

Abstract
The production of induced pluripotent stem cells (iPSCs) from somatic cells provides a means to create valuable tools for basic research and may also produce a source of patient-matched cells for regenerative therapies. iPSCs may be generated using multiple protocols and derived from multiple cell sources. Once generated, iPSCs are tested using a variety of assays including immunostaining for pluripotency markers, generation of three germ layers in embryoid bodies and teratomas, comparisons of gene expression with embryonic stem cells (ESCs) and production of chimeric mice with or without germline contribution2. Importantly, iPSC lines that pass these tests still vary in their capacity to produce different differentiated cell types2. This has made it difficult to establish which iPSC derivation protocols, donor cell sources or selection methods are most useful for different applications.
The most stringent test of whether a stem cell line has sufficient developmental potential to generate all tissues required for survival of an organism (termed full pluripotency) is tetraploid embryo complementation (TEC)3-5. Technically, TEC involves electrofusion of two-cell embryos to generate tetraploid (4n) one-cell embryos that can be cultured in vitro to the blastocyst stage6. Diploid (2n) pluripotent stem cells (e.g. ESCs or iPSCs) are then injected into the blastocoel cavity of the tetraploid blastocyst and transferred to a recipient female for gestation (see Figure 1). The tetraploid component of the complemented embryo contributes almost exclusively to the extraembryonic tissues (placenta, yolk sac), whereas the diploid cells constitute the embryo proper, resulting in a fetus derived entirely from the injected stem cell line.
Recently, we reported the derivation of iPSC lines that reproducibly generate adult mice via TEC1. These iPSC lines give rise to viable pups with efficiencies of 5-13%, which is comparable to ESCs3,4,7 and higher than that reported for most other iPSC lines8-12. These reports show that direct reprogramming can produce fully pluripotent iPSCs that match ESCs in their developmental potential and efficiency of generating pups in TEC tests. At present, it is not clear what distinguishes between fully pluripotent iPSCs and less potent lines13-15. Nor is it clear which reprogramming methods will produce these lines with the highest efficiency. Here we describe one method that produces fully pluripotent iPSCs and "all- iPSC" mice, which may be helpful for investigators wishing to compare the pluripotency of iPSC lines or establish the equivalence of different reprogramming methods.
Keywords: Stem Cell Biology, Issue 69, Molecular Biology, Developmental Biology, Medicine, Cellular Biology, Induced pluripotent stem cells, iPSC, stem cells, reprogramming, developmental potential, tetraploid embryo complementation, mouse
Protocol
This method was used in the research reported in Boland et al. Nature. 461, 91-96 (2009).1
1. Preparation of Lentivirus
This protocol employs doxycycline-inducible lentiviral shuttle vectors that encode for Oct4, Sox2, Klf4, and c-Myc under control of a tetO response element. Transgenes are activated by the reverse tetracycline trans-activating protein, rtTAM2.216, which induces reprogramming factor expression in the presence of doxycycline. This system allows for tightly controlled, high expression of reprogramming factors. The lentiviral vectors used here are self-inactivating and thus cannot replicate following genomic integration. However, caution is required when working with lentiviruses and should be performed in laboratories compliant with BSL2 (USA) and S2 (Europe) standards.
Thaw HEK293T cells and passage at least once before transfection. Cells should be maintained at subconfluent density in HEK medium at 37 °C, 5% CO2 in a humidified environment. Routinely passage cells every 2 days with a split ratio of 1:6-1:10.
Seed ~8 x 106 HEK293T cells/T150 with 25 ml HEK medium. Use one T150 for each lentivirus preparation.
The following day transfect HEK293T cells by calcium phosphate precipitation. (Note: in our hands calcium phosphate precipitation routinely results in 80-90% transfection efficiency; however, cationic lipid transfection reagents such as Lipofectamine 2,000 may also be used). Prepare two 15 ml conical tubes for each virus to be prepared. Label the tubes "A" and "B". Tube A, 10 μg of each of: the lentiviral shuttle vector encoding reprogramming factors (or rtTAM2.2), the viral packaging vectors, and plasmid encoding the viral envelope protein, VSVg. Add 186 μl 2 M CaCl2 to Tube A, and bring the volume to 1.5 ml with sterile H2O. Tube B: 1.5 ml 2x HBS (pre-warmed to room temperature).
Pipette the mixture in Tube A until it is a homogenous solution. Add solution A to solution B dropwise and let stand at room temperature for 2-3 min.
Aspirate growth medium of HEK293T cells and replace with 22 ml pre-warmed HEK medium without penicillin and streptomycin.
Pipette combined solutions AB (calcium phosphate precipitate) directly to HEK293T cells and distribute evenly by gentle rocking.
24 hr after transfection of HEK-293T cells with lentivirus, remove growth medium and replace it with 25 ml of fresh, pre-warmed HEK medium. Return transfected HEKs to the incubator.
48 hr after transfection of HEK-293T cells with lentivirus, collect growth media containing lentiviral particles from the transfected HEKs. Remove particulate debris from the harvested viral solution by centrifugation at 3,000 x g for 5 min at 4 °C.
Concentrate virus by ultracentrifugation through a 20% sucrose cushion (2 ml sucrose/25 ml viral supernatant) for 2 hr at 112,000 x g at 4 °C. Suspend viral pellet in 0.4 ml MEF medium at 4 °C for 15-30 min with gentle rocking. Store viral particles in single use aliquots (i.e. 50 μl) at -80 °C.
2. Preparation of Mouse Embryonic Fibroblasts (MEF) for Reprogramming
Note: The protocol outlined here relates to the derivation of iPSCs from E 13.5 mouse embryonic fibroblasts for use in TEC assays. While other groups have generated all-iPSC mice from adult donor cell sources, we have not tested this method on other cell types and cannot be certain that donor cell type is not a factor.
Set up mouse timed matings. On embryonic day 13.5 (E13.5), euthanize the pregnant female and dissect the embryos from the uterine horns. Place and store embryos in 1x PBS (pre-chilled to 4 °C) on ice.
Remove the extraembryonic tissues (i.e. chorion, amnion and placenta). Decapitate the embryo and remove the tail (optional - if needed for genotyping) and limbs. Scoop out the internal organs, using forceps or a scoop shaped spatula and mince the remaining carcass with the blade of a scalpel or sharp scissors.
Wash the minced carcass in 5 ml pre-chilled 1x PBS. Centrifuge at 200 x g for 5 min.
Aspirate the supernatant. Suspend pellet in 5 ml 0.25% Trypsin-EDTA and incubate at 37 °C with vigorous shaking for 20-30 min.
Add 5 ml of MEF medium (pre-warmed to 37 °C), mix and centrifuge at 200 x g for 5 min.
Aspirate supernatant and resuspend pellet in pre-warmed MEF medium.
Plate dissociated MEFs in 2-3 wells of a 6-well plate pre-coated with 0.1% gelatin. This is considered passage 1.
Passage MEFs at a dilution of 1:4-1:5 every 48 hr. MEFs at passage 3 are ready for lentiviral transduction.
3. Derivation of iPSC Lines
The day before lentiviral transduction; seed ~3 x 105 primary MEFs into one well of a 6-well plate pre-coated with 0.1% gelatin.
Day 1: Primary MEFS should be 80-90% confluent for lentiviral transduction. Add lentiviral particles directly to MEF media and incubate with MEFs overnight at 37 °C, 5% CO2 in a humidified environment.
The following day (day 2) aspirate the medium and wash twice with 3 ml of 1x PBS to remove viral particles.
Add 0.5 ml pre-warmed 0.25% Trypsin-EDTA to the cells and incubate at 37 °C for 3-5 min with occasional rocking.
Triturate to achieve a single-cell suspension. Observe cells by light microscopy to ensure a single cell suspension.
Transfer MEFs into a 15 ml conical tube containing 5 ml MEF media. Centrifuge at 200 x g for 5 min. Aspirate the supernatant and gently resuspend cells in MEF medium.
Evenly split the cell suspension between two wells of a 6 well plate pre-coated with 0.1% gelatin.
Rock the plate back and forth, side to side, and once in a circular motion to achieve an even distribution of cells throughout the well. Incubate overnight at 37 °C, 5% CO2 in a humidified environment.
Day 3: Repeat steps 4-9 except split the cells evenly from one well to 3 wells of a 6 well plate pre-coated with 0.1% gelatin. This will yield 6 wells of transduced primary fibroblasts.
Day 4: Add doxycycline (dox) at a concentration of 10 μg/ml to 5/6 wells. One well should remain untreated to serve as a control.
Add VPA at 1.9 mM to 3 of the 5 wells treated with dox. VPA reduces the proliferation rate of MEFs. Dense cultures of MEFs tolerate prolonged exposure to VPA whereas subconfluent cultures tend to senesce within 2-5 days. Therefore, MEFS should be 100% confluent when VPA is added. Note: We use VPA in our reprogramming experiments because it is a known epigenetic modifier, and has been shown to increase the efficiency of iPSC generation17 although the effects and mechanisms of VPA action with respect to generating fully pluripotent iPSC lines are not known.
Day 5: Aspirate the medium, wash and trypsinize the cells as before. Passage the cells treated with dox/VPA to a 15 cm2 tissue culture dish pre-coated with 0.1% gelatin and each of the other conditions to pre-coated 10 cm2 dishes in ES cell medium supplemented with fresh dox and VPA.
Replenish cells every day with ESC medium supplemented with fresh dox and VPA. ESC-like colonies should begin to appear after ~7 days in the dox/VPA treatment and after ~10 days in the dox alone treatment. No colonies should appear in the absence of dox treatment.
Once colonies possess a bright refractive, well-defined border and contain 30-50 cells, manually isolate the colonies with a gel loading pipette tip and transfer to a U-bottom 96 well plate containing 20 μl of 0.25% trypsin-EDTA. Trypsinize to single cells and transfer to feeders in a flat bottom 96 well plate in 150 μl ESC medium containing dox/VPA or dox alone.
Continue to clonally expand the isolated iPSC lines on feeders in ESC medium. Remove dox and VPA on day 19 after their addition (post-transduction day 23). Discard iPSC lines that do not maintain self-renewal or proliferation rates similar to ESC controls.
It may be helpful to characterize your iPSC lines in relation to ESCs before attempting to perform TEC. We have characterized our lines by 1) expression of endogenous pluripotency markers (SSEA-1, Oct4, Sox2, Nanog) by immunocytochemistry, 2) karyotype analysis by chromosome counting and 3) embryoid body formation. One may also perform lentiviral-specific RT-qPCR to confirm that the proviral transgenes are not expressed in the iPSCs. However, we have identified fully pluripotent iPSCs using only morphology, immunostaining and karyotyping. In our experiments, selection of iPSC lines based on ESC-like morphology and growth characteristics results in the majority of the lines expressing pluripotency markers while we typically identify several lines with potentially abnormal karyotypes.
4. Preparation of iPSCs for Blastocyst Injection
Passage number of a PSC line has been shown to affect its pluripotency18 although this may be line dependent19. We have used iPSCs of from passages 8-14 to produce adult all-iPSC mice.
Thaw iPSCs and plate on feeders in ESC medium. Passage the cells at least once on feeders before use for injection.
One well of a 6-well plate containing 70-80% confluent iPSCs will provide more than a sufficient number of cells for injection. Aspirate growth medium and wash the cells with ~3 ml 1x PBS (without Ca2+/Mg2+).
Add 0.5 ml pre-warmed 0.05% Trypsin-EDTA to the cells and incubate at 37 °C for 10 min with occasional rocking.
Triturate to achieve a single-cell suspension. Observe cells by light microscopy to ensure a single cell suspension. The iPSCs need to be in single cell suspension as colonies/cell aggregates will clog the injection pipette.
Once a single cell suspension has ben achieved, add 1.0 ml ESC media to the well and return the plate to the 37 °C incubator. Incubate for ~15 min or until the majority of feeders have begun to adhere.
Gently remove the medium containing the iPSCs taking care not to dislodge the weakly adherent feeders.
Place the iPSCs in a 15 ml conical tube containing 5 ml ESC medium. Centrifuge at 200 x g for 5 min. Aspirate the supernatant and remove the remainder of ES cell medium with a micropipette. Tap the tube to dislodge the pellet and gently resuspend cells in 0.2-0.5 ml pre-chilled FHM medium. Store cells on ice until and during injection into tetraploid blastocysts.
5. Generation of Tetraploid Blastocysts
Procedures performed in this section have been described in detail elsewhere5,6,20. Here we outline our technique, optimized for the BTX Electro Cell Manipulator ECM 2001.
Set up embryo donor mice by priming 23-28 day old female mice (C57BL/6J-Tyrc-2J /BALB/cByJ F1) with PMS and HCG. Administer 5 IU of PMS at 2PM and 5 IU of HCG 47 hr later. After HCG injection, set up female mice with C57BL/6J-Tyrc-2J /BALB/cByJ F1 stud males. Check the following day for vaginal plugs.
Euthanize plugged female mice and collect oviducts. Collect 1-cell embryos by placing oviducts in FHM with Hyaluronidase and gently tearing the ampulae. Allow the cumulus masses to sit in FHM/Hyaluronidase for 5-7 min.
Collect 1-cell embryos using a mouth pipette and wash through drops of FHM media before placing them in KSOM-AA culture. Culture at 37 °C, 5% CO2 under mineral oil overnight and select 2-cell embryos the day of electrofusion, discard all other embryos.
Place a BTX Microslide in a 10 cm Petri dish. Pour enough room temperature electrofusion media to submerge the slide in the solution, but not so much that the poles of the electrode are completely submerged.
Switch on ECM 2001 and BTX Enhancer 400. Connect the ECM's cables to the microslide's electrode and fix the cables to the side of the Petri dish to prevent unintended movement of the slide.
Run one manual pulse to get a reading on the BTX enhancer and note the voltage of the AC/DC currents being applied. AC current will control the speed at which the embryos will align between the electrodes, DC current will fuse the blastomeres, and pulse time will set the length of the DC pulse. A good starting point is AC 3V, DC 100V, and time 0.05 msec. The optimal DC varies in the range of 90-150 volts.
Using a mouth pipette, take about 30-40 two-cell embryos from KSOM-AA culture and wash them through several drops of electrofusion medium. Draw fresh electrofusion media from the microslide dish into mouth pipette and take embryos from the wash. Place them in the 1 mm gap between the electrodes on the microslide. Be careful that they are aligned down the middle of the gap and that they are not in contact with each other.
Apply AC current by pressing the manual pulse button. The embryos will rotate in the AC field, until the plane of blastomere contact is parallel to the electrodes. If embryos are not aligned in a few seconds, increase AC setting.
After embryos have aligned, press the manual pulse button again to apply the DC pulse.
With electrofusion medium in the pipette, collect the embryos from the microslide. Wash embryos through several drops of KSOM-AA and place them in KSOM-AA culture at 37 °C, 5% CO2. The blastomere fusion should be completed in less than 30 min in culture.
Repeat steps 7-11 for remaining 2-cell embryos. After subsequent fusion groups, monitor and select embryos with fused blastomeres. Successfully fused embryos will appear to be in 1-cell stage. Discard lysed and 2-cell embryos after 30 min in culture. If fusion rate is below 80%, increase voltage and/or time in increments of 5V and 0.01 msec. If lysis is above 20%, decrease DC voltage and/or time accordingly. The optimal settings in our experiments were AC 4V, DC 146V, and 0.07 msec. These settings consistently yielded 90% or higher fusion rates with little or no lysis.
Continue to culture fused embryos in microdrops of KSOM-AA under mineral oil at 37 °C, 5% CO2. You should expect 85-95% of fused embryos to form tetraploid (4n) blastocysts after 48 hr of incubation.
6. Microinjection of iPSCs into Tetraploid Blastocysts
We use a Nikon TE-2000U inverted microscope equipped with DIC optics and Narishige micromanipulators for blastocyst injection. Each tetraploid blastocyst is injected with 10-12 iPSCs using a standard protocol for ESC injection into mouse blastocysts that has been demonstrated in a previous JoVE publication5,20,21
Place a 20 μl drop of FHM in the center of a concave microscope slide and cover it with 150 μl of mineral oil.
Lower the holding pipette and microinjection needle into the FHM drop. Allow 2-3 min for both needles to partially fill with FHM.
Wash 20-30 tetraploid blastocysts through drops of FHM and transfer to the FHM drop on the microscope slide.
Mouth pipette iPSC mixture into the drop. It may be necessary to dilute the cell mixture in a drop of FHM beforehand if cells are too concentrated or aggregated.
Pick up 100-200 cells with the injection needle.
Hold the blastocyst with the inner cell mass in the 9 o'clock position. Inject cells into blastocoel by penetrating the zona pellucida and trophoblast at the 3 o'clock position. Inject 16-18 cells per blastocyst.
Return iPSC complemented blastocysts to KSOM-AA culture.
7. Transfer of Complemented Tetraploid Blastocysts into the Uterine Horns of Recipient Mice
Complemented tetraploid blastocysts are surgically transferred to the uterine horns of female recipient mice according to the guidelines of the researcher's institute, using the standard technique20 which we shall briefly summarize. Select female CD-1 mice at the pro-estrus stage and set them up for mating with vasectomized males. Check for vaginal plugs the next morning. Females are ready for uterine embryo transfer two days after the plug was detected (2.5 dpc).
One day before recipient females are mated with vasectomized males, set up additional CD-1 females with non-vasectomized males to be used as foster mothers for all-iPSC mice retrieved by Caesarian section.
8. Caesarean Section and Fostering of iPSC-derived Pups
The transfer of TC embryos typically results in multiple resorptions after the implantation, even if the iPSC or ESC line has a high developmental potential. As a result, one can expect not more than 4 viable pups (usually 1-2) per recipient. These small litters are usually neglected by recipients. To increase the level of neonatal care and the rate of survival, we perform C-sections and fostering according to the standard protocols20. To perform the Caesarean section, euthanize recipient mice 16 days after embryo transfer at 7-8PM (recipient 18.5 dpc) and dissect pups from the uterine horns. Foster viable pups to CD-1 mothers that delivered litters the same day.
Representative Results
In step 3, "Derivation of iPSCs from MEFs", one should observe morphological heterogeneity and immature iPSC colony formation starting 4-5 days after doxycycline/VPA addition and mature colonies between 7-10 days (Figure 2). The production of one-cell tetraploid embryos in step 5 is highly efficient (Figure 3). We routinely observe up to 95% of treated two-cell embryos successfully fuse to produce tetraploid one-cell embryos. The protocol followed to inject iPSCs into tetraploid blastocysts (Step 6, Figure 4) is similar to the protocol for injection of ESCs into diploid blastocysts to generate chimeric mice, and can be performed by an experienced microinjectionist. The number of live pups born depends on the cell line (Table 1).
| MEF preprogramming efficiency 0.01-0.03% | ||||
| Efficiency of iPSC mouse production by TEC | ||||
| Name | Description | Blastocysts injected | Live Newborn | Live Adult |
| iMZ-21 | iPSC | 867 | 53 (6.1%) | 19 (2.2%) |
| iMZ-9 | iPSC | 195 | 7 (3.6%) | 4 (2.1%) |
| iMZ-11 | iPSC | 338 | 1(0.3%) | 0 (0%) |
Table 1. Representative Results.
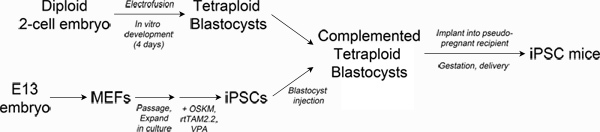 Figure 1. Schematic of experimental design. Top left: Production of tetraploid blastocysts. Fertilized two-cell embryos from albino mice are electrofused to generate tetraploid one-cell embryos, which are cultured in vitro to the blastocyst stage. Bottom left: Reprogramming. Mouse embryonic fibroblasts are transduced with lentiviral particles encoding Oct4, Sox2, Klf4 and c-Myc and the reverse tetracycline transactivating protein, rtTAM2.2. Addition of doxycycline results in transgene expression and the initiation of reprogramming to iPSCs. Right: Production of iPSC mice. iPSCs derived from pigmented mice are injected into the blastocoel of tetraploid blastocysts and then surgically implanted into pseudo-pregnant recipient mice. Newborn iPSC mice are delivered by Caesarian section and cross-fostered. Click here to view larger figure.
Figure 1. Schematic of experimental design. Top left: Production of tetraploid blastocysts. Fertilized two-cell embryos from albino mice are electrofused to generate tetraploid one-cell embryos, which are cultured in vitro to the blastocyst stage. Bottom left: Reprogramming. Mouse embryonic fibroblasts are transduced with lentiviral particles encoding Oct4, Sox2, Klf4 and c-Myc and the reverse tetracycline transactivating protein, rtTAM2.2. Addition of doxycycline results in transgene expression and the initiation of reprogramming to iPSCs. Right: Production of iPSC mice. iPSCs derived from pigmented mice are injected into the blastocoel of tetraploid blastocysts and then surgically implanted into pseudo-pregnant recipient mice. Newborn iPSC mice are delivered by Caesarian section and cross-fostered. Click here to view larger figure.
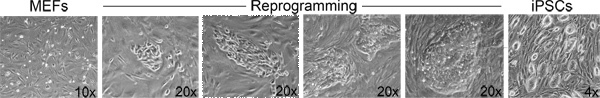 Figure 2. Morphological changes associated with reprogramming. From left to right: Examples of the morphological progression from fibroblasts to iPSC colonies during the course of a reprogramming experiment. Click here to view larger figure.
Figure 2. Morphological changes associated with reprogramming. From left to right: Examples of the morphological progression from fibroblasts to iPSC colonies during the course of a reprogramming experiment. Click here to view larger figure.
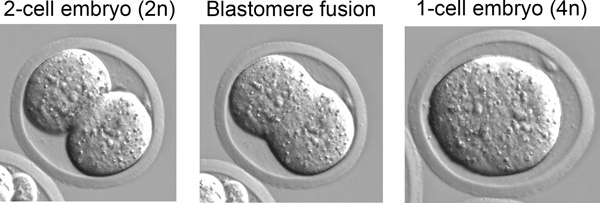 Figure 3. Production of tetraploid embryos. Diploid two-cell embryos are subjected to an electric pulse resulting in blastomere fusion and generation of one-cell tetraploid embryos.
Figure 3. Production of tetraploid embryos. Diploid two-cell embryos are subjected to an electric pulse resulting in blastomere fusion and generation of one-cell tetraploid embryos.
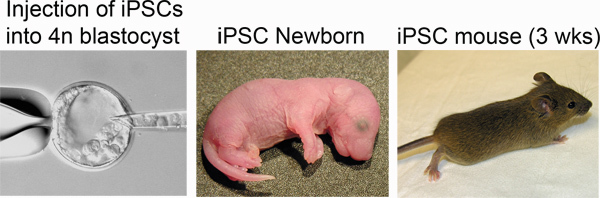 Figure 4. Production of iPSC mice. Left: iPSCs are injected into the blastocoel of a tetraploid blastocyst. Middle: Newborn iPSC mice are distinguished by pigmented eyes. Right: iPSC mouse at three weeks post-delivery.
Figure 4. Production of iPSC mice. Left: iPSCs are injected into the blastocoel of a tetraploid blastocyst. Middle: Newborn iPSC mice are distinguished by pigmented eyes. Right: iPSC mouse at three weeks post-delivery.
Discussion
Generating mice from iPSC lines using TEC assays provides a stringent functional test for the pluripotency of an iPSC line. This test may be useful to assess the relative efficacy of different reprogramming methods or to identify iPSC lines that may be most useful for generating certain cell types in vitro. Mice generated from iPSCs may be used to stringently test the long-term stability and tumorigenicity of iPSC-derived tissues. This protocol will be useful to investigators wishing to generate fully pluripotent iPSC lines or iPSC mice or to compare the relative utility of different reprogramming methods.
The mechanisms that control the generation and identification of fully pluripotent iPSCs remain poorly understood and it is possible that some iPSC lines produced using this method will not pass the TEC test. Many factors may vary between experiments including genetic backgrounds, lentiviral titer, patterns of lentiviral insertion, cell cycle parameters of the donor population, inter-laboratory differences in various steps of the TEC procedure and variable propensities of iPSCs to harbor genetic or epigenetic aberrations. To best ensure success, we take care to establish appropriate levels of lentiviral gene expression in iPSC derivation experiments by testing viral dilutions on control MEFs to ensure that each virus is sufficiently concentrated to produce detectable gene expression at least 80% and ideally 100% of the MEFs. This allows us to identify lines with multiple copies of different lentiviruses while limiting toxicity to the MEFs and producing colonies without overcrowding the wells. It should be noted that multiple other protocols have been shown to produce iPSCs with full developmental potential, using multiple methods and donor cell sources suggesting that multiple paths to full pluripotency may exist1,8-13,15. At present, however, no definitive biomarker of fully pluripotent iPSC has been identified and therefore the TEC assay remains the gold standard test of whether an iPSC line can generate all cell lineages in an organism.
Disclosures
No conflicts of interest declared.
Acknowledgments
Support to KKB, MJB, JLH and KLN was provided by the California Institute for Regenerative Medicine, the Pew Charitable Trusts Biomedical Scholars Program, the Esther B. O'Keeffe Family Foundation and the Shapiro Family Foundation. KKB is a Donald E. and Delia B. Baxter Foundation Faculty Scholar.
References
- Boland MJ, et al. Adult mice generated from induced pluripotent stem cells. Nature. 2009;461:91–94. doi: 10.1038/nature08310. [DOI] [PubMed] [Google Scholar]
- Robinton DA, Daley GQ. The promise of induced pluripotent stem cells in research and therapy. Nature. 2012;481:295–305. doi: 10.1038/nature10761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagy A, Rossant J, Nagy R, Abramow-Newerly W, Roder JC. Derivation of completely cell culture-derived mice from early-passage embryonic stem cells. Proceedings of the National Academy of Sciences of the United States of America. 1993;90:8424–8428. doi: 10.1073/pnas.90.18.8424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eggan K, et al. Hybrid vigor, fetal overgrowth, and viability of mice derived by nuclear cloning and tetraploid embryo complementation. Proc. Natl. Acad. Sci. U.S.A. 2001;98:6209–6214. doi: 10.1073/pnas.101118898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eggan K, Jaenisch R. Generation of embryonic stem (ES) cell-derived embryos and mice by tetraploid-embryo complementation. Springer; 2006. [Google Scholar]
- McLaughlin KJ. Production of tetraploid embryos by electrofusion. Methods Enzymol. 1993;225:919–930. doi: 10.1016/0076-6879(93)25058-a. [DOI] [PubMed] [Google Scholar]
- Humpherys D, et al. Epigenetic instability in ES cells and cloned mice. Science. 2001;293:95–97. doi: 10.1126/science.1061402. [DOI] [PubMed] [Google Scholar]
- Kang L, Wang J, Zhang Y, Kou Z, Gao S. iPS cells can support full-term development of tetraploid blastocyst-complemented embryos. Cell Stem Cell. 2009;5(09):135–138. doi: 10.1016/j.stem.2009.07.001. [DOI] [PubMed] [Google Scholar]
- Zhao XY, et al. iPS cells produce viable mice through tetraploid complementation. Nature. 2009;461:86–90. doi: 10.1038/nature08267. [DOI] [PubMed] [Google Scholar]
- Kang L, et al. Viable mice produced from three-factor induced pluripotent stem (iPS) cells through tetraploid complementation. Cell Res. 2011;21:546–549. doi: 10.1038/cr.2010.164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao X-Y, et al. Viable Fertile Mice Generated from Fully Pluripotent iPS Cells Derived from Adult Somatic Cells. Stem Cell Reviews and Reports. 2010;6:390–397. doi: 10.1007/s12015-010-9160-3. [DOI] [PubMed] [Google Scholar]
- Han J, et al. Tbx3 improves the germ-line competency of induced pluripotent stem cells. Nature. 2010;463:1096–1100. doi: 10.1038/nature08735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stadtfeld M, et al. Ascorbic acid prevents loss of Dlk1-Dio3 imprinting and facilitates generation of all-iPS cell mice from terminally differentiated B cells. Nat. Genet. 2012;44:398–405. doi: 10.1038/ng.1110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stadtfeld M, et al. Aberrant silencing of imprinted genes on chromosome 12qF1 in mouse induced pluripotent stem cells. Nature. 2010;465:175–181. doi: 10.1038/nature09017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carey BW, et al. Reprogramming factor stoichiometry influences the epigenetic state and biological properties of induced pluripotent stem cells. Cell Stem Cell. 2011;9:588–598. doi: 10.1016/j.stem.2011.11.003. [DOI] [PubMed] [Google Scholar]
- Go WY, Ho SN. Optimization and direct comparison of the dimerizer and reverse tet transcriptional control systems. The Journal of Gene Medicine. 2002;4:258–270. doi: 10.1002/jgm.271. [DOI] [PubMed] [Google Scholar]
- Huangfu D, et al. Induction of pluripotent stem cells by defined factors is greatly improved by small-molecule compounds. Nature. 2008;26:795–797. doi: 10.1038/nbt1418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li X-y, et al. Passage number affects the pluripotency of mouse embryonic stem cells as judged by tetraploid embryo aggregation. Cell and Tissue Research. 2007;327:607–614. doi: 10.1007/s00441-006-0354-6. [DOI] [PubMed] [Google Scholar]
- George SHL, et al. Developmental and adult phenotyping directly from mutant embryonic stem cells. Proceedings of the National Academy of Sciences. 2007;104:4455–4460. doi: 10.1073/pnas.0609277104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagy A, Gertsenstein M, Vintersten K, Behringer R. Manipulating the mouse embryo: A laboratory manual. Cold Spring Harbor Laboratory Press; 2003. [Google Scholar]
- Kirak O, et al. Transnuclear Mice with Pre-defined T Cell Receptor Specificities Against Toxoplasma gondii Obtained Via SCNT. J. Vis. Exp. 2010. p. e2168. [DOI] [PMC free article] [PubMed]