mutant. pSET2, a S. suis-E. coli shuttle vector, carrying spcr, was used in the construction of the complementary bacterium.
S. suis strains were grown in Todd–Hewitt broth (THB) or on Todd-Hewitt agar (THA) (Becton Dickinson, Sparks, MD, USA) at 37°C under aerobic conditions. E. coli (DH5α) was cultured in Luria-Bertani broth or agar (Becton Dickinson) at 37°C for 8 h. When necessary, spectinomycin was added to the culture media at the following concentrations: 50 µg/ml for E. coli and 100 µg/ml for S. suis.
Construction of Δhp0197 and Complementation
The isogenic hp0197 mutant was derived from the SS2 strain 05ZY using the strategy of allelic replacement mutagenesis. Approximately 600 bp of hp0197 flanking sequence was obtained by PCR using the 05ZY chromosome as a template with the primer pairs HP0197L1/HP0197L2 (left arm) or HP0197R1/HP0197R2 (right arm). An egfp-conding region was amplified from plasmid pEGFP-N1 with primer pairs EGFP-1/EGFP-2 to exchange egfp with the hp0197 coding sequence (Table S4). All PCR amplicons were digested with the appropriate restriction enzymes and were sequentially ligated into the temperature-sensitive S. suis-E. coli shuttle vector pSET4s ( Fig. 1 ) for constructing the knockout vector pSET4s::hp0197. To obtain the isogenic hp0197 mutant, competent cells of S. suis were subjected to electrotransformation with pSET4s::hp0197 and were grown at 28°C in the presence of spectinomycin selection as described by Takamatsu et al. [65]. Bacteria at the mid-logarithmic growth phase were diluted with THB containing spectinomycin and cultured at 28°C to early logarithmic phase. The culture was then shifted to 37°C and incubated for 4 h. Subsequently, the cells were spread on THA and incubated at 28°C. Temperature resistant colonies were screened at 37°C for the loss of vector-mediated spectinomycin resistance, and the putative mutants were detected in which the hp0197 gene was replaced by egfp gene as a consequence of homologous recombination via a double cross-over. The deletion of hp0197 in SS2 was further confirmed by PCR analysis with the HP0197-P1/HP0197-P2 and EGFP-L/EGFP-R primer pairs, and by immunoblotting with antiserum raised against purified recombinant HP0197 protein [51].
To compliment hp0197, a DNA fragment containing the hp0197 gene and its predicted upstream promoter was amplified by PCR with primers cHP0197-1/cHP0197-2 (Table S4). The obtained DNA fragment was digested with the appropriate restriction enzymes and was cloned into the E. coli-S. suis shuttle vector pSET2 to generate the recombinant plasmid pSET2::Chp0197. The plasmid was then introduced into Δhp0197 to screen the complemented cΔhp0197 strain on THA with selection pressure of spectinomycin. The HP0197 expression was confirmed by the immunoblotting with the antiserum as previously described.
Experimental Infections of Mice and Pigs
This study was performed in strict accordance with the Guide for the Care and Use of Laboratory Animals Monitoring Committee of Hubei Province, China, and the protocol was approved by the Committee on the Ethics of Animal Experiments at the College of Veterinary Medicine, Huazhong Agricultural University (Permit Number: 00010399). All efforts were made to minimize the suffering of the animals.
Thirty 6-week-old female CD-1 mice were randomly assigned to 3 groups with 10 mice in each group. Group 1 was inoculated by intraperitoneal injection of 1 ml of a WT strain suspension at 109 CFU/ml, and groups 2 and 3 received the same dose of the Δhp0197 and cΔhp0197, respectively. The mice were monitored constantly three times a day for 7 days for clinical signs, and were assigned clinical scores as described by Dominguez-Punaro [66]. Mice exhibiting extreme lethargy or neurological signs were considered moribund and were humanely euthanized. The survived animals were sacrificed via carbon dioxide inhalation at the end of the experiment. To evaluate the bacterial load, blood samples were collected daily from the tail vein and at the time of euthanasia by cardiac puncture and were plated on THA plates.
A total of 20 healthy pigs (ages 4–5 weeks) from a herd that was free of SS2 were assigned to three groups. One hour before inoculation, all pigs were given 2 ml of 2% acetic acid intranasally to enhance the severity of the S. suis challenge [67]. Pigs in group 1 (n = 8) and group 2 (n = 8) were inoculated intranasally with 2 ml of 107 CFU of WT and Δhp0197 strains, respectively. Group 3 (n = 4) was inoculated with PBS as a control. Clinical signs of infection and the presence of S. suis in the blood were monitored during the trial. Clinical symptoms were quantified by clinical scores as described by Li et al [68]. Briefly, a daily clinical score (from 0 to 8) was derived as the sum of the attitude and locomotion scores for each pig. The scores were based upon signs of nervous, musculoskeletal, and respiratory diseases. Attitude scores were given as follows: 0 = normal attitude and response to stimuli; 1 = inactive and slow to respond, with oculonasal secretions; 2 = only responsive to repeated stimuli; 3 = recumbent, nonresponsive, and unaware of surroundings; and 4 = dead. Locomotion scores were given as follows: 0 = normal gait and posture; 1 = slight lack of coordination, lameness, and/or joint swelling but able to stand without assistance; 2 = clearly uncoordinated or lame but able to stand without assistance; 3 = severe lameness and/or severe ataxia; and 4 = dead. Pigs with a clinical score of >2 on either scale were euthanized by lethal injection. The survived animals in all three test groups were sacrificed at day 7 pi and were examined for pathological lesions.
Killing by Murine Phagocytes
Murine phagocytes were isolated as previously described [69]. Blood samples were collected by venous puncture from mice, and cell populations were separated by Ficoll 400-Hypaque density gradient centrifugation. The murine phagocytes were isolated by sedimentation in 6% dextran and the contaminating erythrocytes were removed by lysis with 0.83% ammonium chloride. The collected phagocytes were counted by trypan blue staining, and then were adjusted to a density of 1×106 cells/ml. For each sample, 100 µl of bacteria at a concentration of approximately 1×104 CFU/ml was opsonized with an equal volume of complete normal murine serum for 30 min at 37°C and was mixed in a microcentrifuge tube with 100 µl of murine phagocytes. The mixture was incubated for 90 min at 37°C in a 5% CO2 environment. The bacteria were not toxic to the phagocyte under these conditions [18]. After incubation, the cells were lysed with sterile water. Viable bacterial counts were determined by plating the bacteria onto THA. The parallel experiment without phagocytes has been done to evaluate the killing effects of serum.
Electron Microscopy
For morphological analysis of the capsule structure, samples from early exponential grown bacteria were fixed according to a lysine/acetate-based formaldehyde/glutaraldehyde ruthenium red-osmium fixation procedure as previously described [70]. In addition, the capsule thickness was measured from randomly selected bacteria using the iTEM software.
Microarray-based Comparative Transcriptomic Analysis of Δhp0197
Based on the whole genome sequence of 05ZYH33, specific 40- to 60-mer oligo-nucleotides were designed to cover every putative ORF (2194 in total) in the genome using the eArray EArray Online Microarray Design Tool (https://earray.chem.agilent.com/earray/) and were printed six times on the surface of each microarray slide (Agilent).
Total RNAs was extracted with Trizol reagent (Invitrogen) from Δhp0197 or WT strain cultures grown in THB to an OD600 of 0.35, and were purified using an RNeasy Mini Kit (Qiagen). RNA was reverse-transcribed and was then transcribed into cRNA. Purified cRNA from Δhp0197 or WT strain were labeled with Cy5 NHS ester or Cy3 NHS ester (GE healthcare), respectively and were purified again. After fragmentation, microarray slides were hybridized with 1 µg of Cy5- or Cy3-labeled cDNAs at 65°C for 17 h with 10 rpm rotation. All hybridization slides were scanned by an Agilent Microarray Scanner System (G2565BA) after appropriate washing. The p-values and corrected p-values was calculated by Benjamin-Hochberg FDR estimation (GeneSpring software 11.0, Agilent) in the four independent bio-replicates. Only those genes with more than twice of change ratio and corrected a p-value <0.05 were selected as differentially expressed genes.
qRT-PCR Evaluation
To validate the data from the microarray analysis, 12 genes and the hp0197 flanking genes (SSU98_0196 and SSU98_0198) (Table S5) were selected to evaluate the expression changes in WT and Δhp0197 by qRT-PCR with SYBR Green detection [71]. Three independent cultures of each strain (WT and Δhp0197) grown in THB to an OD600 of 0.35 were sampled and quenched in liquid nitrogen. Total RNAs were extracted with Trizol reagent (Invitrogen) and were purified using an RNeasy Mini Kit (Qiagen). The quality and quantity were determined by a 2100 Agilent bioanalyzer and by Nanodrop ND1000 UV spectroscopy. Quantitative analysis was performed in triplicate with an ABI PRISM 7500 sequence detection system using RNA from three independent cultures of each strain (WT and Δhp0197). 16SrRNA was chosen as the internal control. Various Ct values were normalized to the average Ct value for 16SrRNA. The mean fold changes in target gene expression was calculated as described by Livak & Schmittgen [72].
Determination of CcpA Activity by Electrophoretic Mobility Shift Assay (EMSA)
The purified FLAG-tagged HPr from log-phase and stable-phase of WT and Δhp0197 were compared to determine the binding of CcpA to cre as described by Lance M Hellman and Michael G Fried [73]. Purified recombinant His-tagged CcpA (1.5 µg), along with 1.5 µg of purified HPr and 0.3 µg of DNA harboring cre, were prepared and subjected to electrophoresis under native conditions in a polyacrylamide gel. After electrophoresis, the distribution of species containing nucleic acid was determined by EB staining.
Statistical Analysis
Unless otherwise specified, data were analyzed by two-tailed, unpaired t-tests, and all assays were repeated at least three times. The data were expressed the means ± standard deviations. For in vivo virulence experiments, survival was analyzed with the Log-Rank test. For all tests, a value of P<0.05 was considered as the threshold for significance.
Supporting Information
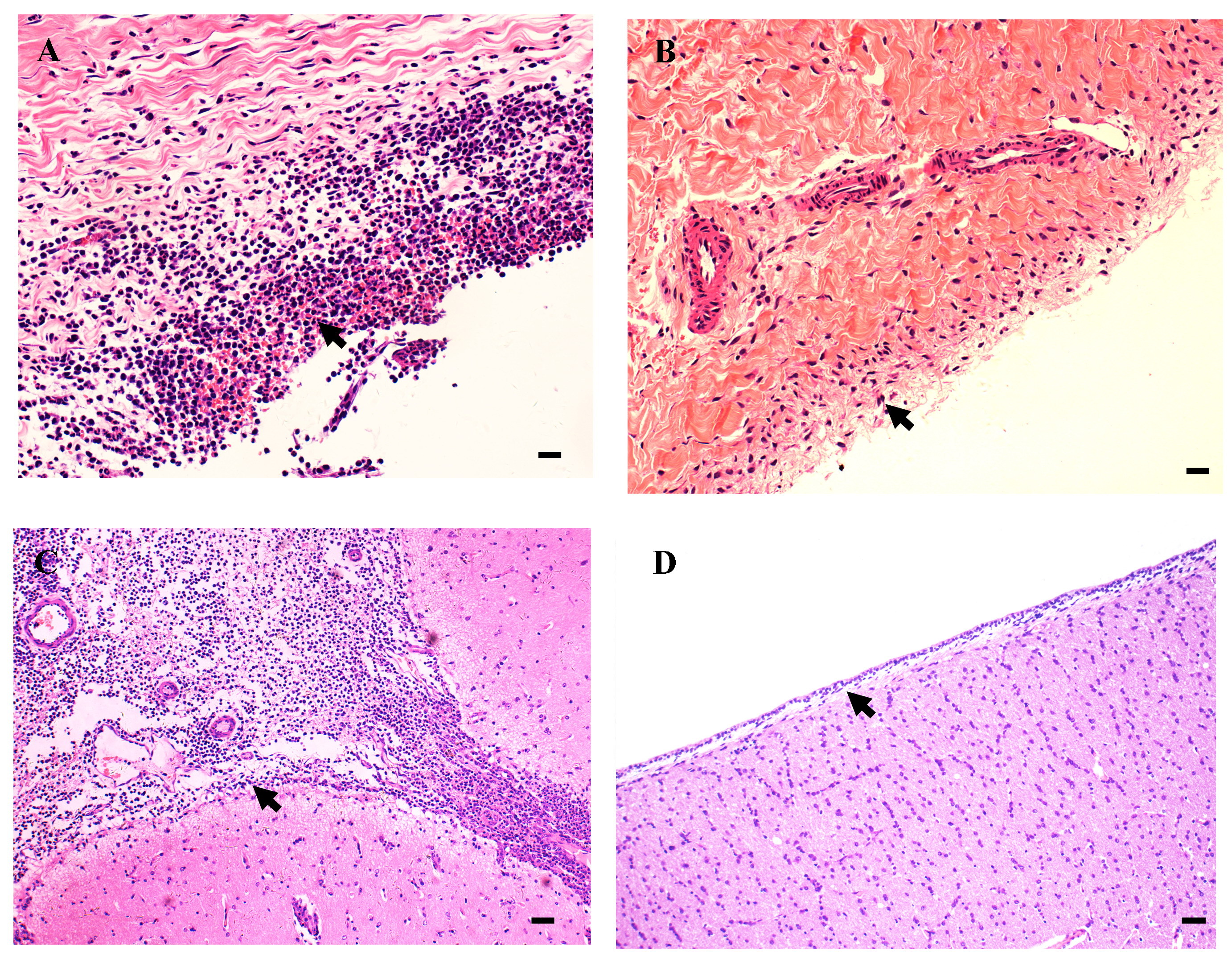
Relevant histopathological findings could be observed from WT-infected pigs (day 3 pi) while no significant histopathological findings could be observed from a representative pig from the Δ hp0197 group (day 7 pi). From a pig in the WT group, moderate inflammatory cells were observed in the inner layer of the dura mater (A), along with leptomeningitis with intense inflammation consisting of neutrophils and macrophages (C). No significant findings were observed from the pigs in the Δhp0197 mutant group (B and D). Scale bar = 100 µm.
(JPG)
{kind=link}
2-DE map of whole cellular proteins from the WT (A) and Δ hp0197 (B) strains. Sixteen spots corresponding to 9 proteins were found differentially expressed (Table S3).
(TIF)
Reduced expression levels of genes in Δ hp0197 compared to WT confirmed by microarray analysis.
(DOC)
Increased expression levels of genes in Δ hp0197 compared to WT confirmed by microarrays analysis.
(DOC)
Differential expression levels of proteins in Δ hp0197 compared to WT as identified by 2-DE/MS.
(DOC)
Primer sequences used for this study.
(DOC)
Primer sequences used for qRT-PCR.
(DOC)
Acknowledgments
We are grateful to Daisuke Takamatsu (National Institute of Animal Health and National Agriculture and Food Research Organization) for kindly providing pSET2 and pSET4s vectors. All of the microarray experiments were preformed with the help of the National Engineering Center for Biochip in Shanghai. We thank Yanxiu Liu for her careful revision of the language of this manuscript.
Funding Statement
This work was supported by the National Natural Science Foundation of China (30901076), the 973 program (2012CB518805) and the 863 program (2011AA10A210). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Staats JJ, Feder I, Okwumabua O, Chengappa MM (1997) Streptococcus suis: past and present. Vet Res Commun 21: 381–407. [DOI] [PubMed] [Google Scholar]
- 2. Lun ZR, Wang QP, Chen XG, Li AX, Zhu XQ (2007) Streptococcus suis: an emerging zoonotic pathogen. Lancet Infect Dis 7: 201–209. [DOI] [PubMed] [Google Scholar]
- 3. Hill JE, Gottschalk M, Brousseau R, Harel J, Hemmingsen SM, et al. (2005) Biochemical analysis, cpn60 and 16S rDNA sequence data indicate that Streptococcus suis serotypes 32 and 34, isolated from pigs, are Streptococcus orisratti. Vet Microbiol 107: 63–69. [DOI] [PubMed] [Google Scholar]
- 4. Wertheim HF, Nghia HD, Taylor W, Schultsz C (2009) Streptococcus suis: an emerging human pathogen. Clin Infect Dis 48: 617–625. [DOI] [PubMed] [Google Scholar]
- 5. Tang J, Wang C, Feng Y, Yang W, Song H, et al. (2006) Streptococcal toxic shock syndrome caused by Streptococcus suis serotype 2. PLoS Med 3: e151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Yu H, Jing H, Chen Z, Zheng H, Zhu X, et al. (2006) Human Streptococcus suis outbreak, Sichuan, China. Emerg Infect Dis 12: 914–920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Segura M (2009) Streptococcus suis: An Emerging Human Threat. J Infect Dis 199: 4–6. [DOI] [PubMed] [Google Scholar]
- 8. Holden MT, Hauser H, Sanders M, Ngo TH, Cherevach I, et al. (2009) Rapid evolution of virulence and drug resistance in the emerging zoonotic pathogen Streptococcus suis. PLoS One 4: e6072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Ye C, Zheng H, Zhang J, Jing H, Wang L, et al. (2009) Clinical, Experimental, and Genomic Differences between Intermediately Pathogenic, Highly Pathogenic, and Epidemic Streptococcus suis. J Infect Dis 199: 97–107. [DOI] [PubMed] [Google Scholar]
- 10. Zheng X, Zheng H, Lan R, Ye C, Wang Y, et al. (2011) Identification of Genes and Genomic Islands Correlated with High Pathogenicity in Streptococcus suis Using Whole Genome Tilling Microarrays. PLoS One 6: e17987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Li M, Shen X, Yan J, Han H, Zheng B, et al. (2011) GI-type T4SS-mediated horizontal transfer of the 89K pathogenicity island in epidemic Streptococcus suis serotype 2. Mol Microbiol 79: 1670–1683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Takamatsu D, Wongsawan K, Osaki M, Nishino H, Ishiji T, et al. (2008) Streptococcus suis in humans, Thailand. Emerg Infect Dis 14: 181–183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Wertheim HF, Nguyen HN, Taylor W, Lien TT, Ngo HT, et al. (2009) Streptococcus suis, an important cause of adult bacterial meningitis in northern Vietnam. PLoS One 4: e5973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Mai NT, Hoa NT, Nga TV, Linh le D, Chau TT, et al. (2008) Streptococcus suis meningitis in adults in Vietnam. Clin Infect Dis 46: 659–667. [DOI] [PubMed] [Google Scholar]
- 15. Nghia HD, Hoa NT, Linh le D, Campbell J, Diep TS, et al. (2008) Human case of Streptococcus suis serotype 16 infection. Emerg Infect Dis 14: 155–157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Smith TC, Capuano AW, Boese B, Myers KP, Gray GC (2008) Exposure to Streptococcus suis among US swine workers. Emerg Infect Dis 14: 1925–1927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Gottschalk M, Xu J, Calzas C, Segura M (2010) Streptococcus suis: a new emerging or an old neglected zoonotic pathogen? Future Microbiol 5: 371–391. [DOI] [PubMed] [Google Scholar]
- 18. Chabot-Roy G, Willson P, Segura M, Lacouture S, Gottschalk M (2006) Phagocytosis and killing of Streptococcus suis by porcine neutrophils. Microb Pathog 41: 21–32. [DOI] [PubMed] [Google Scholar]
- 19. Smith HE, Damman M, van der Velde J, Wagenaar F, Wisselink HJ, et al. (1999) Identification and characterization of the cps locus of Streptococcus suis serotype 2: the capsule protects against phagocytosis and is an important virulence factor. Infect Immun 67: 1750–1756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Van Calsteren MR, Gagnon F, Lacouture S, Fittipaldi N, Gottschalk M (2010) Structure determination of Streptococcus suis serotype 2 capsular polysaccharide. Biochem Cell Biol 88: 513–525. [DOI] [PubMed] [Google Scholar]
- 21. Staats JJ, Plattner BL, Stewart GC, Changappa MM (1999) Presence of the Streptococcus suis suilysin gene and expression of MRP and EF correlates with high virulence in Streptococcus suis type 2 isolates. Vet Microbiol 70: 201–211. [DOI] [PubMed] [Google Scholar]
- 22. Brassard J, Gottschalk M, Quessy S (2004) Cloning and purification of the Streptococcus suis serotype 2 glyceraldehyde-3-phosphate dehydrogenase and its involvement as an adhesin. Vet Microbiol 102: 87–94. [DOI] [PubMed] [Google Scholar]
- 23. Feng Y, Pan X, Sun W, Wang C, Zhang H, et al. (2009) Streptococcus suis enolase functions as a protective antigen displayed on the bacterial cell surface. J Infect Dis 200: 1583–1592. [DOI] [PubMed] [Google Scholar]
- 24. Zhang A, Chen B, Mu X, Li R, Zheng P, et al. (2009) Identification and characterization of a novel protective antigen, Enolase of Streptococcus suis serotype 2. Vaccine 27: 1348–1353. [DOI] [PubMed] [Google Scholar]
- 25. de Greeff A, Buys H, Verhaar R, Dijkstra J, van Alphen L, et al. (2002) Contribution of fibronectin-binding protein to pathogenesis of Streptococcus suis serotype 2. Infect Immun 70: 1319–1325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Chen B, Zhang A, Xu Z, Li R, Chen H, et al. (2011) Large-scale identification of bacteria-host crosstalk by affinity chromatography: capturing the interactions of Streptococcus suis proteins with host cells. J Proteome Res 10: 5163–5174. [DOI] [PubMed] [Google Scholar]
- 27. Haataja S, Tikkanen K, Hytonen J, Finne J (1996) The Gal alpha 1–4 Gal-binding adhesin of Streptococcus suis, a gram-positive meningitis-associated bacterium. Adv Exp Med Biol 408: 25–34. [DOI] [PubMed] [Google Scholar]
- 28. Haataja S, Tikkanen K, Liukkonen J, Francois-Gerard C, Finne J (1993) Characterization of a novel bacterial adhesion specificity of Streptococcus suis recognizing blood group P receptor oligosaccharides. J Biol Chem 268: 4311–4317. [PubMed] [Google Scholar]
- 29. Haataja S, Tikkanen K, Nilsson U, Magnusson G, Karlsson KA, et al. (1994) Oligosaccharide-receptor interaction of the Gal alpha 1–4Gal binding adhesin of Streptococcus suis. Combining site architecture and characterization of two variant adhesin specificities. J Biol Chem 269: 27466–27472. [PubMed] [Google Scholar]
- 30. Tikkanen K, Haataja S, Francois-Gerard C, Finne J (1995) Purification of a galactosyl-alpha 1–4-galactose-binding adhesin from the gram-positive meningitis-associated bacterium Streptococcus suis. J Biol Chem 270: 28874–28878. [DOI] [PubMed] [Google Scholar]
- 31. Tikkanen K, Haataja S, Finne J (1996) The galactosyl-(alpha 1–4)-galactose-binding adhesin of Streptococcus suis: occurrence in strains of different hemagglutination activities and induction of opsonic antibodies. Infect Immun 64: 3659–3665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Kouki A, Haataja S, Loimaranta V, Pulliainen AT, Nilsson UJ, et al. (2011) Identification of a novel streptococcal adhesin P (SadP) recognizing galactosyl-{alpha}1–4-galactose-containing glycoconjugates: convergent evolution of bacterial pathogens to binding of the same host receptor. J Biol Chem 286: 38854–38864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Baums CG, Kaim U, Fulde M, Ramachandran G, Goethe R, et al. (2006) Identification of a novel virulence determinant with serum opacification activity in Streptococcus suis. Infect Immun 74: 6154–6162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Fittipaldi N, Sekizaki T, Takamatsu D, Harel J, Dominguez-Punaro Mde L, et al. (2008) D-alanylation of lipoteichoic acid contributes to the virulence of Streptococcus suis. Infect Immun 76: 3587–3594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Fittipaldi N, Sekizaki T, Takamatsu D, Dominguez-Punaro Mde L, Harel J, et al. (2008) Significant contribution of the pgdA gene to the virulence of Streptococcus suis. Mol Microbiol 70: 1120–1135. [DOI] [PubMed] [Google Scholar]
- 36. Zhang A, Mu X, Chen B, Liu C, Han L, et al. (2010) Identification and characterization of IgA1 protease from Streptococcus suis. Vet Microbiol 140: 171–175. [DOI] [PubMed] [Google Scholar]
- 37. Zhang A, Mu X, Chen B, Han L, Chen H, et al. (2011) IgA1 protease contributes to the virulence of Streptococcus suis. Vet Microbiol 148: 436–439. [DOI] [PubMed] [Google Scholar]
- 38. Schreur PJ, Rebel JM, Smits MA, van Putten JP, Smith HE (2011) TroA of Streptococcus suis Is Required for Manganese Acquisition and Full Virulence. J Bacteriol 193: 5073–5080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Tang Y, Zhang X, Wu W, Lu Z, Fang W (2012) Inactivation of the sodA gene of Streptococcus suis type 2 encoding superoxide dismutase leads to reduced virulence to mice. Vet Microbiol. [DOI] [PubMed]
- 40.Pian Y, Gan S, Wang S, Guo J, Wang P, et al.. (2012) Ssfhb, a novel factor H-binding surface protein, contributing to the anti-phagocytic ability and virulence of Streptococcus suis. Infect Immun. [DOI] [PMC free article] [PubMed]
- 41. Hu Q, Liu P, Yu Z, Zhao G, Li J, et al. (2010) Identification of a cell wall-associated subtilisin-like serine protease involved in the pathogenesis of Streptococcus suis serotype 2. Microb Pathog 48: 103–109. [DOI] [PubMed] [Google Scholar]
- 42. Bonifait L, Dominguez-Punaro MD, Vaillancourt K, Bart C, Slater J, et al. (2010) The cell envelope subtilisin-like proteinase is a virulence determinant for Streptococcus suis. BMC Microbiol 10: 42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Bonifait L, Vaillancourt K, Gottschalk M, Frenette M, Grenier D (2011) Purification and characterization of the subtilisin-like protease of Streptococcus suis that contributes to its virulence. Vet Microbiol 148: 333–340. [DOI] [PubMed] [Google Scholar]
- 44. Li M, Wang C, Feng Y, Pan X, Cheng G, et al. (2008) SalK/SalR, a two-component signal transduction system, is essential for full virulence of highly invasive Streptococcus suis serotype 2. PLoS ONE 3: e2080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Pan X, Ge J, Li M, Wu B, Wang C, et al. (2009) The orphan response regulator CovR: a globally negative modulator of virulence in Streptococcus suis serotype 2. J Bacteriol 191: 2601–2612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Rodriguez-Ortega MJ, Luque I, Tarradas C, Barcena JA (2008) Overcoming function annotation errors in the Gram-positive pathogen Streptococcus suis by a proteomics-driven approach. BMC Genomics 9: 588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Chen C, Tang J, Dong W, Wang C, Feng Y, et al. (2007) A glimpse of streptococcal toxic shock syndrome from comparative genomics of S. suis 2 Chinese isolates. PLoS ONE 2: e315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Zhang A, Xie C, Chen H, Jin M (2008) Identification of immunogenic cell wall-associated proteins of Streptococcus suis serotype 2. Proteomics 8: 3506–3515. [DOI] [PubMed] [Google Scholar]
- 49. Jing HB, Yuan J, Wang J, Yuan Y, Zhu L, et al. (2008) Proteome analysis of Streptococcus suis serotype 2. Proteomics 8: 333–349. [DOI] [PubMed] [Google Scholar]
- 50. Geng H, Zhu L, Yuan Y, Zhang W, Li W, et al. (2008) Identification and characterization of novel immunogenic proteins of Streptococcus suis serotype 2. J Proteome Res 7: 4132–4142. [DOI] [PubMed] [Google Scholar]
- 51. Zhang A, Chen B, Li R, Mu X, Han L, et al. (2009) Identification of a surface protective antigen, HP0197 of Streptococcus suis serotype 2. Vaccine 27: 5209–5213. [DOI] [PubMed] [Google Scholar]
- 52. Ruggiero A, Tizzano B, Pedone E, Pedone C, Wilmanns M, et al. (2009) Crystal structure of the resuscitation-promoting factor (DeltaDUF)RpfB from M. tuberculosis. J Mol Biol 385: 153–162. [DOI] [PubMed] [Google Scholar]
- 53. Benga L, Fulde M, Neis C, Goethe R, Valentin-Weigand P (2008) Polysaccharide capsule and suilysin contribute to extracellular survival of Streptococcus suis co-cultivated with primary porcine phagocytes. Vet Microbiol 132: 211–219. [DOI] [PubMed] [Google Scholar]
- 54. Segura M, Gottschalk M, Olivier M (2004) Encapsulated Streptococcus suis inhibits activation of signaling pathways involved in phagocytosis. Infect Immun 72: 5322–5330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Lecours MP, Gottschalk M, Houde M, Lemire P, Fittipaldi N, et al. (2011) Critical Role for Streptococcus suis Cell Wall Modifications and Suilysin in Resistance to Complement-Dependent Killing by Dendritic Cells. J Infect Dis 204: 919–929. [DOI] [PubMed] [Google Scholar]
- 56. Shelburne SA 3rd, Keith D, Horstmann N, Sumby P, Davenport MT, et al (2008) A direct link between carbohydrate utilization and virulence in the major human pathogen group A Streptococcus. Proc Natl Acad Sci U S A 105: 1698–1703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Willenborg J, Fulde M, de Greeff A, Rohde M, Smith HE, et al. (2011) Role of glucose and CcpA in capsule expression and virulenceof Streptococcus suis. Microbiology 157: 1823–1833. [DOI] [PubMed] [Google Scholar]
- 58. Sonenshein AL (2007) Control of key metabolic intersections in Bacillus subtilis. Nat Rev Microbiol 5: 917–927. [DOI] [PubMed] [Google Scholar]
- 59. Schumacher MA, Allen GS, Diel M, Seidel G, Hillen W, et al. (2004) Structural basis for allosteric control of the transcription regulator CcpA by the phosphoprotein HPr-Ser46-P. Cell 118: 731–741. [DOI] [PubMed] [Google Scholar]
- 60. Feng Y, Zhang H, Ma Y, Gao GF (2010) Uncovering newly emerging variants of Streptococcus suis, an important zoonotic agent. Trends Microbiol 18: 124–131. [DOI] [PubMed] [Google Scholar]
- 61. Mandanici F, Gomez-Gascon L, Garibaldi M, Olaya-Abril A, Luque I, et al. (2010) A surface protein of Streptococcus suis serotype 2 identified by proteomics protects mice against infection. J Proteomics 73: 2365–2369. [DOI] [PubMed] [Google Scholar]
- 62. Chen B, Zhang A, Li R, Mu X, He H, et al. (2010) Evaluation of the protective efficacy of a newly identified immunogenic protein, HP0272, of Streptococcus suis. FEMS Microbiol Lett 307: 12–18. [DOI] [PubMed] [Google Scholar]
- 63. Li R, Zhang A, Chen B, Teng L, Wang Y, et al. (2010) Response of swine spleen to Streptococcus suis infection revealed by transcription analysis. BMC Genomics 11: 556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Dominguez-Punaro Mde L, Segura M, Radzioch D, Rivest S, Gottschalk M (2008) Comparison of the susceptibilities of C57BL/6 and A/J mouse strains to Streptococcus suis serotype 2 infection. Infect Immun 76: 3901–3910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Takamatsu D, Osaki M, Sekizaki T (2001) Construction and characterization of Streptococcus suis-Escherichia coli shuttle cloning vectors. Plasmid 45: 101–113. [DOI] [PubMed] [Google Scholar]
- 66. Dominguez-Punaro MC, Segura M, Plante MM, Lacouture S, Rivest S, et al. (2007) Streptococcus suis serotype 2, an important swine and human pathogen, induces strong systemic and cerebral inflammatory responses in a mouse model of infection. J Immunol 179: 1842–1854. [DOI] [PubMed] [Google Scholar]
- 67. Pallares FJ, Schmitt CS, Roth JA, Evans RB, Kinyon JM, et al. (2004) Evaluation of a ceftiofur-washed whole cell Streptococcus suis bacterin in pigs. Can J Vet Res 68: 236–240. [PMC free article] [PubMed] [Google Scholar]
- 68. Li Y, Gottschalk M, Esgleas M, Lacouture S, Dubreuil JD, et al. (2007) Immunization with recombinant Sao protein confers protection against Streptococcus suis infection. Clin Vaccine Immunol 14: 937–943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Luo Y, Dorf ME (2001) Isolation of mouse neutrophils. Curr Protoc Immunol Chapter 3: Unit 3 20. [DOI] [PubMed]
- 70. Benga L, Goethe R, Rohde M, Valentin-Weigand P (2004) Non-encapsulated strains reveal novel insights in invasion and survival of Streptococcus suis in epithelial cells. Cell Microbiol 6: 867–881. [DOI] [PubMed] [Google Scholar]
- 71. Schmittgen TD, Zakrajsek BA (2000) Effect of experimental treatment on housekeeping gene expression: validation by real-time, quantitative RT-PCR. J Biochem Biophys Methods 46: 69–81. [DOI] [PubMed] [Google Scholar]
- 72. Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25: 402–408. [DOI] [PubMed] [Google Scholar]
- 73. Hellman LM, Fried MG (2007) Electrophoretic mobility shift assay (EMSA) for detecting protein-nucleic acid interactions. Nat Protoc 2: 1849–1861. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Relevant histopathological findings could be observed from WT-infected pigs (day 3 pi) while no significant histopathological findings could be observed from a representative pig from the Δ hp0197 group (day 7 pi). From a pig in the WT group, moderate inflammatory cells were observed in the inner layer of the dura mater (A), along with leptomeningitis with intense inflammation consisting of neutrophils and macrophages (C). No significant findings were observed from the pigs in the Δhp0197 mutant group (B and D). Scale bar = 100 µm.
(JPG)
2-DE map of whole cellular proteins from the WT (A) and Δ hp0197 (B) strains. Sixteen spots corresponding to 9 proteins were found differentially expressed (Table S3).
(TIF)
Reduced expression levels of genes in Δ hp0197 compared to WT confirmed by microarray analysis.
(DOC)
Increased expression levels of genes in Δ hp0197 compared to WT confirmed by microarrays analysis.
(DOC)
Differential expression levels of proteins in Δ hp0197 compared to WT as identified by 2-DE/MS.
(DOC)
Primer sequences used for this study.
(DOC)
Primer sequences used for qRT-PCR.
(DOC)