

Abstract
Partner notification (PN or contact tracing) is an important aspect of treating bacterial sexually transmitted infections (STIs), such as Chlamydia trachomatis. It facilitates the identification of new infected cases that can be treated through individual case management. PN also acts indirectly by limiting onward transmission in the general population. However, the impact of PN, both at the level of individuals and the population, remains unclear. Since it is difficult to study the effects of PN empirically, mathematical and computational models are useful tools for investigating its potential as a public health intervention. To this end, we developed an individual-based modeling framework called Rstisim. It allows the implementation of different models of STI transmission with various levels of complexity and the reconstruction of the complete dynamic sexual partnership network over any time period. A key feature of this framework is that we can trace an individual’s partnership history in detail and investigate the outcome of different PN strategies for C. trachomatis. For individual case management, the results suggest that notifying three or more partners from the preceding 18 months yields substantial numbers of new cases. In contrast, the successful treatment of current partners is most important for preventing re-infection of index cases and reducing further transmission of C. trachomatis at the population level. The findings of this study demonstrate the difference between individual and population level outcomes of public health interventions for STIs.
Introduction
Partner notification (PN, also known as contact tracing) is an integral part of managing several sexually transmitted infections (STIs). The process of PN for curable STIs includes informing sexual partners of infected people of their exposure, administering presumptive treatment and providing advice about the prevention of future infection [1]. PN has multiple objectives and operates at both individual and population levels [2]. One objective is the identification of new infected index cases who can be treated through individual case management. Another objective is to reduce infection prevalence by preventing onward transmission in the population. While PN is often described as an effective control intervention for different STIs, the relative effects at the level of individuals and the population are not well understood.
PN is widely used for Chlamydia trachomatis infections. C. trachomatis is the most common bacterial STI in many developed countries and is primarily found among sexually active young adults [3]. The majority of infections is asymptomatic and remains undiagnosed. Treatment and prevention of C. trachomatis are of particular importance to women since infection can lead to serious reproductive tract complications [4]. The importance of notifying current partners of infected index cases has been illustrated in both reviews of empirical studies [5] and modeling studies [6], [7] where PN has been shown to reduce the probability of re-infection of index cases. Notification of previous partners of index cases is also recommended, particularly as part of screening programs that aim to limit transmission and to reduce the overall prevalence of C. trachomatis in the population [8]. If PN is applied as an integral part of a screening intervention, it is expected to result in a more substantial reduction in the prevalence of C. trachomatis than would be expected by screening alone [9], [10]. However, we are not aware of any empirical studies that have reported the effect of PN on the population prevalence of C. trachomatis.
Several countries recommend different PN look-back periods because there is still uncertainty about the most appropriate strategy. For example, the US Centers for Disease Control and Prevention (CDC) recommends notifying partners with whom the index case has had sexual contact within the previous 60 days [11]. If no sexual contact occurred during this period, the most recent partner should be notified. Zimmermann-Rogers et al. [12] had previously pointed out that a PN period of 180 days or more would help to identify more infected cases. The UK National Guideline for the Management of Genital Tract Infection with C. trachomatis recommends notifying partners of an asymptomatic index case within a period of 6 months [13]. The same is standard in Sweden, but recommendations from the National Board of Health and Welfare might change, based on a recent study that found that extending PN periods could improve the identification of new C. trachomatis cases [14].
Mathematical and computational models for detailed examination of the effects of PN on the identification of new index cases and the reduction in onward transmission need to allow partnerships to be represented explicitly and an individual’s partnership history to be documented beyond the current partner. The impact of PN has been investigated as a general concept [15], [16] and for specific bacterial STIs such as Neisseria gonorrhoeae and C. trachomatis [9], [10], [17], [18]. These studies did not consider a dynamic sexual partnership network and/or did not follow the partnership history over a prolonged look-back period, however. Complex PN strategies that involve tracing both current and previous partners of index cases need to be investigated using stochastic, individual-based models. This structure allows partnerships to be represented explicitly, a dynamic sexual partnership network to be reconstructed over any given time period, and an individual’s partnership history beyond the current partner to be kept on record [19]. A disadvantage of existing individual-based models is that they were usually designed to address one specific research question [20]–[22] and can often not be readily adapted to different situations. In contrast, deterministic population-based models, which are based on ordinary differential equations (ODEs), offer great flexibility in altering model assumptions and for model parameterization. These models, however, cannot track individuals so their potential for studying PN interventions is limited. A modelling framework that combines the tractability of ODE models with the properties of individual-based models would be a powerful tool for examining the impact of different assumptions about model structure on the effects of PN. The importance of comparing different models and assumptions has previously been shown for chlamydia screening [8] and human papillomavirus vaccination [23].
In this paper, we present a novel, stochastic, individual-based modeling framework called Rstisim (from R STI Simulator), which allows the implementation in an individual-based manner of models of STI transmission that are described by ODEs. We apply and compare three basic models of C. trachomatis transmission with different assumptions about the sexual partnership dynamics. Based on these models, we investigate some general properties of PN for C. trachomatis. Two strategies of PN are considered: one in which partners can be notified in order of their recency, and one in which partners within a certain time period can be notified. Together, they allow us to draw some general conclusions about the effects of PN for C. trachomatis at both the level of individuals and the population level.
Results
Modeling Sexual Contacts
We first derive the deterministic, population-based descriptions (ODEs) of three different models that describe heterosexual partnership dynamics with increasing levels of complexity (see Methods). These models are then implemented at an individual level in Rstisim, which allows the sexual partnership network over different time periods to be reconstructed. The instantaneous contact model is based on the assumption that sexual contacts happen instantaneously [24], but this model cannot account for a sexual partnership network at cross-section. In contrast, the pair model assumes that women and men form partnerships that last for a certain period [25]. Sex acts that might lead to transmission of the infection occur throughout the duration of the partnership. The triple model accounts for the fact that individuals can have two sexual partnerships at the same time (concurrency). Despite the simple nature of the models, they exhibit rich dynamics, particularly in the case of the triple model, where chains of contacts can occur at cross-section (Fig. 1). In all models, closely connected groups or bigger circular structures emerge over a period of one year.
Figure 1. Sexual partnership networks from the three individual-based models.

The ‘instantaneous contact’ model has no connectivity at cross-section but exhibits variation and connected components of different size within a period of one year. The ‘pair model’ and ‘triple model’ illustrate the connectivity at cross-section and larger connected components during a period of one year. Note that in all three models, the average number of new partnerships formed within one year is equal. For illustrative purposes, the population size was limited to 100, resulting in higher connected networks compared to larger population sizes. Different sexes are indicated by filled and empty circles.
Number of Sex Partners and Chlamydia Transmission
The models were parameterized to reflect the sexual behavior of the general population of young adults by using data from Britain’s second National Survey of Sexual Attitudes and Lifestyles (Natsal-2) for 16–25 year old women and men [26]. All three models are adjusted so that the total number of new heterosexual contacts or partnerships and the total number of realized sex acts are equal (Table 1). For the triple model, we define the level of concurrency,  , as the ratio of individuals that have two partnerships to all individuals in a partnership. By varying the level of concurrency between 0–100% we found that
, as the ratio of individuals that have two partnerships to all individuals in a partnership. By varying the level of concurrency between 0–100% we found that 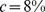 provides the best match between the simulations and data from Natsal-2 describing the gaps and overlaps between sexual partnerships (Fig. S1). Assuming 8% concurrency at cross-section in the triple model results in a cumulative incidence of concurrency of
provides the best match between the simulations and data from Natsal-2 describing the gaps and overlaps between sexual partnerships (Fig. S1). Assuming 8% concurrency at cross-section in the triple model results in a cumulative incidence of concurrency of  15% over the last year. This is in line with the Natsal-2 estimate for 16–24 year old women (15.2%, 95% confidence interval (CI) 12.7, 18.1%) but slightly lower than for men (20.8%, 95% CI 17.8, 24.3%) [26]. The per sex act transmission probabilities of C. trachomatis were calibrated to obtain an endemic prevalence of 3% and are in good agreement with empirical estimates [27]–[29]. The transmission probabilities decrease with increasing complexity of the models (Table 1) since both re-infection and concurrency facilitate the spread of STIs [6], [30].
15% over the last year. This is in line with the Natsal-2 estimate for 16–24 year old women (15.2%, 95% confidence interval (CI) 12.7, 18.1%) but slightly lower than for men (20.8%, 95% CI 17.8, 24.3%) [26]. The per sex act transmission probabilities of C. trachomatis were calibrated to obtain an endemic prevalence of 3% and are in good agreement with empirical estimates [27]–[29]. The transmission probabilities decrease with increasing complexity of the models (Table 1) since both re-infection and concurrency facilitate the spread of STIs [6], [30].
Table 1. Parameters that determine the dynamics of sexual partnerships and the transmission of C. trachomatis.
| Instantaneous | Pair | Triple | |
| contact model | model | model | |
| Assumed parameters | |||
| Mean number of new heterosexual partnershipsper individual [26] | 1.04 y−1 | 1.04 y−1 | 1.04 y−1 |
| Mean number of total heterosexual partnershipsper individual [26] | – | 1.70 y−1 | 1.70 y−1 |
| Level of concurrency, c | – | – | 8% |
| Proportion of individuals that are in a partnershipat cross-section | 0% | 67% | 62% |
| Frequency of sex acts, f | – | 1 per week | 1 per week |
Mean duration of infection, 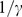 [6], [39]
[6], [39]
|
1 y | 1 y | 1 y |
Prevalence of C. trachomatis, 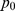 [56]
[56]
|
3% | 3% | 3% |
| Derived parameters | |||
| Contact or pair formation rate, ρ | 0.52 y−1 | 1.56 y−1 | 1.36 y−1 |
| Relative probability of accepting a partnershipif already in a pair, α | – | – | 0.28 |
| Mean duration of partnership, 1/σ | – | 0.65 y | 0.65 y |
| Average number of sex acts per partnership, n | 35 | 35 | 35 |
| Transmission probability per sex act, π | 15% | 11% | 9% |
The different models (instantaneous contact, pair and triple model) are adjusted so that the total number of newly formed contacts or partnerships and the total number of realized sex acts are equal. To achieve the same steady-state prevalence of C. trachomatis, the transmission probability is varied between the models while keeping the mean duration of the infection the same. This ensures that the incidence of C. trachomatis infection is equal in all models. The level of concurrency at cross-section is defined as the ratio of individuals that have more than one partnership to all individuals in a partnership. Some parameters are given as rounded values.
Partner Notification Strategies
The key reason to implement the models in our individual-based modeling framework is to be able to follow an individual’s history of current and previous partners (Fig. 2). The instantaneous contact model and the pair model describe serial monogamy. The triple model illustrates complex partnership dynamics in which a new partnership can replace a previous one, or where short episodes of concurrency can occur.
Figure 2. Partner notification strategies.

An illustrative example of an individual’s history of contacts or partnerships is shown for each model. One strategy is to notify partners of an index case in order of their recency (time since their partnership ended). Another strategy is to notify all partners from a certain time period.
Individual level effect of partner notification
At the level of individuals, PN for C. trachomatis identifies new index cases. It is therefore important to know how many partners of an index case are infected with C. trachomatis. A Swedish study found that the proportion of C. trachomatis-positive partners of an index case decreases with increasing duration since they last had sexual intercourse [14]. In the simulations, everyone who is C. trachomatis-positive at cross-section is defined as an index case. This corresponds to infected individuals that would be detected through random screening. We can now go through all current and previous partners of an index case and ‘test’ whether or not they are infected. Using the same time periods as in the study by Carré et al. [14], all three models exhibit a similar C. trachomatis-positivity of partners with the pair model being within the 95% CI of the data (Fig. 3).
Figure 3. Simulated and empirical data of C. trachomatis-positivity in partners of index cases.

Black crosses correspond to published data of the proportion of positive partners out of those with a positive test results together with the 95% CI [14]. The other symbols represent simulated data for each of the three different models. In the simulations, it is assumed that the steady-state prevalence of C. trachomatis is 3%. Means of 100 simulation runs are shown. Standard errors are small and omitted for better visibility.
This behavior can be examined in more detail with Rstisim. First, we investigate in order of recency the proportion of partners who are infected (Fig. 4A). The models give consistent results with 67–75% of the most recent partners of an index case being infected with C. trachomatis. The less recent a partnership or contact, the lower the probability that the partner is infected. The proportion of infected individuals up to the third most recent partner is still substantially higher than the population prevalence.
Figure 4. Proportion of C. trachomatis-positive partners of index cases.

The proportion of partners of an index case who are infected with C. trachomatis is shown at a steady-state prevalence of 3% (dashed line). (A) The proportion of infected partners in order of their recency. (B) The proportion of infected partners in order of their breakup date. For each strategy, means of 100 simulation runs are shown. Standard errors are small and omitted for better visibility.
We can also group the partners of an index case by the time period since the partnership has ended in greater detail than in the empirical study by Carré et al. [14] (Fig. 4B). The results of the pair and triple models differ from those of the instantaneous contact model. The instantaneous contact model does not have current partners by definition and the pair and triple model result in a lower proportion of infected partners whose partnership has ended within the last year. First, this can be explained through the high per contact transmission probability in the instantaneous contact model. Second, transmission in the pair and triple model can occur before the partnership ends. This makes it possible for the partners to have cleared the infection by the time they will be notified. In all three models, as far back as 18 months, a substantial proportion of partners (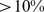 ) are infected with C. trachomatis. This shows that extending PN periods beyond one year yields more new index cases for individual case management than would be found through random screening.
) are infected with C. trachomatis. This shows that extending PN periods beyond one year yields more new index cases for individual case management than would be found through random screening.
Population level effect of partner notification
At the population level, PN can prevent onward transmission of C. trachomatis and reduce the overall prevalence of the infection. Here, we investigate the effects of the different PN strategies if they are implemented as part of a population-wide screening program. After the simulations approach the steady-state prevalence of 3%, we introduce random screening of the whole population of young adults. Every woman and man receives screening at a rate of 0.1 per year, i.e., every 10 years on average. If PN is performed, each notified partner will be tested and successfully treated with a probability of 50% [31], [32].
The simulations show that screening reduces the prevalence of C. trachomatis (Fig. 5). After 5 years of screening, assuming that there is no PN, the prevalence is reduced to about 70% of the baseline in the pair and triple model and 60% in the instantaneous contact model (Fig. 6). The effect of screening is smaller in the pair and triple model because index cases in ongoing partnerships engage in sex acts with the same untreated partner after treatment and can be re-infected [6]. This cannot occur in the instantaneous contact model. If PN is performed for at least the most recent partner, the prevalence is reduced to below 60% of the baseline prevalence (Fig. 6A). Increasing the PN period in the instantaneous contact model results in a slight but steady decrease in prevalence (Fig. 6B). However, in the pair and triple model the strongest effect of PN stems from notifying the current partner only. Thus, under more realistic assumptions of sexual partnership dynamics, notification of current partners is sufficient to achieve most of the additional reduction in prevalence at the population level.
Figure 5. Representative time plots of the prevalence of C. trachomatis after the start of a screening intervention.

The dots indicate the prevalence at the beginning (dotted line) and after 5 years of the screening intervention. The dashed line indicates the steady-state prevalence of 3% in absence of screening. Every individual receives screening at a rate of 0.1 per year and no partner notification is performed. The colored lines represent five individual simulation runs.
Figure 6. Population level effect of partner notification.

The reduction in the prevalence of C. trachomatis is given after screening the population for 5 years at a rate of 0.1 per year. (A) The prevalence of C. trachomatis for increasing numbers of notified partners, in order of their recency. (B) The prevalence of C. trachomatis for different partner notification periods. There is a 50% probability that each notified partner will be tested and successfully treated. For each strategy, means of 100 simulation runs are shown. Standard errors are small and omitted for better visibility.
Sensitivity Analyses
Heterogeneity in sexual behavior
We have presented results from three models that assume a homogeneous and closed heterosexual population of young adults. The assumption of homogenous mixing provides a good description of the C. trachomatis-positivity in partners of index cases (Fig. 3, and refs. [33], [34]). Nevertheless, C. trachomatis transmission can be influenced by heterogeneity in sexual behavior, even within a narrow age group. We therefore developed a model where we stratify the population of 16–25 year old women and men into two risk classes (see Fig. S2 in Text S1 and S3 in Text S1). The average number of new heterosexual partners per year is the same as in the models that assume homogeneous mixing. The results from the model with risk classes are consistent with the conclusions drawn from the simpler models. The C. trachomatis-positivity in partners from more than 18 months ago is low (Fig. S4B in Text S1). Together, this underlines that notifying three or more partners from the last 18 months can be helpful in finding new index cases. Compared to the models that assume homogeneous mixing, the proportion of previous partners of index cases that are infected with C. trachomatis is lower for the most recent partner but higher for the third and subsequent partners (Fig. S4A in Text S1). The population level effect of PN primarily stems from notifying the most recent partner as for the homogeneous mixing models (Fig. S5A in Text S1).
Sex-specific differences in duration of infection
The baseline models assume the same disease parameters for women and men. The infectious duration in men is not well established but it has been suggested that it is shorter than in women [35]. We therefore investigate the scenario where the infectious duration in men is half that of in women. We used the pair model for the sensitivity analysis as it incorporates sexual partnerships explicitly while being more tractable than the triple model. Simulations show that our overall conclusions on the effect of PN remain the same (Fig. S6 and S7 in Text S1).
Differences in uptake of screening and partner notification
The uptake of screening and successful PN are influenced by public health interventions in different settings. The baseline scenario with a screening rate of 0.1 per year is conservative [36], but our simulations show that this can result in a substantial decrease in prevalence if men are screened as well. For the sensitivity analysis, we used again the pair model to investigate the effect of higher screening rates, screening targeted to women only and different probabilities of successful PN (Fig. S8 and S9 in Text S1). Higher screening rates and higher probabilities of successful PN result in a more substantial decrease in the prevalence of C. trachomatis. The general conclusions about the effects of PN are not altered.
Discussion
We studied the effects of PN for C. trachomatis, both at the level of individuals and the population, using a novel, individual-based modeling framework. The model simulations suggest that, while extending PN periods beyond one year helps to find new index cases, most of the additional effect that PN has on reducing transmission in a general heterosexual population of young adults is achieved by notifying the current or most recent partner.
To our knowledge, this is the first C. trachomatis modeling study that investigates the effects of PN of previous partners for realistic look-back periods commonly applied by health care providers. To this end, we developed the modeling framework Rstisim with which one can reconstruct the entire sexual partnership network over any given time period. There are other publicly available software packages that offer some of the features of Rstisim. For example, STI transmission can be simulated using the R package statnet, which generates sexual contact networks based on exponential random graph models [37]. Rstisim allows implementation in an individual-based manner of both simple models formulated by ODEs and more sophisticated models where, for example, the behavior of an individual depends on the previous sexual history [38]. Due to this flexibility, we were able to alter and test different assumptions about sexual partnership dynamics. We used three different models in this study, which shows that taking partnerships explicitly into account is a necessary complexity to study the effects of the two PN strategies. The distinction between current and previous partners cannot be made in a model where contacts are assumed to happen instantaneously, but was necessary for understanding the effect of PN on limiting onward transmission in the population. The additional assumption of concurrency (triple model) did not affect our results in this model of a general heterosexual population of young adults.
Our goal was to investigate some general properties of PN for C. trachomatis in three basic models that have different assumptions about the sexual partnership dynamics. The models were kept deliberately simple so they can be directly compared and adjusted to exhibit the same numbers of partners and sex acts, capturing the partner change rates that are observed in population-based studies. We thus made some simplifying assumptions. First, we treat the population of 16–25 year olds as a closed population that mixes homogeneously. This assumption probably resulted in realistic dynamics in our study because it was restricted to the age group that drives chlamydia transmission in the wider population. Nevertheless, heterogeneity in sexual behavior can influence the transmission of STIs but the overall findings from a model with two levels of sexual activity in the sensitivity analysis (Text S1) were consistent with the simpler models. Second, we did not account for differences in sexual behavior between women and men. We acknowledge that the distribution of women and men between risk groups could differ and men report concurrent partnerships more often than women. Third, we assumed the frequency of sex acts per partnership to be independent of the number of partners in the model with concurrency. We expect that a lower frequency of sex acts per partner for individuals with two partnerships would have a minor effect on the results since the assumption of concurrency did not strongly affect our results in the first place. Fourth, there could also be sex-specific differences in the infection parameters. Our sensitivity analysis showed similar results for the pair model when the infectious duration in men was assumed to be half that in women. The average infectious duration of C. trachomatis in men is not well-established, however, and more reliable estimates are needed. Lastly, we did not consider the possibility of temporal immunity against C. trachomatis infection after natural clearance. Modeling studies have shown that long lasting immunity can reduce the effect of treatment through screening or PN [6], [39] and even result in a rebound in prevalence [40]. It remains to be determined, however, whether the duration of immunity against C. trachomatis infection is long enough to cause these effects [41].
Mathematical and computational models of STI transmission have become increasingly complex over the past two decades. It is possible to incorporate detailed descriptions of sexual behavior and infection characteristics in such models, but this can result in widely different predictions of model outcomes [8], [42]. Such differences can arise because, as model complexity increases, it becomes more difficult to obtain the reliable data for model parameterization. The choice of model complexity should be in balance between incorporating necessary features while keeping the model tractable [43], [44]. Simpler models remain very powerful for studying the impact of public health interventions against STIs [45]. For C. trachomatis, simple models can give a good description of the transmission dynamics [39] and provide conclusions in line with more detailed models that include age and risk stratification [8], [46]. In this study, we focused on a quantitative description of the sexual partnership dynamics between young adults, and the pair and triple model appear to describe the observed durations of partnerships and the gaps between them remarkably well.
Our simulation study illustrates that the choice of PN strategy for C. trachomatis in a general heterosexual population of young adults depends on the public health context in which it is applied. At the individual level, our results suggest that tracing as many as three partners from the preceding 18 months can be helpful in finding new index cases. This is in line with findings from Sweden [14]. Second, at the population level, PN of current partners of an index case should be prioritized: notifying previous partners has little effect on limiting onward transmission because previous partners of an index case are, if C. trachomatis-positive, likely to have been infected for a long time. Hence, there is a high probability that they have already transmitted the infection to other people before they cleared the infection spontaneously. In a scenario where screening is targeted towards high-risk individuals, it is likely that notifying previous partners of index cases would have a stronger effect on limiting onward transmission due to higher partner change rates among these individuals [47].
There are only a few modeling studies that have specifically investigated the effects of PN for C. trachomatis. Our findings confirm those of Kretzschmar et al. [9], [10], who also consider a dynamic sexual partnership network but restrict PN to current partners only. PN can render a screening program more effective, especially if screening rates are high (see Text S1). Armbruster & Brandeau [18] found that increasing contact tracing capacity results in a substantial reduction of C. trachomatis prevalence, although with diminishing returns. They investigate the effects of PN for index cases who seek treatment for symptoms and assume a static sexual network with a relatively high prevalence of C. trachomatis. In our study with a dynamic sexual network, previous partners of an index case do not contribute to re-infection which might explain why our study found that notifying previous partners had little additional effect on reducing C. trachomatis transmission.
There are several open questions that might be addressed in future studies. As noted above, one could consider targeted screening scenarios towards groups with frequent partner change rates, which might result in different PN recommendations for different groups. It would also be interesting to study the effects of different PN strategies for other bacterial STIs, such as N. gonorrhoeae and syphilis. The Rstisim framework will allow researchers to investigate these and other questions in sexual partnership networks with different levels of complexity. This will ultimately lead to a better understanding of the full potential of PN and the identification of the optimal strategies to deal with C. trachomatis and other STIs. In this study, we have shown that PN for C. trachomatis can have different effects at the level of individuals and at the population.
Methods
Individual-based Modeling Framework
Rstisim is a stochastic, individual-based (or agent-based) modeling framework that can simulate the transmission of an arbitrary STI in a sexual partnership network of any level of complexity (see Text S2 for a brief description). It is written in C++ and can be downloaded at http://www.stat.nus.edu.sg/~staar/rstisim as a package for the R software environment for statistical computing [48]. A key concept of the partnership formation in Rstisim is that every individual can be assigned a contact or pair formation rate at which they will initiate a partnership. Every individual can also accept a partnership that was initiated by another individual. This offers various ways to implement partnership formation rules. For example, it provides the flexibility to implement classical models of STI transmission dynamics that are formulated by ODEs [24], as well as more sophisticated models where contact rates depend on the individual’s previous sexual partnership history. At any point in time, all information on individuals, partnerships and infections can be accessed and the sexual contact network can be graphically depicted using the network package [49].
Modeling C. trachomatis Transmission
To investigate the transmission of C. trachomatis in a heterosexual partnership network we implemented three different models in Rstisim: an ‘instantaneous contact model’, a ‘pair model’ and a ‘triple model’ where individuals can have concurrent partnerships. We keep the models deliberately simple (assuming homogeneous mixing), which facilitates the derivation of exact parameter values and allows for a direct comparison of the different assumptions about sexual partnership dynamics. Note that Rstisim also allows sexual contacts between men, or between women. We do not consider them for simplicity. Here, we provide the ODEs of the contact and partnership dynamics (and if applicable the transmission dynamics) for all three models. The structure of these models is then directly implemented into Rstisim where event times (partnership formation and dissolution, sex acts and clearance of C. trachomatis) are drawn from exponential distributions around mean values (Table 1). We assume a total population size 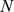 that is equally divided into females and males. The difference in sex is indicated by the subscripts 0 and 1.
that is equally divided into females and males. The difference in sex is indicated by the subscripts 0 and 1.
Instantaneous contact model
Since contacts are assumed to happen instantaneously, individuals always remain single and are denoted by 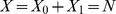 . Assuming an SIS-type (susceptible-infected-susceptible) infection [24], the transmission dynamics can be written as follows:
. Assuming an SIS-type (susceptible-infected-susceptible) infection [24], the transmission dynamics can be written as follows:
| (1) |
| (2) |
The superscripts 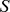 and
and 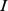 indicate whether individuals are susceptible or infected. The rate at which people have contacts is denoted by
indicate whether individuals are susceptible or infected. The rate at which people have contacts is denoted by 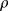 and the transmission probability per contact is given by
and the transmission probability per contact is given by 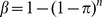 where
where  is the transmission probability per sex act and
is the transmission probability per sex act and  is the number of sex acts per contact. The average duration of an infection is given by
is the number of sex acts per contact. The average duration of an infection is given by 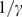 . Note that a contact occurs through initiation by single
. Note that a contact occurs through initiation by single 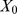 but also if
but also if 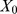 accepts a contact of
accepts a contact of 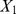 .
.
Pair model
We assume that singles 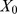 can form a pair,
can form a pair, 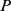 , with a single of the opposite sex
, with a single of the opposite sex 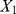 [25]. The transmission dynamics of an STI can then be described as follows:
[25]. The transmission dynamics of an STI can then be described as follows:
| (3) |
| (4) |
| (5) |
| (6) |
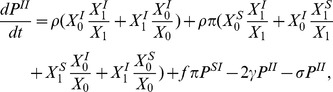 |
(7) |
where 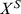 and
and 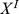 again denote susceptible and infected singles, respectively. Every single
again denote susceptible and infected singles, respectively. Every single 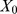 initiates a partnership at a pair formation rate
initiates a partnership at a pair formation rate 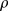 but can also accept a partnership that is initiated by a single
but can also accept a partnership that is initiated by a single 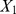 . This results in susceptible-susceptible pairs
. This results in susceptible-susceptible pairs 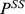 , susceptible-infected pairs
, susceptible-infected pairs 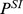 and infected-infected pairs
and infected-infected pairs 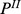 . The average duration of a partnership is given by
. The average duration of a partnership is given by 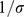 . Every partnership begins with an initial sex act where C. trachomatis can be transmitted at rate
. Every partnership begins with an initial sex act where C. trachomatis can be transmitted at rate  . In an ongoing partnership, transmission occurs at rate
. In an ongoing partnership, transmission occurs at rate 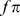 where
where 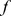 is the frequency of sex acts and
is the frequency of sex acts and  is the transmission probability per sex act.
is the transmission probability per sex act.
Triple model
The pair model framework can be extended so that individuals can have two sexual partnerships at the same time (concurrency) [50]. This allows us to derive an exact formulation of sexual partnership dynamics, compared to purely stochastic descriptions [51], [52] or moment closure approximations [53], [54]. To account for concurrency, we assume that not only singles 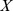 but also individuals that are in one and only one pair
but also individuals that are in one and only one pair 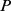 can accept another partnership with a probability
can accept another partnership with a probability  (which is relative to the probability that a single
(which is relative to the probability that a single 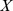 accepts). Such an event can result in a triple
accepts). Such an event can result in a triple 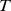 (Fig. 7), which represents the basic unit of concurrent partnerships (an individual with two sexual partnerships at cross-section). Triples can then be elongated to form chains of contacts. Transmission between an infected and susceptible individual happens in the same way as in the pair model. The frequency of sex acts per partnership is constant, i.e., individuals that have two concurrent partnerships have twice as many sex acts per unit of time compared to individuals who are in only one partnership. A full description of the transmission dynamics would require keeping track of the various types of chains together with the infection status of the respective individuals. Here, we concentrate on the overall sexual partnership dynamics that can be described by following the number of singles
(Fig. 7), which represents the basic unit of concurrent partnerships (an individual with two sexual partnerships at cross-section). Triples can then be elongated to form chains of contacts. Transmission between an infected and susceptible individual happens in the same way as in the pair model. The frequency of sex acts per partnership is constant, i.e., individuals that have two concurrent partnerships have twice as many sex acts per unit of time compared to individuals who are in only one partnership. A full description of the transmission dynamics would require keeping track of the various types of chains together with the infection status of the respective individuals. Here, we concentrate on the overall sexual partnership dynamics that can be described by following the number of singles 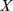 , pairs
, pairs  and triples
and triples 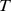 only. The chains are an emergent property of the partnership dynamics and it is therefore not necessary to explicitly consider them in the equations.
only. The chains are an emergent property of the partnership dynamics and it is therefore not necessary to explicitly consider them in the equations.
| (8) |
| (9) |
| (10) |
Figure 7. Schematic depiction of the formation and dissolution of a triple.

Top: An infected single 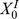 (unconnected white circle) forms a new pair with a susceptible of the opposite sex (black circle in existing pair) which results in a triple
(unconnected white circle) forms a new pair with a susceptible of the opposite sex (black circle in existing pair) which results in a triple 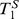 indicating that a susceptible of sex 1 is involved in two pairs. Mid: Transmission can now occur through sexual contacts between the newly formed pair, rendering the triple into
indicating that a susceptible of sex 1 is involved in two pairs. Mid: Transmission can now occur through sexual contacts between the newly formed pair, rendering the triple into 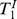 . Bottom: The triple
. Bottom: The triple 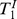 can break up through dissolution of one (
can break up through dissolution of one (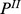 ) or the other pair (
) or the other pair (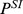 ). Note that pair formation between one single
). Note that pair formation between one single 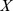 and one pair
and one pair 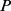 results in two pairs
results in two pairs 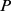 and one triple
and one triple 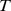 (and vice versa for the pair dissolution). The mathematical representation of each structure is given in rectangles. Subscript 0 denotes white circles (e.g. females) and subscript 1 denotes black circles (e.g. males).
(and vice versa for the pair dissolution). The mathematical representation of each structure is given in rectangles. Subscript 0 denotes white circles (e.g. females) and subscript 1 denotes black circles (e.g. males).
The total population size is now given by 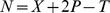 where
where 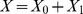 and
and 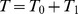 . Note that in the term
. Note that in the term 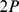 , the individuals who form a triple
, the individuals who form a triple  are counted twice and need to be subtracted. We define the level of concurrency at cross-section,
are counted twice and need to be subtracted. We define the level of concurrency at cross-section, 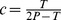 , as the ratio of individuals that have more than one partnership to all individuals in a partnership.
, as the ratio of individuals that have more than one partnership to all individuals in a partnership.
Sexual behavior data and parameter derivation
To parameterize the heterosexual partnership dynamics, we use data from Britain’s second National Survey of Sexual Attitudes and Lifestyles (Natsal-2), a population-based probability sample survey undertaken between 1999–2001 [26]. In order to directly compare the different models, we adjust the contact or pair formation rate, 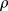 , so that each model exhibits the same number of realized contacts or partnerships as given for the group of 16–25 year old women and men in Natsal-2 (Table 1). The contact rate in the instantaneous contact model is half the mean number of new heterosexual partners per year because every individual can be an initiator and an acceptor of a partnership. Assuming the sexual partnership dynamics has approached steady-state, the pair formation rate for the pair model is given by
, so that each model exhibits the same number of realized contacts or partnerships as given for the group of 16–25 year old women and men in Natsal-2 (Table 1). The contact rate in the instantaneous contact model is half the mean number of new heterosexual partners per year because every individual can be an initiator and an acceptor of a partnership. Assuming the sexual partnership dynamics has approached steady-state, the pair formation rate for the pair model is given by 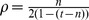 , where
, where  is the average number of new heterosexual partners per year and
is the average number of new heterosexual partners per year and  is the total number of heterosexual partners per year. For the triple model, the pair formation rate is given by
is the total number of heterosexual partners per year. For the triple model, the pair formation rate is given by 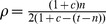 , where
, where  is the level of concurrency. Similarly, the partnership dissolution rate for both the pair and triple model is given by
is the level of concurrency. Similarly, the partnership dissolution rate for both the pair and triple model is given by 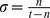 . Note that
. Note that 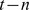 corresponds to the proportion of individuals in a partnership. We also assume the average number of sex acts per partnership to be equal in all three models. The average infectious duration is assumed to be 1 year. This takes into account that most infections are asymptomatic and can persist for more than a year [39], [55]while some infections are shorter due to symptoms. We assume equal transmissibility for both sexes. C. trachomatis prevalence rates in 18–24 year olds in Natsal-2 were 3.0% in women and 2.7% in men in Britain as a whole [56]. Similar levels have been observed in young adults in the US, with no significant difference between women and men [57]. In addition, levels of concordance of C. trachomatis-positivity in women and men in heterosexual partnerships are very similar [58]. The transmission probability per sex act for each model was calibrated so that the steady state prevalence of C. trachomatis was 3%. In the sensitivity analysis, we consider a model where the infectious duration in men is half of that in women, which resulted in a somewhat lower prevalence in men compared to women (see Text S1). Data from Natsal-2 were weighted to adjust for unequal selection probabilities and to correct for the age and gender profile in the population, and mean values were taken for women and men together. Parameter solutions were obtained in Mathematica [59]. For the triple model, the desired transmission probability to obtain the steady-state prevalence could only be approximated through numerical simulations. For the definition of gaps and overlaps between partnerships (Fig. S1), we refer to Althaus et al. [8]. Simulation time strongly depends on the population size (see Fig. S10 in Text S2) which was set to 20′000 if not otherwise indicated.
corresponds to the proportion of individuals in a partnership. We also assume the average number of sex acts per partnership to be equal in all three models. The average infectious duration is assumed to be 1 year. This takes into account that most infections are asymptomatic and can persist for more than a year [39], [55]while some infections are shorter due to symptoms. We assume equal transmissibility for both sexes. C. trachomatis prevalence rates in 18–24 year olds in Natsal-2 were 3.0% in women and 2.7% in men in Britain as a whole [56]. Similar levels have been observed in young adults in the US, with no significant difference between women and men [57]. In addition, levels of concordance of C. trachomatis-positivity in women and men in heterosexual partnerships are very similar [58]. The transmission probability per sex act for each model was calibrated so that the steady state prevalence of C. trachomatis was 3%. In the sensitivity analysis, we consider a model where the infectious duration in men is half of that in women, which resulted in a somewhat lower prevalence in men compared to women (see Text S1). Data from Natsal-2 were weighted to adjust for unequal selection probabilities and to correct for the age and gender profile in the population, and mean values were taken for women and men together. Parameter solutions were obtained in Mathematica [59]. For the triple model, the desired transmission probability to obtain the steady-state prevalence could only be approximated through numerical simulations. For the definition of gaps and overlaps between partnerships (Fig. S1), we refer to Althaus et al. [8]. Simulation time strongly depends on the population size (see Fig. S10 in Text S2) which was set to 20′000 if not otherwise indicated.
Supporting Information
Gaps and overlaps between sexual partnerships. The emergent gaps and overlaps from the triple model correspond well with population-based data of 16–25 year olds from Natsal-2.
(PDF)
Sensitivity analysis. First, this file contains the description of a Chlamydia trachomatis transmission model with heterogeneity in sexual behavior (risk class model) together with the effects of screening and partner notification (PN) in this model. Second, the results of screening and PN are shown for the pair model, assuming that the infectious duration in men is shorter than in women. Third, the effects of different rates of screening uptake and probability of PN in the pair model are shown.
(PDF)
Description of Rstisim. This file provides a brief description of the individual-based modeling framework Rstisim. It contains examples on how to define the partnership formation and an infection. It also contains a section on simulation times for different models.
(PDF)
Acknowledgments
We thank Catherine Mercer and the other investigators of the Natsal-2 study for providing access to the data, and Pelham Barton for his help during the early development of Rstisim. We also thank Kali Tal for providing editorial help with the manuscript and three anonymous reviewers for their critical comments and suggestions.
Funding Statement
This study is part of a project funded by the NIHR Health Technology Assessment Programme (project number 07/42/02). The views and opinions expressed therein are those of the authors and do not necessarily reflect those of the Department of Health. JCMH and SAH acknowledge financial support by the Swiss National Science Foundation (project numbers 320030-118424, 320030-135654 and PDFMP3-124952). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.World Health Organization, UNAIDS (1999) Sexually transmitted diseases: policies and principles for prevention and care. World Health Organization, Geneva, UNAIDS best practice collection edition.
- 2. Low N, Broutet N, Adu-Sarkodie Y, Barton P, Hossain M, et al. (2006) Global control of sexually transmitted infections. Lancet 368: 2001–2016. [DOI] [PubMed] [Google Scholar]
- 3.World Health Organization (2001) Global prevalence and incidence of selected curable sexually transmitted infections: overview and estimates. World Health Organization, Geneva.
- 4. Cates WJ, Wasserheit JN (1991) Genital chlamydial infections: epidemiology and reproductive sequelae. Am J Obstet Gynecol 164: 1771–1781. [DOI] [PubMed] [Google Scholar]
- 5. Trelle S, Shang A, Nartey L, Cassell JA, Low N (2007) Improved effectiveness of partner notification for patients with sexually transmitted infections: systematic review. BMJ 334: 354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Heijne JCM, Althaus CL, Herzog SA, Kretzschmar M, Low N (2011) The role of reinfection and partner notiffication in the effcacy of Chlamydia screening programs. J Infect Dis 203: 372–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Heijne JCM, Herzog SA, Althaus CL, Tao G, Kent CK, et al.. (2012) Insights into the timing of repeated testing after treatment for Chlamydia trachomatis: data and modelling study. Sexually Transmitted Infections. [DOI] [PubMed]
- 8. Althaus CL, Turner KME, Schmid BV, Heijne JCM, Kretzschmar M, et al. (2012) Transmission of Chlamydia trachomatis through sexual partnerships: a comparison between three individual-based models and empirical data. J R Soc Interface 9: 136–146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Kretzschmar M, Welte R, van den Hoek A, Postma MJ (2001) Comparative model-based analysis of screening programs for Chlamydia trachomatis infections. Am J Epidemiol 153: 90–101. [DOI] [PubMed] [Google Scholar]
- 10. Kretzschmar M, Satterwhite C, Leichliter J, Berman S (2012) Effects of screening and partner notification on Chlamydia positivity in the United States: a modeling study. Sex Transm Dis 39: 325–31. [DOI] [PubMed] [Google Scholar]
- 11. Centers for Disease Control and Prevention (2010) Sexually transmitted diseases treatment guidelines, 2006. MMWR Recomm Rep 59: 1–110. [PubMed] [Google Scholar]
- 12. Zimmerman-Rogers H, Potterat JJ, Muth SQ, Bonney MS, Green DL, et al. (1999) Establishing efficient partner notification periods for patients with chlamydia. Sex Transm Dis 26: 49–54. [DOI] [PubMed] [Google Scholar]
- 13.BASHH Clinical Effectiveness Group (2006) 2006 UK National Guideline for the Management of Genital Tract Infection with Chlamydia trachomatis.
- 14. Carré H, Boman J, Osterlund A, Gärdén B, Nylander E (2008) Improved contact tracing for Chlamydia trachomatis with experienced tracers, tracing for one year back in time and interviewing by phone in remote areas. Sex Transm Infect 84: 239–42. [DOI] [PubMed] [Google Scholar]
- 15. Müller J, Kretzschmar M, Dietz K (2000) Contact tracing in stochastic and deterministic epidemic models. Math Biosci 164: 39–64. [DOI] [PubMed] [Google Scholar]
- 16. House T, Keeling MJ (2010) The impact of contact tracing in clustered populations. PLoS Comput Biol 6: e1000721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Eames KTD, Keeling MJ (2003) Contact tracing and disease control. Proc Biol Sci 270: 2565–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Armbruster B, Brandeau ML (2007) Contact tracing to control infectious disease: when enough is enough. Health Care Manag Sci 10: 341–55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Ghani AC, Garnett GP (2000) Risks of acquiring and transmitting sexually transmitted diseases in sexual partner networks. Sex Transm Dis 27: 579–87. [DOI] [PubMed] [Google Scholar]
- 20. Kretzschmar M, van Duynhoven YT, Severijnen AJ (1996) Modeling prevention strategies for gonorrhea and Chlamydia using stochastic network simulations. Am J Epidemiol 144: 306–317. [DOI] [PubMed] [Google Scholar]
- 21. Ghani AC, Swinton J, Garnett GP (1997) The role of sexual partnership networks in the epidemiology of gonorrhea. Sex Transm Dis 24: 45–56. [DOI] [PubMed] [Google Scholar]
- 22. Gray RT, Beagley KW, Timms P, Wilson DP (2009) Modeling the Impact of Potential Vaccines on Epidemics of Sexually Transmitted Chlamydia trachomatis Infection. J Infect Dis 199: 1680–1688. [DOI] [PubMed] [Google Scholar]
- 23. Van de Velde N, Brisson M, Boily MC (2010) Understanding differences in predictions of HPV vaccine effectiveness: A comparative model-based analysis. Vaccine 28: 5473–84. [DOI] [PubMed] [Google Scholar]
- 24.Keeling MJ, Rohani P (2008) Modeling Infectious Diseases in Humans and Animals. Princeton: Princeton University Press.
- 25. Dietz K, Hadeler KP (1988) Epidemiological models for sexually transmitted diseases. J Math Biol 26: 1–25. [DOI] [PubMed] [Google Scholar]
- 26. Johnson AM, Mercer CH, Erens B, Copas AJ, McManus S, et al. (2001) Sexual behaviour in Britain: partnerships, practices, and HIV risk behaviours. Lancet 358: 1835–1842. [DOI] [PubMed] [Google Scholar]
- 27. Katz BP (1992) Estimating transmission probabilities for chlamydial infection. Stat Med 11: 565–577. [DOI] [PubMed] [Google Scholar]
- 28. Tu W, Ghosh P, Katz BP (2011) A Stochastic Model for Assessing Chlamydia trachomatis Transmission Risk Using Longitudinal Observational Data. J R Stat Soc Ser A Stat Soc 174: 975–989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Althaus CL, Heijne JCM, Low N (2012) Towards More Robust Estimates of the Transmissibility of Chlamydia trachomatis. Sex Transm Dis 39: 402–4. [DOI] [PubMed] [Google Scholar]
- 30. Morris M, Kretzschmar M (1997) Concurrent partnerships and the spread of HIV. AIDS 11: 641–648. [DOI] [PubMed] [Google Scholar]
- 31. Tayal SC, Ochogwu SA, Crindon S (2010) Audit of partner notification for Chlamydia infection in the genitourinary medicine clinic at the University Hospital of Hartlepool: 2004–2008. Int J STD AIDS 21: 516–8. [DOI] [PubMed] [Google Scholar]
- 32.Herzog SA, McClean H, Carne CA, Low N (2011) Variation in partner notification outcomes for chlamydia in UK genitourinary medicine clinics: multilevel study. Sex Transm Infect. [DOI] [PubMed]
- 33. Khan A, Fortenberry JD, Juliar BE, Tu W, Orr DP, et al. (2005) The prevalence of chlamydia, gonorrhea, and trichomonas in sexual partnerships: implications for partner notification and treatment. Sex Transm Dis 32: 260–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Rogers SM, Miller WC, Turner CF, Ellen J, Zenilman J, et al. (2008) Concordance of chlamydia trachomatis infections within sexual partnerships. Sex Transm Infect 84: 23–8. [DOI] [PubMed] [Google Scholar]
- 35. Korenromp EL, Sudaryo MK, de Vlas SJ, Gray RH, Sewankambo NK, et al. (2002) What proportion of episodes of gonorrhoea and chlamydia becomes symptomatic? Int J STD AIDS 13: 91–101. [DOI] [PubMed] [Google Scholar]
- 36. Chow JM (2012) Measuring the uptake and impact of Chlamydia screening programs easier said than done. Sex Transm Dis 39: 89–91. [DOI] [PubMed] [Google Scholar]
- 37.Handcock MS, Hunter DR, Butts CT, Goodreau SM, Morris M (2003) statnet: Software tools for the Statistical Modeling of Network Data. Seattle, WA. Version 2.0. [DOI] [PMC free article] [PubMed]
- 38. De Blasio BF, Svensson A, Liljeros F (2007) Preferential attachment in sexual networks. Proc Natl Acad Sci U S A 104: 10762–10767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Althaus CL, Heijne JC, Roellin A, Low N (2010) Transmission dynamics of Chlamydia trachomatis affect the impact of screening programmes. Epidemics 2: 123–131. [DOI] [PubMed] [Google Scholar]
- 40. Brunham RC, Pourbohloul B, Mak S, White R, Rekart ML (2005) The unexpected impact of a Chlamydia trachomatis infection control program on susceptibility to reinfection. J Infect Dis 192: 1836–1844. [DOI] [PubMed] [Google Scholar]
- 41. Batteiger BE, Xu F, Johnson RE, Rekart ML (2010) Protective immunity to Chlamydia trachomatis genital infection: evidence from human studies. J Infect Dis 201 Suppl 2S178–89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Eaton JW, Johnson LF, Salomon JA, Bärnighausen T, Bendavid E, et al. (2012) HIV Treatment as Prevention: Systematic Comparison of Mathematical Models of the Potential Impact of Antiretroviral Therapy on HIV Incidence in South Africa. PLoS Med 9: e1001245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. May RM (2004) Uses and abuses of mathematics in biology. Science 303: 790–793. [DOI] [PubMed] [Google Scholar]
- 44. Regan DG, Wilson DP (2008) Modelling sexually transmitted infections: less is usually more for informing public health policy. Trans R Soc Trop Med Hyg 102: 207–208. [DOI] [PubMed] [Google Scholar]
- 45. Bogaards JA, Kretzschmar M, Xiridou M, Meijer CJLM, Berkhof J, et al. (2011) Sex-specific immunization for sexually transmitted infections such as human papillomavirus: insights from mathematical models. PLoS Med 8: e1001147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Regan DG, Wilson DP, Hocking JS (2008) Coverage is the key for effective screening of Chlamydia trachomatis in Australia. J Infect Dis 198: 349–358. [DOI] [PubMed] [Google Scholar]
- 47. Mercer CH, Aicken CRH, Brook MG, Estcourt CS, Cassell JA (2011) Estimating the likely public health impact of partner notification for a clinical service: an evidence based algorithm. Am J Public Health 101: 2117–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.R Development Core Team (2009) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0.
- 49.Butts CT, Handcock MS, Hunter DR (2008) network: Classes for Relational Data. Irvine, CA. R package version 1.4–1.
- 50.Dietz K, Tudor D (1992) Triangles in heterosexual HIV transmission. In: Jewell N, Kietz K, VT F, editors, AIDS Epidemiology: Methodological Issues, Birkhäuser, Boston. 143–155.
- 51. Morris M, Kretzschmar M (1995) Concurrent partnerships and transmission dynamics in networks. Social Networks 17: 299–318. [Google Scholar]
- 52. Kretzschmar M, Morris M (1996) Measures of concurrency in networks and the spread of infectious disease. Math Biosci 133: 165–195. [DOI] [PubMed] [Google Scholar]
- 53. Bauch C, Rand DA (2000) A moment closure model for sexually transmitted disease transmission through a concurrent partnership network. Proc Biol Sci 267: 2019–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Ferguson NM, Garnett GP (2000) More realistic models of sexually transmitted disease transmission dynamics: sexual partnership networks, pair models, and moment closure. Sex Transm Dis 27: 600–9. [DOI] [PubMed] [Google Scholar]
- 55. Molano M, Meijer CJLM, Weiderpass E, Arslan A, Posso H, et al. (2005) The natural course of Chlamydia trachomatis infection in asymptomatic Colombian women: a 5-year follow-up study. J Infect Dis 191: 907–916. [DOI] [PubMed] [Google Scholar]
- 56. Fenton KA, Korovessis C, Johnson AM, McCadden A, McManus S, et al. (2001) Sexual behaviour in Britain: reported sexually transmitted infections and prevalent genital Chlamydia trachomatis infection. Lancet 358: 1851–1854. [DOI] [PubMed] [Google Scholar]
- 57. Datta SD, Sternberg M, Johnson RE, Berman S, Papp JR, et al. (2007) Gonorrhea and chlamydia in the United States among persons 14 to 39 years of age, 1999 to 2002. Ann Intern Med 147: 89–96. [DOI] [PubMed] [Google Scholar]
- 58. Quinn TC, Gaydos C, Shepherd M, Bobo L, Hook EW 3rd, et al (1996) Epidemiologic and microbiologic correlates of Chlamydia trachomatis infection in sexual partnerships. JAMA 276: 1737–42. [PubMed] [Google Scholar]
- 59.Wolfram Research, Inc (2008) Mathematica Edition: Version 7.0. Champaign, Illinois: Wolfram Research, Inc.
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Gaps and overlaps between sexual partnerships. The emergent gaps and overlaps from the triple model correspond well with population-based data of 16–25 year olds from Natsal-2.
(PDF)
Sensitivity analysis. First, this file contains the description of a Chlamydia trachomatis transmission model with heterogeneity in sexual behavior (risk class model) together with the effects of screening and partner notification (PN) in this model. Second, the results of screening and PN are shown for the pair model, assuming that the infectious duration in men is shorter than in women. Third, the effects of different rates of screening uptake and probability of PN in the pair model are shown.
(PDF)
Description of Rstisim. This file provides a brief description of the individual-based modeling framework Rstisim. It contains examples on how to define the partnership formation and an infection. It also contains a section on simulation times for different models.
(PDF)