

Abstract
Experiments suggest that biodiversity enhances the ability of ecosystems to maintain multiple functions, such as carbon storage, productivity, and buildup of nutrient pools (multifunctionality). However, the relationship between biodiversity and multifunctionality has never been assessed globally in natural ecosystems. We report on the first global empirical study relating plant species richness and abiotic factors to multifunctionality in drylands, which collectively cover 41% of Earth’s land surface and support over 38% of the human population. Multifunctionality was positively and significantly related to species richness. The best-fitting models accounted for over 55% of the variation in multifunctionality, and always included species richness as a predictor variable. Our results suggest that preservation of plant biodiversity is crucial to buffer negative effects of climate change and desertification in drylands.
Two decades of research have revealed causal linkages between biodiversity and univariate measures of ecosystem functioning, such as primary productivity or nitrogen accumulation, in many terrestrial and aquatic habitats (1-4). These relationships suggest that the loss of biodiversity may impair the functioning of natural ecosystems, and thus diminish the number and quality of services they provide (5-7). Ecosystems are valued for their ability to maintain multiple functions and services simultaneously (multifunctionality, 8). If the maintenance of biodiversity is to be justified as a strategy for enhancing ecosystem services (5, 9), it is essential to understand how biodiversity affects multifunctionality (8-10). Existing knowledge comes from controlled small-scale experiments from a limited number of ecosystems, mainly in North America and Europe (8-12). Furthermore, biodiversity is by no means the only, or even the primary, driver of ecosystem functioning, which is also influenced by other biotic and abiotic factors (13, 14). Given this complexity, a rigorous examination is needed of the role of biodiversity in maintaining multifunctionality at a large number of sites that represent a wide range of spatial variability in resource availability, abiotic factors, and species richness and composition (15).
Arid, semi-arid and dry-subhumid ecosystems (hereafter “drylands”) constitute some of the largest terrestrial biomes, collectively covering 41% of Earth’s land surface and supporting over 38% of the global human population (16). Drylands host many endemic plant and animal species (5), and include about 20% of the major centers of global plant diversity and over 30% of the designated endemic bird areas (17). These ecosystems are also highly vulnerable to global environmental change and desertification (16, 18). Nevertheless, the relationship between biodiversity and ecosystem functioning has seldom been studied in drylands (19). We evaluated how the richness of perennial vascular plants (hereafter species richness) and a range of key abiotic factors (climate, slope, elevation, and soil texture) relate to multifunctionality in 224 dryland ecosystems sampled from all continents except Antarctica (map S1). We surveyed 30 m × 30 m plots, large enough to represent the main ecosystem features at each site, and assessed 14 ecosystem functions related to the cycling and storage of carbon (C; organic C, β-glucosidase, pentoses, hexoses, aromatic compounds, and phenols), nitrogen (N; total N, NO3−-N, NH +4-N, aminoacids, proteins, and potential N transformation rate) and phosphorus (P; available inorganic P and phosphatase). These functions were chosen because they deliver some of the fundamental supporting and regulating ecosystem services (9, 18, 20), and because they are used to identify the onset of desertification processes (21). Our survey captured a substantial range of the climatic conditions, ecosystem types and soil classes found in drylands worldwide (fig. S1, map S1).
We first evaluated the direct relationship between species richness and multifunctionality at the global scale using both non-spatial (ordinary least squares [OLS]) and spatial (simultaneous autoregression, [SAR]) regression models (20). Because we did not experimentally control for other abiotic and biotic factors that are known to affect ecosystem functioning, significant relationships would indicate potentially strong effects of richness on multifunctionality. To quantify multifunctionality, we calculated Z-scores (standardized deviates) of the 14 functions evaluated (20). The multifunctionality index M for each plot was the average Z-score for all functions measured within the plot. This index measures all functions on a common scale of standard deviation units, has good statistical properties, and is well-correlated with previously proposed indices for quantifying multifunctionality (20, fig. S4). Multifunctionality was positively and significantly (P < 0.05) related to species richness, according to both OLS and SAR models (Fig. 1A). Separate analyses of functions related to the C, N and P cycles (20) also yielded positive and significant relationships with species richness in all cases when using OLS regression (Figs. 1B-D). When SAR regressions were used, significant relationships were found only for functions related to C cycling (Figs. 1B-D).
Figure 1.

Relationship between perennial plant species richness and ecosystem multifunctionality (A) measured in a global survey of drylands. Similar relationships for carbon (B), nitrogen (C) and phosphorus (D) cycling are shown. Red and green lines are the fitted lines from ordinary least squares (OLS) and spatial autoregressive (SAR) regressions, respectively. Results of regressions are as follows: A) OLS, R2 = 0.030, P = 0.009; SAR, R2 = 0.022, P = 0.027; B) OLS, R2 = 0.029, P = 0.011; SAR, R2 = 0.022, P = 0.027; C) OLS, R2 = 0.018, P = 0.044; SAR, R2 = 0.014, P = 0.082; and D) OLS, R2 = 0.032, P = 0.008; SAR, R2 = 0.016, P = 0.061.
We then evaluated whether observed effects of species richness were important compared to those of abiotic factors with a multi-model inference approach based on information theory and OLS regression (22). We built separate models using the multifunctionality index M and functions from the N, C and P cycles as dependent variables, and seven abiotic variables (sand content, slope, elevation and four components derived from a principal component analysis of 21 available climatic variables, 20) plus species richness as potential independent variables. Among the 255 possible models resulting from all possible combinations of these independent variables, we selected the set of best-fitting models that minimized the second-order Akaike information criterion (AICc). Collinearity among independent variables in these models was negligible (20, table S15). Whenever a model included species richness as an important predictor, we compared its AICc to that of the corresponding model without species richness; differences < 2.0 in AICc between alternative models indicate they are approximately equivalent in explanatory power (22). To account for potential effects of spatial autocorrelation between sites, latitude and longitude were included in all the models (23).
The best and most parsimonious models (smallest AICc and fewest variables with comparable AICc, respectively) describing global multifunctionality contained 9 and 7 predictor variables (Table 1). Both models explained more than 55% of the variance found in multifunctionality, and included species richness. In both cases, removal of species richness as a predictor variable substantially reduced the model fit (Table 1). These results were virtually identical to those obtained with SAR regression and OLS models that included quadratic terms, to account for potential autocorrelation and non-linear effects, respectively (20, tables S2 and S3), and for models that used other multifunctionality indices proposed in the literature (20, table S13). Species richness was also an important factor in separate models of C and N cycling (tables S4, S5, S7, S8, S10 and S11), but had weaker effects on P cycling (tables S6, S9 and S12). Overall, the general result that species richness makes important contributions to multifunctionality was robust to the analytical methods used and to the choice of multifunctionality index.
Table 1.
Best-fitting regression models of ecosystem multifunctionality. Each column represents a different predictor variable (red = perennial plant species richness; green = abiotic variables, blue = climatic variables, gold = geographic variables). Of all 255 possible models, the best eight models are presented, ranked according to the second-order Akaike information criterion (AICc). AICc measures the relative goodness of fit of a given model; the lower its value, the more likely this model is correct. Unshaded cells indicate variables that were not included in a particular model. The first and third models of the table are the best and most parsimonious models, respectively; the same models without species richness had R2 = 0.539, AICc = 293.236, Δ AICc = 10.486, and R2 = 0.515, AICc = 300.078, ΔAICc = 17.328, respectively.
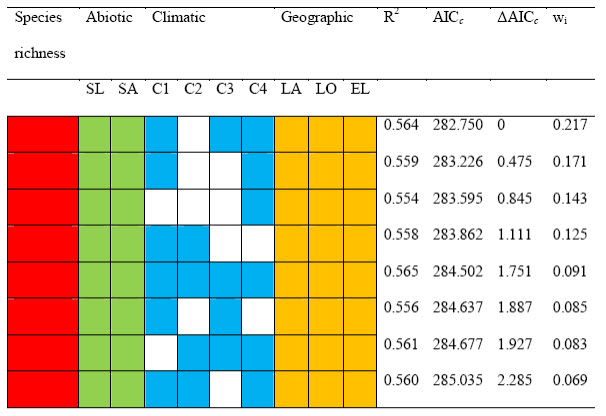
R2 = percent variation in multifunctionality explained by the model, ΔAICc = difference between the AICc of each model and that of the best model, wi = Akaike weights, C1, C2, C3, C4 = first, second, third and fourth components of a principal component analysis conducted with climatic variables, SA = sand content, SL = slope angle (sqrt-transformed), EL = elevation (sqrt-transformed), LA =latitude, and LO = longitude.
To quantify the relative importance of the different predictors of multifunctionality, we summed the Akaike weights for each predictor across all the models in which it occurred (20, 22); the larger this sum, the more important a given variable is relative to the other variables used in the same models. By this criterion, the two most important predictors of multifunctionality were annual mean temperature (reflected in large negative loadings for the 4th principal component of the climatic variables, 20) and sand content of the soil (Fig. 2A). Both variables were negatively related to multifunctionality: higher ecosystem functionality was found at cooler temperatures and lower sand content (table S14). The importance of species richness was very similar to that of mean temperature and sand content. Indeed, species richness was more important than climatic variables such as mean annual rainfall, and mean temperature and rainfall in the driest quarter (reflected in loadings on the first and third principal components of the climatic variables, respectively, 20). Similar results were obtained when functions related to the C and N cycles were evaluated separately (Figs. 2B-C). Species richness was less important to P cycling than other abiotic factors such as sand content, elevation, and annual rainfall (Fig. 2D).
Figure 2.

Relative importance of perennial plant species richness (red column) and other predictor variables in models of ecosystem multifunctionality (A), and carbon (B), nitrogen (C) and phosphorus (D) cycling. The height of each bar is the sum of the Akaike weights of all models that included the predictor of interest, taking into account the number of models in which each predictor appears. Variable abbreviations as in Table 1.
The positive effects of species richness on multifunctionality may be mediated through increased net primary production (NPP), which has cascading effects on multiple organisms and ecosystem processes (1, 24). However, the relationship between plant species richness and NPP is uncertain (25), and NPP could not be measured in this study. We speculate instead that complementarity in the use of resources such as water (2, 9), which has been demonstrated in drylands and can occur without changes in NPP (26, 27), accounts for correlations between species richness and multifunctionality. Our results also implicate soil water conditions, which are largely affected by temperature and soil texture (28), as an important driver of multifunctionality.
By itself, species richness accounted for only a small fraction of the observed variation in multifunctionality of drylands (Fig. 1). However, the best-fitting models accounted for over 55% of this variation, and always included species richness (Table 1). The unexplained variation likely reflects factors not measured in our global survey, including the intensity of herbivory, historical patterns of land use, the presence of keystone and invasive species, and differences in components of biodiversity such as soil fauna, whose changes along environmental gradients do not necessarily track those of plant richness (28-30).
Climate change models predict increases in average annual temperature in drylands up to 4°C by the end of the XXIth century (31). Our results suggest that such an increase will reduce the ability of dryland ecosystems to perform multiple functions related to C, N and P cycling. Ongoing climate change is also likely to reduce local species richness (32), and to increase the extent of areas affected by desertification (16, 18), both of which will negatively impact ecosystem functioning. However, these outcomes are uncertain because of the complex interactions and contrasting effects of increases in temperature, which we found to reduce multifunctionality, and in atmospheric carbon dioxide concentrations, which can ameliorate water stress in dryland vegetation and potentially minimize biodiversity losses (33, 34). Because the quality and quantity of ecosystem services depend largely on ecosystem functions such as those measured in this study (5, 9), increased plant species richness may enhance the services provided by dryland ecosystems. Our findings also suggest that such richness may be particularly important for maintaining ecosystem functions linked to C and N cycling, which sustain carbon sequestration and soil fertility (18, 28). Because land degradation is often accompanied by the loss of soil fertility (16, 18), plant species richness may also promote ecosystem resistance to desertification.
The consistent effects of species richness on multifunctionality over and above those of climate and abiotic factors highlight the importance of plant biodiversity as a driver of multifunctionality in drylands. The positive relationship between species richness and multifunctionality found is consistent with experimental results obtained in temperate grasslands, microbial, biological soil crust and aquatic communities (8-12). Collectively, these results suggest that the correlation between species richness and multifunctionality may be a general pattern in nature that reflects a cause-and-effect linkage.
One-sentence summary: Plant species richness is positively and significantly related to ecosystem multifunctionality in drylands at a global scale.
Supplementary Material
Acknowledgements
We thank all the technicians that assisted with field and laboratory work, T. Navarro and Z. Noumi for their help with plant species identification, and J. Bascompte, J. F. Reynolds, K. J. van Groenigen, W. van der Puten and two anonymous reviewers for helpful comments. This research was funded by the European Research Council under the European Community’s Seventh Framework Programme (FP7/2007-2013)/ERC Grant agreement 242658 (BIOCOM). CYTED funded networking activities (EPES, Acción 407AC0323).
Footnotes
Supporting Online Material www.sciencemag.org Materials and Methods Figs. S1 to S10 Tables S1 to S15 References (35-137) Database S1 Map S1
The data used in the primary analyses are available in the Supporting Online Material. The authors declare no competing financial interests. F.T.M. and A.E. designed the study. Field data were collected by all authors but A.E., A.G., N.J.G., B.G., E.V. and M.B. Laboratory analyses were done by V.O., A.G., M.B., M.D.B., E.V. and B.G. Data analyses were done by F.T.M., assisted by N.J.G. The paper was written by F.T.M., and all authors contributed to the subsequent drafts.
References and Notes
- 1.Cardinale BJ, et al. Am. J. Bot. 2011;98:572. doi: 10.3732/ajb.1000364. [DOI] [PubMed] [Google Scholar]
- 2.Hooper DU, et al. Ecol. Monogr. 2005;75:3. [Google Scholar]
- 3.Hector A, et al. Science. 1999;286:1123. doi: 10.1126/science.286.5442.1123. [DOI] [PubMed] [Google Scholar]
- 4.Flombaum P, Sala OE. Proc. Nat. Acad. Sci. USA. 2008;105:6087. doi: 10.1073/pnas.0704801105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Millennium Ecosystem Assessment . Ecosystems and Human Well-being: Biodiversity Synthesis. World Resources Institute; Washington, DC: 2005. [Google Scholar]
- 6.Naeem S, Bunker DE, Hector A, Loreau M, Perrings C, editors. Biodiversity, Ecosystem Functioning and Human Wellbeing. An Ecological and Economic Perspective. Oxford University Press; Oxford: 2009. [Google Scholar]
- 7.Guo Z, et al. PloS One. 2010;5:e13113. [Google Scholar]
- 8.Zavaleta ES, et al. Proc. Nat. Acad. Sci. USA. 2010;107:1443. doi: 10.1073/pnas.0906829107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Isbell F, et al. Nature. 2011;477:199. doi: 10.1038/nature10282. [DOI] [PubMed] [Google Scholar]
- 10.Hector A, Bagchi R. Nature. 2007;448:188. doi: 10.1038/nature05947. [DOI] [PubMed] [Google Scholar]
- 11.Gamfeldt L, Hillebrand H, Jonsson PR. Ecology. 2008;89:1223. doi: 10.1890/06-2091.1. [DOI] [PubMed] [Google Scholar]
- 12.Maestre FT, Castillo-Monroy AP, Bowker M, Ochoa-Hueso R. J Ecol. 2011 doi: 10.1111/j.1365-2745.2011.01918.x. [Google Scholar]
- 13.Maestre FT, et al. Phil. Trans. R. Soc. B. 2010;365:2057. doi: 10.1098/rstb.2010.0016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Godbold JA, Solan M. Mar. Ecol. Progr. Ser. 2009;396:273. [Google Scholar]
- 15.Wardle DA, Jonsson M. Front. Ecol. Environ. 2010;8:10. [Google Scholar]
- 16.Reynolds JF, et al. Science. 2007;316:847. doi: 10.1126/science.1131634. [DOI] [PubMed] [Google Scholar]
- 17.White RP, Nackoney J. Drylands, People, and Ecosystem Goods and Services: A Web-Based Geospatial Analysis. World Resources Institute; Washington, DC: 2003. http://www.wri.org/publication/content/8241) [Google Scholar]
- 18.Millennium Ecosystem Assessment . Ecosystems and Human Well-being: Desertification Synthesis. World Resources Institute; Washington, DC: 2005. [Google Scholar]
- 19.A search on the ISI Web of Science® (18 November 2011) using the keywords “species AND (diversity OR richness) AND (community OR ecosystem) AND (function OR functioning OR production OR productivity OR biomass OR predation OR decomposition OR herbivory)” yielded 14136 documents, from which only 2.5% of them contained the word “arid”.
- 20.Materials and methods are available as supporting material on Science Online.
- 21.Maestre FT, Escudero A. Ecology. 2009;90:1729. doi: 10.1890/08-2096.1. [DOI] [PubMed] [Google Scholar]
- 22.Burnham KP, Anderson DR. Model Selection and Multimodel Inference: a Practical Information-Theoretic Approach. ed. 2. Springer; New York: 2002. [Google Scholar]
- 23.Diniz-Filho JAF, Rangel TFLVB, Bini LM. Global Ecol. Biogeogr. 2008;17:479. [Google Scholar]
- 24.Pringle RM, Young TP, Rubenstein DI, McCauley DJ. Proc. Nat. Acad. Sci. USA. 2007;104(193) doi: 10.1073/pnas.0609840104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Adler PB, et al. Science. 2011;333:1750. doi: 10.1126/science.1204498. [DOI] [PubMed] [Google Scholar]
- 26.O'Connor TG, Haines LM, Snyman HA. J Ecol. 2001;89:850. [Google Scholar]
- 27.Hooper DU, Vitousek PM. Ecol. Monogr. 1998;68:121. [Google Scholar]
- 28.Whitford WG. Ecology of Desert Systems. Academic Press; San Diego: 2002. [Google Scholar]
- 29.Wu T, Ayres E, Bardgett RD, Wall DH, Garey JR. Proc. Nat. Acad. Sci. USA. 2011;108:17720. doi: 10.1073/pnas.1103824108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Evans RD, Rimer R, Sperry L, Belnap J. Ecol. Appl. 2001;11:1301. [Google Scholar]
- 31.Solomon S, et al. Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press; Cambridge: 2007. [Google Scholar]
- 32.Sala O, et al. Science. 2000;287:1770. doi: 10.1126/science.287.5459.1770. [DOI] [PubMed] [Google Scholar]
- 33.Morgan JA, et al. Nature. 2011;476:202. doi: 10.1038/nature10274. [DOI] [PubMed] [Google Scholar]
- 34.Woodward FI, Kelly CK. Ecol. Lett. 2008;11:1229. doi: 10.1111/j.1461-0248.2008.01240.x. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.