

Abstract
Stem cells must proliferate while maintaining ‘stemness’; however, much remains to be learned about how factors that control the division of stem cells influence their identity. Multiple stem cell types display cell cycles with short G1 phases, thought to minimize susceptibility to differentiation factors. Drosophila female germline stem cells (GSCs) have short G1 and long G2 phases, and diet-dependent systemic factors often modulate G2. We previously observed that Cyclin E (CycE), a known G1/S regulator, is atypically expressed in GSCs during G2/M; however, it remained unclear whether CycE has cell cycle-independent roles in GSCs or whether it acts exclusively by modulating the cell cycle. In this study, we detected CycE activity during G2/M, reflecting its altered expression pattern, and showed that CycE and its canonical partner, Cyclin-dependent kinase 2 (Cdk2), are required not only for GSC proliferation, but also for GSC maintenance. In genetic mosaics, CycE- and Cdk2-deficient GSCs are rapidly lost from the niche, remain arrested in a G1-like state, and undergo excessive growth and incomplete differentiation. However, we found that CycE controls GSC maintenance independently of its role in the cell cycle; GSCs harboring specific hypomorphic CycE mutations are not efficiently maintained despite normal proliferation rates. Finally, CycE-deficient GSCs have an impaired response to niche bone morphogenetic protein signals that are required for GSC self-renewal, suggesting that CycE modulates niche-GSC communication. Taken together, these results show unequivocally that the roles of CycE/Cdk2 in GSC division cycle regulation and GSC maintenance are separable, and thus potentially involve distinct sets of phosphorylation targets.
Keywords: BMP signaling, Cdk2, Cyclin E, Drosophila, Cell cycle, Germline stem cell
INTRODUCTION
Stem cells have extensive proliferative potential, but must also maintain a balance between self-renewal and production of differentiated daughters. Many progenitor/stem cell populations, including Drosophila germline stem cells (GSCs), C. elegans germline progenitors and mammalian embryonic stem cells, have cell cycles in which the G1 phase is very short or absent (Fox et al., 2011; Hsu et al., 2008; Singh and Dalton, 2009). Decreasing G1 length has been proposed as a strategy employed by various types of mammalian embryonic and adult stem cells to limit their sensitivity to differentiation signals (Lange and Calegari, 2010; Orford and Scadden, 2008; Singh and Dalton, 2009). Other lines of evidence in Drosophila neuroblasts and follicle stem cells (FSCs), and C. elegans germline progenitors, however, suggest that the canonical cell cycle regulator Cyclin E (CycE) can function to maintain stem cells independently of the cell cycle (Berger et al., 2010; Fox et al., 2011; Jeong et al., 2011; Singh and Dalton, 2009; Wang and Kalderon, 2009). The relationship between cell cycle regulation and stem cell maintenance across different systems is therefore incompletely defined, and the range of mechanisms involved remains poorly understood.
GSCs in the adult Drosophila ovary have relatively short G1 and long G2 phases, and multiple diet-dependent signals regulate G2 (Ables and Drummond-Barbosa, 2010; Hsu et al., 2008; LaFever et al., 2010). GSCs self-renew and generate cystoblasts via asymmetric cell division (Fig. 1A). Cystoblasts undergo four rounds of incomplete mitosis to produce 16-cell germline cysts (composed of one oocyte and 15 nurse cells) that are subsequently enveloped by follicle cells derived from FSCs (Ables et al., 2012). Although core cell cycle machinery components, including Cyclin A (CycA) and Cyclin B (CycB) in females and Cdc25 in males, influence GSC maintenance (Chen et al., 2009; Inaba et al., 2011; Wang and Lin, 2005), it is largely unknown how factors that control proliferation of GSCs modulate their self-renewal.
Fig. 1.

CycE expression peaks during G2, leading to an atypical CycE activity pattern in GSCs. (A) Drosophila germarium. GSCs in a niche composed of terminal filament (grey), cap cells and a subset of escort cells (yellow) give rise to cystoblasts that form 16-cell cysts. (A′) Distribution of GSCs displaying specific fusome morphologies relative to cell cycle phases (supplementary material Fig. S1, Table S1). (B-G) Expression of CycE (B-D) and Dup (E-G) in GSCs. (B,C,E,F) CycE (red; B,C) or Dup (green; E,F) expression in GSCs (outlined) with ‘round’ (B,E) or ‘fusing’ (C,F) fusomes; insets show GSC fusomes, visible in adjacent optical slices. (D,G) Average CycE (D) or Dup (G) fluorescence intensity in GSCs according to fusome morphology. Bars represent s.e.m. *P<0.05, compared with ‘fusing’; **P<0.05, compared with ‘bar’, ‘dumbbell’, ‘fusing’ and ‘exclamation point’; ***P<0.05, compared with all other fusome morphologies. (H) Maximum intensity projection (6 μm thickness) showing MPM2 (green) labeling the histone locus body in GSCs (white outlines) and nascent cystoblasts (yellow outlines). In mitotically dividing cysts (e.g. eight-cell cystocyte indicated by an arrowhead), MPM2 expression oscillates, as described previously (Narbonne-Reveau and Lilly, 2009). (I) MPM2 (white) labeling the histone locus body (arrows) in wild-type germline cells (marked by GFP; green), but absent from a GFP-negative CycE-null GSC (circled). (B,C,E,F,I) 1B1, fusomes (blue); LamC, cap cell nuclear envelopes (blue). LamC is also prominent in CycE mutant germ cells. Scale bars: 5 μm in all main panels or 2.5 μm in insets in C,F. (J) Quantification of MPM2-positive wild-type versus CycE-deficient GSCs. Numbers in bars represent the number of GSCs analyzed.
CycE, a known regulator of the G1/S transition in somatic cells (Möröy and Geisen, 2004), is atypically expressed in Drosophila female GSCs (Hsu et al., 2008). In ovarian follicle cells and germline cysts, CycE levels oscillate, peaking in G1 and rapidly decreasing during S (Calvi et al., 1998; Hsu et al., 2008; Lilly and Spradling, 1996). By contrast, CycE expression in GSCs is not limited to G1, as CycE is frequently detected with CycB (a G2/M marker) or during M phase (Hsu et al., 2008). It remained unclear, however, whether CycE has specialized cell cycle-independent roles in GSCs or whether it acts exclusively by modulating the cell cycle.
Here, we demonstrate that CycE controls the maintenance of GSCs by modulating their response to niche signals. CycE activity is broadly evident during G2 and M, reflecting its expression pattern. In addition to their role in GSC proliferation, CycE and its canonically associated kinase, Cyclin-dependent kinase 2 (Cdk2; also known as Cdc2c), are required for GSC maintenance. GSCs lacking CycE or Cdk2 function are rapidly lost from the niche, and become unusually large while arrested in a G1-like state. Although CycA and CycB levels are decreased in CycE mutant GSCs, our data suggest that the loss of CycE-deficient GSCs is not simply a consequence of alterations in other cyclins, or of lengthening of G1. Instead, CycE controls GSC maintenance at least in part independently of its role in the cell cycle. Specifically, GSCs harboring two hypomorphic CycE mutations (Wang and Kalderon, 2009) display normal rates of proliferation, but fail to be efficiently maintained. Finally, we show that CycE-deficient GSCs have an impaired response to niche bone morphogenetic protein (BMP) signals, which are known to be required for GSC maintenance (Chen et al., 2011; Xie and Spradling, 1998). We speculate that the cell cycle-independent function of CycE in modulating the responsiveness of GSCs to key signals could potentially be conserved in other stem cells or cancers.
MATERIALS AND METHODS
Drosophila strains and culture conditions
Flies were maintained at 22°-25°C in standard medium. For genetic mosaic analyses using flipase (FLP)/FLP recognition target (FRT) (Xu and Rubin, 1993), we obtained the following alleles on FRT-containing chromosomes: CycEAR95, CycEWX, CycE1F36 and CycEJP (Wang and Kalderon, 2009); CycEKG00239 (Drosophila Genetic Resource Center); and Cdk23 (Sukhanova and Du, 2008). CycB2 (Jacobs et al., 1998) was recombined with FRT40A. y w is used as wild type. Other genetic tools are described in FlyBase (Ashburner and Drysdale, 1994).
Genetic mosaic generation and GSC analyses
Genetic mosaics were generated by FLP/FRT-mediated recombination in 2- to 3-day-old females carrying a mutant allele in trans to a wild-type allele (linked to a Ubi-GFP marker) on homologous FRT arms, and a hs-FLP transgene, as described (Ables and Drummond-Barbosa, 2010). Wild-type, instead of mutant, alleles were used for control mosaics. GSCs were identified based on the juxtaposition of their fusomes to adjacent cap cells (Ables and Drummond-Barbosa, 2010; Hsu and Drummond-Barbosa, 2009).
GSC loss was measured as the percentage of total germline-mosaic germaria showing evidence of recent GSC loss, namely the presence of GFP-negative cystoblasts/cysts in the absence of the GFP-negative mother GSC, 8 or 12 days after recombination (Method I) (Ables and Drummond-Barbosa, 2010; Hsu and Drummond-Barbosa, 2009). Alternatively, we quantified the frequency of total analyzed germaria containing at least one GFP-negative GSC 4, 8 or 12 days after recombination (Method II) (Xie and Spradling, 1998). Results were subjected to χ2 analysis.
To quantify proliferation, dissected ovaries were incubated for 1 hour at room temperature in Grace's Insect Media (Lonza) containing 10 μm 5-ethynyl-2′-deoxyuridine (EdU; Invitrogen), immediately preceding fixation and staining. EdU was detected using AlexaFluor-594 via Click-It chemistry, following the manufacturer's recommendations (Invitrogen). The number of EdU-positive GSCs was measured as a percentage of total GFP-negative GSCs analyzed. Results were subjected to χ2 analysis. The number of GFP-negative GSC progeny was calculated as the total number of GFP-negative cystoblasts and cysts divided by the total number of GFP-negative GSCs in each germarium. Results were averaged across 24 to 33 germaria.
Immunofluorescence and microscopy
Ovaries were dissected, fixed, washed and blocked as described previously (Ables and Drummond-Barbosa, 2010), except where noted. The following primary antibodies were used overnight at 4°C: mouse anti-Hts [1B1, Developmental Studies Hybridoma Bank (DSHB); 1:10], mouse anti-Lamin C (LamC) (LC28.26, DSHB; 1:100), rabbit anti-GFP (TP-401, Torrey Pines Biolabs; 1:2500), rabbit anti-pMad (#1880, Epitomics; 1:100), mouse anti-CycB (F2F4, DSHB; 1:20), mouse anti-Bam (DSHB; 1:10), rat anti-E-Cadherin (DCAD2, DSHB; 1:3), mouse anti-Orb (4H8, DSHB; 1:20), rabbit anti-cleaved Caspase 3 (#9661S, Cell Signaling Technology; 1:50), guinea pig anti-CycE (a gift from T. Orr-Weaver, Whitehead Institute, Cambridge, MA, USA; 1:1000) (Hsu et al., 2008), mouse anti-CycE (8B10; 1:10) (Richardson et al., 1995), mouse anti-MPM2 (#05-368, Millipore; 1:100), guinea pig anti-Dup (1:1000) (Whittaker et al., 2000) and rabbit anti-C(3)G (1:3000) (Hong et al., 2003). For anti-E-Cadherin labeling, fixed ovaries were permeabilized for 30 minutes in 0.5% Triton X-100 in PBS prior to blocking. For anti-Bam labeling, ovaries were fixed for 10 minutes in 4% formaldehyde, permeabilized in 0.3% Triton X-100 in PBS, then blocked in 0.5% BSA in 0.3% Triton X-100 in PBS prior to primary antibody incubation. Following a 2-hour incubation with Alexa Fluor 488-, 568- or 633-conjugated goat species-specific secondary antibodies (Invitrogen; 1:200), ovaries were stained with 0.5 μg/ml 4′-6-diamidino-2-phenylindole (DAPI) (Sigma). Ovaries were mounted in 90% glycerol containing 20 mg/ml n-propyl gallate (Sigma). Confocal z-stacks (1 μm optical sections) were collected with a Zeiss LSM 700 microscope using ZEN 2009 software. Images were analyzed using Zeiss ZEN or AxioVision software, and minimally and equally enhanced via histogram using Adobe Photoshop CS4. DNA content was measured as the total DAPI fluorescence intensity across 3-7 single z-planes in AxioVision, using ‘round’ fusome wild-type GSCs in G2 or early mitosis (4n) as controls. To compare pMad levels between GFP-negative mutant GSCs and neighboring GFP-positive wild-type GSCs, fluorescence intensity was quantified using AxioVision by manually demarcating individual GSC nuclei and measuring densitometric mean (grey units/pixel) at the largest nuclear diameter. E-cadherin levels at the GSC-cap cell junction were similarly measured using morphometric analysis of single z-planes or maximum intensity projections. As a measure of cell size, we calculated the cross-sectional area of GFP-negative single cells across their largest diameter in control versus mutant genetic mosaics using AxioVision. Statistical analysis was performed using the Student's two-tailed t-test or one-way ANOVA in Microsoft Excel or GraphPad Prism.
RESULTS
Cyclin E expression peaks during G2 in GSCs
CycE, a canonical G1/S cyclin (Möröy and Geisen, 2004), is atypically expressed during the G2 and M phases in GSCs, based on colocalization with CycB and mitotic figures (Hsu et al., 2008). To examine CycE levels carefully as GSCs progress through the cell cycle, we quantified CycE fluorescence intensity within wild-type GSCs displaying different forms of the fusome, a germline-specific organelle that undergoes morphological changes according to the cell cycle (Fig. 1A′; supplementary material Fig. S1, Table S1) (de Cuevas and Spradling, 1998; Hsu et al., 2008). Our analysis (n=467 GSCs) showed that CycE reaches its highest levels during G2, specifically when GSCs have ‘round’ fusomes (Fig. 1A′-D). CycE levels remain relatively high through the ‘plug’ fusome stage, which corresponds to GSCs in G1 (Fig. 1D). During subsequent cell cycle stages, exhibiting ‘bar’ (G1/S), ‘dumbbell’ (S), ‘fusing’ (very early G2) and ‘exclamation point’ (G2) fusome morphologies (Fig. 1A′), CycE expression becomes significantly lower (Fig. 1C,D). Thus, CycE expression peaks during G2 prior to the onset of M and is downregulated as cells progress from G1 into S, remaining low into early G2.
Cyclin E appears to be active during G2 in GSCs
The high levels of CycE during G2 in GSCs might represent CycE actively complexed with Cdk2; alternatively, CycE might remain inactive until G1, following a more canonical pattern of activity. Our analysis of the DNA replication licensing factor Double parked (Dup), the mammalian Cdt1 homolog (Whittaker et al., 2000), supports the former possibility. During eye development, Dup is co-expressed with CycE in late G1 but is then rapidly degraded (in part as a result of CycE/Cdk2 activity), while CycE expression extends into S (Thomer et al., 2004). In GSCs, Dup also mirrors CycE in its pattern of high G2 expression; however, in contrast to CycE, Dup expression is negligible in G1 (Fig. 1E-G). Thus, Dup and CycE are co-expressed in GSCs (albeit in an atypical pattern) and the decline in Dup expression precedes that of CycE, reminiscent of their relationship in the developing eye.
To more directly monitor CycE activity in GSCs, we used MPM2 antibodies that recognize conserved mitotic phospho-epitopes (Davis et al., 1983) and label the Drosophila histone locus body, reflecting CycE activity in the embryo, endocycling follicle cells and mitotically dividing cysts (Calvi et al., 1998; Narbonne-Reveau and Lilly, 2009; White et al., 2007). We observed MPM2-positive spherical structures corresponding to the histone locus body in early germ cells (Fig. 1H,I), and confirmed that MPM2 labeling is dependent on CycE in GSCs (Fig. 1I,J). In contrast to the oscillating presence of MPM2 epitopes in dividing germline cysts (Narbonne-Reveau and Lilly, 2009), MPM2 stains all GSCs, regardless of their cell cycle phase (n=110 GSCs; Fig. 1H,J), suggesting that CycE is active throughout most of the cell cycle. The constant phosphorylation of the histone locus body detected by MPM2 in GSCs (in contrast to the dynamic CycE pattern) potentially reflects more stable phospho-epitopes in the histone locus body relative to other targets or, alternatively, a lower CycE activity threshold requirement for the phosphorylation of a subset of targets. Taken together, these results suggest that some of the cell cycle events (i.e. high CycE activity, Dup expression) normally associated with G1 occur during G2 in GSCs. These observations could potentially help explain the short G1 in GSCs.
CycE- and Cdk2-deficient GSCs arrest in a G1-like state
Given the atypical GSC patterns of CycE expression and activity, we examined the roles of CycE in GSCs. We first asked whether CycE is required for GSC proliferation. We analyzed genetic mosaic females carrying CycE-deficient GSCs (recognized by the absence of green fluorescent protein, GFP), using EdU as an S-phase marker (Fig. 2). Not surprisingly, unlike wild-type GSCs in control ‘mock’ mosaics (Fig. 2A,E), null CycEAR95 GSCs (Fig. 2B,E) failed to incorporate EdU. CycEAR95 mosaics did not contain GFP-negative multicellular germline cysts (Fig. 2B; data not shown), indicating that CycE is also required for division of GSC daughters. By contrast, hypomorphic CycE1F36 (Park et al., 2007) or CycEWX (Wang and Kalderon, 2009) GSCs incorporated EdU at control frequencies (Fig. 2C,E) and produced equivalent number of progeny to control GSCs (control: 3.9±1.2 progeny per GSC; CycE1F36: 4.0±1.2 progeny per GSC; CycEWX: 3.8±1.6 progeny per GSC). Furthermore, formation of GFP-negative cysts and overall morphology of CycE1F36 and CycEWX mosaic germaria were comparable with controls (Fig. 2C; also see supplementary material Fig. S6C,D). Thus, CycE function is required for proliferation of GSCs and their progeny, but CycE1F36 and CycEWX GSCs retain sufficient activity to support normal proliferation rates.
Fig. 2.
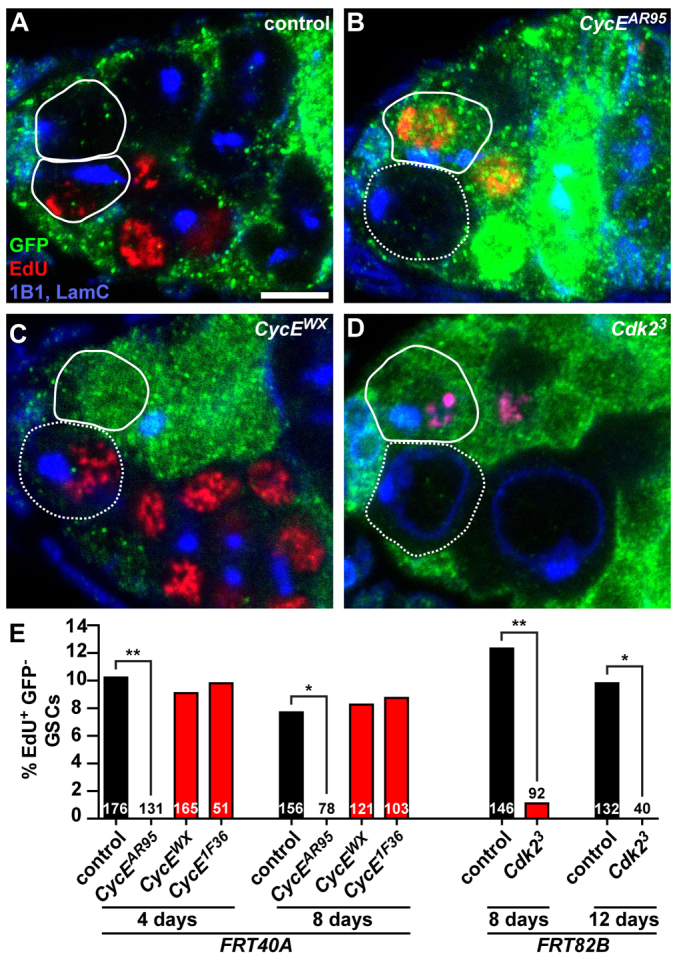
CycE and Cdk2 are required for GSC proliferation. (A-D) Mosaic germaria showing control (A), null CycEAR95 (B), hypomorphic CycEWX (C) and null Cdk23 (D) GFP-negative clones. GFP (green), wild-type control cells; EdU (red), cells in S phase; 1B1 (blue), fusomes; LamC (blue), cap cell nuclear envelopes. Scale bar: 5 μm. (E) Percentage of GFP-negative GSCs that are positive for EdU in mosaic germaria at 4 or 8 days after clone induction. Numbers in the bars represent the number of GFP-negative GSCs analyzed. *P≤0.01; **P≤0.001.
CycE functions in a complex with a cyclin-dependent kinase, Cdk2, to regulate the cell cycle (Möröy and Geisen, 2004); therefore, we asked whether Cdk2 was also required for GSC proliferation. Null Cdk23 GSCs in mosaic germaria showed a drastically reduced frequency of EdU incorporation relative to control GSCs at 8 days after clone induction (1% versus 12%, respectively; Fig. 2D,E). The residual EdU incorporation in Cdk23 GSCs presumably results from wild-type Cdk2 perdurance, as we find no EdU-positive Cdk23 GSCs at 12 days (Fig. 2E). Null Cdk23 mosaics also failed to form normal GFP-negative cysts (Fig. 2D), phenocopying CycEAR95 mosaics (Fig. 2B). These results are consistent with a requirement for CycE/Cdk2 activity during GSC proliferation.
Although CycE/Cdk2 canonically regulates the G1/S transition (Möröy and Geisen, 2004), the atypical GSC expression of CycE in G2 and M prompted us to determine the stage of cell cycle arrest in null CycEAR95 GSCs. CycEAR95 GSCs had approximately half the DAPI fluorescence [95,948±21,708 (s.d.) grey units; n=11] of control 4n germline cells in G2 or early mitosis [197,795±43,576 (s.d.) grey units; n=18], indicating that CycEAR95 GSCs have a 2n DNA content. We also examined the mitotic cyclins CycA and CycB, which are expressed during S/G2 and G2/M, respectively (Hsu et al., 2008; Lilly et al., 2000; Wang and Lin, 2005). As previously described (Lilly et al., 2000), CycA expression was detected in the cytoplasm and enriched on ‘round’ fusomes of wild-type GFP-positive GSCs (supplementary material Fig. S2). By contrast, CycEAR95 GSCs did not express CycA (n=22 GSCs). Similarly, Although CycB was detected in neighboring GFP-positive control GSCs with the ‘round’ fusome morphology typical of cells in G2 and M (Hsu et al., 2008), CycB was absent from all CycE-deficient GSCs examined (Fig. 3A,A′; n=60 CycEAR95 GSCs). A large percentage of CycEAR95 GSCs have either a ‘plug’-like (Fig. 3C; 20%) or abnormally stretched (Fig. 3D; 24%) fusome (see Fig. 1A), whereas 53% of CycEAR95 GSCs (n=86) have a ‘round’-like fusome (Fig. 3B,C). These results indicate that fusome morphology becomes aberrant in the absence of CycE, consistent with the reported detection of active CycE at the fusome (Ohlmeyer and Schüpbach, 2003). Finally, we observe that null CycEAR95 or Cdk23 GSCs appear to grow continuously, reaching significantly larger sizes than wild-type GSCs (supplementary material Fig. S3). Taken together, these data indicate that GSCs completely lacking CycE activity arrest in an abnormal G1-like state.
Fig. 3.
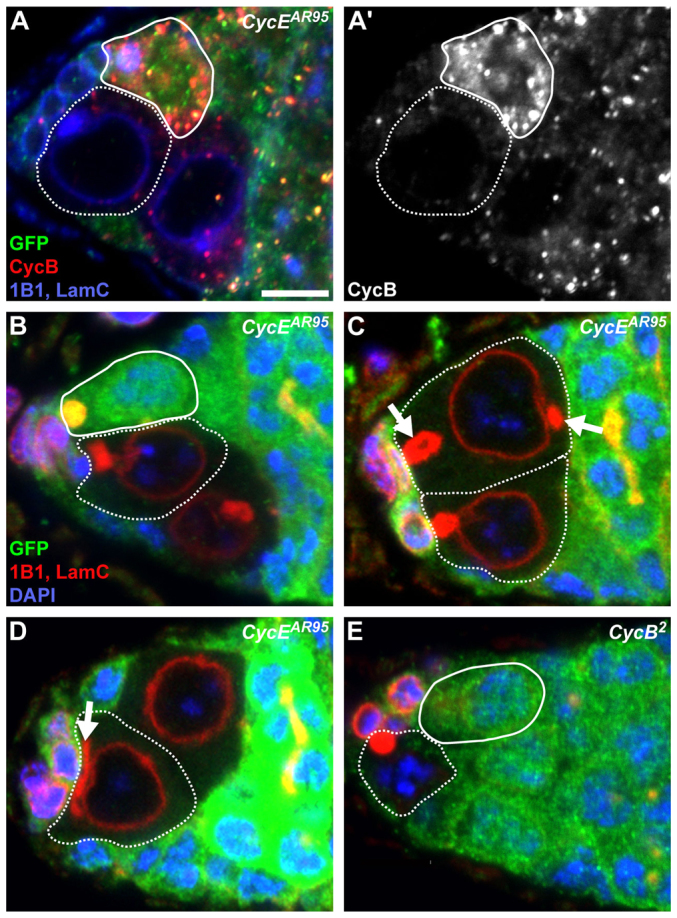
GSCs lacking CycE arrest in a G1-like state showing a phenotype distinct from that of CycB-deficient GSCs. (A,A′) Mosaic germarium showing a GFP-negative CycEAR95 clone. CycB (red) is shown in greyscale in A′. GFP (green), wild-type control cells; 1B1 (blue), fusomes; LamC (blue), cap cell nuclear envelopes. (B-E) CycEAR95 (B-D) or CycB2 (E) mosaic germaria showing GFP-negative clones. Arrows indicate unusual fusome morphology (see supplementary material Fig. S1 for comparison). GFP (green), wild-type control cells; 1B1 (red), fusomes; LamC (red), cap cell nuclear envelopes; DAPI (blue), nuclei. Solid outline indicates wild-type GSCs; dashed outlines indicate mutant GSCs. Scale bar: 5 μm.
CycE and Cdk2 are required for GSC maintenance
The unusual expression and activity profiles of CycE in GSCs suggest that CycE might have specialized roles in these cells. To test whether CycE is required for GSC maintenance, we measured GSC loss in control versus CycE mutant mosaic germaria (Fig. 4; see Materials and methods). We calculated the percentage of germaria with a mosaic germline that contained GFP-negative cystoblasts/cysts, but lacked an accompanying mother GFP-negative GSC (i.e. GSC loss event). In control ‘mock’ mosaics, where all cells are wild type, few GSC loss events were observed (Fig. 4A,E; supplementary material Table S2). In striking contrast, null CycEAR95 or strong hypomorphic CycEKG00239 mosaic germaria frequently displayed large, single, GFP-negative cells, without an accompanying GFP-negative GSC (Fig. 4B,E; supplementary material Table S2). Germaria mosaic for the hypomorphic CycEWX or CycE1F36 alleles showed GSC loss at a small but significant frequency (Fig. 4C,E; supplementary material Table S2). This finding is in contrast to a previous report that CycEWX GSCs are well maintained (Wang and Kalderon, 2009); this discrepancy is likely to be due to the different sensitivities of methods used to measure GSC loss (for example, compare Method I and Method II in supplementary material Table S2). We also found frequent GSC loss in null Cdk23 mosaic germaria (Fig. 4D,E; supplementary material Table S2), comparable with that of null CycEAR95 mosaics. Similar results were obtained by measuring the percentage of total analyzed germaria carrying a GFP-negative GSC over time (supplementary material Table S2). These data indicate that CycE/Cdk2 is required for maintenance of the GSC fate.
Fig. 4.

CycE and Cdk2 are required for GSC maintenance; this role is at least in part genetically separable from cell cycle regulation. (A-D) Mosaic germaria showing control (A), null CycEAR95 (B), hypomorphic CycEWX (C) and null Cdk23 (D) GFP-negative clones. GFP (green), wild-type control cells; 1B1 (red), fusomes; LamC (red), cap cell nuclear envelopes; DAPI (blue), nuclei. GSCs are outlined. Arrows indicate GSC loss events. Scale bar: 5 μm. (E) Percentage of germline-mosaic germaria with a GSC loss event 8 or 12 days after clone induction. Numbers in the bars represent the number of germline-mosaic germaria analyzed. *P<0.0001.
CycE mutant GSCs have a distinct phenotype relative to GSCs lacking CycB
CycB and CycA are known to modulate GSC maintenance (Chen et al., 2009; Hsu et al., 2008; Wang and Lin, 2005); therefore, we considered the possibility that CycE-deficient GSCs are lost due to the absence of these mitotic cyclins (see Fig. 3A,A′; supplementary material Fig. S2). Although we cannot formally rule out that absence of CycA contributes to the maintenance defect of CycEAR95 GSCs, an excess, rather than lack, of CycA has been shown to impair GSC maintenance (Chen et al., 2009). As previously reported (Wang and Lin, 2005), null CycB2 GSCs are rapidly lost from the niche (supplementary material Fig. S4). The phenotype of CycEAR95 GSCs, however, is markedly distinct from that resulting from loss of CycB function (Fig. 3). CycB2 GSCs are normally sized (Fig. 3E; supplementary material Fig. S3), display a compact DNA morphology (in contrast to the more diffuse DNA morphology of CycEAR95 GSCs; Fig. 3), and are found with either ‘round’ or ‘fusing’ fusome morphologies, consistent with a G2 arrest (Fig. 3E; n=32 GSCs). Thus, it seems improbable that CycEAR95 GSCs fail to be maintained simply as a secondary consequence of CycB absence.
CycE has a role in GSC maintenance that is at least partially distinct from its role in proliferation
Null CycE GSCs have impaired proliferation and maintenance; however, it remains unclear whether the maintenance role of CycE is secondary to its effects on cell cycle progression. Lower proliferation rates per se do not cause GSC loss because GSCs defective for insulin signaling in genetic mosaics divide more slowly due to a prolonged G2, but are well maintained (Hsu and Drummond-Barbosa, 2009; Hsu et al., 2008; LaFever et al., 2010). It remained possible, however, that the G1-like arrest of GSCs deficient for CycE (see Fig. 3) was responsible for their loss. Nevertheless, hypomorphic CycEWX and CycE1F36 GSCs in mosaic germaria are lost at a significantly higher rate than control GSCs (Fig. 4C,E), despite their normal proliferation rates (see Fig. 2C,E), demonstrating that the role of CycE in GSC maintenance is separable from its role in regulating GSC proliferation. Additionally, CycEWX GSCs do not grow excessively (see supplementary material Fig. S3), indicating that the loss of CycEAR95 GSCs is not simply a consequence of their large size. Thus, we conclude that CycE has a cell cycle-independent role in controlling GSC maintenance. Given that the GSC loss phenotype is less severe for CycEWX and CycE1F36 than for stronger CycEAR95 or CycEKG00239 alleles, however, we cannot exclude the possibility of a partial contribution of CycE to GSC maintenance that is secondary to its cell cycle role.
Absence of CycE does not lead to apoptosis
We next asked whether loss of CycE or cell cycle disruption in null CycEAR95 GSCs could result in their apoptosis. We analyzed CycEAR95 mosaic germaria using the early apoptosis marker cleaved Caspase 3 (Yu et al., 2002), but did not detect it in any GFP-negative CycEAR95 germ cell (Fig. 5A-C; n=28 CycEAR95 GSCs). Although other forms of cell death are theoretically possible, they are unlikely, because the frequency of observed GSC loss events increases with time (see Fig. 4E), consistent with inefficient clearing of CycE-deficient cells from mosaic germaria. Indeed, CycEAR95 germ cells are frequently detected near early follicle cells (Fig. 5B), and are occasionally encapsulated by follicle cells (Fig. 5D). Intriguingly, however, we found that the percentage of CycEAR95 mosaic germaria containing at least one GFP-positive cell with cleaved Caspase 3 was significantly increased relative to control ‘mock’ mosaics (Fig. 5B,B′,C). We also noticed many examples of CycEAR95 mosaic germaria containing GFP-negative, yet no GFP-positive, germ cells (Fig. 5A). Taken together, these results indicate that CycE-deficient cells are not lost through apoptosis. Furthermore, they suggest that CycE null cells, perhaps owing to their excessive growth, outcompete neighboring wild-type cells for nutrients, survival factors, or other resources, resulting in the death of otherwise normal wild-type cells.
Fig. 5.
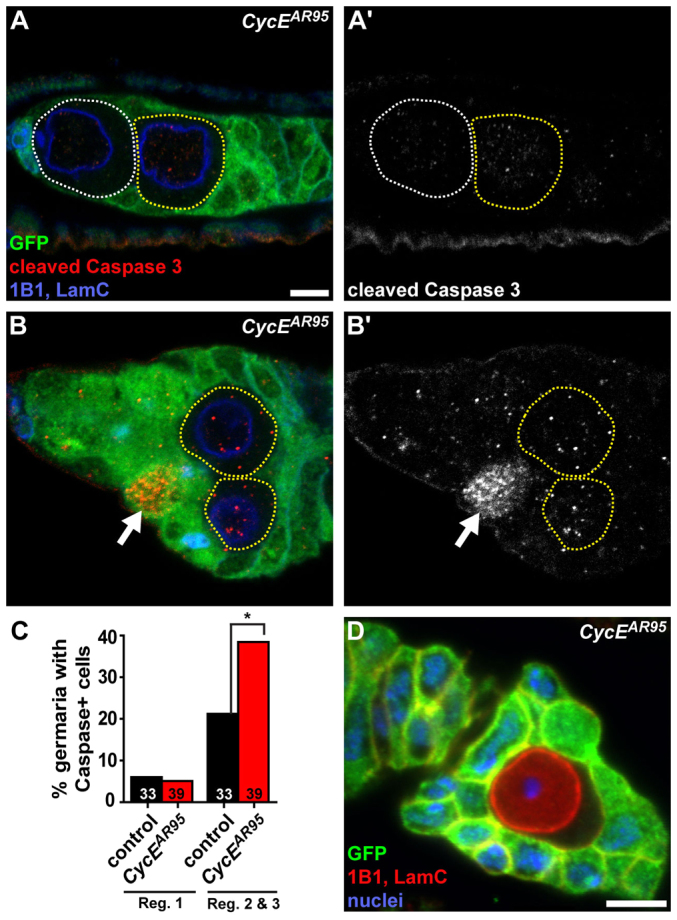
CycE mutant GSCs are not lost through apoptosis. (A-B′) Mosaic germaria showing GFP-negative CycEAR95 clones. GFP (green), wild-type control cells; cleaved Caspase 3 (red) (greyscale images in A′,B′), early apoptosis marker; 1B1 (blue), fusomes; LamC (blue), cap cell nuclear envelopes. CycEAR95 GSCs are outlined in white; CycEAR95 cystoblast-like cells are outlined in yellow. Arrows indicate a GFP-positive wild-type germ cell positive for cleaved Caspase 3. (C) Percentage of germline-mosaic germaria with at least one Caspase-positive cell in region 1 (which contains GSCs and cystoblasts) or regions 2 and 3 (which normally contain more differentiated cysts) in ‘mock’ control (black) or CycEAR95 (red) mosaic germaria. Numbers in bars represent number of germline-mosaic germaria analyzed. *P<0.01. (D) Single GFP-negative CycEAR95 germ cell enveloped by follicle cells in mosaic ovariole. GFP (green), wild-type control cells; 1B1 (red), follicle cell membranes. LamC (red) stains germ cell nuclear envelope; DAPI (blue), nuclei. Scale bars: 5 μm.
CycE does not control E-cadherin levels at the GSC-cap cell junction
Our observations that single CycEAR95 germ cells do not apoptose and are detected away from the niche led us to hypothesize that the absence of CycE in GSCs may cause their loss as a result of downregulation of E-cadherin, which is required for anchoring GSCs to the niche (Song et al., 2002). We found no significant difference, however, in E-cadherin mean fluorescence intensity at the cap cell-GSC interface between CycEAR95 and adjacent wild-type GSCs (Fig. 6A-C), suggesting that the loss of CycEAR95 GSCs is not due to reduced E-cadherin levels. In fact, the total E-cadherin fluorescence intensity at the cap cell-GSC interface in CycEAR95 GSCs is significantly greater than that of control GSCs (Fig. 6D), consistent with the large size and apparent larger niche contact area of CycEAR95 GSCs (e.g. see Fig. 6B; supplementary material Fig. S3). These results are in contrast to the proposed connection between CycE and E-cadherin in FSCs, although in those studies, E-cadherin levels were not directly measured in CycE mutant FSCs (Wang and Kalderon, 2009).
Fig. 6.
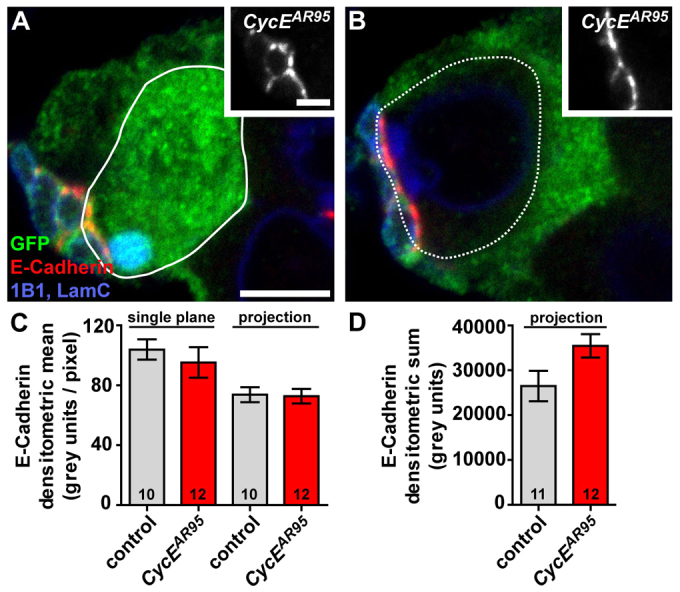
CycE AR95 GSCs have normal levels of E-cadherin at the GSC-cap cell interface. (A,B) Anterior region of a mosaic germarium showing a GFP-positive wild-type GSC (A) and a GFP-negative CycEAR95 GSC (B). GFP (green), wild-type control cells; E-cadherin (red; greyscale images shown in insets); 1B1 (blue), fusomes; LamC (blue) cap cell nuclear envelopes. GSCs are outlined. Scale bars: 5 μm (or 2.5 μm in insets). (C,D) E-cadherin levels at the GSC-cap cell junction, measured by densitometric mean (C) or sum (D) of pixel intensity in single optical slices or maximum intensity projections in CycEAR95 or adjacent wild-type GSCs. Numbers in the bars represent number of GSCs measured. Bars represent s.e.m.
Null CycEAR95 GSCs have a reduced response to BMP signals, but fail to fully differentiate
CycE mutant GSCs are not lost due to apoptosis or reduced E-cadherin levels; therefore, we asked whether premature differentiation might explain the rapid loss of CycEAR95 GSCs. BMP ligands produced by the niche are required for GSC maintenance (Chen et al., 2011; Xie and Spradling, 1998). To test the ability of CycE mutant GSCs to respond to these signals, we measured the levels of phosphorylated Mothers against decapentaplegic (pMad), a well-established reporter of BMP pathway activation (Ables and Drummond-Barbosa, 2010; Kai and Spradling, 2003). CycEAR95 GSCs consistently displayed lower levels of pMad than neighboring wild-type GSCs (Fig. 7A,A′,B). Wild-type cystoblasts also express low levels of pMad once they transition from pre-cystoblast to cystoblast (Rangan et al., 2011). Nevertheless, CycE AR95 cystoblast-like cells express lower levels of pMad than neighboring wild-type cystoblasts of similar proximity to the niche (Fig. 7A,A′,B). These data indicate that, in the absence of CycE function, GSCs do not respond properly to niche BMP signals.
Fig. 7.

CycE AR95 GSCs have reduced BMP signaling, but do not fully differentiate. (A,A′) CycEAR95 mosaic germaria showing GFP-negative clones. pMad (red; greyscale image in A′) is a BMP signaling marker. Arrowheads indicate a CycEAR95 cystoblast-like cell. (B) Average level of pMad expression in control or CycEAR95 GSCs in mosaic germaria. Numbers in the bars represent the number of GSCs analyzed. Bars represent s.e.m. **P<0.01. (C-D′) CycEAR95 mosaic germaria. Bam (red; greyscale image in C′,D′) is an early differentiation marker. Arrows indicate GFP-positive wild-type germline cysts that express high levels of Bam. In A,C,D, GFP (green) indicates control cells, 1B1 (blue) indicates fusomes, and LamC (blue) indicates cap cell nuclear envelopes. Control wild-type GSCs have solid white outlines; CycEAR95 GSCs are indicated by dashed white outlines; control cystoblasts are indicated by solid yellow outlines; CycEAR95 cystoblast-like cells are indicated by dashed yellow outlines. (E) Model for genetically separable roles of CycE/Cdk2 in regulating the female GSC cell cycle and maintenance. CycE function promotes GSC responsiveness to niche BMP signals; in addition, high CycE expression during G2 and M probably results in rapid entry into S following M phase, which limitis excessive growth during G1. An indirect contribution of CycE/Cdk2 to GSC function through its effect on the cell cycle is also possible. It remains unknown what mechanisms lead to the atypical CycE pattern in GSCs. Scale bars: 5 μm.
The reduced BMP signaling in CycEAR95 GSCs could be an indirect consequence of the G1-like cell cycle stage in which they are arrested (see Fig. 3), or they may reflect a more direct role of CycE in modulating BMP responsiveness. We therefore measured pMad levels throughout the cell cycle of wild-type GSCs and found no cell cycle-dependent pattern (supplementary material Fig. S5). These observations suggest that CycE modulates BMP signaling independently of the cell cycle, in accordance with the impaired maintenance of CycEWX and CycE1F36 GSCs in the absence of proliferation defects (see Figs 2, 4).
BMP signaling inhibits GSC differentiation at least in part through transcriptional repression of bag of marbles (bam) (Chen et al., 2011). Low pMad levels in CycEAR95 GSCs might therefore indicate that these cells have prematurely differentiated to a cystoblast or later fate, despite failing to divide again because of the separate role of CycE in proliferation. As in wild-type GSCs, however, Bam is undetectable in CycEAR95 GSCs, despite low pMad levels (Fig. 7C,C′), reminiscent of the situation in GSCs mutant for pelota, a gene that controls self-renewal through repression of a Bam-independent differentiation pathway (Xi et al., 2005). Thus, although the reduction in BMP signaling observed in CycEAR95 GSCs is not sufficient to cause Bam upregulation, it might derepress a Bam-independent differentiation pathway.
We also examined the fate of CycEAR95 cystoblast-like cells. These cells express Bam at the moderate levels found in wild-type cystoblasts (Fig. 7C,D), but never show the high levels of Bam observed in wild-type two-, four- or eight-cell cysts (Fig. 7C,D; n=48 CycEAR95 cystoblast-like cells), suggesting that CycEAR95 cystoblast-like cells only partially differentiate. Accordingly, CycEAR95 cystoblast-like cells, despite their large size and frequent proximity to follicle cells, do not express the oocyte-specific markers Orb (Lantz et al., 1994) or C(3)G (Hong et al., 2003) (supplementary material Fig. S6A,B; n=73 or 86 CycEAR95 cystoblast-like cells, respectively). We considered the possibility that CycEAR95 cystoblast-like cells fail to differentiate due to impaired function of the gap junction component Innexin 4/Zero population growth (Zpg); however, zpg-deficient cystoblasts die at very early stages of differentiation (Gilboa et al., 2003), unlike CycEAR95 cystoblast-like cells. Instead, the failure of CycEAR95 cystoblast-like cells to fully differentiate is probably a consequence of their inability to divide, as CycEWX and CycE1F36 germ cells appear to differentiate normally (supplementary material Fig. S6C,D). In summary, these results show that CycE controls the responsiveness of GSCs to BMP signals, probably in a cell cycle-independent way, and that the arrest of CycEAR95 GSCs is not sufficient to cause their terminal differentiation or the upregulation of differentiation factors. In fact, the role of CycE in promoting proliferation/S-phase entry appears to be required for the complete differentiation of germ cells.
DISCUSSION
In this study, we uncover cell cycle-dependent and -independent roles of a core cell cycle machinery component, CycE, in adult Drosophila female GSCs. We demonstrate that CycE and its canonical binding partner Cdk2 promote GSC identity, and unequivocally show that the role of CycE in GSC maintenance is, at least in part, genetically separable from regulation of the GSC division cycle. Importantly, we show that CycE controls the responsiveness of GSCs to niche-derived BMP signals. In addition, our results suggest that high CycE levels in G2/M are accompanied by an altered pattern of CycE activity that probably contributes to the short length of G1 in GSCs, limiting their excessive growth. We propose the model that CycE/Cdk2 regulates two distinct sets of targets, activated by different thresholds of CycE/Cdk2 activity to stimulate cell cycle progression and promote stem cell maintenance (Fig. 7E).
Cell cycle-independent roles of CycE in stem cell maintenance
Many correlative studies address the connection between the stem cell division cycle and ‘stemness’ itself (Lange and Calegari, 2010; Orford and Scadden, 2008; Singh and Dalton, 2009); however, the precise nature of this connection and the contribution of individual cell cycle regulators to stem cell self-renewal remain largely unknown. Specifically, the proposed idea that many stem cells employ a short G1 phase as a strategy to minimize their vulnerability to differentiation factors has been controversial. Knockdown of Cdk2 in human embryonic stem cells causes G1 arrest and differentiation (Neganova et al., 2009). Similarly, inhibition of Cdk2/CycE or other anti-proliferative manipulations lead to differentiation of neuronal precursors (Salomoni and Calegari, 2010). By contrast, prolonging G1 in mouse embryonic stem cells does not promote differentiation (Li et al., 2012), suggesting that the length of G1 is not a sufficient determinant of whether stem cells retain their identity or differentiate.
Several studies in Drosophila and C. elegans stem cells have also addressed the connection between the cell cycle and stem cell fate. Loss of CycB or excess of CycA causes increased female GSC loss (Chen et al., 2009; Wang and Lin, 2005), and Cdc25/string is required for male GSC maintenance (Inaba et al., 2011). CycE function is required for FSC maintenance, and analysis of CycEWX in the FSC lineage showed that FSC loss occurs in the absence of follicle cell proliferation defects (Wang and Kalderon, 2009). In this study, however, the cell cycle was not directly examined in FSCs (Wang and Kalderon, 2009), and it is known that the proliferation of FSCs and follicle cells can be distinctly regulated (LaFever et al., 2010). We demonstrate that the role of CycE in GSC maintenance is at least partially distinct from its function in the cell cycle of the GSC, as CycEWX and CycE1F36 GSCs fail to be maintained but exhibit normal proliferation. The CycEWX allele carries a C-terminal domain deletion that results in lower CycE/Cdk2 kinase activity (Wang and Kalderon, 2009), suggesting that GSC maintenance targets require a higher threshold of kinase activity than cell cycle regulatory targets. Although we did not exclude an additional contribution of the cell cycle to GSC maintenance, the separate roles of CycE in GSC maintenance versus proliferation are reminiscent of the situation in the developing Drosophila nervous system and in the C. elegans germline, where CycE controls precursor identity independently of its role in their cell cycle (Berger et al., 2010).
Our studies show that CycE promotes GSC maintenance at least in part by modulating how GSCs respond to niche BMP signals. Similarly, C. elegans CycE genetically interacts with Notch signaling from the niche (Fox et al., 2011). By contrast, CycE controls neuroblast maintenance through Prospero localization (Berger et al., 2010), indicating that CycE operates through diverse mechanisms in different stem cell populations. Moreover, although both Drosophila and C. elegans CycE interact with niche signals, in C. elegans, the absence of CycE results in loss of GSCs by promoting their entry into meiosis (Fox et al., 2011; Jeong et al., 2011). In Drosophila, however, the impairment in GSC maintenance appears to occur at a stage earlier than the meiotic entry decision.
The modulation of BMP signaling by CycE suggests that CycE/Cdk2 might phosphorylate components or regulators of the BMP signaling machinery. Alternatively, CycE/Cdk2 may modify chromatin modifiers, which have also been implicated in modulating BMP signaling in GSCs (Ables and Drummond-Barbosa, 2010; Xi and Xie, 2005). In fact, a genetic screen for dominant modifiers of the hypomorphic CycEJP allele during eye development identified components of the Brahma chromatin remodeling complex as CycEJP suppressors (Brumby et al., 2004). Future studies should identify the sets of CycE/Cdk2 targets required for maintenance versus cell cycle regulation of various stem cell populations.
Other cell cycle-independent roles of cell cycle regulators
As mentioned above, cell cycle regulators, including Cdc25, CycB and CycA have been implicated in GSC maintenance (Chen et al., 2009; Inaba et al., 2011; Wang and Lin, 2005), although it remains unclear whether they control stem cell fate independently of the cell cycle. Yet, cell cycle regulators perhaps function outside of the cell cycle more commonly than previously thought. For example, Drosophila E2F is required for transfer of cytoplasm from nurse cells to oocytes, and for the establishment of oocyte dorsal-ventral polarity (Myster et al., 2000). In postmitotic mammalian neurons, constitutively expressed CycE binds to and inhibits Cdk5 to control synaptic plasticity and memory formation (Odajima et al., 2011). Activation of Cdk5 in adipocytes as a result of high fat-induced obesity also leads to PPARγ phosphorylation and deregulated expression of insulin-sensitizing genes independently of cyclins (Choi et al., 2010). CycE also has a Cdk2-independent role in DNA damage-induced apoptosis of growth-arrested mesenchymal precursors (Ugland et al., 2008). It will be important to evaluate other cell cycle regulators beyond their traditional cell cycle roles in stem cells and more differentiated cell types.
Altered CycE expression pattern: a common feature of progenitor/stem cell populations
The high CycE levels during G2 in GSCs (this study) (Hsu et al., 2008) appear to be a common feature shared by multiple types of precursors/stem cells. In Drosophila, early syncytial embryonic divisions (Richardson et al., 1993; Sauer et al., 1995) and early histoblast divisions during metamorphosis (Ninov et al., 2009) lack a recognizable G1 owing to pre-accumulated stores of CycE. Mitotic germ cells in the C. elegans gonad divide rapidly with little or no G1, probably as a result of high CycE levels throughout the cell cycle (Brodigan et al., 2003; Fox et al., 2011). CycE is constitutively expressed in mammalian embryonic stem cells, where G1 is also very short (Singh and Dalton, 2009). The short G1 in GSCs is probably a direct consequence of high CycE levels during G2 and M. For example, the high expression of the CycE target Dup (the Drosophila Cdt1 homolog) during G2/M probably contributes to rapid entry into S phase following mitosis. Indeed, a recent study in embryonic stem cells suggests that high levels of human CDT1 during G2 and M phases leads to efficient DNA replication licensing after mitotic exit (Ballabeni et al., 2011). It is difficult, however, to address experimentally the specific consequences of having a short G1 in GSCs and other types of precursors/stem cells.
The loss of CycEWX and CycE1F36 GSCs (which have normal proliferation) indicates that CycE has a role in GSC maintenance that is genetically separable from its role in maintaining a short G1. Nevertheless, it remains possible that a short G1 contributes independently to GSC identity, consistent with the higher rate of GSC loss observed for null CycE mutant GSCs arrested in G1. It is also conceivable that high CycE activity throughout most of the cell cycle ensures more rapid cell cycles and minimizes excessive cell growth. Indeed, cystoblasts also share the atypical CycE expression and constitutive MPM2 pattern displayed by GSCs (our unpublished observations) (Hsu et al., 2008), and we found that G1-arrested null CycE GSCs or cystoblast-like cells grow excessively (unlike G2-arrested CycB mutant cells), apparently outcompeting neighboring wild-type cells for nutrients. These data are consistent with studies showing that CycE overexpression in mammalian fibroblasts shortens G1, reduces cell size and alleviates the dependence on extrinsic growth factors for the G1/S transition (Ohtsubo and Roberts, 1993; Resnitzky et al., 1994), potentially explaining why G2 is instead the major point of regulation by nutrient-dependent pathways in GSCs (Ables and Drummond-Barbosa, 2010; Hsu et al., 2008; LaFever et al., 2010). Finally, the presence of CycE throughout most of the cell cycle may contribute to more efficient signaling through key pathways, as suggested by our data that CycE stimulates BMP responsiveness in GSCs.
Regulation of CycE expression and activity in GSCs
The molecular mechanisms responsible for the atypical CycE pattern in GSCs remain unclear. In Drosophila somatic cells, CycE expression is activated by the transcription factor E2F, and its activity is repressed by the p21/p27 homolog, Dacapo (de Nooij et al., 1996; Duronio et al., 1998; Duronio and O'Farrell, 1995). It is therefore conceivable that altered E2F or Dacapo function in GSCs may contribute to the atypical CycE expression and activity. Other non-canonical regulators of CycE expression could also help shape its unusual expression pattern in GSCs. For example, in Drosophila neuroblasts, CycE expression is under control of homeotic genes (Berger et al., 2005). In the ovaries of mutants in mei-P26 (which encodes a Trim-NHL domain protein), CycE is highly expressed throughout oogenesis, suggesting that mei-P26 might modulate CycE (Neumüller et al., 2008). Several signaling molecules important for GSC maintenance, including the BMP signal Decapentaplegic, Hedgehog and Notch, regulate CycE in a complex context-dependent manner during Drosophila eye development (Duman-Scheel et al., 2002; Escudero and Freeman, 2007; Firth and Baker, 2005; Nicholson et al., 2011). It will be interesting to determine the role of extrinsic factors in establishing the atypical CycE pattern in GSCs.
Alternatively, CycE degradation might be differentially regulated in GSCs. Blocking the ability of the Skp/Cul/F-box (SCF) complex to target CycE for degradation can lead to its constitutive expression. For example, mutation of CycE degrons that control binding to SCFbw7 in knock-in mice disrupts CycE periodicity and causes its accumulation and increased activity (Minella et al., 2008). Similarly, mutation of the human F-box protein CDC4 in a cancer cell line leads to very high levels of CycE (Strohmaier et al., 2001). In early germ cells in the Drosophila ovary (Doronkin et al., 2003), mutation of the COP9 signalosome, an SCF regulator, results in uniformly high CycE expression; however, its function has not been directly examined in GSCs.
CycE in cancer cells
Cancer cells often express constitutively high levels of CycE (Siu et al., 2012). Some evidence suggests that high CycE levels in cancer cells may lead to genomic instability (Siu et al., 2012). For example, CycE overexpression in rat embryo fibroblasts or human breast epithelial cells results in chromosomal instability (Spruck et al., 1999). Mice expressing a stable version of CycE have an increased incidence of chromosomal breaks and translocations, and accelerated tumorigenesis in the absence of p53 (Loeb et al., 2005). Activated Ras promotes genetic instability and cancer formation by inhibiting SCFbw7-mediated degradation of CycE (Minella et al., 2005). These observations suggest that in normal precursors/stem cells with atypical CycE patterns, there must be mechanisms in place to ensure genomic stability despite high CycE levels. Nevertheless, there may also be roles of CycE that are shared between normal stem cells and cancers. For example, analogous to the function of CycE in stimulating BMP signaling in Drosophila GSCs, CycE may contribute to the stem cell-like potential of cancers to generate new cells, perhaps by boosting the signaling activity of key tumorigenic pathways.
Supplementary Material
Acknowledgements
We thank D. Kalderon, B. Duronio, M. Lilly, H. Lin, T. Orr-Weaver, H. Richardson, the Bloomington and Kyoto Stock Centers, and the Developmental Studies Hybridoma Bank for fly stocks and antibodies; and M. de Cuevas, A. Armstrong, K. Laws and anonymous reviewers for critical reading of this manuscript.
Footnotes
Funding
This work was supported by the National Institutes of Health (NIH) [R01 GM069875] and by the American Cancer Society [RSG-DDC-112316]. E.T.A. was supported by a National Research Service Award [F32 GM086031] from the NIH. Deposited in PMC for release after 12 months.
Competing interests statement
The authors declare no competing financial interests.
Supplementary material
Supplementary material available online at http://dev.biologists.org/lookup/suppl/doi:10.1242/dev.088583/-/DC1
References
- Ables E. T., Drummond-Barbosa D. (2010). The steroid hormone ecdysone functions with intrinsic chromatin remodeling factors to control female germline stem cells in Drosophila. Cell Stem Cell 7, 581-592 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ables E. T., Laws K. M., Drummond-Barbosa D. (2012). Control of adult stem cells in vivo by a dynamic physiological environment: diet-dependent systemic factors in Drosophila and beyond. WIREs Dev Biol 1, 657-674 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ashburner M., Drysdale R. (1994). FlyBase – the Drosophila genetic database. Development 120, 2077-2079 [DOI] [PubMed] [Google Scholar]
- Ballabeni A., Park I. H., Zhao R., Wang W., Lerou P. H., Daley G. Q., Kirschner M. W. (2011). Cell cycle adaptations of embryonic stem cells. Proc. Natl. Acad. Sci. USA 108, 19252-19257 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berger C., Pallavi S. K., Prasad M., Shashidhara L. S., Technau G. M. (2005). A critical role for cyclin E in cell fate determination in the central nervous system of Drosophila melanogaster. Nat. Cell Biol. 7, 56-62 [DOI] [PubMed] [Google Scholar]
- Berger C., Kannan R., Myneni S., Renner S., Shashidhara L. S., Technau G. M. (2010). Cell cycle independent role of Cyclin E during neural cell fate specification in Drosophila is mediated by its regulation of Prospero function. Dev. Biol. 337, 415-424 [DOI] [PubMed] [Google Scholar]
- Brodigan T. M., Liu J., Park M., Kipreos E. T., Krause M. (2003). Cyclin E expression during development in Caenorhabditis elegans. Dev. Biol. 254, 102-115 [DOI] [PubMed] [Google Scholar]
- Brumby A., Secombe J., Horsfield J., Coombe M., Amin N., Coates D., Saint R., Richardson H. (2004). A genetic screen for dominant modifiers of a cyclin E hypomorphic mutation identifies novel regulators of S-phase entry in Drosophila. Genetics 168, 227-251 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calvi B. R., Lilly M. A., Spradling A. C. (1998). Cell cycle control of chorion gene amplification. Genes Dev. 12, 734-744 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen D., Wang Q., Huang H., Xia L., Jiang X., Kan L., Sun Q., Chen D. (2009). Effete-mediated degradation of Cyclin A is essential for the maintenance of germline stem cells in Drosophila. Development 136, 4133-4142 [DOI] [PubMed] [Google Scholar]
- Chen S., Wang S., Xie T. (2011). Restricting self-renewal signals within the stem cell niche: multiple levels of control. Curr. Opin. Genet. Dev. 21, 684-689 [DOI] [PubMed] [Google Scholar]
- Choi J. H., Banks A. S., Estall J. L., Kajimura S., Boström P., Laznik D., Ruas J. L., Chalmers M. J., Kamenecka T. M., Blüher M., et al. (2010). Anti-diabetic drugs inhibit obesity-linked phosphorylation of PPARgamma by Cdk5. Nature 466, 451-456 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis F. M., Tsao T. Y., Fowler S. K., Rao P. N. (1983). Monoclonal antibodies to mitotic cells. Proc. Natl. Acad. Sci. USA 80, 2926-2930 [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Cuevas M., Spradling A. C. (1998). Morphogenesis of the Drosophila fusome and its implications for oocyte specification. Development 125, 2781-2789 [DOI] [PubMed] [Google Scholar]
- de Nooij J. C., Letendre M. A., Hariharan I. K. (1996). A cyclin-dependent kinase inhibitor, Dacapo, is necessary for timely exit from the cell cycle during Drosophila embryogenesis. Cell 87, 1237-1247 [DOI] [PubMed] [Google Scholar]
- Doronkin S., Djagaeva I., Beckendorf S. K. (2003). The COP9 signalosome promotes degradation of Cyclin E during early Drosophila oogenesis. Dev. Cell 4, 699-710 [DOI] [PubMed] [Google Scholar]
- Duman-Scheel M., Weng L., Xin S., Du W. (2002). Hedgehog regulates cell growth and proliferation by inducing Cyclin D and Cyclin E. Nature 417, 299-304 [DOI] [PubMed] [Google Scholar]
- Duronio R. J., O'Farrell P. H. (1995). Developmental control of the G1 to S transition in Drosophila: cyclin Eis a limiting downstream target of E2F. Genes Dev. 9, 1456-1468 [DOI] [PubMed] [Google Scholar]
- Duronio R. J., Bonnette P. C., O'Farrell P. H. (1998). Mutations of the Drosophila dDP, dE2F, and cyclin E genes reveal distinct roles for the E2F-DP transcription factor and cyclin E during the G1-S transition. Mol. Cell. Biol. 18, 141-151 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Escudero L. M., Freeman M. (2007). Mechanism of G1 arrest in the Drosophila eye imaginal disc. BMC Dev. Biol. 7, 13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Firth L. C., Baker N. E. (2005). Extracellular signals responsible for spatially regulated proliferation in the differentiating Drosophila eye. Dev. Cell 8, 541-551 [DOI] [PubMed] [Google Scholar]
- Fox P. M., Vought V. E., Hanazawa M., Lee M. H., Maine E. M., Schedl T. (2011). Cyclin E and CDK-2 regulate proliferative cell fate and cell cycle progression in the C. elegans germline. Development 138, 2223-2234 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilboa L., Forbes A., Tazuke S. I., Fuller M. T., Lehmann R. (2003). Germ line stem cell differentiation in Drosophila requires gap junctions and proceeds via an intermediate state. Development 130, 6625-6634 [DOI] [PubMed] [Google Scholar]
- Hong A., Lee-Kong S., Iida T., Sugimura I., Lilly M. A. (2003). The p27cip/kip ortholog dacapo maintains the Drosophila oocyte in prophase of meiosis I. Development 130, 1235-1242 [DOI] [PubMed] [Google Scholar]
- Hsu H. J., Drummond-Barbosa D. (2009). Insulin levels control female germline stem cell maintenance via the niche in Drosophila. Proc. Natl. Acad. Sci. USA 106, 1117-1121 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsu H. J., LaFever L., Drummond-Barbosa D. (2008). Diet controls normal and tumorous germline stem cells via insulin-dependent and -independent mechanisms in Drosophila. Dev. Biol. 313, 700-712 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inaba M., Yuan H., Yamashita Y. M. (2011). String (Cdc25) regulates stem cell maintenance, proliferation and aging in Drosophila testis. Development 138, 5079-5086 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jacobs H. W., Knoblich J. A., Lehner C. F. (1998). Drosophila Cyclin B3 is required for female fertility and is dispensable for mitosis like Cyclin B. Genes Dev. 12, 3741-3751 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jeong J., Verheyden J. M., Kimble J. (2011). Cyclin E and Cdk2 control GLD-1, the mitosis/meiosis decision, and germline stem cells in Caenorhabditis elegans. PLoS Genet. 7, e1001348 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kai T., Spradling A. (2003). An empty Drosophila stem cell niche reactivates the proliferation of ectopic cells. Proc. Natl. Acad. Sci. USA 100, 4633-4638 [DOI] [PMC free article] [PubMed] [Google Scholar]
- LaFever L., Feoktistov A., Hsu H. J., Drummond-Barbosa D. (2010). Specific roles of Target of rapamycin in the control of stem cells and their progeny in the Drosophila ovary. Development 137, 2117-2126 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lange C., Calegari F. (2010). Cdks and cyclins link G1 length and differentiation of embryonic, neural and hematopoietic stem cells. Cell Cycle 9, 1893-1900 [DOI] [PubMed] [Google Scholar]
- Lantz V., Chang J. S., Horabin J. I., Bopp D., Schedl P. (1994). The Drosophila orb RNA-binding protein is required for the formation of the egg chamber and establishment of polarity. Genes Dev. 8, 598-613 [DOI] [PubMed] [Google Scholar]
- Li V. C., Ballabeni A., Kirschner M. W. (2012). Gap 1 phase length and mouse embryonic stem cell self-renewal. Proc. Natl. Acad. Sci. USA 109, 12550-12555 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lilly M. A., Spradling A. C. (1996). The Drosophila endocycle is controlled by Cyclin E and lacks a checkpoint ensuring S-phase completion. Genes Dev. 10, 2514-2526 [DOI] [PubMed] [Google Scholar]
- Lilly M. A., de Cuevas M., Spradling A. C. (2000). Cyclin A associates with the fusome during germline cyst formation in the Drosophila ovary. Dev. Biol. 218, 53-63 [DOI] [PubMed] [Google Scholar]
- Loeb K. R., Kostner H., Firpo E., Norwood T., Tsuchiya K. D., Clurman B. E., Roberts J. M. (2005). A mouse model for cyclin E-dependent genetic instability and tumorigenesis. Cancer Cell 8, 35-47 [DOI] [PubMed] [Google Scholar]
- Minella A. C., Welcker M., Clurman B. E. (2005). Ras activity regulates cyclin E degradation by the Fbw7 pathway. Proc. Natl. Acad. Sci. USA 102, 9649-9654 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Minella A. C., Loeb K. R., Knecht A., Welcker M., Varnum-Finney B. J., Bernstein I. D., Roberts J. M., Clurman B. E. (2008). Cyclin E phosphorylation regulates cell proliferation in hematopoietic and epithelial lineages in vivo. Genes Dev. 22, 1677-1689 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Möröy T., Geisen C. (2004). Cyclin E. Int. J. Biochem. Cell Biol. 36, 1424-1439 [DOI] [PubMed] [Google Scholar]
- Myster D. L., Bonnette P. C., Duronio R. J. (2000). A role for the DP subunit of the E2F transcription factor in axis determination during Drosophila oogenesis. Development 127, 3249-3261 [DOI] [PubMed] [Google Scholar]
- Narbonne-Reveau K., Lilly M. (2009). The Cyclin-dependent kinase inhibitor Dacapo promotes genomic stability during premeiotic S phase. Mol. Biol. Cell 20, 1960-1969 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neganova I., Zhang X., Atkinson S., Lako M. (2009). Expression and functional analysis of G1 to S regulatory components reveals an important role for CDK2 in cell cycle regulation in human embryonic stem cells. Oncogene 28, 20-30 [DOI] [PubMed] [Google Scholar]
- Neumüller R. A., Betschinger J., Fischer A., Bushati N., Poernbacher I., Mechtler K., Cohen S. M., Knoblich J. A. (2008). Mei-P26 regulates microRNAs and cell growth in the Drosophila ovarian stem cell lineage. Nature 454, 241-245 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicholson S. C., Nicolay B. N., Frolov M. V., Moberg K. H. (2011). Notch-dependent expression of the archipelago ubiquitin ligase subunit in the Drosophila eye. Development 138, 251-260 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ninov N., Manjón C., Martín-Blanco E. (2009). Dynamic control of cell cycle and growth coupling by ecdysone, EGFR, and PI3K signaling in Drosophila histoblasts. PLoS Biol. 7, e1000079 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Odajima J., Wills Z. P., Ndassa Y. M., Terunuma M., Kretschmannova K., Deeb T. Z., Geng Y., Gawrzak S., Quadros I. M., Newman J., et al. (2011). Cyclin E constrains Cdk5 activity to regulate synaptic plasticity and memory formation. Dev. Cell 21, 655-668 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohlmeyer J. T., Schüpbach T. (2003). Encore facilitates SCF-Ubiquitin-proteasome-dependent proteolysis during Drosophila oogenesis. Development 130, 6339-6349 [DOI] [PubMed] [Google Scholar]
- Ohtsubo M., Roberts J. M. (1993). Cyclin-dependent regulation of G1 in mammalian fibroblasts. Science 259, 1908-1912 [DOI] [PubMed] [Google Scholar]
- Orford K. W., Scadden D. T. (2008). Deconstructing stem cell self-renewal: genetic insights into cell-cycle regulation. Nat. Rev. Genet. 9, 115-128 [DOI] [PubMed] [Google Scholar]
- Park E. A., Macalpine D. M., Orr-Weaver T. L. (2007). Drosophila follicle cell amplicons as models for metazoan DNA replication: a cyclinE mutant exhibits increased replication fork elongation. Proc. Natl. Acad. Sci. USA 104, 16739-16746 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rangan P., Malone C. D., Navarro C., Newbold S. P., Hayes P. S., Sachidanandam R., Hannon G. J., Lehmann R. (2011). piRNA production requires heterochromatin formation in Drosophila. Curr. Biol. 21, 1373-1379 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Resnitzky D., Gossen M., Bujard H., Reed S. I. (1994). Acceleration of the G1/S phase transition by expression of cyclins D1 and E with an inducible system. Mol. Cell. Biol. 14, 1669-1679 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richardson H. E., O'Keefe L. V., Reed S. I., Saint R. (1993). A Drosophila G1-specific cyclin E homolog exhibits different modes of expression during embryogenesis. Development 119, 673-690 [DOI] [PubMed] [Google Scholar]
- Richardson H., O'Keefe L. V., Marty T., Saint R. (1995). Ectopic cyclin E expression induces premature entry into S phase and disrupts pattern formation in the Drosophila eye imaginal disc. Development 121, 3371-3379 [DOI] [PubMed] [Google Scholar]
- Salomoni P., Calegari F. (2010). Cell cycle control of mammalian neural stem cells: putting a speed limit on G1. Trends Cell Biol. 20, 233-243 [DOI] [PubMed] [Google Scholar]
- Sauer K., Knoblich J. A., Richardson H., Lehner C. F. (1995). Distinct modes of cyclin E/cdc2c kinase regulation and S-phase control in mitotic and endoreduplication cycles of Drosophila embryogenesis. Genes Dev. 9, 1327-1339 [DOI] [PubMed] [Google Scholar]
- Singh A. M., Dalton S. (2009). The cell cycle and Myc intersect with mechanisms that regulate pluripotency and reprogramming. Cell Stem Cell 5, 141-149 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siu K. T., Rosner M. R., Minella A. C. (2012). An integrated view of cyclin E function and regulation. Cell Cycle 11, 57-64 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song X., Zhu C. H., Doan C., Xie T. (2002). Germline stem cells anchored by adherens junctions in the Drosophila ovary niches. Science 296, 1855-1857 [DOI] [PubMed] [Google Scholar]
- Spruck C. H., Won K. A., Reed S. I. (1999). Deregulated cyclin E induces chromosome instability. Nature 401, 297-300 [DOI] [PubMed] [Google Scholar]
- Strohmaier H., Spruck C. H., Kaiser P., Won K. A., Sangfelt O., Reed S. I. (2001). Human F-box protein hCdc4 targets cyclin E for proteolysis and is mutated in a breast cancer cell line. Nature 413, 316-322 [DOI] [PubMed] [Google Scholar]
- Sukhanova M. J., Du W. (2008). Control of cell cycle entry and exiting from the second mitotic wave in the Drosophila developing eye. BMC Dev. Biol. 8, 7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomer M., May N. R., Aggarwal B. D., Kwok G., Calvi B. R. (2004). Drosophila double-parked is sufficient to induce re-replication during development and is regulated by cyclin E/CDK2. Development 131, 4807-4818 [DOI] [PubMed] [Google Scholar]
- Ugland H., Boquest A. C., Naderi S., Collas P., Blomhoff H. K. (2008). cAMP-mediated induction of cyclin E sensitizes growth-arrested adipose stem cells to DNA damage-induced apoptosis. Mol. Biol. Cell 19, 5082-5092 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Z. A., Kalderon D. (2009). Cyclin E-dependent protein kinase activity regulates niche retention of Drosophila ovarian follicle stem cells. Proc. Natl. Acad. Sci. USA 106, 21701-21706 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Z., Lin H. (2005). The division of Drosophila germline stem cells and their precursors requires a specific cyclin. Curr. Biol. 15, 328-333 [DOI] [PubMed] [Google Scholar]
- White A. E., Leslie M. E., Calvi B. R., Marzluff W. F., Duronio R. J. (2007). Developmental and cell cycle regulation of the Drosophila histone locus body. Mol. Biol. Cell 18, 2491-2502 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whittaker A. J., Royzman I., Orr-Weaver T. L. (2000). Drosophila double parked: a conserved, essential replication protein that colocalizes with the origin recognition complex and links DNA replication with mitosis and the down-regulation of S phase transcripts. Genes Dev. 14, 1765-1776 [PMC free article] [PubMed] [Google Scholar]
- Xi R., Xie T. (2005). Stem cell self-renewal controlled by chromatin remodeling factors. Science 310, 1487-1489 [DOI] [PubMed] [Google Scholar]
- Xi R., Doan C., Liu D., Xie T. (2005). Pelota controls self-renewal of germline stem cells by repressing a Bam-independent differentiation pathway. Development 132, 5365-5374 [DOI] [PubMed] [Google Scholar]
- Xie T., Spradling A. C. (1998). decapentaplegic is essential for the maintenance and division of germline stem cells in the Drosophila ovary. Cell 94, 251-260 [DOI] [PubMed] [Google Scholar]
- Xu T., Rubin G. M. (1993). Analysis of genetic mosaics in developing and adult Drosophila tissues. Development 117, 1223-1237 [DOI] [PubMed] [Google Scholar]
- Yu S. Y., Yoo S. J., Yang L., Zapata C., Srinivasan A., Hay B. A., Baker N. E. (2002). A pathway of signals regulating effector and initiator caspases in the developing Drosophila eye. Development 129, 3269-3278 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.