

Abstract
Background
Vector-borne diseases remain a threat to public health, especially in tropical countries. The incompatible insect technique has been explored as a potential control strategy for several important insect vectors. However, this strategy has not been tested in Culex pipiens pallens, the most prevalent mosquito species in China. Previous works used introgression to generate new strains that matched the genetic backgrounds of target populations while harboring a new Wolbachia endosymbiont, resulting in mating competitiveness and cytoplasmic incompatibility. The generation of these incompatible insects is often time-consuming, and the long-term stability of the newly created insect-Wolbachia symbiosis is uncertain. Considering the wide distribution of Cx. pipiens pallens and hence possible isolation of different populations, we sought to test for incompatibilities between natural populations and the possibility of exploiting these incompatibilities as a control strategy.
Methodology/Principal Findings
Three field populations were collected from three geographic locations in eastern China. Reciprocal cross results showed that bi-directional patterns of incompatibility existed between some populations. Mating competition experiments indicated that incompatible males could compete with cognate males in mating with females, leading to reduced overall fecundity. F1 offspring from incompatible crosses maintained their maternal crossing types. All three populations tested positive for Wolbachia. Removal of Wolbachia by tetracycline rendered matings between these populations fully compatible.
Conclusions/Significance
Our findings indicate that naturally occurring patterns of cytoplasmic incompatibility between Cx. pipiens pallens populations can be the basis of a control strategy for this important vector species. The observed incompatibilities are caused by Wolbachia. More tests including field trials are warranted to evaluate the feasibility of this strategy as a supplement to other control measures.
Author Summary
Population suppression is an important component of mosquito control measures. The incompatible insect technique exploits the monogamous mating behavior of female mosquitoes to decrease the percentage of females inseminated by compatible males and hence reduce overall fecundity. Previous studies used genetically engineered Wolbachia-infected mosquitoes as the sources of incompatible males. The long-term stability of these mosquitoes is unknown. In this study, we examined naturally occurring incompatibilities between different field populations of Culex pipiens pallens, a vector of West Nile Virus and filarial worms widely distributed in China. We found that bi-directional patterns of incompatibility existed between some Cx. pipiens pallens populations. Incompatible males could compete against cognate males in mating with females and suppress reproduction. The level of suppression depended on the relative population sizes of incompatible and cognate males. We also found that these incompatibilities were preserved in the offspring from incompatible crosses, indicating that this control strategy is likely to be sustainable when applied repeatedly to successive mosquito generations. Our data also indicate that these incompatibilities were caused by endosymbiont Wolbachia, which is also the basis for cytoplasmic incompatibility in a variety of mosquito species. Our results may help to design simpler and more time-effective control strategies for a number of vector-borne diseases.
Introduction
Vector-borne diseases especially those transmitted by mosquitoes such as malaria, dengue fever, Japanese encephalitis, West Nile fever, Chikungunya and lymphatic filariasis are still important scourges responsible for millions of deaths each year, especially in tropical and developing countries. The lack of effective vaccines for major mosquito-borne diseases and the development of resistance to available chemotherapies both underscore the importance of reducing the numbers of major vector mosquitoes. Insecticides are the major weapons for mosquito control. Earlier efforts using insecticides to reduce malaria and other neglected tropical diseases had been met with success [1], [2]. But, long-term intensive use of insecticides has led to the development of insecticide resistance in important vector mosquito species, jeopardizing the effectiveness of insecticide-based vector control [3]. In addition, the adverse effects of insecticides on human health and the environment could not be ignored [4], [5].
With these concerns, biological approaches are called upon as alternatives to chemical control. One approach is to render the arthropods incapable of disease transmission. This strategy uses transgenesis or paratransgenesis to introduce foreign genes into arthropod populations. The expression of the transgenes makes these arthropods resistant to infections or unsuitable for parasite development to their infective stages [6], [7]. Introduction of endosymbionts such as Wolbachia, maternally inherited obligatory intracellular bacteria, into certain mosquito species can also decrease their vectorial capacity and have yielded novel strategies for population replacement [8]–[11]. Another approach resembles chemical control in that it aims at population reduction. For example, transgenes or biological agents such as Wolbachia can be introduced into mosquito populations to reduce fertility [12]–[15].
Because females of many insect species mate only once in their life-time, unproductive mating effectively precludes their reproduction or reduces their fecundity. Two strategies have been developed to take advantage of their monogamous behaviors, namely the sterile insect technique (SIT) and the incompatible insect technique (IIT). SIT uses radiologically or chemically sterilized males to suppress target populations. IIT uses incompatibilities especially cytoplasmic incompatibility (CI) between males and females to reduce female fecundity. CI is an incompatibility between a sperm and an egg that interferes with the normal development of a zygote. It is caused by endosymbionts in the cytoplasm such as Wolbachia [15]–[17]. Since their initial identification in the ovaries of Culex mosquitoes [18], Wolbachia have been found in the reproductive tissues of the majority of tested arthropod species [19]. Wolbachia cause a variety of reproductive abnormalities besides CI, such as feminization of genetic males, thelytokous parthenogenesis (female offspring being produced from unfertilized eggs), and male-killing [15], [20]. Among these abnormalities, CI has been explored as a potential tool for the development of novel and environmentally friendly strategies for insect controls [9], [21]. Because CI exists between sperm of Wolbachia-infected mosquitoes and eggs of an uninfected or an incompatible strain of Wolbachia-infected mosquitoes, CI-based IIT is applicable to both Wolbachia-negative and Wolbachia-positive target populations. CI prevents embryonic development, thus making the mating events unproductive. Consequently, those mated females are unable to reproduce or have their fecundity greatly reduced.
The application of CI to suppress insect populations predated the identification of Wolbachia as its causative agent. Laven used an introgressed Culex pipiens fatigans strain with maternal cytoplasm derived from a Paris strain to successfully suppress the incompatible local Cx. pipiens fatigans populations in Myanmar [22]. Subsequently, CI-based suppression was trialed for the European cherry fruit fly Rhagoletis cerasi [23] and almond warehouse moth Cadra (Ephestia) cautella [24]. To improve the safety of this suppression strategy, an irradiation step was included in several studies on Culex mosquitoes to minimize inadvertent release of fertile females and minority-type (compatible) males [25]–[27]. Medfly Ceratitis capitata transinfected with Wolbachia derived from R. cerasi have been generated for IIT approaches to suppress medfly populations [28]. CI has also been utilized to develop strategies to control Aedes mosquitoes. Through interspecific hybridization and introgression, an Aedes polynesiensis strain harboring Wolbachia derived from Aedes riversi was generated. This new strain is bi-directionally incompatible with the natural strain [29].
Wolbachia-infected mosquito strains that display novel patterns of CI for population suppression are usually created by introgression or transinfection [30]–[32]. The preparation of incompatible mosquitoes using such methods is often laborious and time-consuming. Because introgression involves putting Wolbachia into new host genetic backgrounds, the long-term stability of such novel mosquito-Wolbachia symbioses is unknown. In addition, although IIT has been applied to a variety of insects, it has not been tested in Culex pipiens pallens, one of the most prevalent mosquito species in China. In this study, using three Cx. pipiens pallens field populations collected in eastern China, we tested the applicability of exploiting incompatibilities between naturally occurring populations to suppress target populations. We report that incompatible males could compete with cognate males in mating with females, resulting in reduced fecundity. Though the incompatibilities were not absolute, they were preserved with hybrid offspring keeping the maternal crossing types. Our data indicate that this strategy may be a viable and sustainable approach over time to suppress populations of Cx. pipiens pallens.
Materials And Methods
Mosquitoes
WX (Wuxi, Jiangsu Province, 31°33′58.47″N, 120°18′9.88″E), NJ (Nanjing, Jiangsu Province, 32°3′30.11″N, 118°47′47.28″E), and TK (Tangkou, Shandong Province, 34°52′34.97″N, 117°22′53.69″E) populations of Cx. pipiens pallens were used in this study (Figure 1) [33]. WX and TK populations were collected in 2008 from July to August. NJ population was collected in July, 2006. For each population, several hundreds of larvae were collected with 350-ml dippers from over 50 larval habitats in public areas of the respective location. Fourth-instar larvae were identified to species by morphology, including the aspect ratio of the air tube and the number of pectens and tufts on it. The larvae were then brought back and reared in the insectary. Mosquitoes were kept at 28°C and 75% relative humidity with 14 h∶10 h light∶dark cycle. Adult mosquitoes were fed 10% (w/v) glucose solution prior to blood meals.
Figure 1. Collecting sites of Cx. pipiens pallens field populations.
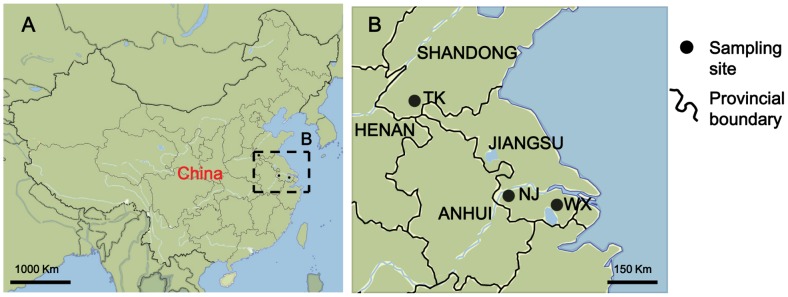
(A) The mosquito populations were collected from eastern China; (B) Three collecting sites Tangkou, Nanjing and Wuxi are shown in detail. The mosquito populations collected are designated TK, NJ and WX, respectively.
Crossing experiments
To separate virgin females and males, pupae from each population were put into 15-ml tubes with water for individual emergence. Afterward, male and female adults were raised in 30.5×30.5×30.5 cm cages. Females 2 days after emergence and males 3 days after emergence were used in mating experiments. Each set of crossings included combination groups of virgin males and females from two different Cx. pipiens pallens populations, with combinations of males and females from the same populations as positive controls. Females and males placed in the same cages were given 2 days to mate. Females were blood fed after mating, then the egg rafts were given 48 hours after oviposition to hatch in separate containers. The numbers of eggs and larvae were counted under stereoscope and the hatching rates (HR) for individual rafts were calculated. Unhatched egg rafts were all examined for fertilization through microscopic observation of embryonic development [34]. Each experiment was performed twice, shown here is a representative result.
Detection of Wolbachia
Total DNA of individual Cx. pipiens pallens mosquito was extracted using a method previously described [35]. Polymerase chain reaction (PCR) was first carried out using generic primers to amplify wsp gene which encodes the major surface protein of Wolbachia to determine the infection status of the mosquito [36]. Studies have reported that the majority of insect Wolbachia belong to supergroups A and B, specific wsp primers were used accordingly for typing [37]. All primers were adopted from published studies. Primers wF and wR (see Table 1 for primer sequences) were used to amplify a 590–632 bp fragment from all Wolbachia strains; primers wAF and wR were used to amplify a 556 bp fragment characteristic of supergroup A; primers wBpipF and wR were used to amplify a 501 bp fragment characteristic of wPip strains of supergroup B; and primers wBcauBF and wR were used to amplify a 466 bp fragment characteristic of the wCauB strains of supergroup B. In addition, PCR was carried out to amplify the ankyrin domain-encoding gene ank2 to differentiate wPip strains into groups I-V based on 313–511 bp amplification fragments [38].
Table 1. Genes used for Wolbachia typing and their PCR amplification primers.
| Gene | Putative product | Primer (5′-3′) | Size of expected amplification fragment (bp) | References |
| wsp | major surface protein | wF-TGGTCCAATAAGTGATGAAGAAAC | 590–632 bp | [36], [37] |
| wAF-TGAAATTTTACCTCTTTTC | 556 bp | |||
| wBpipF-AAGGAACCGAAGTTCATG | 501 bp | |||
| wBcauBF-CCATCTTTTCTAGCTGGA | 466 bp | |||
| wR-AAAAATTAAACGCTACTCCA | ||||
| ank2 | ankyrin domain protein | F-CTTCTTCTGTGAGTGTACGT | 313–511 bp | [38] |
| R2-TCCATATCGATCTACTGCGT | ||||
All PCR reactions were composed as follows: 1 unit of Pyrobest Taq DNA polymerase (Takara, Japan), 5 µl 10× Pyrobest Buffer II (Mg2+ Plus), 4 µl of 2.5 mM dNTPs, 5 ng total DNA, 2 µl of each 10 µM primer, and ddH2O was added to bring up the total volume to 50 µl. The amplification of wsp gene was performed as follows: 32 cycles of 94°C for 30 s, 55°C for 30 s and 72°C for 60 s, followed by a final extension at 72°C for 7 min. The amplification of ank2 gene was performed as follows: 35 cycles of 94°C for 30 s, 52°C for 30 s and 72°C for 60 s, followed by a final extension at 72°C for 7 min. PCR products were resolved by 1% agarose gel electrophoresis and stained with ethidium bromide. PCR products of ten mosquitoes from each population were used for sequencing of wsp and ank2 alleles by chain-termination method (Invitrogen). Sequence analysis was carried out using DNAMAN software. The unique DNA sequences were deposited into GenBank (accession numbers JX050182 - JX050187).
Tetracycline treatment and fluorescence staining
Tetracycline treatment to eliminate Wolbachia from Culex populations was carried out according to published methods [17]. Tetracycline (Amresco) at a concentration of 0.05 mg/ml was used for the treatment through both larval and pupal stages. Eggs were placed on tetracycline water solution to hatch. Surviving larvae were transferred to fresh tetracycline solution every 24 hours. A normal infusion was prepared in parallel and fed to larvae in tetracycline solution. The elimination of Wolbachia was checked by Hoechst 33342 (Sigma) staining [39]. Egg rafts within 1 hour of oviposition were placed in 5 ml fixation buffer (182 mM KCl, 46 mM NaCl, 3 mM CaCl2, 10 mM Tris pH 7.2, 3.7% formaldehyde) and overlaid with 5 ml n-heptane in a 50-ml tube. After incubation at room temperature for 15 min with constant shaking, fixation buffer was replaced with 10 ml methanol. The eggs were incubated at room temperature for 10 min with constant shaking. The n-heptane layer was replaced with 10 ml methanol. The eggs were washed for 2 times with methanol, and stained with Hoechst 33342 (1 µg/ml in PBS) for 15 min before visualization under microscope.
Statistical analysis
Statistical differences in hatching rate among crossing groups were examined using the Student's t-test. Linear regression analysis was conducted to determine the correlation coefficient between hatching rate and percentage of incompatible males. All statistical analyses were carried out using SPSS Statistics 17.0.
Results
Incompatibility exists between Cx. pipiens pallens populations
We first tested the compatibilities between available Cx. pipiens pallens populations collected from three geographic locations in China. Reciprocal crosses among these three field populations were performed as outlined in Table S1. In the mating combinations of NJ and WX populations, no significant incompatibility was detected in either NJ♀×WX♂ or WX♀×NJ♂ cross as compared to NJ♀×NJ♂ and WX♀×WX♂ control groups (Table S1 and Figure 2A). In the mating combinations of TK and WX populations, bi-directional incompatibility was observed, with both TK♀×WX♂ and WX♀×TK♂ crosses having significantly reduced average hatching rate as compared to TK♀×TK♂ and WX♀×WX♂ control groups (Table S1 and Figure 2B). In the WX♀×TK♂ group, 14 larvae hatched out of 21 egg rafts, making the average hatching rate 0.005±0.026. The incompatibility was more pronounced in the TK♀×WX♂ group, with 2 larvae hatched out of a total of 18 egg rafts, resulting in an average hatching rate of 0.001±0.001. In comparison, the hatching rates in the control groups TK♀×TK♂ and WX♀×WX♂ were 0.873±0.028 and 0.856±0.003, respectively.
Figure 2. Reproductive compatibilities between three Cx. pipiens pallens field populations.
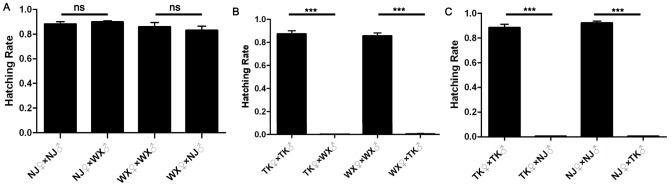
Three Cx. pipiens pallens field populations NJ, WX and TK were used in reciprocal crosses to test their reproductive compatibilities. For each cross, an equal number of virgin females and males were kept in a cage for 48 hours before blood feeding. After oviposition, egg rafts were collected and allowed to hatch. The average hatching rates of these crosses were compared. (A) Mating combinations of NJ and WX populations, (B) Mating combinations of TK and WX populations, (C) Mating combinations of NJ and TK populations. Error bars represent standard errors; ns, non-significant P-value; *** P<0.0001 by t-test.
Similarly, bi-directional incompatibility was detected in the mating combinations of NJ and TK populations (Table S1 and Figure 2C). The average hatching rates for TK♀×NJ♂ and NJ♀×TK♂ crosses were 0.004±0.003 and 0.004±0.002, respectively. In comparison, the average hatching rates were 0.883±0.028 and 0.922±0.015 for TK♀×TK♂ and NJ♀×NJ♂ control groups, respectively.
The incompatibility is not caused by mating failure
The observed bi-directional incompatibility between TK and WX populations and between TK and NJ populations could be a result of mating failure or some post-mating events. To distinguish between these two possibilities, insemination status and embryonic development in incompatible crosses were examined. The female genitalia of Culex mosquitoes include three oval-shaped spermathecal capsules for spermatozoa storage which are connected to vagina through spermathecal ducts. During coitus, some or all of these capsules are filled with seminal fluid from a male and become inflated. The females from incompatible crosses were examined and compared to virgin females. Because three spermathecal capsules from the same individual female had variable sizes before insemination as seen in virgin females, and it was likely that they also varied from mosquito to mosquito, inflation was not always a reliable indicator of successful insemination. Instead, the presence of spermatozoa inside the spermathecal capsules or ovarioles was checked. In all crossing experiments we conducted, the females from both TK♀×NJ♂ and TK♀×WX♂ crosses contained spermatozoa, indicating that they had mated. In addition, the egg rafts were examined 48 hours after oviposition. The eggs from incompatible crosses showed clear embryonic development (Figure S1). Inside the eggs from TK♀×WX♂ cross, there was segment formation along the axis. At the anterior end of the eggs, two red spots representing primitive eyes (stemmata) were visible. Consistent with previous reports, these embryos showed some abnormalities [34]. One evident difference between TK♀×WX♂ cross and compatible crosses was that the incompatible embryos had disorientated bristles. In the eggs from TK♀×NJ♂ cross, similar development was observed. 1–3 (mostly 2) pigmented stemmata were formed. Most stemmata were located in the head region, but some did not seem to have a specific localization. In some eggs, two stemmata were aligned anteroposteriorly instead of being side-by-side. In addition, there was segment formation along the axis in some eggs. Disoriented bristles were also observed in the eggs from TK♀×NJ♂ cross. These observations further confirmed that the females in the incompatible crosses had mated. For those females needed for subsequent mating experiments, only embryonic development inside the eggs was checked to confirm their insemination status.
Mating with incompatible males prevents females from subsequent mating with cognate males
Female monogamy in insects is common but often not absolute. Considering these populations were geographically isolated, it was possible that mating with incompatible males did not preclude subsequent mating of the inseminated females. To test if mating with incompatible males made the females refractory to re-mating, females were retrieved from the incompatible crosses and tested for their ability to mate with cognate males. In the aforementioned crossing experiment, the combination of TK and WX populations displayed higher level of incompatibility than that of TK and NJ populations, so TK and WX populations were chosen in subsequent experiments. TK♀ from TK♀×WX♂ cross and WX♀ from WX♀×TK♂ cross were separated from WX♂ and TK♂, respectively. Each group was equally divided into two subgroups, with one subgroup mixed with cognate males and the other kept alone (Table S2). If subsequent mating could happen, the subgroup mixed with cognate males would become inseminated with both compatible and incompatible spermatozoa and result in higher hatching rates than the subgroup kept separate from cognate males. As shown in Figure 3 and Table S2, the hatching rates in TK♀ subgroup mixed with TK♂ and in TK♀ subgroup kept alone without males were not significantly different (t = −1.013, df = 18, P = 0.324). Similarly, the hatching rates in WX♀ subgroup mixed with WX♂ and in WX♀ subgroup kept alone without males were not significantly different (t = −1.0, df = 6, P = 0.356). These results indicate that both TK♀ and WX♀ became refractory to subsequent mating after they mated with incompatible males.
Figure 3. Effect of subsequent encounter with cognate males on the fecundity of the females retrieved from incompatible crosses.
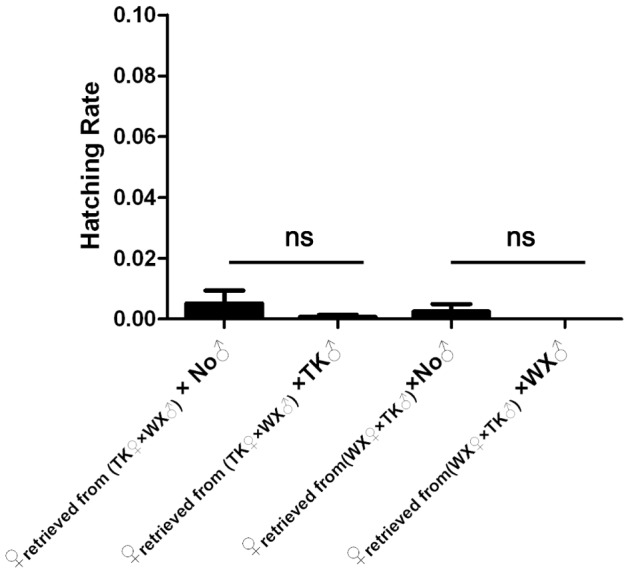
Females were retrieved from the two incompatible crosses as shown in Figure 2B. TK or WX females were equally divided into two subgroups, with one mixed with cognate males and the other kept alone (no male). After a second blood feeding and oviposition, egg rafts were collected and allowed to hatch. The average hatching rates of these subgroups were compared. Error bars represent standard errors; ns, non-significant P-value.
Incompatible males can competitively decrease females mating with compatible males and reduce overall fecundity in a dose-dependent manner
Because these incompatible females and males can successfully mate in the absence of compatible males and produce reduced numbers of offspring, we then tested if incompatible mating would still occur when compatible males were available. We also tested if the reduction in fecundity depended on the number of incompatible males introduced. To that end, two sets of experiments were performed using TK and WX females (Table S3). In each set, an equal number of females were placed in different cages together with no male (blank), equal number of compatible males (positive control), equal number of compatible males plus equal number of incompatible males, or equal number of males plus 3× as many incompatible males. In addition, as in the previous experiment, 48 hours after the first oviposition, egg rafts were checked for embryonic development to ascertain that the females were inseminated. Subsequently, females were collected from each group and divided into two subgroups to be mixed with either no male or cognate males (Table S4). The hatching rates from the second oviposition were compared to determine if mating with mixed male populations made these females refractory to subsequent mating even when only cognate males were available.
The results (Table S3, Figure 4A) show that in the positive control group (TK♀×TK♂) the average hatching rate was 0.861±0.020. When an equal number of WX males were included in the cage [TK♀×(TK♂+WX♂)], the average hatching rate dropped to 0.211±0.071. This value was further decreased to 0.063±0.039 in the group that included 3× as many WX males [TK♀×(TK♂+3×WX♂)]. In comparison, the average hatching rate of TK♀ mixed with only WX♂ was 0.004±0.003 and TK♀ kept alone produced no larva. These data indicate TK♀ mated with TK♂ or WX♂ in the presence of mixed male population, i.e., even in the presence of TK♂, mating between TK♀ and WX♂ still occurred, which resulted in reduced overall fecundity of the groups. The extent of fecundity reduction correlated with the ratio of WX♂ to TK♂.
Figure 4. Incompatible males competitively suppress reproduction in a dose-dependent manner.

Females were mixed with cognate males and an increasing number of incompatible males. They were given 48 hours before blood feeding. The average hatching rates of these crosses were compared. Subsequently, females were retrieved and divided equally into two subgroups, with one mixed with cognate males and the other kept alone. The average hatching rates of these subgroups were compared. (A) TK♀ mixed with TK♂ and WX♂; (B) WX♀ mixed with WX♂ and TK♂; (C) TK♀ retrieved from A were mixed with TK♂ or kept alone; (D) WX♀ retrieved from B were mixed with WX♂ or kept alone. Error bars represent standard errors; ns, non-significant P-value; *** P<0.0001; * P<0.05 by t-test.
Similarly, in parallel experiment with WX♀ (Table S3, Figure 4B), the average hatching rate of WX♀ was reduced from 0.903±0.013 in the positive control group (WX♀×WX♂) to 0.607±0.066 in the group with an equal number of competing TK♂ included [WX♀×(WX♂+TK♂)] and further to 0.407±0.077 in the group with 3× competing TK♂ included [WX♀×(WX♂+3×TK♂)]. The average hatching rate of WX♀ mixed with only TK♂ (WX♀×TK♂) was 0.001±0.001. WX♀ kept alone produced no larva. These data also indicate that the frequency of WX♀×TK♂ mating in the presence of mixed male population can also be increased with an increasing TK♂∶WX♂ ratio.
As shown in Figure 4C, after retrieved TK♀ were mixed with TK♂ or no male, these two subgroups produced similar hatching rates in their second oviposition. Similarly, subsequent mixing of retrieved WX♀ with WX♂ did not significantly increase the hatching rates in the second oviposition compared to the subgroups of retrieved WX♀ kept alone (Figure 4D). These results indicate both TK♀ and WX♀ became refractory to subsequent mating after they mated with mixed male populations. This was similar to the scenario in which their first mating was with incompatible males only. Polyandry was not common or completely absent for these female populations. In addition, the average hatching rates in their subsequent ovipositions correlated with the hatching rates in their first oviposition. A higher number of incompatible males during the first mating event resulted in lower fecundity during the first and subsequent gonotrophic cycles.
The correlation between hatching rate and the proportion of incompatible males was plotted for both TK♀ and WX♀. As shown in Figure 5, based on the curves, the fecundity of TK♀ could be diminished when inundated by excess WX♂. 9× WX♂ can achieve nearly complete suppression of TK♀ fecundity. On the other hand, the maximal fecundity reduction of WX♀ by TK♂ is around 90% in one generation. The r2 value is 0.92 (P<0.05) for TK♀ curve (Figure 5A), and 0.9363 (P<0.05) for WX♀ curve (Figure 5B).
Figure 5. Correlation between group average hatching rate and the percentage of incompatible males.
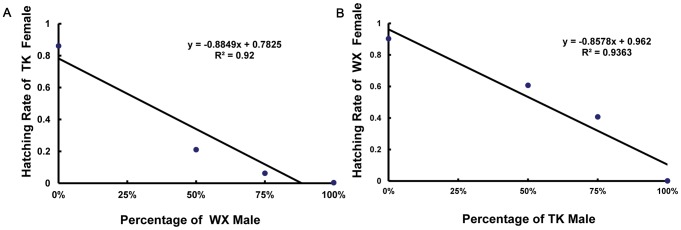
The average hatching rate of a group was plotted against the percentage of incompatible males which was calculated by dividing the number of incompatible males by the total number of males (incompatible and cognate). (A) TK females with TK and WX males; (B) WX females with WX and TK males.
The incompatibility is preserved in the offspring from the crosses
Since the incompatibility between TK and WX populations is not absolute, when incompatible males are released to suppress a target population, some hybrids will be generated. If these hybrids are compatible with the released males, then subsequent male release will help these hybrids to reproduce. This would pose a danger of gradually replacing the target population with a compatible hybrid population or creating a balance between the original target population and the hybrid population. In these cases, the populations are not effectively suppressed by male release. To test if the use of naturally incompatible populations as a control strategy is sustainable, the compatibility between the hybrids and TK and WX populations were measured.
In the case of releasing WX♂ to suppress TK population, both male and female F1 offspring will be generated from TK♀×WX♂ cross, designated as F1♀(TK♀×WX♂) and F1♂(TK♀×WX♂). These hybrids will encounter TK♀, TK♂ and WX♂, resulting in six possible mating combinations. Three extra mating combinations WX♀×F1♂(TK♀×WX♂), WX♀×WX♂ and WX♀×TK♂ that would not accompany this population suppression measure were also included in this set to provide more information about the crossing type of F1♂(TK♀×WX♂). These nine mating combinations were compared for fecundity. As shown in Table S5 and Figure 6A, the mating combinations F1♀(TK♀×WX♂)×F1♂(TK♀×WX♂) and F1♀(TK♀×WX♂)×TK♂ had comparably high hatching rates, while F1♀(TK♀×WX♂)×WX♂ produced no larva. These indicate F1♀(TK♀×WX♂) maintained the crossing type of TK♀. On the other hand, the average hatching rates of TK♀×F1♂(TK♀×WX♂) and TK♀×TK♂ were comparably high, while the hatching rates in WX♀×F1♂(TK♀×WX♂) and WX♀×TK♂ crosses were comparably low, indicating that F1♂(TK♀×WX♂) maintained the crossing type of TK♂. These results demonstrate that in TK♀×WX♂ cross, both male and female F1 offspring maintained their maternal crossing type.
Figure 6. Reproductive compatibilities of F1 offspring from incompatible crosses.
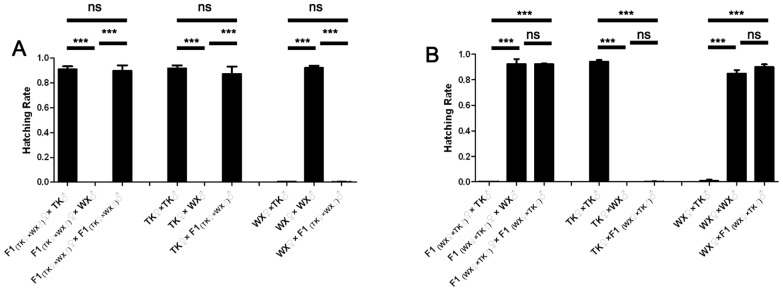
F1 offspring from TK and WX crosses were mixed with TK, WX or the F1. The hatching rates were compared. (A) F1 from TK♀×WX♂ cross; (B) F1 from WX♀×TK♂ cross. Error bars represent standard errors; ns, non-significant P-value; *** P<0.0001 by t-test.
Reciprocally, the strategy of using TK♂ to suppress WX population was tested for sustainability. To test the crossing type of F1♀(WX♀×TK♂) and F1♂(WX♀×TK♂) generated from the cross between WX♀ and TK♂, nine possible mating combinations were compared for hatching rate. As shown in Table S5 and Figure 6B, the average hatching rates of F1♀(WX♀×TK♂)×F1♂(WX♀×TK♂) and F1♀(WX♀×TK♂)×WX♂ were comparably high, while the hatching rate was low for F1♀(WX♀×TK♂)×TK♂ group. These indicate F1♀(WX♀×TK♂) maintained the crossing type of WX♀. On the other hand, the average hatching rates of WX♀×F1♂(WX♀×TK♂) and WX♀×WX♂ were comparably high, while the average hatching rates of TK♀×WX♂ and TK♀×F1♂(WX♀×TK♂) were comparably low. These indicate F1♂(WX♀×TK♂) maintained the crossing type of WX♂. In WX♀×TK♂ cross, both male and female F1 offspring maintained their maternal crossing type.
Taken together, when using incompatible males to suppress a target Cx. pipiens pallens population, hybrid offspring may be generated if the incompatibility is not absolute. However, these hybrids maintain their maternal crossing types. This phenomenon indicates that both original target population and inadvertently generated hybrids are subject to suppression by the released incompatible males, suggesting that this control strategy is sustainable.
The three Cx. pipiens pallens populations are all infected with Wolbachia
The crossing types observed in these Cx. pipiens pallens populations were inherited maternally, suggesting the possibility that Wolbachia was the causal factor. To detect possible infections of Wolbachia, PCR was carried out using total DNA extracted from WX, NJ and TK populations. The primers were selected to amplify the wsp gene [36], [37] and the ank2 gene [38] according to published studies. For each population, 100% infection rate was detected (30 positive out of 30 tested mosquitoes for each population). The wsp sequence can distinguish supergroup A, wPip strains of supergroup B and wCauB strains of supergroup B. The ank2 sequence can further distinguish five phylogenetic groups (wPip-I to wPip-V) of wPip strains [38]. PCR results show that using the generic wF-wR primer pair a fragment around 600 bp was amplified from all three natural populations of Cx. pipiens pallens (Figure S2A). A fragment around 500 bp was amplified from all three populations using the wPip-specific wBpipF-wR primer pair. Neither wAF-wR primer pair (specific for supergroup A) nor wBcauBF-wR primer pair (specific for wCauB of supergroup B) generated any amplification product. These results are consistent with previous reports that most mosquito-infecting Wolbachia are wPip strains of supergroup B. This was confirmed by subsequent sequencing analysis, which also revealed that the wsp genes from these three populations are identical (GenBank accession numbers JX050185 - JX050187). PCR and sequencing analysis of ank2 gene revealed that NJ population was infected by Wolbachia of wPip-III group (GenBank accession number JX050182), while WX and TK populations were both infected by Wolbachia of wPip-IV group (GenBank accession numbers JX050183 and JX050184). The ank2 genes from WX and TK populations are identical (Figure S2B and Figure S3).
The incompatibilities between different Cx. pipiens pallens populations are caused by Wolbachia
To confirm that the observed incompatibilities were caused by Wolbachia, the mosquitoes were treated with tetracycline [17]. The elimination of Wolbachia was checked by Hoechst 33342 staining [39]. As shown in Figure S4, eggs from untreated mosquitoes had strong fluorescence at both anterior and posterior ends, indicating Wolbachia were concentrated at these poles. In contrast, eggs from tetracycline-treated mosquitoes had even distribution of background fluorescence. No strong fluorescence was observed around the micropyle or at the posterior end. These results indicate that Wolbachia were removed from these mosquito populations. In addition, tetracycline-treated strains all tested negative for Wolbachia by PCR using wsp-specific primers wF and wR (data not shown). Crossing experiments were carried out using both Wolbachia-positive and Wolbachia-negative populations. WX females were crossed with Wolbachia-positive TK males (TK♂), Wolbachia-negative TK males (TKtet♂) and WX males. Similarly, TK females were crossed with Wolbachia-positive WX males (WX♂), Wolbachia-negative WX males (WXtet♂) and TK males. As shown in Figure 7, the hatching rate of WX♀×TKtet♂ cross was not significantly different from that of WX♀×WX♂ cross, but was significantly higher than that of WX♀×TK♂ cross. The hatching rate of TK♀×WXtet♂ cross was not significantly different from that of TK♀×TK♂ cross, but was significantly higher than that of TK♀×WX♂ cross. These results indicate that the bi-directional incompatibility between TK and WX populations is dependent on the presence of Wolbachia, i.e., it is Wolbachia-induced CI.
Figure 7. Elimination of Wolbachia rendered TK and WX populations fully compatible.
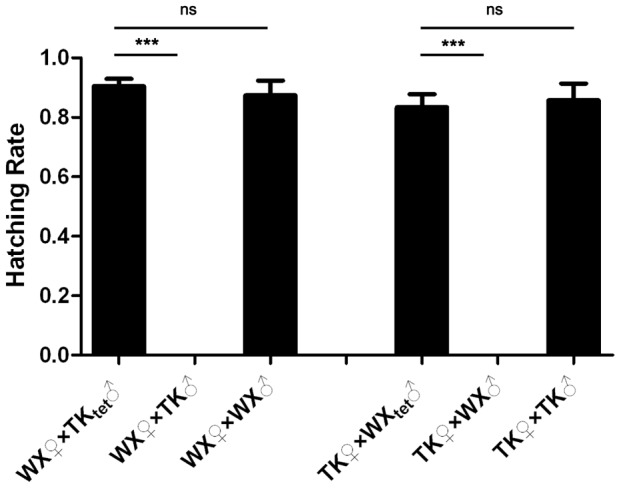
WX females were crossed with an equal number of tetracycline-treated TK males (TKtet♂), untreated TK males (TK♂) or WX males (WX♂). TK females were crossed with an equal number of tetracycline-treated WX males (WXtet♂), untreated WX males (WX♂) or TK males (TK♂). Hatching rates were calculated for these crosses. Error bars represent standard errors; ns, non-significant P-value; *** P<0.0001 by t-test.
Accession numbers
The GenBank accession numbers for sequences mentioned in the paper are ankyrin domain protein ank2 genes of Wolbachia in Nanjing, Tangkou and Wuxi populations of Cx. pipiens pallens (JX050182 - JX050184), and surface protein precursor wsp genes of Wolbachia in Nanjing, Tangkou and Wuxi populations of Cx. pipiens pallens (JX050185 - JX050187).
Discussion
Biological control is a low-pollution component of integrated pest management. A variety of strategies have been developed to suppress populations of insect pests. Taking advantage of many insects' monogamous mating behavior, SIT uses sterile male insects to compete with normal males for their mates, which results in some females producing no or a reduced number of offspring. Two common methods to produce sterile male insects are irradiation and chemical treatments. SIT has been reported to be successful in a number of trials [13], [40]–[42]. One potential problem with SIT is the dosage of radiation or sterilizing agent used to treat male insects. Insufficient dosages might result in the release of fertile males while overtreatment might damage the males so much that they cannot compete with normal male insects. In addition, the residual chemicals in treated insects could potentially cause pollution to the environment and harm untargeted insect species [43], [44].
Alternatively, IIT exploits incompatibilities between insect populations and uses incompatible male insects to compete with compatible insects for their mates. Because IIT relies on the genetic traits of incompatible populations, it could be more reproducible than SIT. Most IITs are based on Wolbachia-induced cytoplasmic incompatibility. Infection with Wolbachia has been reported in many species, including some disease vectors such as Aedes albopictus and Culex quinquefasciatus ( = Cx. pipiens fatigans) [15], [21], [30]. For naturally uninfected insects, artificial Wolbachia infection would generate an incompatible strain that could be used to suppress target populations [28]. Introgression using Wolbachia-infected females and Wolbachia-negative target population males can also be used to generate an incompatible strain. For Wolbachia-positive target populations, a common approach is introgression with females from another strain infected with an incompatible Wolbachia. Sometimes, antibiotic treatment to eliminate Wolbachia from target strain is needed to facilitate the introgression process [29]. The advantages of this strategy is the generation of insect population with a desired host genetic background (often to match that of the target population) to increase the likelihood of survival in the target environment and successful competition with native males. However, both artificial infection and introgression can be technically difficult. Even with successful establishment of an incompatible strain, because of the placement of Wolbachia in a different host genetic background, the long-term stability of this new symbiosis needs to be tested.
In our study, we exploited incompatibilities between naturally existent populations from different geographic locations to suppress mosquito populations. This spares the time and effort to create incompatible populations. Another advantage is that the stability of Wolbachia can be assured since this symbiotic relationship has been naturally selected for a long time. IIT involves repeated release of incompatible males that can competitively inseminate females in a target population. Since these males are not meant to have offspring, any potential adverse effect of their genetic makeup on their fitness in the target environment would not be an issue so long as these males can live long enough to mate and are sexually competitive. We tested the mating behavior of three populations collected from different locations. When mixed with males of a different population, females became inseminated, as spermatozoa were present in the spermathecal capsules of these females. TK females mixed with NJ males or WX males laid comparable number of egg rafts, although these eggs had much lower hatching rates. NJ females or WX females also laid comparable numbers of egg rafts when mixed with TK males compared to when they were mixed with males of their own populations. Egg rafts with low hatching rate also showed clear embryonic development. All these data indicate that these males and females from different populations could successfully mate. Geographical and chronic isolation did not appear to create a mating barrier between these populations.
IIT works effectively only if the incompatible males are competitive enough. Although these mosquito populations could mate with each other, we also tested if the females had strong preference of their cognate males in the presence of both cognate and incompatible males using a mating competition experiment. Our results indicated significant mating preference was not observed. In Figure 5, a linear curve would indicate random mating, while a strong preference of cognate males would result in an arch curve with the median value greater than the average of the two extrema. Both curves are basically linear, supporting that these females chose their mates randomly. With an increasing ratio of incompatible to cognate males, the number of offspring decreased. Our results are consistent with previous reports from other groups that assortative mating usually does not occur in natural populations when numerous Wolbachia strains coexist within those populations [45]. These data suggest that although Wolbachia may cause CI in mosquitoes and skew survival in favor of those embryos that harbor the same or compatible Wolbachia, it does not affect the mate selection significantly. Instead, insects including mosquitoes select their mates based on their own genetic traits [46], [47]. The selection of suitable populations for mosquito control would depend on finding a population whose genome is compatible with that of target population, while also having a Wolbachia infection that induces cytoplasmic incompatibility in that context.
Another factor that influences IIT success is the monogamous mating behavior of females. After female mosquitoes are inseminated, they usually become refractory to re-mating. This phenomenon has been attributed to the effects of proteins in the seminal fluid received from male mosquitoes. But female monogamy is not always absolute. Polyandry has been reported in a number of mosquito species, such as Aedes aegypti and Culex tarsalis. The likelihood of female re-mating increases after these females go through gonotrophic cycles [48], [49], possibly due to the waning of seminal fluid proteins. One potential obstacle to the use of mosquitoes from different geographic locations to suppress target populations is the incompatibility between ligands in seminal fluid of one population and receptors in females of the other. In this scenario, insemination would not prevent the female mosquitoes from mating with reproductively compatible males. In our study, TK female mosquitoes became refractory to re-mating after being inseminated by WX or NJ male mosquitoes, even after these females had gone through a gonotrophic cycle, indicating seminal fluid proteins of WX or NJ population can act on receptors of TK population to cause post-coital behavioral changes in TK females. These results suggest that the seminal fluid proteins and their receptors are conserved enough between these populations, or their interactions are flexible enough to tolerate certain mutations. Further studies are needed to reveal the interactions between seminal fluid proteins and their receptors of different mosquito populations. Potentially, there are other factors causing female monogamy. If so, these factors seem to be functional across different populations.
Population suppression usually requires repeated effort. It is important for IIT to work for many generations. We tested if hybrids of incompatible populations became compatible with their parental populations. Our data indicate the incompatibility between different mosquito populations is hereditarily stable. This suggests that using males from incompatible natural populations to suppress a target population is sustainable. Although in our study, F1 offspring from incompatible crosses maintained the maternal crossing types, it should be noted that the preservation of maternal crossing type is not always the case. For example, F1♀ from the cross of bi-directionally incompatible Cx. quinquefasciatus strains Bei and Pel became fully compatible with Pel males [50]. This cautions us that when choosing incompatible populations, the crossing types of F1 hybrids should be carefully tested as well.
Although the climates at the sites where these mosquitoes originated from are different due to their latitudes, etc., these mosquitoes did not demonstrate significant behavioral differences in the same artificial environment of our insectary. More trials are needed to determine how they would behave and interact in semi-field and field trials. It remains to be tested whether incompatible male mosquitoes released into different natural environments will remain sexually competitive, and reduce the fecundity of local females.
Both maternal inheritance of crossing types and elimination of incompatibility by tetracycline treatment indicate that the observed incompatibility is caused by Wolbachia. To test if these mosquito populations are infected with different strains of Wolbachia, specific DNA fragments were PCR amplified from three mosquito populations. We sequenced wsp and ank2 genes, two specific genes commonly used to type Wolbachia. However, sequence analysis shows that wsp gene is identical in these three mosquito populations. The ank2 gene is identical between TK and WX populations, while ank2 gene of NJ population is different from TK or WX population. These Wolbachia are all wPip strains of supergroup B. It is currently unclear how they differ from each other. The definitive answer would require more sequencing or other typing tests which are beyond the scope of the current study. ANK genes (including ank2) were initially proposed to be involved in CI [50]. This hypothesis was challenged by subsequent analysis that failed to find any association of ANK polymorphism and CI [38]. Our data show NJ and WX populations are bi-directionally compatible, yet their ank2 genes are different. On the other hand, TK and WX populations are bi-directionally incompatible, yet their ank2 genes are identical (or at least the sequenced segments). These provide support to the view that homology between these typing markers does not correlate to the level of CI, suggesting molecular markers that are polymorphic and more closely associated with CI factors are yet to be found [38], [51]. Nuclear contribution to incompatibility has also been reported [50]. Being isolated from each other in the wild, these mosquitoes might have accumulated enough mutations in their genomes and/or even mitochondrial DNA to make some crosses incompatible. These differences in the host could directly cause incompatibility or more likely cause incompatibility by modulating Wolbachia gene expression. Consequently, although the incompatibilities between these populations are dependent on Wolbachia, contribution of host genomes cannot be ruled out at present. Our observation of embryonic development of incompatible eggs is in accordance with previously reported Wolbachia-mediated CI. It has been reported that incompatible eggs from two Wolbachia-positive populations have higher level of development than incompatible eggs from Wolbachia-negative females (which are usually undistinguishable from unfertilized eggs) [34]. We observed stemmata, bristles and segmentation in both TK♀×WX♂ and TK♀×NJ♂ crosses. This would be consistent with the multi-factorial mod resc model: even incompatible Wolbachia provides partial rescue in the eggs [15].
Although incompatibility between insect populations (including CI) is not fully understood, its applicability is promising. Our study proves that IIT is a feasible control strategy for Cx. pipiens pallens. The use of naturally occurring populations without genetic manipulation will save time and effort, and require less technical knowhow. This is an advantage for many less developed regions that deserves consideration. The wide distribution of mosquitoes in varied environments may be turned against them because it provides rich diversity in incompatibility; so that it is likely to find naturally incompatible and sexually competitive strains for many target populations. The conclusions from our study on Cx. pipiens pallens might offer reference to control measures of other mosquitoes, as well.
Supporting Information
Embryonic development of eggs from incompatible crosses. Eggs from incompatible crosses TK♀×WX♂ and TK♀×NJ♂ were compared to unfertilized eggs and eggs from compatible cross TK♀×TK♂ using microscopy. (A) unfertilized egg of TK♀ 48 hours after oviposition, (B) egg from TK♀×NJ♂ 48 hours after oviposition, (C) egg from TK♀×WX♂ 48 hours after oviposition, (D) egg from compatible cross (TK♀×TK♂) 36 hours after oviposition, (E) Egg from TK♀×TK♂ 48 hours after oviposition. Both TK♀×WX♂ and TK♀×NJ♂ eggs displayed signs of embryonic development. In (E), a first-instar larva already released into water, leaving behind an empty eggshell with an opened operculum.
(TIF)
Detection of Wolbachia in NJ, WX and TK populations. (A) PCR amplification of Wolbachia wsp gene. Four PCR reactions were performed in parallel for each DNA specimen (Lanes 1, 4, 7, 10 for TK population; Lanes 2, 5, 8, 11 for NJ population; Lanes 3, 6, 9, 12 for WX population). Lanes 1–3 were amplified with primers wF and wR, Lanes 4–6 were amplified with primers wAF and wR, Lanes 7–9 were amplified with primers wBpipF and wR, Lanes 10–12 were amplified with primers wBcauBF and wR. (B) PCR amplification of Wolbachia ank2 gene. Lane 1: TK population, Lane 2: NJ population, Lane 3: WX population.
(TIF)
Alignment of ank2 sequences from NJ, WX and TK populations. Alignment of amplified fragments of Wolbachia ank2 genes from NJ, WX and TK populations with two published sequences ank2-c (GenBank accession number AM397070.1) and ank2-d (GenBank accession number AM397071.1). NJ sequence shares 100% homology with ank2-c, WX and TK sequences are identical and share 100% homology with ank2-d.
(TIF)
Elimination of Wolbachia from TK, WX and NJ populations by tetracycline treatment. Eggs of tetracycline-treated and untreated mosquitoes were stained with Hoechst 33342 and visualized by fluorescence microscopy. (A) Wolbachia are concentrated at both anterior and posterior ends of untreated eggs indicated by strong fluorescence. (B) The same egg as in (A) under visible light. (C) Tetracycline-treated egg shows no Wolbachia distribution at either end of the egg. (D) The same egg as in (C) under visible light. Shown here are results of TK population. Treatment of WX and NJ populations had similar results. Scale bar, 0.1 mm.
(TIF)
Reciprocal crosses among Cx. pipiens pallens field populations.
(PDF)
Second round mating combinations of females retrieved from incompatible crosses.
(PDF)
Mating combinations of TK and WX females with both cognate and an increasing number of incompatible males.
(PDF)
Second round mating combinations of females retrieved from mixed male populations.
(PDF)
Mating combinations of F1 offspring of TK and WX crosses.
(PDF)
Crosses of tetracycline-treated and untreated WX and TK populations.
(PDF)
Acknowledgments
We thank Zhikuan Jiang, Chao Chen, Maoqing Gong and Huayun Zhou for their help with mosquito collection, and Lei Ma for maintaining the insectary.
Funding Statement
This work was supported by the Natural Science Foundation of Jiangsu Higher Education Institutions (Grant 07KJD180137 to DZ) and in part by the Priority Academic Development Program of Jiangsu Higher Education Institutions. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.WHO (2010) Working to Overcome the Global Impact of Neglected Tropical Diseases: First WHO Report on Neglected Tropical Diseases. Geneva: World Health Organization.
- 2.WHO (2010) World Malaria Report 2010. Geneva: World Health Organization.
- 3. Hemingway J, Ranson H (2000) Insecticide resistance in insect vectors of human disease. Annu Rev Entomol 45: 371–391. [DOI] [PubMed] [Google Scholar]
- 4. Martinez FD, Trejo-Acevedo A, Betanzos AF, Espinosa-Reyes G, Alegria-Torres JA, et al. (2012) Assessment of DDT and DDE levels in soil, dust, and blood samples from Chihuahua, Mexico. Arch Environ Contam Toxicol 62: 351–358. [DOI] [PubMed] [Google Scholar]
- 5. Wassie F, Spanoghe P, Tessema DA, Steurbaut W (2012) Exposure and health risk assessment of applicators to DDT during indoor residual spraying in malaria vector control program. J Expo Sci Environ Epidemiol 22: 549–558. [DOI] [PubMed] [Google Scholar]
- 6. Moreira LA, Wang J, Collins FH, Jacobs-Lorena M (2004) Fitness of anopheline mosquitoes expressing transgenes that inhibit Plasmodium development. Genetics 166: 1337–1341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Kokoza V, Ahmed A, Cho WL, Jasinskiene N, James AA, et al. (2000) Engineering blood meal-activated systemic immunity in the yellow fever mosquito, Aedes aegypti . Proc Natl Acad Sci U S A 97: 9144–9149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Moreira LA, Iturbe-Ormaetxe I, Jeffery JA, Lu G, Pyke AT, et al. (2009) A Wolbachia symbiont in Aedes aegypti limits infection with dengue, Chikungunya, and Plasmodium . Cell 139: 1268–1278. [DOI] [PubMed] [Google Scholar]
- 9. Hoffmann AA, Montgomery BL, Popovici J, Iturbe-Ormaetxe I, Johnson PH, et al. (2011) Successful establishment of Wolbachia in Aedes populations to suppress dengue transmission. Nature 476: 454–457. [DOI] [PubMed] [Google Scholar]
- 10. Blagrove MS, Arias-Goeta C, Failloux AB, Sinkins SP (2012) Wolbachia strain wMel induces cytoplasmic incompatibility and blocks dengue transmission in Aedes albopictus . Proc Natl Acad Sci U S A 109: 255–260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Walker T, Johnson PH, Moreira LA, Iturbe-Ormaetxe I, Frentiu FD, et al. (2011) The wMel Wolbachia strain blocks dengue and invades caged Aedes aegypti populations. Nature 476: 450–453. [DOI] [PubMed] [Google Scholar]
- 12. Wise de Valdez MR, Nimmo D, Betz J, Gong HF, James AA, et al. (2011) Genetic elimination of dengue vector mosquitoes. Proc Natl Acad Sci U S A 108: 4772–4775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Alphey L, Benedict M, Bellini R, Clark GG, Dame DA, et al. (2010) Sterile-insect methods for control of mosquito-borne diseases: an analysis. Vector Borne Zoonotic Dis 10: 295–311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Harris AF, Nimmo D, McKemey AR, Kelly N, Scaife S, et al. (2011) Field performance of engineered male mosquitoes. Nat Biotechnol 29: 1034–1037. [DOI] [PubMed] [Google Scholar]
- 15. Werren JH (1997) Biology of Wolbachia . Annu Rev Entomol 42: 587–609. [DOI] [PubMed] [Google Scholar]
- 16. Yen JH, Barr AR (1971) New hypothesis of the cause of cytoplasmic incompatibility in Culex pipiens L. Nature 232: 657–658. [DOI] [PubMed] [Google Scholar]
- 17. Yen JH, Barr AR (1973) The etiological agent of cytoplasmic incompatibility in Culex pipiens . J Invertebr Pathol 22: 242–250. [DOI] [PubMed] [Google Scholar]
- 18. Hertig M, Wolbach SB (1924) Studies on Rickettsia-like micro-organisms in insects. J Med Res 44: 329–374. [PMC free article] [PubMed] [Google Scholar]
- 19. Hilgenboecker K, Hammerstein P, Schlattmann P, Telschow A, Werren JH (2008) How many species are infected with Wolbachia?–A statistical analysis of current data. FEMS Microbiol Lett 281: 215–220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Stouthamer R, Breeuwer JA, Hurst GD (1999) Wolbachia pipientis: microbial manipulator of arthropod reproduction. Annu Rev Microbiol 53: 71–102. [DOI] [PubMed] [Google Scholar]
- 21. Sinkins SP (2004) Wolbachia and cytoplasmic incompatibility in mosquitoes. Insect Biochem Mol Biol 34: 723–729. [DOI] [PubMed] [Google Scholar]
- 22. Laven H (1967) Eradication of Culex pipiens fatigans through cytoplasmic incompatibility. Nature 216: 383–384. [DOI] [PubMed] [Google Scholar]
- 23. Boller EF, Russ K, Vallo V, Bush GL (1976) Incompatible races of European cherry fruit fly Rhagoletis cerasi (Diptera: Tephritidae): their origin and potential use in biological control. Entomol Exp Appl 20: 237–247. [Google Scholar]
- 24. Brower JH (1979) Suppression of laboratory populations of Ephestia cautella (Walker) (Lepidoptera: Pyralidae) by release of males with cytoplasmic incompatibility. J Stored Prod Res 15: 1–4. [Google Scholar]
- 25. Arunachalam N, Curtis CF (1985) Integration of radiation with cytoplasmic incompatibility for genetic control in the Culex pipiens complex (Diptera: Culicidae). J Med Entomol 22: 648–653. [DOI] [PubMed] [Google Scholar]
- 26. Shahid MA, Curtis CF (1987) Radiation sterilization and cytoplasmic incompatibility in a “tropicalized” strain of the Culex pipiens complex (Diptera: Culicidae). J Med Entomol 24: 273–274. [DOI] [PubMed] [Google Scholar]
- 27. Sharma VP, Subbarao SK, Adak T, Razdan RK (1979) Integration of gamma irradiation and cytoplasmic incompatibility in Culex pipiens fatigans . J Med Entomol 15: 155–156. [Google Scholar]
- 28. Zabalou S, Riegler M, Theodorakopoulou M, Stauffer C, Savakis C, et al. (2004) Wolbachia-induced cytoplasmic incompatibility as a means for insect pest population control. Proc Natl Acad Sci USA 101: 15042–15045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Brelsfoard CL, Sechan Y, Dobson SL (2008) Interspecific hybridization yields strategy for South Pacific filariasis vector elimination. PLoS Negl Trop Dis 2: e129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Atyame CM, Pasteur N, Dumas E, Tortosa P, Tantely ML, et al. (2011) Cytoplasmic incompatibility as a means of controlling Culex pipiens quinquefasciatus mosquito in the islands of the south-western Indian Ocean. PLoS Negl Trop Dis 5: e1440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Chambers EW, Hapairai L, Peel BA, Bossin H, Dobson SL (2011) Male mating competitiveness of a Wolbachia-introgressed Aedes polynesiensis strain under semi-field conditions. PLoS Negl Trop Dis 5: e1271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Hancock PA, Sinkins SP, Godfray HC (2011) Strategies for introducing Wolbachia to reduce transmission of mosquito-borne diseases. PLoS Negl Trop Dis 5: e1024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Chen L, Zhong D, Zhang D, Shi L, Zhou G, et al. (2010) Molecular ecology of pyrethroid knockdown resistance in Culex pipiens pallens mosquitoes. PLoS One 5: e11681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Duron O, Weill M (2006) Wolbachia infection influences the development of Culex pipiens embryo in incompatible crosses. Heredity (Edinb) 96: 493–500. [DOI] [PubMed] [Google Scholar]
- 35. Collins FH, Mendez MA, Rasmussen MO, Mehaffey PC, Besansky NJ, et al. (1987) A ribosomal RNA gene probe differentiates member species of the Anopheles gambiae complex. Am J Trop Med Hyg 37: 37–41. [DOI] [PubMed] [Google Scholar]
- 36. Braig HR, Zhou W, Dobson SL, O'Neill SL (1998) Cloning and characterization of a gene encoding the major surface protein of the bacterial endosymbiont Wolbachia pipientis . J Bacteriol 180: 2373–2378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Zhou W, Rousset F, O'Neil S (1998) Phylogeny and PCR-based classification of Wolbachia strains using wsp gene sequences. Proc R Soc Lond B Biol Sci 265: 509–515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Duron O, Boureux A, Echaubard P, Berthomieu A, Berticat C, et al. (2007) Variability and expression of ankyrin domain genes in Wolbachia variants infecting the mosquito Culex pipiens . J Bacteriol 189: 4442–4448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Rasgon JL, Scott TW (2003) Wolbachia and cytoplasmic incompatibility in the California Culex pipiens mosquito species complex: parameter estimates and infection dynamics in natural populations. Genetics 165: 2029–2038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Vanderplank FL (1947) Experiments in the hybridization of tsetse flies (Glossina, Diptera) and the possibility of a new method of control. Transactions of the Royal Entomological Society (London) 98: 1–18. [Google Scholar]
- 41. Baumhover AH, Graham AJ, Bitter BA, Hopkins DE, New WD, et al. (1955) Screw-worm control through release of sterilized flies. J Econ Entomol 48: 462–466. [Google Scholar]
- 42. Lindquist DA, Abusowa M, Hall MJ (1992) The New World screwworm fly in Libya: a review of its introduction and eradication. Med Vet Entomol 6: 2–8. [DOI] [PubMed] [Google Scholar]
- 43. Lofgren CS, Dame DA, Breeland SG, Weidhaas DE, Jeffery G, et al. (1974) Release of chemosterilized males for the control of Anopheles albimanus in El Salvador. 3. Field methods and population control. Am J Trop Med Hyg 23: 288–297. [DOI] [PubMed] [Google Scholar]
- 44. Bracken GK, Dondale CD (1972) Fertility and survival of Achaearanea tepidariorum (Araneida: Theridiidae) on a diet of chemosterilized mosquitoes. Can Entomol 104: 1709–1712. [Google Scholar]
- 45. Atyame CM, Duron O, Tortosa P, Pasteur N, Fort P, et al. (2011) Multiple Wolbachia determinants control the evolution of cytoplasmic incompatibilities in Culex pipiens mosquito populations. Mol Ecol 20: 286–298. [DOI] [PubMed] [Google Scholar]
- 46. Thistle R, Cameron P, Ghorayshi A, Dennison L, Scott K (2012) Contact Chemoreceptors Mediate Male-Male Repulsion and Male-Female Attraction during Drosophila Courtship. Cell 149: 1140–1151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Arbuthnott D (2009) The genetic architecture of insect courtship behavior and premating isolation. Heredity (Edinb) 103: 15–22. [DOI] [PubMed] [Google Scholar]
- 48. Young ADM, Downe AER (1983) Influence of mating on sexual receptivity and oviposition in the mosquito, Culex tarsalis . Physiological Entomology 8: 213–217. [Google Scholar]
- 49. Williams RW, Berger A (1980) The relation of female polygamy to gonotrophic activity in the ROCK strain of Aedes aegypti . Mosquito News 40: 597–604. [Google Scholar]
- 50. Sinkins SP, Walker T, Lynd AR, Steven AR, Makepeace BL, et al. (2005) Wolbachia variability and host effects on crossing type in Culex mosquitoes. Nature 436: 257–260. [DOI] [PubMed] [Google Scholar]
- 51. Duron O, Raymond M, Weill M (2010) Many compatible Wolbachia strains coexist within natural populations of Culex pipiens mosquito. Heredity (Edinb) 106: 986–993. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Embryonic development of eggs from incompatible crosses. Eggs from incompatible crosses TK♀×WX♂ and TK♀×NJ♂ were compared to unfertilized eggs and eggs from compatible cross TK♀×TK♂ using microscopy. (A) unfertilized egg of TK♀ 48 hours after oviposition, (B) egg from TK♀×NJ♂ 48 hours after oviposition, (C) egg from TK♀×WX♂ 48 hours after oviposition, (D) egg from compatible cross (TK♀×TK♂) 36 hours after oviposition, (E) Egg from TK♀×TK♂ 48 hours after oviposition. Both TK♀×WX♂ and TK♀×NJ♂ eggs displayed signs of embryonic development. In (E), a first-instar larva already released into water, leaving behind an empty eggshell with an opened operculum.
(TIF)
Detection of Wolbachia in NJ, WX and TK populations. (A) PCR amplification of Wolbachia wsp gene. Four PCR reactions were performed in parallel for each DNA specimen (Lanes 1, 4, 7, 10 for TK population; Lanes 2, 5, 8, 11 for NJ population; Lanes 3, 6, 9, 12 for WX population). Lanes 1–3 were amplified with primers wF and wR, Lanes 4–6 were amplified with primers wAF and wR, Lanes 7–9 were amplified with primers wBpipF and wR, Lanes 10–12 were amplified with primers wBcauBF and wR. (B) PCR amplification of Wolbachia ank2 gene. Lane 1: TK population, Lane 2: NJ population, Lane 3: WX population.
(TIF)
Alignment of ank2 sequences from NJ, WX and TK populations. Alignment of amplified fragments of Wolbachia ank2 genes from NJ, WX and TK populations with two published sequences ank2-c (GenBank accession number AM397070.1) and ank2-d (GenBank accession number AM397071.1). NJ sequence shares 100% homology with ank2-c, WX and TK sequences are identical and share 100% homology with ank2-d.
(TIF)
Elimination of Wolbachia from TK, WX and NJ populations by tetracycline treatment. Eggs of tetracycline-treated and untreated mosquitoes were stained with Hoechst 33342 and visualized by fluorescence microscopy. (A) Wolbachia are concentrated at both anterior and posterior ends of untreated eggs indicated by strong fluorescence. (B) The same egg as in (A) under visible light. (C) Tetracycline-treated egg shows no Wolbachia distribution at either end of the egg. (D) The same egg as in (C) under visible light. Shown here are results of TK population. Treatment of WX and NJ populations had similar results. Scale bar, 0.1 mm.
(TIF)
Reciprocal crosses among Cx. pipiens pallens field populations.
(PDF)
Second round mating combinations of females retrieved from incompatible crosses.
(PDF)
Mating combinations of TK and WX females with both cognate and an increasing number of incompatible males.
(PDF)
Second round mating combinations of females retrieved from mixed male populations.
(PDF)
Mating combinations of F1 offspring of TK and WX crosses.
(PDF)
Crosses of tetracycline-treated and untreated WX and TK populations.
(PDF)