

Abstract
The opportunistic pathogen Pseudomonas aeruginosa utilizes type III secretion system (T3SS) to translocate effector proteins into eukaryotic host cells that subvert normal host cell functions to the benefit of the pathogen, and results in serious infections. T3SS in P. aeruginosa is controlled by a complex system of regulatory mechanisms and signaling pathways. In this study, we described that Crc, an RNA-binding protein, exerts a positive impact on T3SS in P. aeruginosa, as evidenced by promoter activity assays of several key T3SS genes, transcriptomics, RT-PCR, and immunoblotting in crc mutant. We further demonstrated that the regulatory function of Crc on the T3SS was mediated through the T3SS master regulator ExsA and linked to the Cbr/Crc signaling system. Expression profiling of the crc mutant revealed a downregulation of flagship T3SS genes as well as 16 other genes known to regulate T3SS gene expression in P. aeruginosa. On the basis of these data, we proposed that Crc may exert multifaceted control on the T3SS through various pathways, which may serve to fine-tune this virulence mechanism in response to environmental changes and nutrient sources.
Keywords: Crc, exsA, Pseudomonas aeruginosa, type III secretion system
Introduction
Pseudomonas aeruginosa is a ubiquitous environmental bacterium capable of causing serious and often life-threatening infections in cystic fibrosis patients and immunocompromised humans. This opportunistic pathogen is equipped with a large number of virulence mechanisms and one of the major virulence factors is the type III secretion system (T3SS), which facilitates the delivery/translocation of bacterial effector proteins into eukaryotic host cells to inhibit phagocytosis, resulting in cytotoxicity and tissue destructions in the host (Ghosh 2004; Sato and Frank 2004; Journet et al. 2005; Yahr and Wolfgang 2006).
Regulation of T3SS in P. aeruginosa is complicated and it involves both genetic and environmental factors. There are about 43 coordinately regulated genes encoding the type III secretion and translocation machinery, regulatory functions, type III effectors, and effector-specific chaperones (Frank 1997). Expression of the T3SS genes is tightly regulated and under the direct transcriptional control of the master regulator ExsA, a member of the AraC family of transcriptional activators (Yahr and Frank 1994; Hovey and Frank 1995). The activity of ExsA is controlled by interaction with the anti-activator protein ExsD (McCaw et al. 2002), and the ExsD–ExsA interaction can be disrupted by tight binding between the anti-anti-activator ExsC and ExsD (Dasgupta et al. 2004). ExsC activity is in turn controlled by a secreted protein ExsE (Rietsch et al. 2005; Urbanowski et al. 2005). In addition to these T3SS-specific regulatory proteins, other regulators and proteins are also known to affect the expression of T3SS genes. They include, nonexhaustively, the membrane-associated adenylcyclase CyaB; cyclic AMP (cAMP)-binding protein Vfr (Wolfgang et al. 2003); stationary-phase sigma factor RpoS (Hogardt et al. 2004); alginate biosynthesis proteins MucA/AlgU/AlgR (Wu et al. 2004); two-component systems RtsM/RetS and GacA/GacS; RNA-binding protein RsmA (Goodman et al. 2004; Laskowski et al. 2004; Zolfaghar et al. 2005); small proteins PtrB and PtrC (Wu and Jin 2005; Jin et al. 2011); type IV pili biogenesis protein FimL (Whitchurch et al. 2005); transcriptional activator PsrA (Shen et al. 2006); tryptophan synthase TrpAB and tryptophan dioxygenase KynA (Shen et al. 2008); nitrite reductase NirS (van Alst et al. 2009); magnesium transporter MgtE (Anderson et al. 2010); transmembrane protein FlhA (Bange et al. 2010) and cAMP phosphodiesterase CpdA (Fuchs et al. 2010). However, it is not clear how these proteins exert their influence on T3SS.
Expression of P. aeruginosa T3SS is also influenced by various environmental conditions, such as calcium depletion and direct contact with host cells or serum (Yahr et al. 1995; Frank 1997). Quorum-sensing signals N-butyryl-homoserine lactone (BHL) and Pseudomonas quinolone signal (PQS) as well as other small molecules, such as acetyl-CoA, histidine, and spermidine have also been reported to influence the expression of T3SS genes (Wolfgang et al. 2003; Rietsch et al. 2004; Bleves et al. 2005; Rietsch and Mekalanos 2006; Zhou et al. 2007; Singh et al. 2010). Osmotic stress, metabolic signals, and specific metabolites such as tryptophan catabolites were found to have a profound influence on the expression of T3SS genes, although the molecular regulatory mechanisms linking these metabolite/stresses remains obscure (Aspedon et al. 2006; Shen et al. 2008). It is plausible that P. aeruginosa has evolved several complicated regulatory mechanisms and signaling pathways to fine-tune the regulation of T3SS in response to environmental changes.
Crc is a catabolite repression control protein involved in carbon regulation of several pathways in the Pseudomonas species (MacGregor et al. 1991, 1996; Morales et al. 2004). Recently, Crc was shown to have RNA-binding properties and it is capable of binding to the A-rich region of target mRNAs to inhibit their translation initiation (Moreno et al. 2007, 2009, 2010; Sonnleitner et al. 2009). In P. aeruginosa, the activity of Crc is modulated by a small RNA CrcZ, which is able to bind to and sequester the Crc protein with high affinity and thus relieve repression of Crc targets. The expression of CrcZ is controlled by CbrA/CbrB two-component system in response to different carbon sources (Sonnleitner et al. 2009). In addition to catabolite repression, Crc was found to be central in the regulation of virulence and virulence-related processes, and these include cell motility, biofilm formation, and antibiotic resistance. Inactivation of the crc gene impaired swimming, swarming, and twitching motilities, and increased exopolysaccharides production and biofilm formation in P. aeruginosa (O'Toole et al. 2000; Linares et al. 2010). The crc mutant also exhibited reduced virulence to Dictyostelium discoideum and reduced expression of quorum sensing-regulated virulence determinants, such as cyanide, pyoverdine, and elastase, but increased pyocyanin production (Linares et al. 2010; Yeung et al. 2011). Proteomic analysis revealed that three proteins, NirS, GltA, and AceE, which were reported to be involved in T3SS modulation, were differentially expressed in the crc mutant and this correlated with a downregulation in the protein levels of the four T3SS secretion proteins, PscN, PopD, PcrV, and ExoS from the crc mutant culture supernatants (Linares et al. 2010). Real-time quantitative polymerase chain reaction (RT-qPCR) on cbrA, cbrB, crcZ, and crc mutant revealed altered expression of several T3SS genes during P. aeruginosa infection of HBE cell line (Yeung et al. 2011). However, it remains unknown how Crc modulates T3SS. Here, we present a detailed analysis and systematic study of Crc on its influence on the T3SS and propose that Crc modulates T3SS through multiple regulatory pathways.
Materials and Methods
Bacterial strains, media, and growth conditions
Pseudomonas aeruginosa strains used in this study are derived from the prototrophic laboratory strain PAO1. These strains and other bacteria used in this study are listed in Table 1. Bacteria were routinely grown at 37°C in Luria–Bertani broth (LB) or LB medium supplemented with the chelating reagent nitrilotiracetic acid (NTA) at a final concentration of 5.0 mmol/L for induction of T3SS. Mins (Nicas and Iglewski 1984) and BSM (Sonnleitner et al. 2009) minimal medium was also used. Three different carbon sources, mannitol, glucose, and succinate were tested in BSM minimal medium at final concentration of 40 mmol/L and with/without addition of 1 mmol/L spermidine, which induces the transcription of T3SS genes. Antibiotics were used when necessary at the following concentrations: carbenicillin, 300 μg/mL for P. aeruginosa, 200 μg/mL for Escherichia coli; tetracycline, 100 μg/mL for P. aeruginosa and 10 μg/mL for E. coli; and gentamicin, 50 μg/mL for P. aeruginosa and 5 μg/mL for E. coli.
Table 1.
Bacterial strains and plasmids used in this study1
| Strain or plasmid | Relevant genotype or phenotype | Source or reference |
|---|---|---|
| Pseudomonas aeruginosa | ||
| PAO1 | Prototrophic laboratory strain | Laboratory collection |
| PAO1pClacZ | PAO1 with chromosome integration of exsCEBA promoter fused with lacZ | Zhou et al. (2007) |
| p39 | Transposon mutant with disruption of crc gene in PAO1pClacZ | This study |
| P39(crc) | P39 containing pUCP-crc | This study |
| Δcrc | PAO1 with in-frame deletion of crc | This study |
| Δcrc(crc) | Δcrc containing pUCP-crc | This study |
| ΔcrcpClacZ | PAO1pClacZ with in-frame deletion of crc | This study |
| ΔcrcpClacZ(crc) | ΔcrcpClacZ containing pUCP-crc | This study |
| ΔcbrBpClacZ | PAO1pClacZ with in-frame deletion of cbrB | This study |
| ΔCrcZpClacZ | PAO1pClacZ with deletion of CrcZ | This study |
| exsA::Tn | Transposon mutant with disruption of exsA gene in PAO1pClacZ | This study |
| PAO1pTlacZ | PAO1 with chromosome integration of exoT promoter fused with lacZ | Zhou et al. (2007) |
| PAO1pDlacZ | PAO1 containing a vector of pME-PexsD–lacZ | This study |
| PAO1pD’-‘lacZ | PAO1 containing a vector of pME-PexsD’-‘lacZ | This study |
| PAO1pN’-‘lacZ | PAO1 containing a vector of pME-PpopN’-‘lacZ | This study |
| Escherichia coli | ||
| DH5α | F−ϕ80dlacZΔM15 endA1hsdR17 (rk− mk−. supE44 thi-1 gyrA96 Δ(lacZYA-argF) | Gibco |
| S17-1 | recA pro (RP4-2Tet::Mu Kan::Tn7) | Simon et al. (1983) |
| Plasmid | ||
| pBT20 | Mariner transposon vector; Gmr, Apr | Kulasekara et al. (2005) |
| pUCP19 | E. coli – P. aeruginosa shuttle vector with the lac promoter (Plac), Ampr/Cbr | ATCC 87110 |
| pUCP-crc | pUCP19 containing crc under the control of Plac | This study |
| pUCP-exsA | pUCP19 containing exsA under the control of Plac | This study |
| pEX18Gm | P. aeruginosa gene replacement vector; sacB, Gmr/Cbr | Hoang et al. (1998) |
| pEX-de crc | pEX18Gm carrying the crc flanking region with the gene being deleted | This study |
| pEX-de cbrB | pEX18Gm carrying the cbrB flanking region with the gene being deleted | This study |
| pEX-de CrcZ | pEX18Gm carrying the CrcZ flanking region with the gene being deleted | This study |
| pME6010 | pVS1-p15A shuttle vectors | Heeb et al. (2000) |
| pME2-lacZ | pME6010 carrying a full-length lacZ | Dong et al. (2008) |
| pME-PexsDlacZ | pME2-lacZ carrying exsD promoter in front of lacZ | This study |
| pME-PexsD’-‘lacZ | PexsD’-‘lacZ translational fusion | This study |
| pME-PpopN’-‘lacZ | PpopN’-‘lacZ translational fusion | This study |
Ampr, ampicillin resistant; Cbr, carbenicillin resistant; Gmr, gentamicin resistant.
Plasmid and mutant construction
The plasmids used in this study are listed in Table 1. The Mariner transposon carried by plasmid pBT20 (Kulasekara et al. 2005) was used to generate the transposon mutants and the mutants were screened for genes involved in the regulation of exsCEBA expression as described previously (Zhou et al. 2007). The genes disrupted by transposon insertion were identified by PCR (Caetano-Annoles 1993). All deletion mutants of P. aeruginosa were produced by using pEX18Gm plasmid (Hoang et al. 1998) containing the PCR fragment flanking each gene for in-frame deletion of the internal sequence following the methods described previously (Dong et al. 2008). The deletion mutants were confirmed by PCR.
For complementation of the mutant and in trans expression, the coding regions of the native gene were amplified from PAO1 genomic DNA and the PCR products were cloned at HindIII and BamH1 sites in the shuttle vector pUCP19 (ATCC 87110). To construct the PAO1pDlacZ reporter, a 568-bp fragment spanning −409 to +159 bp relative to the translational start site of the exsD gene was amplified from the genomic DNA of P. aeruginosa strain PAO1 by PCR. This fragment contains the engineered HindIII and EcoRI sites. The HindIII- and EcoRI-digested fragments were then ligated into the corresponding sites of pME2-lacZ (Dong et al. 2008). The PexsD’-‘lacZ and PpopN’-‘lacZ translational fusion reporter plasmids were constructed by using the fragments (−409 to +159 for exsD and −243 to +120 for popN) amplified from PAO1 genomic DNA and ligated into pME'lacZ translational fusion vector (Heeb et al. 2000). These constructs were introduced into P. aeruginosa by electroporation, and the transformants were selected on LB agar plates containing relevant antibiotics.
Immunoblotting detection
Whole-cell lysate samples were prepared from different bacterial strains grown in LB broth supplemented with NTA to an absorbance (A600) of ≍1.3–1.5. After sonication, samples were analyzed by 12% SDS–PAGE followed by either InstantBlue (Expedeon, Cambridgeshire, U.K.) staining or transferred to nitrocellulose membranes for detection by immunoblotting with antibodies against ExsA, ExsD (generously provided by Dr. Sylvie Elsen, iRTSV, France), E. coli β-galactosidase, and E. coli RNA polymerase (RNAP), respectively, as indicated.
RNA extraction, microarray, RT-PCR, and real-time PCR analysis
Overnight cultures of P. aeruginosa strains were diluted in Mins medium and incubated at 37°C until the OD600 reached about 1.5. Total RNA samples were then isolated using the RNeasy miniprep kit (Qiagen). Following DNase I (Promega) digestion, RNA samples were quantified and analyzed for purity by agarose gel electrophoresis and by PCR to make sure that no contaminating genomic DNA was present. cDNA was synthesized by using random primers (Invitrogen), SuperScript III (Invitrogen) and biotin-ddUTP according to the protocol from Affymetrix (Santa Clara, CA). Target hybridization, washing, and staining were performed following the manufacturer's instructions on an Affymetrix GeneChip fluidics station. GeneChip arrays were scanned using an Affymetrix probe array scanner. The data were analyzed by using Affymetrix Expression Console Software and the RMA algorithm was used for comparing gene expression between mutant and wild-type strain. Two independent experiments were performed and the results were showed as fold change. Reverse transcription PCR (RT-PCR) was performed using OneStep RT-PCR Kit (Qiagen) and RT-qPCR analysis was carried out by using QuantiTect SYBR Green PCR kit (Qiagen) on the LightCycler® 4.0 system (Roche) according to the manufacturer's instruction.
Quantitative β-galactosidase assay
For measurement of β-galactosidase activity, P. aeruginosa lacZ reporter strains were grown in different media as indicated with shaking at 37°C until reaching an OD600 of 1.0–1.5. Bacterial cells were collected and measured for β-galactosidase activity. Results were given as Miller units of β-galactosidase activity per OD600.
Cytotoxicity assay
The cytotoxicity of P. aeruginosa strain PAO1 and its crc deletion mutant were measured using the Cell Counting Kit-8 (Dojindo Molecular Technologies, Rockville, MD), which is based on the dehydrogenase activity detection in viable cells. Bacterial strains were grown in LB medium with 5.0 mmol/L NTA till an OD600 of 1.0. The bacterial cells were washed and diluted in Dulbecco's Modified Eagle Medium and then inoculated into the monolayer of HeLa or A549 cells in a 96-well plate at a multiplicity of infection of 50. Absorbance at 450 nm was measured and the cell viability was calculated according to the manufacturer's protocol.
Results
Mutation of the carbon metabolism regulator Crc resulted in reduced transcription of exsCEBA
To identify the genes that regulate the expression of T3SS in P. aeruginosa, the promoter of exsCEBA operon was fused to the reporter gene lacZ and the construct was integrated into the chromosome of the strain PAO1 to generate the reporter strain PAO1pClacZ as described previously (Zhou et al. 2007). A transposon mutant library of strain PAO1pClacZ was generated by random insertional mutagenesis using Mariner transposon, and the library was screened for genes implicated in the regulation of T3SS in P. aeruginosa under T3SS-inducing condition (by addition of 5 mmol/L calcium-depleted chelating regent – NTA to LB medium). One mutant, p39, was found to have dramatically reduced exsCEBA promoter-directed β-galactosidase activity (Fig. 1A). PCR and DNA sequencing analysis revealed that p39 contains a transposon insertion in the coding region of crc gene. To confirm the role of Crc in the modulation of exsCEBA expression, the wild-type crc gene was amplified by PCR from the genomic DNA of P. aeruginosa PAO1 and the fragment was cloned into a multicopy vector pUCP19 under the control of the lac promoter. The construct was introduced into the mutant p39 and the β-galactosidase activity assay showed that expression of exsCEBA was restored in the complemented strain (Fig. 1A). For further validation, we generated the crc in-frame deletion mutant using P. aeruginosa PAO1pClacZ as the parental strain. The assay results confirmed that deletion of crc reduced the expression of exsCEBA and in trans expression of the wild-type crc allele in Δcrc mutant restored the expression of exsCEBA to the level of its parental strain PAO1pClacZ (Fig. 1A, right). The β-galactosidase activity assay results were further supported by Western blot analysis on the abundance of the β-galactosidase protein (Fig. 1B). As Mins minimal medium was reported to be capable of maximizing the induction of T3SS (Nicas and Iglewski 1984), it is interesting to determine whether disruption of crc could still hamper exsCEBA expression in a strongly T3SS-inducing condition. The results showed the same pattern as observed in the LB + NTA medium (Fig. 1C). The findings indicate that Crc is critical for the expression of exsCEBA operon under various growth conditions. Taken together, the above results indicate that Crc positively regulates the expression of exsCEBA.
Figure 1.
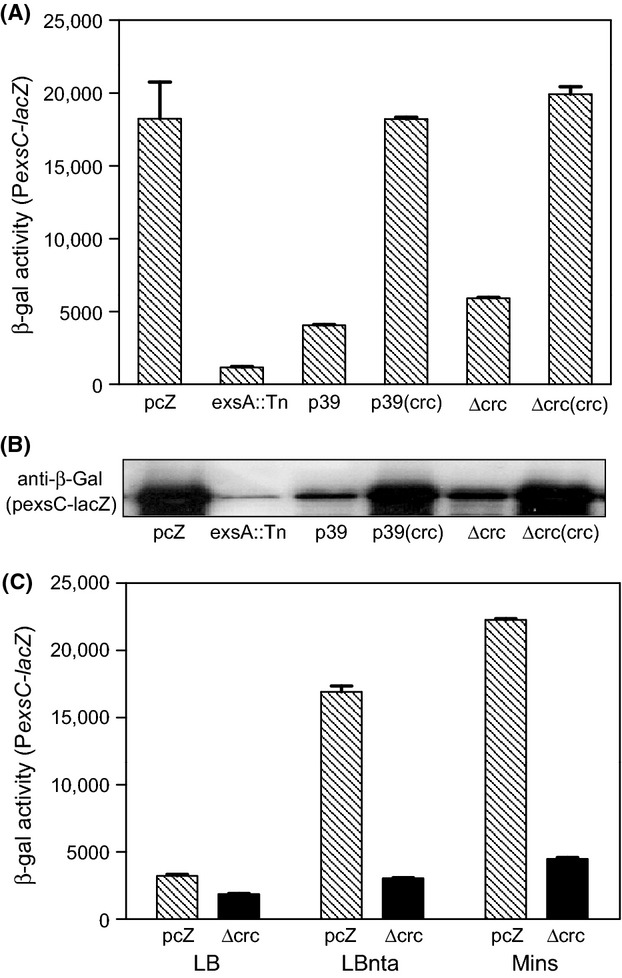
Mutation in the carbon metabolism regulator, Crc, reduced the expression of exsCEBA. (A) Transcriptional expression of PexsC–lacZ in crc transposon mutant (p39) and deletion mutant (Δcrc) as well as their corresponding crc-complemented strains under type III secretion system-inducing condition. The parental strain PAO1pClacZ (pcZ), which carried a single copy of PexsC–lacZ transcriptional reporter gene on its chromosome and its exsA transposon mutant (exsA::Tn) were used as positive and negative controls, respectively. (B) Immunoblotting detection of exsCEBA promoter-directed β-galactosidase. Total proteins were prepared from strain PAO1pClacZ (pcZ) and its derivatives. The expression levels were assayed by immunoblotting using antibodies against β-galactosidase. (C) Expression of PexsC–lacZ in crc deletion mutant under different growth conditions as indicated. Bacterial cultures were grown in Luria–Bertani (LB) medium, LB with 5.0 mmol/L nitrilotiracetic acid (LBnta), or Mins minimal medium. The data were the means of three replicates and error bar represents standard deviations.
Disruption of crc impaired the expression of exoT, exsD, and popN and attenuated T3SS-dependent cytotoxicity
In addition to analyzing the expression of the master regulator exsCEBA operon in Δcrc mutant, we also tested the impact of Crc on expression of other T3SS genes. We first checked the transcription of exsD and exoT by measuring their promoter-directed β-galactosidase activity. exsD encodes an anti-activator of ExsA whose expression is positively regulated by ExsA (McCaw et al. 2002), and ExoT is a bifunctional type III cytotoxin exoenzyme T. The assay results showed that their expression was reduced drastically in the Δcrc mutant compared with the parental strain, either cultured in LB-NTA (Fig. 2A and B) or Mins medium (data not shown). Next, we tested the translational expression of exsD and popN by in-frame gene fusions to a truncated lacZ reporter gene (Heeb et al. 2000). As both proteins of ExsD and PopN are known for their negative regulatory role on T3SS, we wondered whether Crc exerts its influence on T3SS by inhibiting their translation. The β-galactosidase activity assay results showed that the translational expression of both genes was downregulated in the Δcrc mutant (Fig. 2C and D).
Figure 2.
Reduced expression of exoT, exsD, and popN in crc deletion mutant. (A) and (B) Transcriptional expression of exsD (A) and exoT (B) in wild-type PAO1 and Δcrc mutant carrying a vector with PexsD–lacZ or PexoT–lacZ transcriptional reporter gene. (C) and (D) The translational expression of exsD (C) and popN (D) in wild-type PAO1 and Δcrc mutant carrying a vector with PexsD’-‘lacZ or PpopN’-‘lacZ translational fusion reporter genes. Bacteria were grown at 37°C in Luria–Bertani medium containing 5.0 mmol/L nitrilotiracetic acid. (E) Reduced cytotoxicity in crc mutant in comparison with wild-type PAO1. Cytotoxicity was assayed using Cell Counting Kit-8 (Dojindo Molecular Technologies), which is based on the dehydrogenase activity detection in viable cells. HeLa and A549 cells were infected with bacterial cells at multiplicity of infection of 50. Absorbance at 450 nm was measured and the cell viability was calculated. The data were the means of four (A–D) or five (E) replicates and error bar represents standard deviations.
To test T3SS-dependent cytotoxicity toward mammalian cells, wild-type PAO1 and Δcrc mutant were used to infect the human epithelial cell lines Hela and A549. After 4-h infection, the cells were measured for dehydrogenase activity. As shown in Figure 2E, the cell viability in the sample infected with Δcrc strain was higher than that challenged with the wild-type PAO1, indicating that the Δcrc mutant was less cytotoxic. This result is agreeable with Linares's result that Crc reduces virulence in Dictyostelium (Linares et al. 2010).
Crc modulates T3SS by influencing the expression level of ExsA
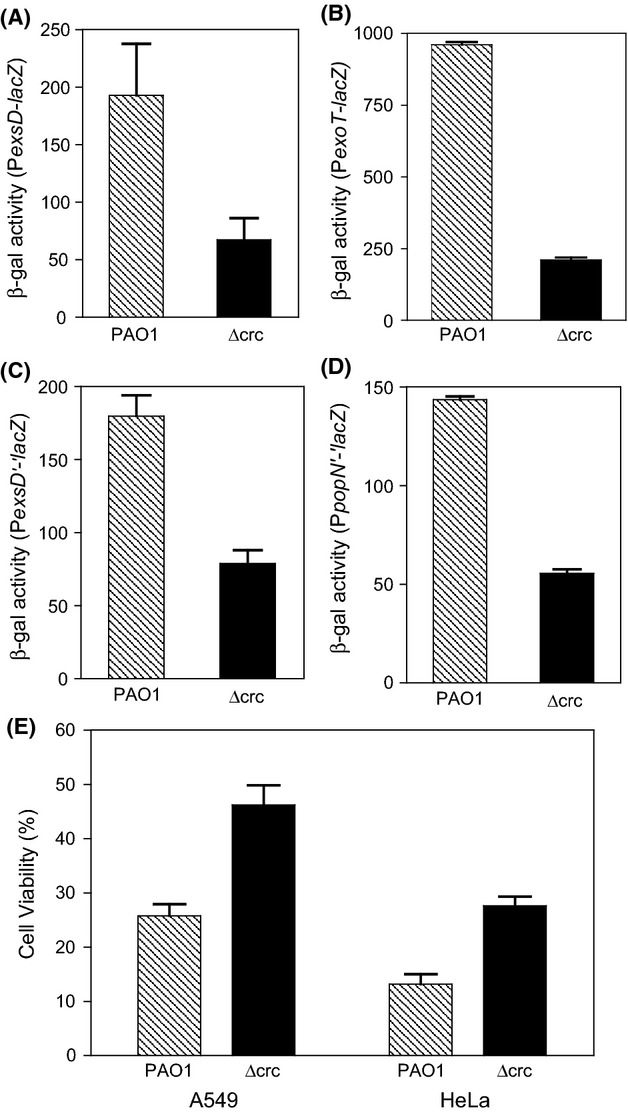
As ExsA is a master regulator of T3SS, we tested whether the positive regulation of Crc on T3SS was exerted through ExsA. Immunoblotting using antibody against ExsA yielded much weaker signals from both of crc transposon and crc deletion mutants compared with the parental strain; whereas the crc-complemented strains showed stronger signals (Fig. 3A, anti-ExsA), indicating that the influence of Crc on T3SS was mediated by ExsA. Similarly, immunoblots of ExsD presented the same patterns (Fig. 3A, anti-ExsD), which correlates with previous observation that the expression of ExsD is under the control of the master regulator ExsA (McCaw et al. 2002).
Figure 3.
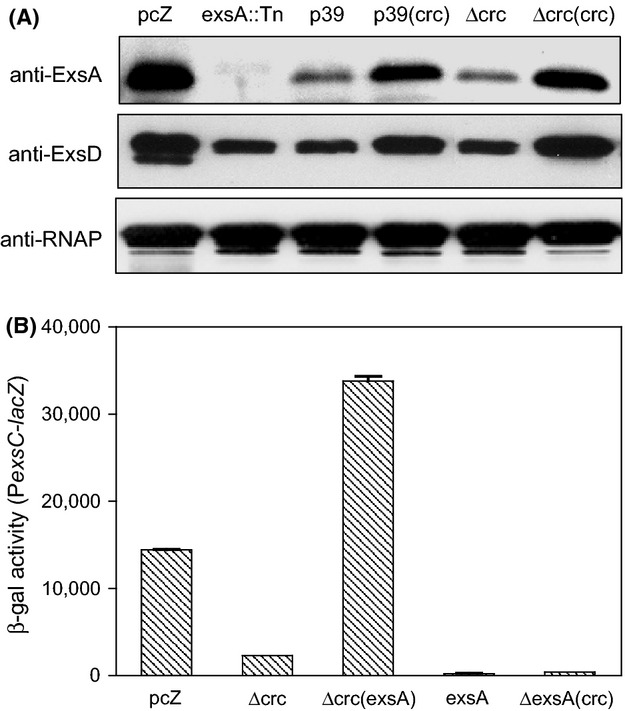
Influence of Crc on type III secretion system (T3SS) dependent on ExsA expression. (A) Immunoblotting detection of the expression level of ExsA and ExsD. Total proteins were prepared from strain PAO1pClacZ (pcZ) and its derivatives. The expression levels were assayed by immunoblotting using antibodies against ExsA, and ExsD as well as RNA polymerase, which serves as internal control. (B) In trans expression of exsA restored the T3SS gene expression in Δcrc mutant, but overexpression of crc in ΔexsA mutant failed to induce the expression of T3SS genes. In these in trans expression experiments, the empty vector pUCP19 was introduced into corresponding control strains for comparison analysis. Bacterial strains were grown under T3SS-inducing condition (Luria–Bertani + 5 mmol/L nitrilotiracetic acid).
To determine whether Crc regulates T3SS solely through ExsA, we tested the T3SS expression patterns by in trans expression of exsA in Δcrc mutant and by introducing wild-type crc into ΔexsA mutant, respectively. β-Galactosidase activity assay showed that overexpression of exsA in Δcrc mutant resulted in enhanced expression of exsCEBA, whereas overexpression of crc in ΔexsA had no effect on T3SS gene expression (Fig. 3B). Cumulatively, these results suggest that the influence of Crc on T3SS is through its direct or indirect regulatory role on the transcriptional expression of exsA.
Effect of different carbon sources and Cbr/Crc signaling system on the regulation of T3SS
Crc and CbrA/CbrB two-component system have been demonstrated to play a role in catabolite repression and enabling Pseudomonas spp. to utilize various carbon sources (Nishijyo et al. 2001; Rojo 2010). To test whether different carbon sources could have a possible effect on T3SS in P. aeruginosa, three different carbon sources, that is, succinate, glucose, and mannitol were used and the expression of exsCEBA was measured. Among them, succinate is a preferred substrate, which can cause strong catabolite repression in Pseudomonas, glucose causes mild catabolite repression, whereas mannitol is unable to cause catabolite repression (Sonnleitner et al. 2009). These three carbon sources were each supplemented in BSM minimal medium, at a final concentration of 40 mmol/L with or without addition of 1 mmol/L spermidine, which is a known host signal that induces the transcription of T3SS genes in P. aeruginosa (Zhou et al. 2007). Δcrc mutant grew poorly in mannitol-supplemented medium, but addition of spermidine improved its growth (data not shown). Importantly, β-galactosidase activity assay of wild-type strain showed that maximal expression of exsCEBA was detected when succinate was used as carbon source, which seems to suggest that catabolite repression promotes T3SS expression. This notion was reinforced by the findings that glucose was better than mannitol in promoting the expression of exsCEBA. As expected, the influence of carbon source on T3SS expression was abolished when crc was deleted (Fig. 4A). Interestingly, we found that addition of spermidine partially restored the T3SS response to catabolite repression-dependent induction in the Δcrc mutant background (Fig. 4A). As the function of Crc is under the control of CbrA/CbrB two-component system and small RNA CrcZ, we next tested their effects on the expression of exsCEBA. Under T3SS-inducing conditions, the cbrB deletion mutant showed only a minor reduction in expression of exsCEBA, while the crcZ deletion mutant displayed an increased expression in comparison with their parental strain (Fig. 4B). These results indicate that the Cbr/Crc signaling system is involved in the regulation of T3SS in addition to its role in catabolite repression in P. aeruginosa.
Figure 4.
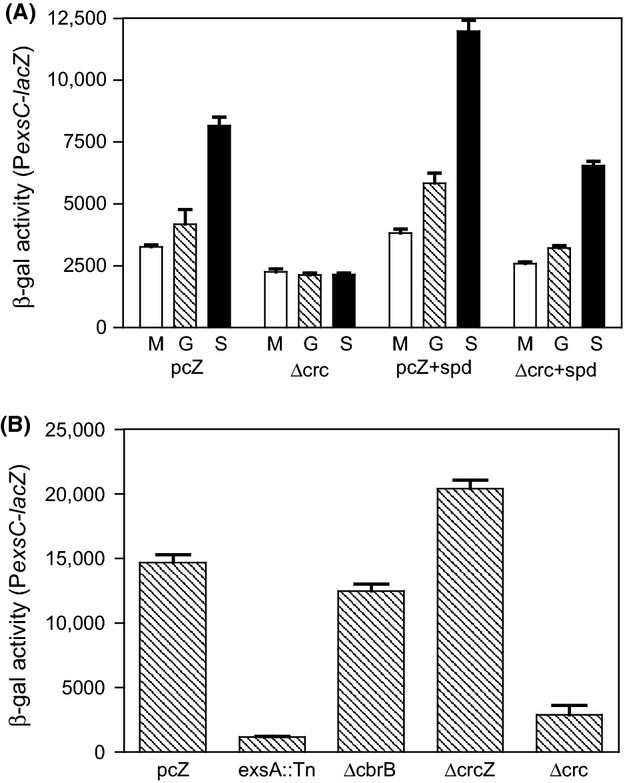
Effect of different carbon sources and Cbr/Crc signaling system on type III secretion system (T3SS) gene expression. (A) The transcriptional expression of PexsC–lacZ in response to different carbon sources with/without external addition of T3SS-inducer spermidine. Crc deletion mutant (Δcrc) and its parental strain PAO1pClacZ (pcZ) were grown in BSM minimal medium supplemented with either 40 mmol/L mannitol (M), 40 mmol/L glucose (G), or 40 mmol/L succinate (S) with/without 1 mmol/L spermidine (spd). (B) Effect of Cbr/Crc signaling system on the expression of exsCEBA. Bacterial strains were grown in Luria–Bertani medium with 5.0 mmol/L nitrilotiracetic acid at 37°C and their parental strain PAO1pclacZ (pcZ) and exsA mutant were used as positive and negative controls, respectively. The data were the means of three (A) or six (B) replicates and error bar represents standard deviations.
Transcriptome analysis revealed the mutation in crc downregulated the transcription of T3SS and T3SS-related genes
To investigate the possible regulatory mechanisms of Crc on T3SS, global gene expression profiles of crc mutant and its parental strain PAO1pClacZ were examined by using the P. aeruginosa whole-genome microarray (Affymetrix) under calcium-depleted conditions. The results showed that a total of 425 genes were differentially regulated by at least twofold in mRNA expression by Crc. Among them, 208 genes were downregulated (Table S1) and 217 were upregulated in the Δcrc mutant (Table S2). Agreeable with the genetic analysis data, the transcriptional expression levels of 26 T3SS genes, including exsCEBA, exsDpscBCDEFGHIJK, pcrVHpopBD, exoT, and exoS etc., were decreased more than 1.5-fold in the Δcrc mutant (Fig. 5A). RT-PCR analysis confirmed the reduced expression of exsC, exsD, pscF, popB, and exoT in Δcrc mutant (Fig. S1).
Figure 5.
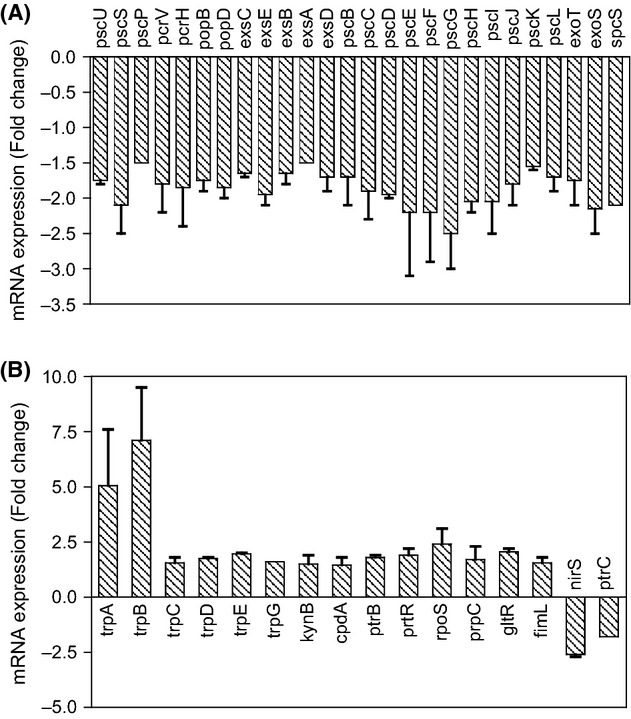
Expression of type III secretion system (T3SS) and T3SS-related genes in Δcrc mutant by transcriptome analysis. (A) Decreased transcripts of 26 T3SS genes in Δcrc mutant in comparison with its parental strain PAO1pClacZ. (B) Expression of T3SS-related genes in Δcrc mutant. Bacteria were cultured in Mins medium at 37°C and the cells were collected at OD600 ≍ 1.5. The data are shown as the transcriptional fold changes (Δcrc vs. PAO1pClacZ). The microarray was repeated twice and the mean data were presented.
In addition, several genes associated with the regulation of T3SS (Diaz et al. 2011) were also found to be influenced by Crc (Fig. 5B). These genes include the trp operon trpABCBEG, as well as kynB, cpdA, ptrB, ptrR, prpC, rpoS, gltR, fimL, nirS, and ptrC (see Discussion section for the description of these genes). Apart from nirS and ptrC, which were downregulated in crc mutant, other genes were upregulated in the mutant (≥1.5-fold change). Furthermore, mgtE and aceE were found also to be moderately upregulated by 1.4- and 1.3-fold, respectively. To confirm the expression patterns of these genes in Δcrc mutant and its parental strain, RT-PCR and real-time PCR were performed. The results confirmed that their expressions were influenced by Crc (Fig. 6).
Figure 6.
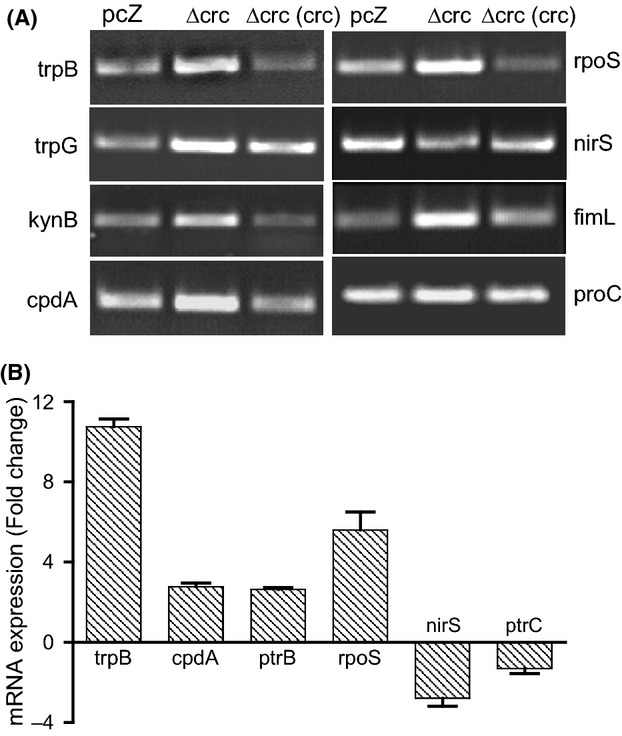
Transcriptional analysis by using real time polymerase chain reaction (RT-PCR) and real-time quantitative polymerase chain reaction (RT-qPCR) for selected type III secretion system (T3SS)-related genes. (A) RT-PCR analysis of the selected T3SS-related genes in Δcrc mutant, crc-complemented strain Δcrc(crc) and parental strain PAO1pclacZ (pcZ). The gene proC was used as internal control. (B) RT-qPCR analysis of several selected genes on their relative expression in Δcrc mutant in comparison with its parental strain PAO1pclacZ. Their expression levels were normalized against the reference gene proC and the data presented were the average of three experiments.
Discussion
Crc has been known to repress the expression of genes involved in assimilation of sugars, nitrogenated compounds, aromatic compounds, and hydrocarbons and in the transport of catabolites when preferred substrates, such as succinate, are present (Wolff et al. 1991; MacGregor et al. 1996; Hester et al. 2000). Here we showed that Crc positively regulated the expression of many T3SS genes in P. aeruginosa under T3SS-inducing condition. Detailed genetic and transcriptomic analyses indicate that Crc may exert a multifaceted effect on T3SS through various pathways to respond to different environmental signals. Our results provide further information to establish the links and connections between Cbr/Crc catabolite repression signaling system and T3SS regulatory network. It is possible that relief of catabolite repression by inactivation of Crc might have changed the intracellular level of some catabolites and such changes may repress the expression of T3SS genes. In fact, some catabolic pathways, such as histidine utilization, were reported to be severely depressed by catabolite repression when succinate was added in the medium (Potts and Clarke 1976), and the reduced level of histidine due to overexpression of genes involved in histidine transport and metabolism prevents expression of T3SS genes (Rietsch et al. 2004). In this study, we found that the tryptophan catabolic pathway is depressed by Crc-mediated catabolite repression (Figs. 5B, 6) and tryptophan metabolites, such as indole-3-acetic acid, kynurenine and PQS were reported to inhibit T3SS expression (Shen et al. 2008; Singh et al. 2010). These findings strongly suggest that catabolite repression may be implicated in modulation of T3SS in P. aeruginosa.
Crc is an RNA-binding protein, which was known to bind to an A-rich region located close to the ribosomal-binding site of target mRNAs to inhibit translation initiation (Moreno et al. 2007, 2009, 2010; Sonnleitner et al. 2009). On the basis of the above information and combining a previous bioinformatic study, which predicted 215 candidate genes containing A-rich motifs (AANAANAA) on their 5′ end of mRNA (Browne et al. 2010), we tried to locate key T3SS genes with the A-rich motif within or close to their ribosomal binding site. However, we could not find any. It seemed unlikely that the regulation of Crc on T3SS is through direct binding to the ribosomal-binding site of the key T3SS genes. However, when the same search was conducted on the Crc regulon, 56 genes containing the A-rich motif in their promoter region were identified (Table S3), but none of them belongs to T3SS regulon. These data may provide further information of the mechanism of regulation of other genes by Crc in P. aeruginosa.
Although Crc is known to inhibit target mRNA translation, only a few targets have been experimentally demonstrated, and these include benR, alkS, xylR, and xylB from P. putida and amiE from P. aeruginosa (Moreno et al. 2007, 2009, 2010; Sonnleitner et al. 2009). Our transcriptome profiles indicated that Crc indeed serves as a global regulator in controlling the expression of many genes, including that of the type III secretion system. Crc may influence on gene expression indirectly, but we still could not rigorously eliminate the possibility that Crc may also have a direct role in transcriptional regulation through its influence on mRNA stability. The fact that the crc regulon makes up to nearly 10% of the genome suggests that the mode of regulation by Crc is pleiotropic rather than specific. Comparing our transcriptomic data with Linares's proteomic data (both with fold change threshold set at 1.5), we found only 13 genes from our transcriptomic data in common with that of the proteomics data, and the latter has reported 66 differentially expressed proteins in the crc mutant (Linares et al. 2010). The same trend was observed in P. putida, where only an overlap of 23 genes were identified to be differentially transcribed in the transcriptomic assay of the total of 64 proteins controlled by Crc as shown by proteomics methods (Moreno et al. 2009). The difference between transcriptomic and proteomic analysis could reflect the different culture conditions, experimental errors, and/or posttranscriptional regulation, and the inherent disparities between protein and mRNA molecular stabilities. Our microarray data may provide the basic information for further exploring the different mechanisms of action for Crc.
In addition to the key regulatory genes of T3SS, several other genes, including trp, kyn, cpdA, ptrB, ptrR, prpC, rpoS, gltR, fimL, nirS, and ptrC, have been documented to play a role in the modulation of T3SS (Diaz et al. 2011), but their molecular regulatory mechanisms are largely unknown. Our microarray result suggested that in addition to the 26 T3SS genes with much reduced expression in crc mutant (Fig. 5A; Table S1), 16 genes known to be involved in the modulation of T3SS were also affected by Crc (Figs. 5B, 6; Tables S1, S2). Among these genes, seven genes (six trp genes and kynB) which are involved in tryptophan synthesis and catabolism were upregulated in crc mutant, in particular trpAB whose expression was increased by five- and sevenfold, respectively, in comparison with the parental strain (Fig. 5B; Table S2). Inactivation of either trpA or kynA gene resulted in a decrease in the T3SS-inhibiting activity, and further experiments found that the tryptophan derivatives, indole-3-acetic acid and 3-hydroxykynurenine, contribute to the inhibitory activity on T3SS expression (Shen et al. 2008). PQS, another tryptophan-derived quorum-sensing signaling molecule, was recently reported to cause inhibition of T3SS at elevated concentrations (Singh et al. 2010). We observed that deletion of trpAB in the crc mutant partially rescued the T3SS-deficient phenotype of Δcrc and deletion of trpAB in PAO1pClacZ also increased exsCEBA expression (Fig. S2A). Furthermore, the Δcrc mutant also showed increased PQS production (Fig. S2B). Thus, Crc might have modulated the T3SS in P. aeruginosa through inhibition of the tryptophan synthesis and catabolism pathway.
In addition, ptrB, cpdA, rpoS, and prpC, which were documented to possess negative regulatory effects on the expression of T3SS genes, were upregulated in the crc mutant (Figs. 5B, 6). ptrB encodes a small protein, which functions as a repressor of the expression of T3SS genes and the expression of ptrB is specifically repressed by the transcriptional regulator PrtR (Wu and Jin 2005). The expression of prtR was also upregulated in the crc mutant (Fig. 5B). cpdA encodes a cAMP phosphodiesterase, which hydrolyzes cAMP and decreases the intracellular cAMP concentration in P. aeruginosa which, in turn, reduces cAMP-Vfr-dependent virulence factor production, including T3SS (Fuchs et al. 2010). Inactivation of the stationary-phase sigma factor rpoS gene was found to be able to increase the expression of T3SS genes (Hogardt et al. 2004). Mutation in the citrate synthetase gene prpC resulted in an enhanced induction of T3SS (Rietsch and Mekalanos 2006). Notably, one gene, nirS, which encodes a nitrite reductase, was downregulated in the crc mutant (Fig. 5B). nirS was reported to have a positive effect on T3SS as its deletion mutant was defective in the expression of T3SS components in P. aeruginosa PA14 (van Alst et al. 2009). It is possible that the positive influence of Crc on the expression of T3SS genes may come from its global effect on the expression of these genes. However, regulation of Crc on other three T3SS-related genes, gltR, fimL, and ptrC seems inconsistent with its positive role in the modulation of T3SS. The gltR gene encodes a regulator for glucose transporter and was reported to have a positive effect on the expression of T3SS (Wolfgang et al. 2003). fimL encodes a protein required for type IV pili biogenesis and was also found to be required for T3SS-mediated cytotoxicity, and such activity is likely to occur, at least in part, via modulation of vfr expression and cAMP production (Whitchurch et al. 2005). ptrC gene encodes a small peptide and functions as T3SS repressor (Jin et al. 2011). Both gltR and fimL were found to be upregulated in crc mutant, but ptrC was shown to be downregulated in crc mutant (Fig. 5B). In addition, our microarray result also showed that among the 425 Crc-regulated genes, at least 39 of them encode proteins involved in transport (Figs. S1 and S2) and it is possible that some of them may have an influence on T3SS.
In summary, our data from detailed genetic and transcriptomic analysis confirmed the critical role of Crc in the regulation of T3SS in P. aeruginosa. The effect of Crc on T3SS was exerted through ExsA and was also linked to the Cbr/Crc signaling pathway. Furthermore, Crc seemed to modulate T3SS by participating in many known T3SS-related regulatory mechanisms. This complex and multifaceted effect of Crc may serve to fine-tune the T3SS to accurately sense and respond to environmental signals and nutrient sources, thereby creating a powerful virulence machinery (Fig. 7). Our results provide further information to establish the links and connections between catabolite repression and the complicated T3SS-regulatory network, which are essential for understanding the sophisticated regulatory mechanisms of the pathogen in infection and adaptation.
Figure 7.
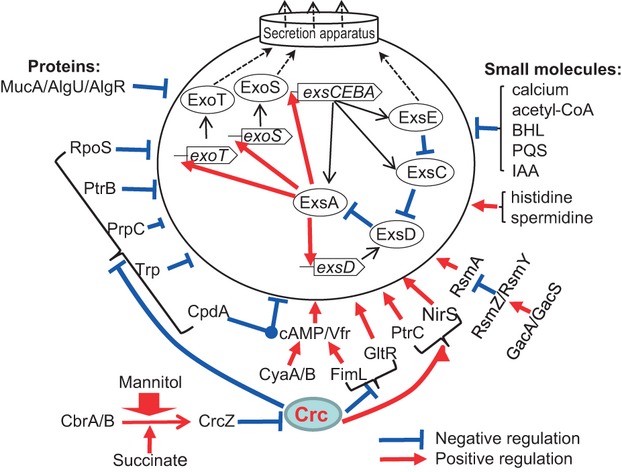
A working model of Crc in modulation of type III secretion system (T3SS) in Pseudomonas aeruginosa. Under the control of CrcZ and CbrAB two-component system, Crc positively influences the ExsA-dependent T3SS through modulating the expression of many genes and the production of some catabolites, which collectively contribute to regulation of T3SS. The key T3SS genes and encoding proteins are shown inside of the circle. The proteins and small molecules with documented regulatory role on T3SS are shown at outside the circle. Dashed line arrows indicate translocation, and solid arrows indicate positive regulation and the T bars indicate negative regulation. BHL, N-butanoyl-l-homoserine lactone; PQS, Pseudomonas quinolone signal; IAA, indole-3-acetic acid.
Acknowledgments
We thank Dr Sylvie Elsen, iRTSV, France for the anti-ExsA and anti-ExsD antiserum.
Conflict of Interest
None declared.
Supporting Information
Additional Supporting Information may be found in the online version of this article:
Figure S1. RT-PCR analysis of the selected T3SS genes in Δcrc mutant and parental strain PAO1pclacZ (pcZ).
Figure S2. Effect of trpAB deletion in Δcrc mutant (Δcrc/ΔtrpAB) and parental strain PAO1pClacZ on exsCEBA expression (A) and PQS production (B).
Table S1. Downregulated genes in crc mutant under T3SS-induced condition (≤twofold).
Table S2. Up-regulated genes in crc mutant under T3SS-induced condition (≥twofold).
Table S3. Comparison of the crc-regulated genes and predicated crc-binding genes.
References
- van Alst NE, Wellington M, Clark VL, Haidaris CG, Iglewski BH. Nitrite reductase NirS is required for type III secretion system expression and virulence in the human monocyte cell line THP-1 by Pseudomonas aeruginosa. Infect. Immun. 2009;77:4446–4454. doi: 10.1128/IAI.00822-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson GG, Yahr TL, Lovewell RR, O'Toole GA. The Pseudomonas aeruginosa magnesium transporter MgtE inhibits transcription of the type III secretion system. Infect. Immun. 2010;78:1239–1249. doi: 10.1128/IAI.00865-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aspedon A, Palmer K, Whiteley M. Microarray analysis of the osmotic stress response in Pseudomonas aeruginosa. J. Bacteriol. 2006;188:2721–2725. doi: 10.1128/JB.188.7.2721-2725.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bange G, Kümmerer N, Engel C, Bozkurt G, Wild K, Sinning I. FlhA provides the adaptor for coordinated delivery of late flagella building blocks to the type III secretion system. Proc. Natl. Acad. Sci. USA. 2010;107:11295–11300. doi: 10.1073/pnas.1001383107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bleves S, Soscia C, Nogueira-Orlandi P, Lazdunski A, Filloux A. Quorum sensing negatively controls type III secretion regulon expression in Pseudomonas aeruginosa PAO1. J. Bacteriol. 2005;187:3898–3902. doi: 10.1128/JB.187.11.3898-3902.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Browne P, Barret M, O'Gara F, Morrissey JP. Computational prediction of the Crc regulon identifies genus-wide and species-specific targets of catabolite repression control in Pseudomonas bacteria. BMC Microbiol. 2010;10:300. doi: 10.1186/1471-2180-10-300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caetano-Annoles G. Amplifying DNA with arbitrary oligonucleotide primers. PCR Methods Appl. 1993;28:85–92. doi: 10.1101/gr.3.2.85. [DOI] [PubMed] [Google Scholar]
- Dasgupta N, Lykken GL, Wolfgang MC, Yahr TL. A novel anti-anti-activator mechanism regulates expression of the Pseudomonas aeruginosa type III secretion system. Mol. Microbiol. 2004;53:297–308. doi: 10.1111/j.1365-2958.2004.04128.x. [DOI] [PubMed] [Google Scholar]
- Diaz MR, King JM, Yahr TL. Intrinsic and extrinsic regulation of Type III secretion gene expression in Pseudomonas aeruginosa. Front Microbiol. 2011;2:89. doi: 10.3389/fmicb.2011.00089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dong YH, Zhang XF, An SW, Xu JL, Zhang LH. A novel two-component system BqsS-BqsR modulates quorum sensing-dependent biofilm decay in Pseudomonas aeruginosa. Commun. Integr. Biol. 2008;1:88–96. doi: 10.4161/cib.1.1.6717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frank DW. The exoenzyme S regulon of Pseudomonas aeruginosa. Mol. Microbiol. 1997;26:621–629. doi: 10.1046/j.1365-2958.1997.6251991.x. [DOI] [PubMed] [Google Scholar]
- Fuchs EL, Brutinel ED, Klem ER, Fehr AR, Yahr TL, Wolfgang MC. In vitro and in vivo characterization of the Pseudomonas aeruginosa cyclic AMP (cAMP) phosphodiesterase CpdA, required for cAMP homeostasis and virulence factor egulation. J. Bacteriol. 2010;192:2779–2790. doi: 10.1128/JB.00168-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghosh P. Process of protein transport by the type III secretion system. Microbiol. Mol. Biol. Rev. 2004;68:771–795. doi: 10.1128/MMBR.68.4.771-795.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goodman AL, Kulasekara B, Rietsch A, Boyd D, Smith RS, Lory S. A signaling network reciprocally regulates genes associated with acute infection and chronic persistence in Pseudomonas aeruginosa. Dev. Cell. 2004;7:745–754. doi: 10.1016/j.devcel.2004.08.020. [DOI] [PubMed] [Google Scholar]
- Heeb S, Itoh Y, Nishijyo T, Schnider U, Keel C, Wade J, et al. Small, stable shuttle vectors based on the minimal pVS1 replicon for use in gram-negative, plant-associated bacteria. Mol. Plant Microbe Interact. 2000;13:232–237. doi: 10.1094/MPMI.2000.13.2.232. [DOI] [PubMed] [Google Scholar]
- Hester KL, Lehman J, Najar F, Song L, Roe BA, MacGregor CH, et al. Crc is involved in catabolite repression control of the bkd operons of Pseudomonas putida and Pseudomonas aeruginosa. J. Bacteriol. 2000;182:1144–1149. doi: 10.1128/jb.182.4.1144-1149.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoang TT, Karkhoff-Schweizer RR, Kutchma AJ, Schweizer HP. A broad-host-range Flp-FRT recombination system for site-specific excision of chromosomally-located DNA sequences: application for isolation of unmarked Pseudomonas aeruginosa mutants. Gene. 1998;212:77–86. doi: 10.1016/s0378-1119(98)00130-9. [DOI] [PubMed] [Google Scholar]
- Hogardt M, Roeder M, Schreff AM, Eberl L, Heesemann J. Expression of Pseudomonas aeruginosa exoS is controlled by quorum sensing and RpoS. Microbiology. 2004;150:843–851. doi: 10.1099/mic.0.26703-0. [DOI] [PubMed] [Google Scholar]
- Hovey AK, Frank DW. Analyses of the DNA-binding and transcriptional activation properties of ExsA, the transcriptional activator of the Pseudomonas aeruginosa exoenzyme S regulon. J. Bacteriol. 1995;177:4427–4436. doi: 10.1128/jb.177.15.4427-4436.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jin Y, Yang H, Qiao M, Jin S. MexT regulates the type III secretion system through MexS and PtrC in Pseudomonas aeruginosa. J. Bacteriol. 2011;193:399–410. doi: 10.1128/JB.01079-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Journet L, Hughes KT, Cornelis GR. Type III secretion: a secretory pathway serving both motility and virulence. Mol. Membr. Biol. 2005;22:41–50. doi: 10.1080/09687860500041858. [DOI] [PubMed] [Google Scholar]
- Kulasekara HD, Ventre I, Kulasekara BR, Lazdunski A, Filloux A, Lory S. A novel two-component system controls the expression of Pseudomonas aeruginosa fimbrial cup genes. Mol. Microbiol. 2005;55:368–380. doi: 10.1111/j.1365-2958.2004.04402.x. [DOI] [PubMed] [Google Scholar]
- Laskowski MA, Osborn E, Kazmierczak BI. A novel sensor kinase-response regulator hybrid regulates type III secretion and is required for virulence in Pseudomonas aeruginosa. Mol. Microbiol. 2004;54:1090–1103. doi: 10.1111/j.1365-2958.2004.04331.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Linares JF, Moreno R, Fajardo A, Martínez-Solano L, Escalante R, Rojo F, et al. The global regulator Crc modulates metabolism, susceptibility to antibiotics and virulence in Pseudomonas aeruginosa. Environ. Microbiol. 2010;12:3196–3212. doi: 10.1111/j.1462-2920.2010.02292.x. [DOI] [PubMed] [Google Scholar]
- MacGregor CH, Wolff JA, Arora SK, Phibbs PV., Jr Cloning of a catabolite repression control crc. gene from Pseudomonas aeruginosa, expression of the gene in Escherichia coli, and identification of the gene product in Pseudomonas aeruginosa. J. Bacteriol. 1991;173:7204–7212. doi: 10.1128/jb.173.22.7204-7212.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacGregor CH, Arora SK, Hager PW, Dail MB, Phibbs PV., Jr The nucleotide sequence of the Pseudomonas aeruginosa pyrE-crc-rph region and the purification of the crc gene product. J. Bacteriol. 1996;178:5627–5635. doi: 10.1128/jb.178.19.5627-5635.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCaw ML, Lykken GL, Singh PK, Yahr TL. ExsD is a negative regulator of the Pseudomonas aeruginosa type III secretion regulon. Mol. Microbiol. 2002;46:1123–1133. doi: 10.1046/j.1365-2958.2002.03228.x. [DOI] [PubMed] [Google Scholar]
- Morales G, Linares JF, Beloso A, Albar JP, Martínez JL, Rojo F. The Pseudomonas putida Crc global regulator controls the expression of genes from several chromosomal catabolic pathways for aromatic compounds. J. Bacteriol. 2004;186:1337–1344. doi: 10.1128/JB.186.5.1337-1344.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moreno R, Ruiz-Manzano A, Yuste L, Rojo F. The Pseudomonas putida Crc global regulator is an RNA binding protein that inhibits translation of the AlkS transcriptional regulator. Mol. Microbiol. 2007;64:665–675. doi: 10.1111/j.1365-2958.2007.05685.x. [DOI] [PubMed] [Google Scholar]
- Moreno R, Marzi S, Romby P, Rojo F. The Crc global regulator binds to an unpaired A-rich motif at the Pseudomonas putida alkS mRNA coding sequence and inhibits translation initiation. Nucleic Acids Res. 2009;37:7678–7690. doi: 10.1093/nar/gkp825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moreno R, Fonseca P, Rojo F. The Crc global regulator inhibits the Pseudomonas putida pWW0 toluene/xylene assimilation pathway by repressing the translation of regulatory and structural genes. J. Biol. Chem. 2010;285:24412–24419. doi: 10.1074/jbc.M110.126615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicas TI, Iglewski BH. Isolation and characterization of transposon-induced mutants of Pseudomonas aeruginosa deficient in production of exoenzyme S. Infect. Immun. 1984;45:470–474. doi: 10.1128/iai.45.2.470-474.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishijyo T, Haas D, Itoh Y. The CbrA-CbrB two-component regulatory system controls the utilization of multiple carbon and nitrogen sources in Pseudomonas aeruginosa. Mol. Microbiol. 2001;40:917–931. doi: 10.1046/j.1365-2958.2001.02435.x. [DOI] [PubMed] [Google Scholar]
- O'Toole GA, Gibbs KA, Hager PW, Phibbs PV, Jr, Kolter R. The global carbon metabolism regulator Crc is a component of a signal transduction pathway required for biofilm development by Pseudomonas aeruginosa. J. Bacteriol. 2000;182:425–431. doi: 10.1128/jb.182.2.425-431.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Potts JR, Clarke PH. The effect of nitrogen limitation on catabolite repression of amidase, histidase and urocanase in Pseudomonas aeruginosa. J. Gen. Microbiol. 1976;93:377–387. doi: 10.1099/00221287-93-2-377. [DOI] [PubMed] [Google Scholar]
- Rietsch A, Mekalanos JJ. Metabolic regulation of type III secretion gene expression in Pseudomonas aeruginosa. Mol. Microbiol. 2006;59:807–820. doi: 10.1111/j.1365-2958.2005.04990.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rietsch A, Wolfgang MC, Mekalanos JJ. Effect of metabolic imbalance on expression of type III secretion genes in Pseudomonas aeruginosa. Infect. Immun. 2004;72:1383–1390. doi: 10.1128/IAI.72.3.1383-1390.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rietsch A, Vallet-Gely I, Dove SL, Mekalanos JJ. ExsE, a secreted regulator of type III secretion genes in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA. 2005;102:8006–8011. doi: 10.1073/pnas.0503005102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rojo F. Carbon catabolite repression in Pseudomonas: optimizing metabolic versatility and interactions with the environment. FEMS Microbiol. Rev. 2010;34:658–684. doi: 10.1111/j.1574-6976.2010.00218.x. [DOI] [PubMed] [Google Scholar]
- Sato H, Frank DW. ExoU is a potent intracellular phospholipase. Mol. Microbiol. 2004;53:1279–1290. doi: 10.1111/j.1365-2958.2004.04194.x. [DOI] [PubMed] [Google Scholar]
- Shen DK, Filopon D, Kuhn L, Polack B, Toussaint B. PsrA is a positive transcriptional regulator of the type III secretion system in Pseudomonas aeruginosa. Infect. Immun. 2006;74:1121–1129. doi: 10.1128/IAI.74.2.1121-1129.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen DK, Filopon D, Chaker H, Boullanger S, Derouazi M, Polack B, et al. High-cell-density regulation of the Pseudomonas aeruginosa type III secretion system: implications for tryptophan catabolites. Microbiology. 2008;154:2195–2208. doi: 10.1099/mic.0.2007/013680-0. [DOI] [PubMed] [Google Scholar]
- Simon R, Prifer U, Puhler A. A broad range mobilization system for in vivo genetic engineering: transposon mutagenesis in gram-negative bacteria. Biotechnology. 1983;1:784–791. [Google Scholar]
- Singh G, Wu B, Baek MS, Camargo A, Nguyen AA, Slusher NA, et al. Secretion of Pseudomonas aeruginosa type III cytotoxins is dependent on pseudomonas quinolone signal concentration. Microb. Pathog. 2010;49:196–203. doi: 10.1016/j.micpath.2010.05.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sonnleitner E, Abdou L, Haas D. Small RNA as global regulator of carbon catabolite repression in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA. 2009;106:21866–21871. doi: 10.1073/pnas.pnas.0910308106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Urbanowski ML, Lykken GL, Yahr TL. A secreted regulatory protein couples transcription to the secretory activity of the Pseudomonas aeruginosa type III secretion system. Proc. Natl. Acad. Sci. USA. 2005;102:9930–9935. doi: 10.1073/pnas.0504405102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whitchurch CB, Beatson SA, Comolli JC, Jakobsen T, Sargent JL, Bertrand JJ, et al. Pseudomonas aeruginosa fimL regulates multiple virulence functions by intersecting with Vfr-modulated pathways. Mol. Microbiol. 2005;55:1357–1378. doi: 10.1111/j.1365-2958.2005.04479.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolff JA, MacGregor CH, Eisenberg RC, Phibbs PV., Jr Isolation and characterization of catabolite repression control mutants of Pseudomonas aeruginosa PAO. J. Bacteriol. 1991;173:4700–4706. doi: 10.1128/jb.173.15.4700-4706.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolfgang MC, Lee VT, Gilmore ME, Lory S. Coordinate regulation of bacterial virulence genes by a novel adenylate cyclase-dependent signaling pathway. Dev. Cell. 2003;4:253–263. doi: 10.1016/s1534-5807(03)00019-4. [DOI] [PubMed] [Google Scholar]
- Wu W, Jin S. PtrB of Pseudomonas aeruginosa suppresses the type III secretion system under the stress of DNA damage. J. Bacteriol. 2005;187:6058–6068. doi: 10.1128/JB.187.17.6058-6068.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu W, Badrane H, Arora S, Baker HV, Jin S. MucA-mediated coordination of type III secretion and alginate synthesis in Pseudomonas aeruginosa. J. Bacteriol. 2004;186:7575–7585. doi: 10.1128/JB.186.22.7575-7585.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yahr TL, Frank DW. Transcriptional organization of the trans-regulatory locus which controls exoenzyme S synthesis in Pseudomonas aeruginosa. J. Bacteriol. 1994;176:3832–3838. doi: 10.1128/jb.176.13.3832-3838.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yahr TL, Wolfgang MC. Transcriptional regulation of the Pseudomonas aeruginosa type III secretion system. Mol. Microbiol. 2006;62:631–640. doi: 10.1111/j.1365-2958.2006.05412.x. [DOI] [PubMed] [Google Scholar]
- Yahr TL, Hovey AK, Kulich SM, Frank DW. Transcriptional analysis of the Pseudomonas aeruginosa exoenzyme S structural gene. J. Bacteriol. 1995;177:1169–1178. doi: 10.1128/jb.177.5.1169-1178.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yeung AT, Bains M, Hancock RE. The sensor kinase CbrA is a global regulator that modulates metabolism, virulence, and antibiotic resistance in Pseudomonas aeruginosa. J. Bacteriol. 2011;193:918–931. doi: 10.1128/JB.00911-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou L, Wang J, Zhang LH. Modulation of bacterial Type III secretion system by a spermidine transporter dependent signaling pathway. PLoS ONE. 2007;2:e1291. doi: 10.1371/journal.pone.0001291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zolfaghar I, Angus AA, Kang PJ, To A, Evans DJ, Fleiszig SM. Mutation of retS, encoding a putative hybrid two-component regulatory protein in Pseudomonas aeruginosa, attenuates multiple virulence mechanisms. Microbes Infect. 2005;7:1305–1316. doi: 10.1016/j.micinf.2005.04.017. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.