

Abstract
Infectious diseases caused by parasites are a major threat for the entire mankind, especially in the tropics. More than 1 billion people world-wide are directly exposed to tropical parasites such as the causative agents of trypanosomiasis, leishmaniasis, schistosomiasis, lymphatic filariasis and onchocerciasis, which represent a major health problem, particularly in impecunious areas. Unlike most antibiotics, there is no “general” antiparasitic drug available. Here, the selection of antiparasitic drugs varies between different organisms. Some of the currently available drugs are chemically de novo synthesized, however, the majority of drugs are derived from natural sources such as plants which have subsequently been chemically modified to warrant higher potency against these human pathogens. In this review article we will provide an overview of the current status of plant derived pharmaceuticals and their chemical modifications to target parasite-specific peculiarities in order to interfere with their proliferation in the human host.
Keywords: neglected infectious diseases, natural compounds, schistosomiasis, river blindness, trypanosomatids
1. Introduction
Infectious diseases have a significant impact in human heath stock. A variety of these diseases are caused by parasites that belong to the diseases of poverty or the so-called neglected tropical diseases. Amongst others, these include the causative agents of trypanosomiasis, leishmaniasis, schistosomiasis, lymphatic filariasis and onchocerciasis. These parasites are responsible for a high rate of mortality and morbidity each year in the respective endemic countries. Since vaccines and safe and affordable treatments are still lacking, there is an urgent need to discover novel therapeutics against these human health threats. The current situation is aggravated by the fact that most people living in the endemic countries have a low-income profile, making the pharmaceutical market for the development of new medications financially unattractive to the private-research sector. Therefore, natural sources, such as plants, represent a major opportunity to discover new lead molecules [1–4]. In this review we will shed light on the discovery and application of natural plant derived products on human neglected diseases.
2. Nematodes
Nematodes are roundworms that belong to the phylum Nematoda or Nemathelminthes [5]. Even though the total number of nematode species has been estimated to be over 1 million [6], only about 28,000 species have been described [7], with over 16,000 of these species being parasites of plants and animals (including humans). Parasitic nematodes constitute an enormous medical and veterinary burden in some parts of the world; in the tropics, however, they represent a severe public health problem. The most common and persistent parasitic nematodes of humans are the soil-transmitted nematodes—roundworms (Ascaris lumbricoides), whipworms (Trichuris trichiura), hookworms (Necator americanus and Ancylostoma duodenale) and thread worms (Strongyloides stercoralis)—and the filarial nematodes that are responsible for lymphatic filariasis (LF) (Brugia sp, Wuchereria bancrofti) and onchocerciasis (Onchocerca volvulus).
Onchocerciasis or subcutaneous filariasis is a parasitic disease caused by O. volvulus. The disease affects several millions of people in the world and is transmitted from host to host by the blood-feeding “blackfly” Simulium. While more than 90% of all onchocerciasis cases are found in 30 African countries, the rest occur in isolated foci that exist in Yemen and six countries in central and South America World Health Organization [8]. About 37 million persons are infected with O. volvulus, of whom 270,000 are blind and 500,000 visually impaired [9].
LF is a mosquito-borne infection caused by the lymphatic dwelling parasites W. bancrofti, B. malayi and B. timori. These parasites invade and block the lymphatic system of the infected person [10]. Lymphatic vessel dysfunction and damage ultimately lead to clinical symptoms such as lymphoedema, elephantiasis and genital pathology. The WHO estimates that 1.1 billion people live in high risk areas, with 120 million people already infected with LF [11,12]. The filarial nematode W. bancrofti accounts for 91% of LF infections, while B. malayi and B. timori are responsible for the remaining 9% in the Southeast Asia region.
The burden of onchocerciasis and LF causes long term disability, social stigmatization and, in the case of onchocerciasis, forces the affected population to abandon the infested areas which usually have a high agricultural potential [13,14]. Thus, a high burden of onchocerciasis primarily leads to a highly unproductive population and consequently to economic loss and slowdown of country development over the years.
During the last three decades, a lot of progress has been made in the control of onchocerciasis and LF; however, the diseases still constitute a serious public health concern in the endemic countries. From 1987 to date, the control of LF and onchocerciasis has been based on two approaches: vector control using insecticides and mass drug administration of diethylcarbamazine (DEC), ivermectin and albendazole. While ivermectin is the sole drug used in community-directed treatment of onchocerciasis, DEC or ivermectin in combinantion with albendazole is used for the global control of LF [15]. The recommended treatment of filariasis patients is the administration of albendazole combined with ivermectin [16,17]. A combination of DEC and albendazole is also effective [16]. All of these treatments are microfilaricides and only show a limited macrofilaricidal activity.
The development of anthelminthic resistance is a worldwide reality [9,18–20] and also one of the greatest challenges in worm control. Early in the last decade, resistances to insecticides and ivermectin have been reported [9,18]. Additionally, a re-infestation phenomenon was observed in insecticide-treated areas. Due to these limitations, vector control was stopped. Drugs to treat onchocerciasis and LF are commonly used in combination to reduce microfilariae in the blood and skin. However, all of them have their limits. In fact, DEC or ivermectin treatment of infected individuals can cause high adverse effects [21].
Anthelmintic resistance, secondary effects and limited macrofilaricidal activities of the known antifilarial drugs have stimulated the search for alternative treatment. Here, as an alternative source of filaricidal compounds, medicinal plants have received more attention [22]. Medicinal plants have been used as therapies in traditional health care systems since prehistoric times and are still the most important health care source for the majority of the world population [23]. It is estimated that more than 60% of the world’s population rely on traditional herbal medicine to meet their primary health care needs [24].
2.1. Medicinal Plants in the Treatment and Control of Filariasis
2.1.1. Subcutaneous Filariasis
Subcutaneous filariasis or onchocerciasis is a parasitic disease that is caused by O. volvulus. The cattle parasite O. ochengi is the closest known relative of O. volvulus with which it shares the same arthropod vector, Simulium damnosum. The O. ochengi system fills the critical niche between laboratory studies in rodent models and field evaluation of onchocerciasis control in human populations. The cattle—O. ochengi model equals that of human onchocerciasis, with nodules that closely resemble those formed by O. volvulus [25]. Thus it is feasible that medicinal plants, traditionally used by farmers against the bovine parasite, also affect the human parasite O. volvulus.
Nyasse et al. [26] demontrated that polycarpol from Polyalthia suaveolens (Annonaceae) and 3-O-acetyl aleuritolic acid from Discoglypremna caloneura (Euphorbiaceae) exhibited significant inhibitory activities on the vitality of adult male worms of Onchocerca gutturosa.
Further studies were conducted by Cho-Ngwa et al. [27] on O. ochengi. These authors reported microfilaricidal activity of the hexane extract of Homalium africanum (Salicaceae) leaves, the hexane extract of Margaritaria discoidea (Euphorbiaciaea) roots, the methylene chloride extract of H. africanum leaves and the methylene chloride extract of M. discoidea leaves. However, none of the plants used showed macrofilaricidal activity. These Salicaceae and Euphorbiaceae are commonly used in the traditional treatment of onchocerciasis in North West Cameroon.
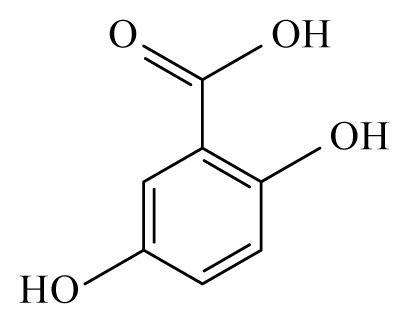
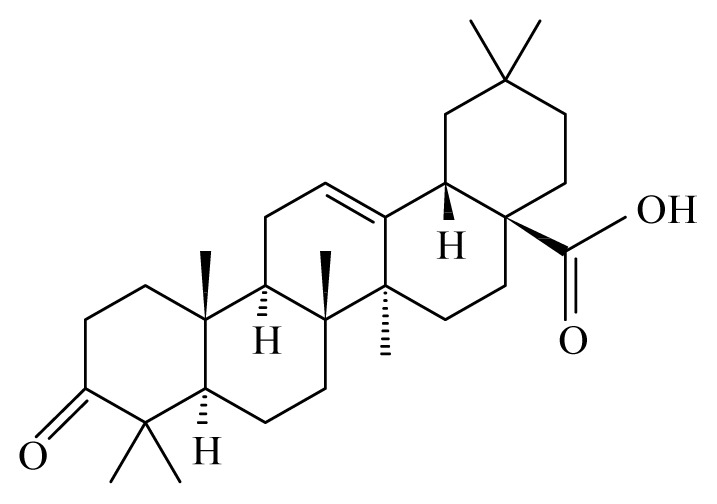
Using the cattle parasite O. ochengi, Ndjonka et al. [28] reported that ethanolic extracts of the bark of Anogeissus leiocarpus (Combretaceae) and Khaya senegalensis (Meliaceae) as well as leaves of K. senegalensis and Euphorbia hirta (Euphorbiaciaea) display high macro- and microfilaricidal activities, while aqueous extracts of leaves from Parquetina nigrescens (Asclepiadaceae) and Annona senegalensis (Annonaceae) displayed a more moderate effect on the worms viability. This was also observed using Caenorhabditis elegans, a highly suitable and free-living model organism for research on nematode parasites. Furthermore, some selected plants showed toxicity not only against wildtype C. elegans but also against drug resistant (ivermectin, levamisole and albendazole) strains [29]. Here, the most promising plant was A. leiocarpus (Combretaceae) with a high toxicity against O. ochengi, C. elegans wildtype and C. elegans drug resistant strains [29]. The phytochemical analysis of an A. leiocarpus extract showed a high amount of tannins, which have been reported to have a certain anthelmintic activity [30,31]. Tannins present in their composition several phenolic groups such as ellagic, gallic and gentisic acids. Gallic and gentisic acids have been reported to be toxic for C. elegans [30]. Recently, we showed that ellagic acid exhibited higher toxicity against C. elegans wildtype and drug resistant strains (Ndjonka and Liebau, personal communication). High microfilaricidal and macrofilaricidal activities were also reported with ellagic acid [29].
Thomsen et al. [32] used the anthelmintic active extracts of Hagenia abyssinica (Rosaceae) to develop a simple and inexpensive bioassay against the non-parasitic nematode C. elegans.
Katiki et al. [33] demonstrated that extracts rich in hydrolysable tannins such as Acer rubrum (Aceraceae), Rosa multiflora (Rosaceae) and Quercus alba (Fagaceae), or extracts rich in both hydrolysable and condensed tannins such as Rhus typhina (Anacardiaceae), were significantly more lethal to adult of C. elegans than extracts containing only condensed tannins such as Lespedeza cuneata (Fabaceae), Salix X sepulcralis (Salicaceae) and Robinia pseudoacacia (Fabaceae).
Our online search on medicinal plants used against onchocerciasis found nine publications since 2002, where a total of 17 plant species, belonging to 10 different families, have been studied. In these studies, only five pure compounds were isolated (Table 1).
Table 1.
List of medicinal plants, products and active compounds known to have antifilarial properties.
| Names | Family | Parts used | Solvent used for extraction | Active compounds | Activities | References |
|---|---|---|---|---|---|---|
| Polyalthia suaveolens | Annonaceae |
 Polycarpol |
O. gutturosa: Significant inhibitory activities on the vitality of adult male worms | [26] | ||
| Discoglypremna caloneura | Euphorbiaceae |
 3-O-acetyl aleuritolic acid |
||||
|
| ||||||
| Homalium africanum | Salicaceae | Leaves | Hexane Methylene chloride | O. ochengi: Microfilaricide | [27] | |
|
| ||||||
| Margaritaria discoidea | Euphorbiaciaea | Roots Leaves | Hexane Methylene chloride | O. ochengi: Microfilaricide | ||
|
| ||||||
| Anogeissus leiocarpus | Combretaceae | Bark, leaves | Ethanol |
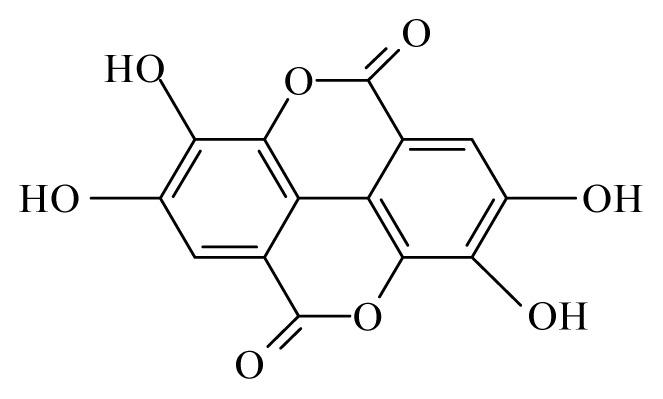 Ellagic acid |
O. ochengi: Microfilaricide and macrofilaricide. C. elegans: High activity on adults and larvae |
[28,29] |
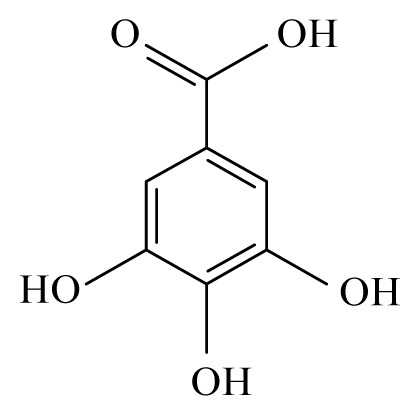 Gallic acid | ||||||
 Gentisic acid | ||||||
|
| ||||||
| Khaya senegalensis | Meliaceae | Bark, leaves | Ethanol |
O. ochengi: Microfilaricide and macrofilaricide. C. elegans: Moderate activity on adults and larvae |
[28] | |
|
| ||||||
| Euphorbia hirta | Euphorbiaciaea | Leaves | Ethanol | C. elegans: Moderate activity on adults and larvae | ||
|
| ||||||
| Parquetina nigrescens | Asclepiadaceae | Water | [28] | |||
|
| ||||||
| Annona senegalensis | Annonaceae | Water | C. elegans: Moderate activity on adults and larvae | [28] | ||
|
| ||||||
| Hagenia abyssinica | Rosaceae | Female flowers | 80% Methanol | [32] | ||
|
| ||||||
| Acer rubrum | Aceraceae | |||||
| Rosa multiflora | Rosaceae | |||||
| Quercus alba | Fagaceae | Leaves | 70% Acetone | C. elegans: Moderate activity on adults and larvae | [33] | |
| Rhus typhina | Anacardiaceae | |||||
|
| ||||||
| Lespedeza cuneata | Fabaceae | |||||
| Salix X sepulcralis | Salicaceae | Leaves | 70% Acetone | C. elegans: Low activity on adults and larvae | ||
| Robinia pseudoacacia | Fabaceae | |||||
| Botryocladia leptopoda | Rhodymeniaceae | Whole alga | 95% Ethanol |
L. sigmodontis and A. viteae: Adults B. malayi: Macrofilaricide and sterilization of female |
[34] | |
|
| ||||||
| Neurolaena lobata | Asteraceae | Leaves | Ethanol | B. pahangi: Macrofilaricide and micrifilaricide | [35] | |
|
| ||||||
| Lantana camara | Verbenaceae | Stem | 95% Ethanol | A. viteae: Microfilaricide (95.04%) and sterilization of female (60.66%) | [36] | |
| B. malayi: Mastomys coucha killed 43.05% of the adult and sterilized 76% females | ||||||
|
| ||||||
| Lantana camara | Verbenaceae | Stem | Chloroform fraction |
 Oleanonic acid |
B. malayi: in M. coucha (Meriones unguiculatus) killed 100% (80%) of the adult | [36] |
| B. malayi: Macrofilaricide | ||||||
|
| ||||||
| Butea monosperma | Fabaceae | Leaves | Water | Polyphenol | B. malayi: Strong inhibition of motility microfilariae, presence of oxidative parameters | [36–38] |
| Roots | Water | B. malayi: Strong inhibition of motility microfilariae | [36,37] | |||
|
| ||||||
| Vitex negundo | Lamiaceae | Roots | Ethanol | Alkaloids, saponin, flavonoids, polyphenol | B. malayi: Strong inhibition of motility microfilariae, presence of oxidative parameters | |
| Aegle marmelo | Rutaceae | Leaves | Ethanol |
 Coumarin Polyphenol |
B. malayi: Strong inhibition of motility microfilariae, presence of oxidative parameters | [36–38] |
| Ricinus communis | Euphorbiaceae | Leaves | 70% Methanol | B. malayi: Moderate inhibition of motility microfilariae | ||
|
| ||||||
| Caesalpinia bonducella | Caesalpiniaceae | Seed kernel | Ethanol | L. sigmodontis in cotton rats S. hispidus: Reduction to up to 96% filariae and 100% female sterilization. Microfilaricide in B. malayi. | ||
| Butanol fraction | L. sigmodontis in cotton rats S. hispidus: Reduction to up to 73.7% microfilariae. 82.5% mortality of macrofilaria and 100% female sterilization. Microfilaricide in B. malayi. | [39] | ||||
| Aqueous fraction | L. sigmodontis in cotton rats S. hispidus: Reduction to up to 90% microfilariae. 82.5% mortality of macrofilaria and 100% female sterilization. Microfilaricide in B. malayi. | |||||
|
| ||||||
| Trachyspermum ammi | Apiaceae | Fruits | Methanol | Phenolic monoterpene | S. digitata: Macrofilaricide B. malayi: Macrofilaricide and females sterilization | [40] |
|
| ||||||
| Piper betle | Piperaceae | Leaves | Methanol | n-Hexane and chloroform fractions | B. malayi: Microfilaricide, moderate activity on macrofilariae and female sterilization. Immunomodulatory properties in mices | [41] |
|
| ||||||
| Xylocarpus granatum | Meliaceae | Fruits | 50% Ethanol |
 Gedunin |
B. malayi: Excellent microfilaricidal and macrofilaricidal efficacies | [42,43] |
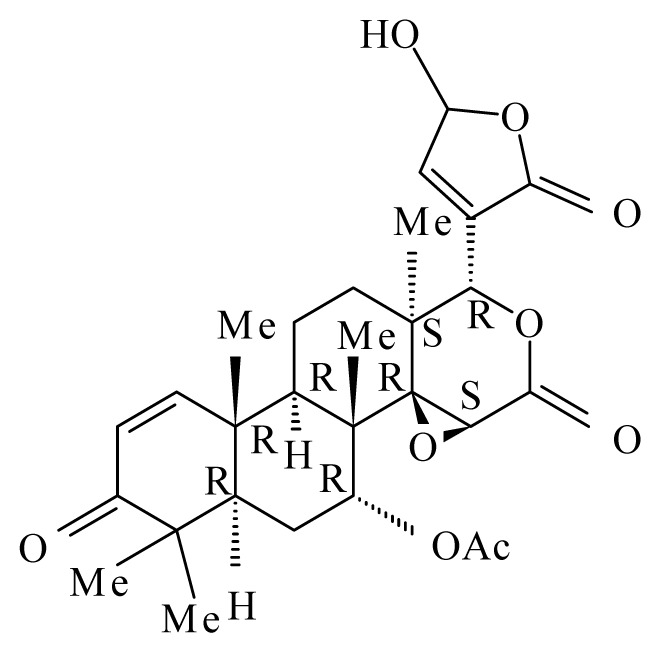 Photogedunin | ||||||
|
| ||||||
| Bauhinia racemosa | Caesalpinaeceae | Leaves | 95% Ethanol |
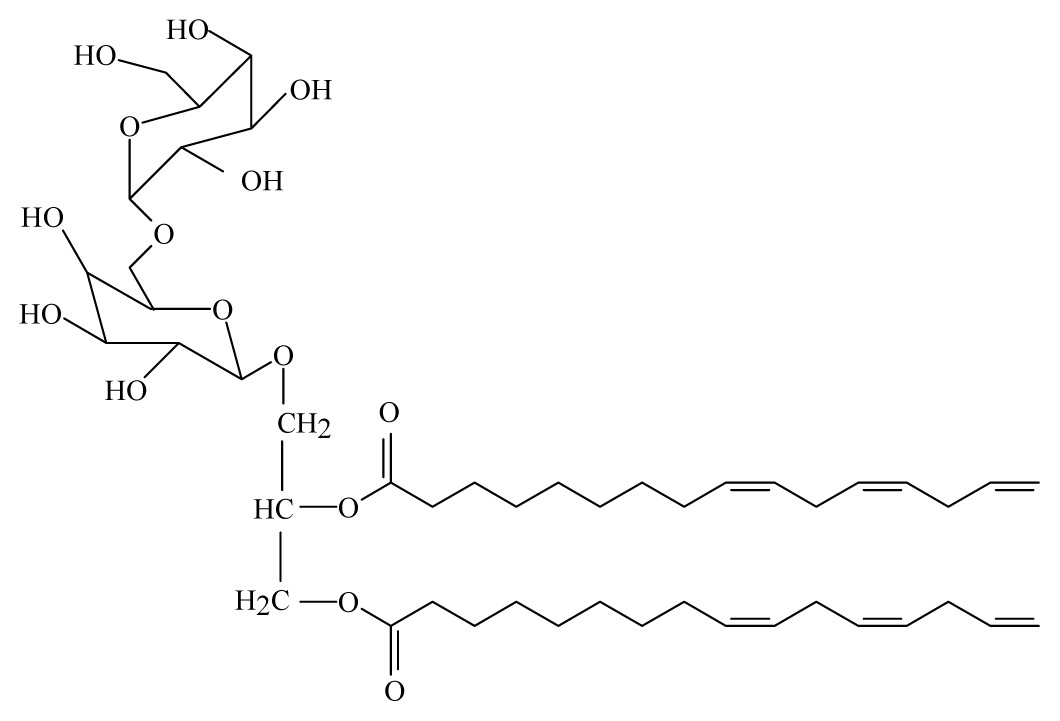 Galactolipid 1 |
B. malayi: In vivo and in vitro antifilarial activity | [44] |
|
| ||||||
| Bauhinia racemosa | Caesalpinaeceae | Leaves | 95% Ethanol |
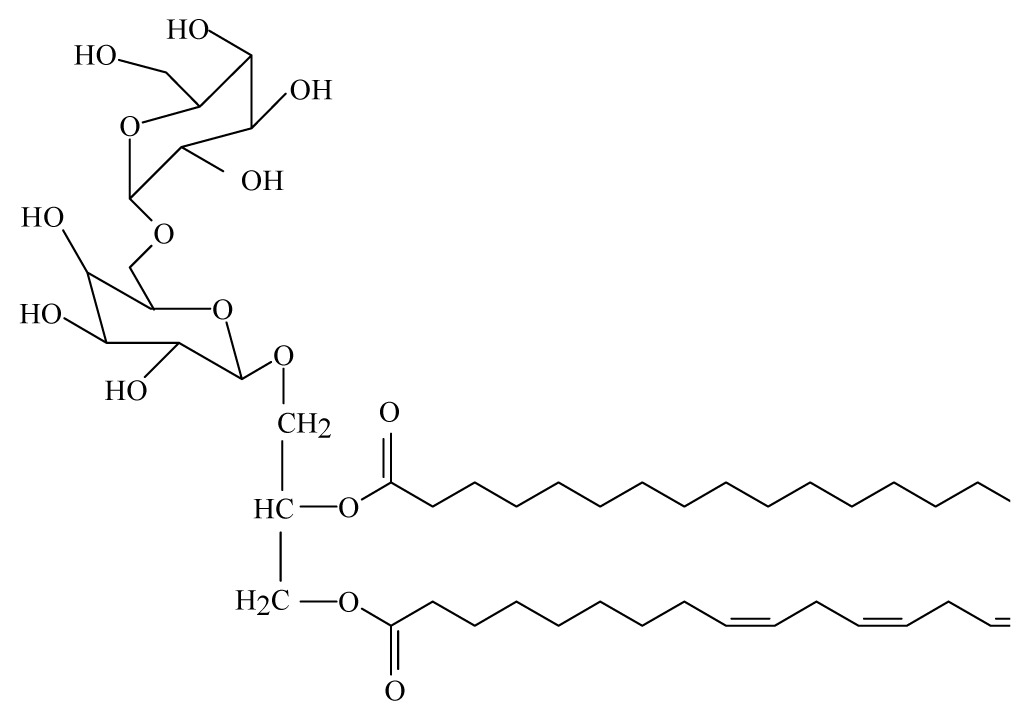 Galactolipid 2 |
B. malayi: In vivo and in vitro antifilarial activity | [44] |
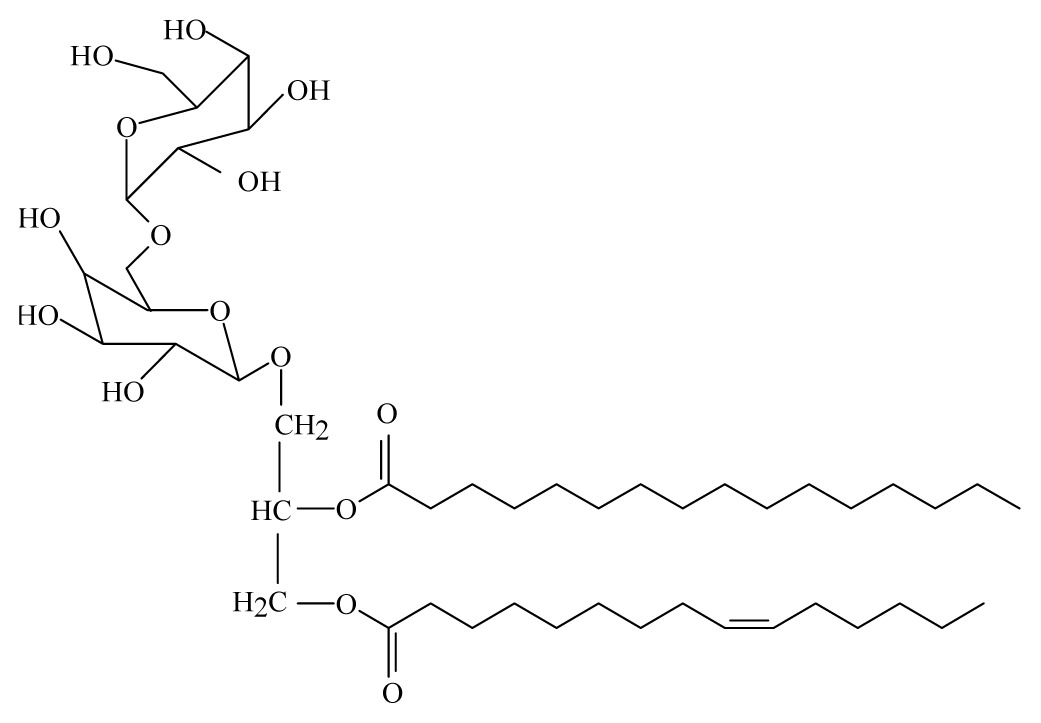 Galactolipid 3 | ||||||
|
| ||||||
| Corinder: Coriandrum sativum | Apiaceae |
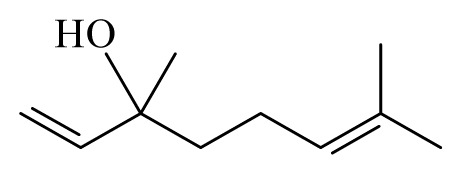 Linalool |
[45] | |||
 α-Pinene | ||||||
|
| ||||||
| Cassia: Cassia | Fabaceae |
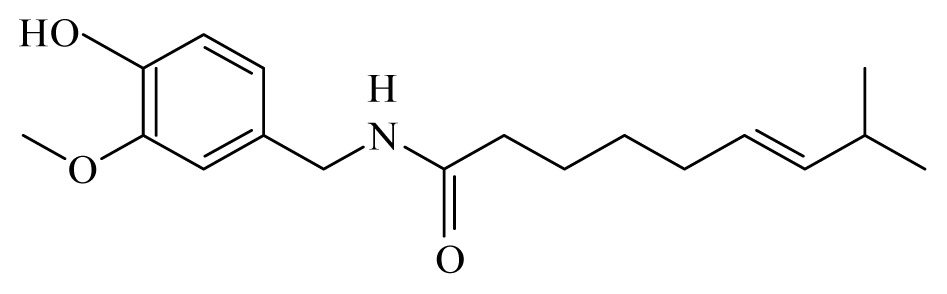 Capsaicin |
[45] | |||
| Turmeric: Curcuma longa | Zingiberaceae |
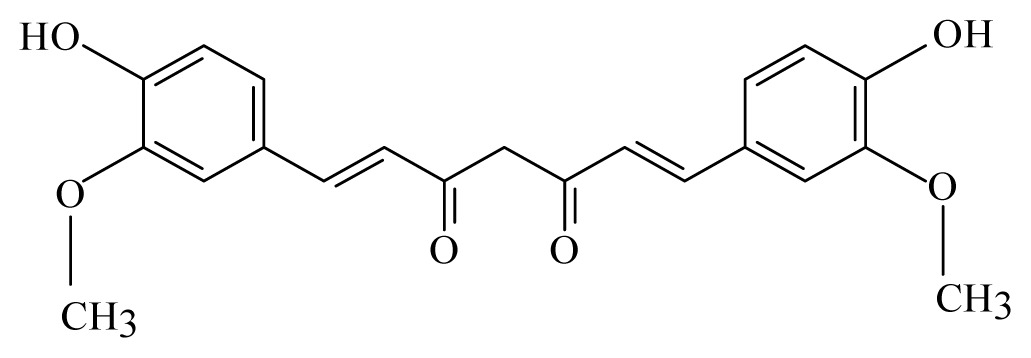 Curcumin |
||||
| Allspice: Pimenta dioica | Myrtaceae |
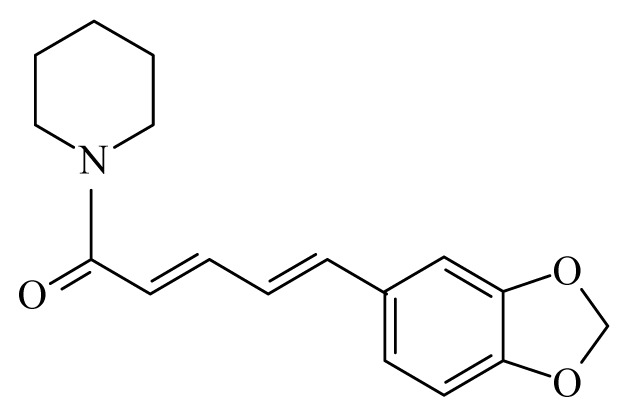 Piperine |
||||
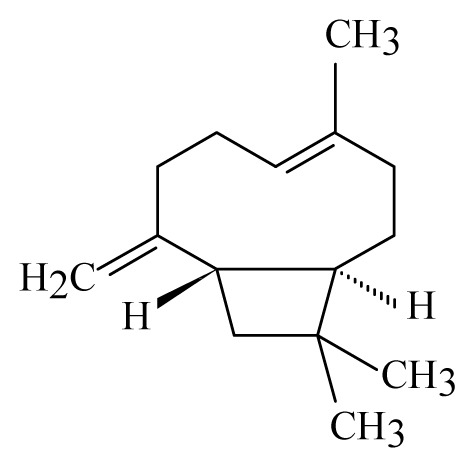 β-Caryophyllene |
||||||
|
| ||||||
| Cinnamon: Cinnamomum | Lauraceae |
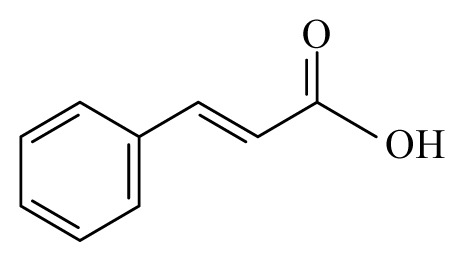 Cinnamic acid β-Caryophyllene |
B. malayi: GST inhibitory activity in vitro | [45] | ||
| Strychnous: Strychnos | Loganiaceae |
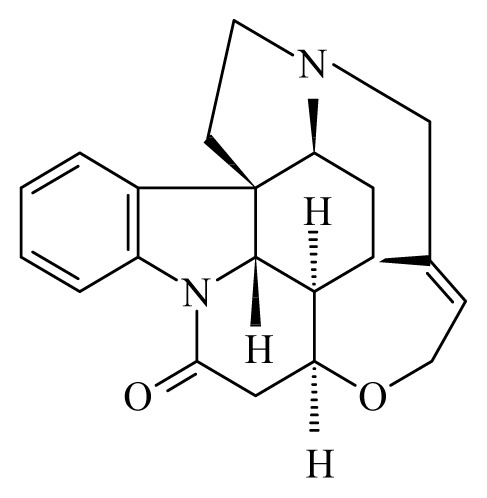 Strychnine |
||||
| Lemongrass: Cymbopogon | Poaceae |
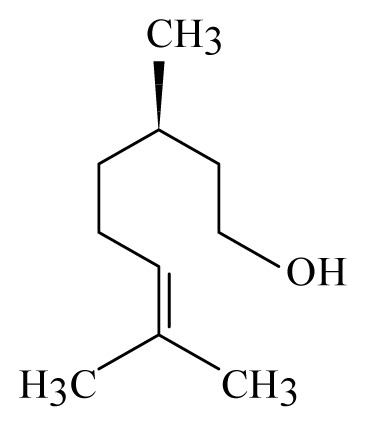 Citronellol |
||||
 Geraniol |
||||||
| Garlic: Allium sativum | Amaryllidaceae |
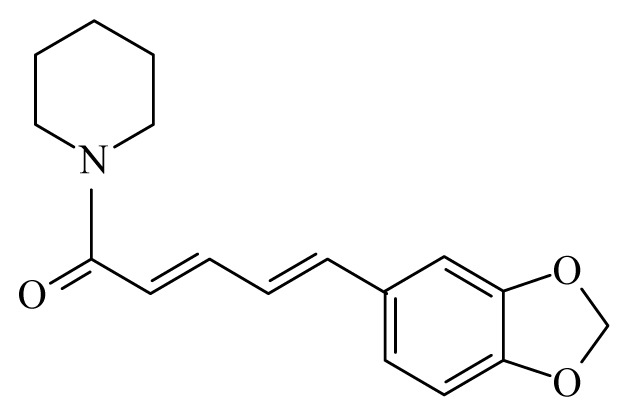 Piperine |
||||
|
| ||||||
| Litsea: Litsea | Lauraceae |
 Vanillin |
[45] | |||
| Vanilla: Vanilla | Orchidaceae |
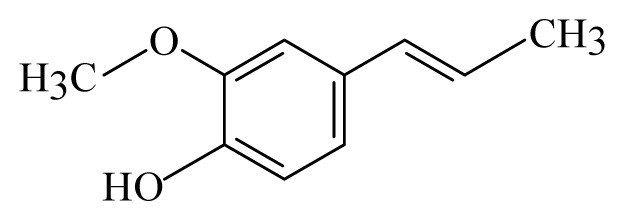 Isoeugenol |
||||
|
| ||||||
| Withania somnifera | Solanaceae Roots |
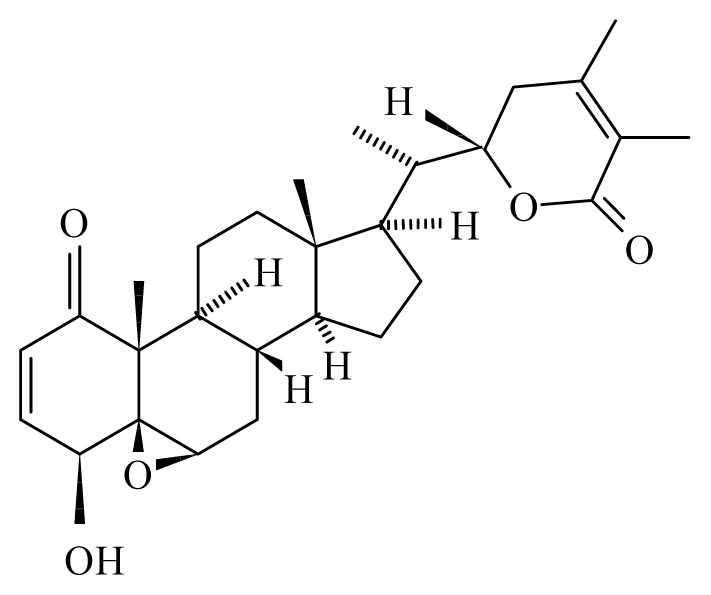 Withanolide |
Protection to the rodent host M. coucha against infection of filarial parasite B. malayi | [46,47] | ||
|
| ||||||
| Nigella sativa | Ranunculaceae | Seeds | B. malayi: Immunomodulatory and therapeutic properties in mices | [48] | ||
Galactolipid 1: (2S)-1, 2-di-O-linolenoyl-3-O-a-galactopyra-nosyl-(1/6)-O-b-galactopy-ranosyl glycerol; Galactolipid 2: (2S)-1-O-linolenoyl-2-O-palmitoyl-3-O-a-galacto-pyranosyl-(1/6)- O-b-galactopyranosyl glycerol; Galactolipid 3: (2S)-1-O oleoyl-2-O-palmitoyl-3-O-a-galacto-pyranosyl-(1/6)-O-b-galactopyranosyl glycerol.
2.1.2. Lymphatic Filariasis
LF is a parasitic disease that is caused by Brugia sp. and W. bancrofti. Current control programs outside sub-Saharan Africa use DEC plus albendazole or DEC alone, while in Africa ivermectin plus albendazole is used because of contraindications for DEC in patients infected with O. volvulus [16,17]. However, there is no effective drug that targets the adult stage of the worms. Based on the observation that filarial worms depend on the endosymbiontic bacteria Wolbachia for metabolic and reproductive activities, doxycycline therapy is suggested by Hoerauf et al. [49] and Taylor et al. [50] for individual drug administration in bancroftian filariasis.
Several plants have been assessed for their antifilarial activity against Brugia sp and W. bancrofti. Lakshmi et al. [34] conducted a study on the antifilarial activity of a marine red alga, Botryocladia leptopoda (Rhodymeniaceae) against experimental infections with the rodent filarial parasites Litomosoides sigmodontis and Acanthocheilonema viteae and the human-pathogenic B. malayi. They show that the ethanolic crude extract and its hexane fraction reduce microfilarial levels and kill a significant proportion of the adult worms from L. sigmodontis and A. viteae. In the case of B. malayi, the macrofilaricidal efficacy was much less than that observed in the rodent parasites, but significant sterilization of the surviving female parasites was caused by the hexane and chloroform fractions of the ethanolic crude extract of B. leptopoda.
Fujimaki et al. [35] screened and provided detailed informations about 11 medicinal plants used in Guatemala for in vitro macrofilaricidal activity against B. pahangi. Among the 11 medicinal plants, the ethanolic extract of leaves of Neurolaena lobata (Asteraceae) showed the highest inhibitory activity against the motility of adult worms. The in vitro assay of the extract of N. lobata showed potential macro- and micro-filaricidal activities.
Misra et al. [36] reported the antifilarial activity in the extract of stem portions of the plant Lantana camara (Verbenaceae). They showed that the ethanolic crude extract of this plant administered orally to the rodent model Mastomys coucha killed 43.05% of adult B. malayi and sterilized 76% of surviving female worms. After fractionation of the ethanolic extract of this Verbenaceae, Misra et al. [36] were able to show that the chloroform fraction contains 34.5% adulticidal activity along with sterilization of 66% of female worms. They also demonstrated that the activity of L. camara depends on the rodent host. Thus, using Meriones unguiculatus (gerbil) rodents as host for B. malayi, lead to an antifilarial activity up to 80%, whereas at the same dosage, all adult worms were killed using M. coucha as host. The authors also reported that L. camara is efficient against the A. viteae, exerting strong microfilaricidal (95.04%) and sterilization (60.66%) efficacy with mild macrofilaricidal action. Two compounds from L. camara, oleanonic acid and oleanolic acid, isolated from hexane and chloroform fractions also showed an in vitro activity against B. malayi adults. Toxicity studies of L. camara in rats showed that all rats remained active and healthy throughout the study [34].
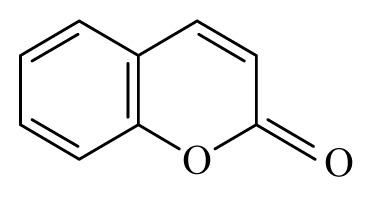
Sahare et al. [37,51] analyzed the in vitro effect of Butea monosperma (Fabaceae), Vitex negundo (Lamiaceae), Aegle marmelo (Rutaceae) and Ricinus communis (Euphorbiaceae) on the motility of B. malayi microfilariae. They reported that aqueous extract of B. monosperma leaves, ethanolic extract of V. negundo root and ethanolic extract A. marmelo leaves exhibited a 100% inhibition of motility of microfilariae at 100 ng/mL concentration as compared to controls. At the same concentration, methanolic extract of R. communis leaves and aqueous extract of B. monosperma roots showed no significant activity on the motility of B. malayi microfilaria. Using thin layer chromatography, Sahare et al. [51] were able to reveal the presence of alkaloids, saponin and flavonoids in the roots of V. negundo and coumarin in the leaves of A. marmelos.
Gaur et al. [39] studied the antifilarial activity of Caesalpinia bonducella (Caesalpiniaceae) seed kernel by using cotton rats Sigmodon hispidus and M. coucha that harbor the filarial parasites L. sigmodontis and B. malayi, respectively. Oral treatment of the rats with ethanolic extract of C. bonducella significantly reduced the number of microfilariae and macrofilariae of L. sigmodontis, with up to 96% filaricidal activities and 100% female sterilizing efficacy. Furthermore, Gaur et al. [39] reported that the butanol and the aqueous fraction exerted 73.7% and 90% reduction of microfilariae number, respectively, and 82.5% mortality in adult worms with 100% female sterilization for both fractions, while two hexane fractions lead to 64% and 95% macrofilaricidal activity and 100% worm sterilization. However, in the B. malayi/M. coucha model, the hexane fraction that demonstrated high macrofilaricidal activity on L. sigmodontis, only showed a gradual microfilaraemia reduction and caused 80% sterilization of female parasites. These results suggest that C. bonducella seed kernel extract and fractions show microfilaricidal, macrofilaricidal and female-sterilizing efficacy against L. sigmodontis and microfilaricidal and female-sterilizing efficacy against B. malayi. This indicates the potential of the plant in providing a lead for new antifilarial drug development [39].
Mathew et al. [40] investigated the in vitro activity of a methanolic extract of fruits of Trachyspermum ammi (Apiaceae) against adult bovine filarial Setaria digitata worms. Using high-performance liquid chromatography, they isolated the active fraction containing a phenolic monoterpene. After characterization, the active molecule showed macrofilaricidal activity and sterilizing efficacy against B. malayi in M. coucha. After extraction of leaves of Piper betle (Piperaceae) in methanol, Singh et al. [41] studied the antifilarial activities of the crude extract, the n-hexane and chloroform fractions of this Piperaceae using B. malayi. They demonstrated that the methanolic crude extract, the hexane and chloroform fractions of P. betle in vivo contain remarkable immunomodulatory properties in BALB/c mice and enhance both humoral as well as cell-mediated immune responses. Furthermore, in vivo antifilarial activity of the crude methanol extract and its active fractions was assessed using B. malayi, experimentally maintained in M. coucha, and the antifilarial results correlated with the immunomodulation potential of the plant. The crude extract and its active fractions suppressed microfilaraemia, showed moderate adulticidal activity and female worm sterilization.
Sharma et al. [38] selected extracts of roots of Vitex negundo, leaves of Butea monosperma, Aegle marmelos and Ricinus communis, to explore the possible involvement of any oxidative mechanisms in the observed antifilarial effect. They observe the presence of polyphenolic compounds and an increase in oxidative parameters in V. negundo, B. monosperma and A. marmelos and conclude that the observed antifilarial action of these herbal extracts might be associated with oxidative stress.
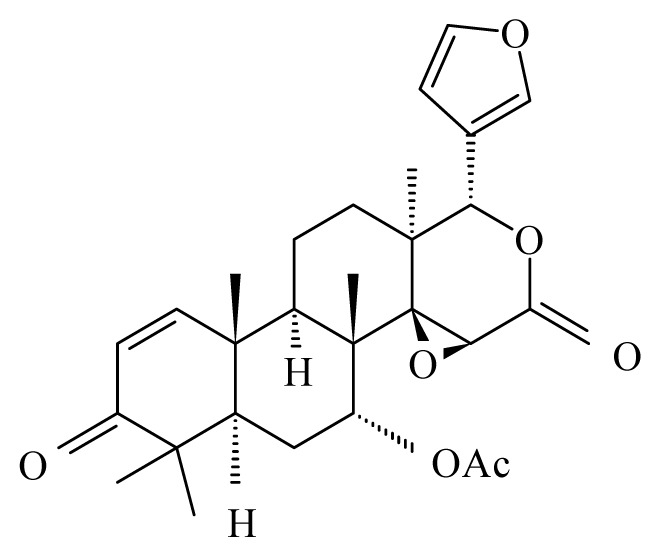
Misra et al. [42] studied the antifilarial activity of fruits of Xylocarpus granatum (Meliaceae) against B. malayi in vivo using the jird model M. unguiculatus and M. coucha. The in vitro effect of crude aqueous extract from different parts of X. granatum on filarial parasites was reported earlier by Zaridah et al. [43]. In their studies, Misra et al. [42] demonstrated that the crude aqueous ethanolic extract was active in vitro and demonstrated a 52.8% and 62.7% adulticidal and embryostatic effect on B. malayi in vivo, transplanted in the jird model (M. unguiculatus) and M. caucha. After fractionation (ethyl acetate fraction, n-butanol fraction, water-soluble fraction and water-insoluble fraction), only the ethyl acetate fraction revealed in vivo activity. Eight pure molecules were isolated from the active fraction, with the two compounds gedunin (IC50 = 0.239 μg/mL, SI = 889.1) and photogedunin (IC50 = 0.213 μg/mL, SI = 1231.4) at 5 × 100 mg/kg by subcutaneous route revealed adulticidal efficacy, with 80% and 70% mortality of the transplanted B. malayi.
Sashidhara et al. [44] isolated galactolipid from Bauhinia racemosa (Caesalpinaeceae). They showed that the leaves of B. racemosa exhibit promising in vitro and in vivo antifilarial activity against B. malayi which can be attributed to the presence of one pure compound, namely (2S)-1,2-di-O-linolenoyl-3-O-a-galactopyranosyl-(1/6)-O-b-galactopy ranosyl glycerol.
Azeez et al. [45] identified 10 spices and medicinal plants, namely coriander (Coriandrum sativum, Apiaceae), cassia (Cassia, Fabaceae), turmeric (Curcuma longa, Zingiberaceae), allspice (Pimenta dioica, Myrtaceae), cinnamon (Cinnamomum, Lauraceae), strychnous (Strychnos, Loganiaceae), lemongrass (Cymbopogon, Poaceae), garlic (Allium sativum, Amaryllidaceae), litsea (Litsea, Lauraceae) and vanilla (Vanilla, Orchidaceae) that contain phytochemicals, with the potential to inhibit the detoxification enzyme glutathione S-transferase (GST) from B. malayi. Molecular docking of these compounds, followed by in vitro GST assay, demonstrated that linalool, alpha-pinene, strychnine, vanillin, piperine, isoeugenol, curcumin, beta-caryophyllene, cinnamic acid, capsaicin, citronellol and geraniol effectively inhibit the B. malayi GST and therefore mark them as lead compounds.
Kushwaha et al. [46] reported that chemotypical variations in Withania somnifera (Solanaceae) lead to differentially modulated immune responses in BALB/c mice. Furthermore, the published findings [48] on the different immunomodulatory and immunotherapeutic potentials for the crude oil of Nigella sativa (Ranunculaceae) seeds and its active ingredients led Kushwaha et al. [47] to hypothesize that immunostimulation prior to pathogen invasion might provide protection against filarial infection. They demonstrated that crude root extracts of chemotypes NMITLI-101 (101R), NMITLI-118 (118R) and NMITLI-128 (128R) as well as withanolide withaferin A of W. somnifera offer protection to the rodent host M. coucha against infection of filarial parasite B. malayi. They also showed that treatment with W. somnifera extracts negatively affected not only larval establishment in the host but also led to a defective embryogenesis in female worms. This study revealed the potent immunoprophylactic property of W. somnifera.
Our online search on medicinal plants used against LF found 16 publications since 2002, where a total of 24 plant species, belonging to 20 different families, have been studied on. In these studies 24 pure compounds were isolated (Table 1).
2.2. Schistosomiasis
Schistosomiasis (also known as bilharzia, bilharziosis or snail fever) is a chronic parasitic disease caused by trematode parasites of the genus Schistosoma. Schistosomiasis is endemic in 77 countries, with higher prevalence in the tropical and the subtropical belt of the globe [52]. Is it estimated that about 800 million people are at risk of infection due to their exposure to contaminated water and 390–600 million people are infected with these parasites [52–55]. There are two main clinical forms of schistosomiasis: the intestinal form, which is caused by five different species (S. mansoni, S. japonicum, S. mekongi, S. guineensis, S. intercalatum) and the urogenital form, which is caused by S. haematobium [56].
The infection of Schistosoma sp. occurs when the larvae of the parasites are liberated by the infected snail (intermediary host), get in contact with the human host and subsequently penetrate the skin. Once inside the human hosts, the pathogens differentiate into schistosomules, which migrate via the bloodstream to the liver and develop into male and female mature forms. After mating, the worms migrate again and relocate at the mesenteric intestinal veins or the venous plexus of the urinary system. The females release the eggs, which are able to pass epithel of the blood vessels and reach the intestinal lumen, the bladder or urethra lumen in order to be expelled by feces or urine. Some of these eggs also remain in these tissues. The damage of blood vessels, together with immune reactions against the retained eggs are responsible for the clinical forms of schistosomiasis.
The first approaches towards controlling the infection were initiated in the beginning of the 20th century; however, there is still no consensus about the best strategy for controlling this disease [57]. Traditionally, the main prevention and control strategies are based on the control of the transmitting snails of the genus Biomphalaria, improving the sanitation conditions and the treatment of patients [56,58,59]. It is generally agreed that no individual method is sufficient to block transmission. Consequently, combinatory approaches are considered for interruption of the life cycle of Schistosoma sp. [54,60–62].
Currently, a main strategy to control schistosomiasis is based on the periodic treatment of people living at risk areas with anti-schistosomicidal drugs in order to reduce morbidity and transmission [63]. Moreover, it was recently suggested that schistosomiasis, as well as some other neglected diseases could be controlled by large scale treatment of the whole population with safe and efficient drugs in areas endemic for more than one parasitic disease [63].
2.2.1. Currently Available Drugs for the Treatment of Schistosomiasis
In former times, chemotherapy against Schistosoma sp. infection depended on antimonials. Due to serious side effect caused by these drugs, the use as schistosomicidal drugs was discontinued and currently the treatment mainly relies on three compounds: metrifonate, oxamniquine and praziquantel [64,65].
2.2.1.1. Metrifonate
Metrifonate (O,O-dimetil-2,2,2-tricloro-1-hydroxyethylphosphonate) was derived from an organophosphorus insecticide (dimethyl(2,2,2-trichloro-1-hydroxyethyl phosphonate). Metrifonate is a reversible inhibitor of the acetylcholinesterase, the enzyme that is responsible for acetylcholine catabolism. This inhibitory activity at low concentrations causes a selective paralysis of the parasite’s muscles, making it susceptible of being carried out by the bloodstream to the liver (in case of S. mansoni) or to the lungs (in case of S. haematobium). Since high concentrations of metrifonate are toxic for humans due to the diminished level of erythrocitary cholinesterase activity, the use is restricted to 3 doses. Recent results indicate that following the reduction of the concentration of the drug, S. mansoni is able to go back to the mesenteric veins within the intestine and re-establish the infection. However, S. haematobium adults remain in the lung and are not able to move back to the bladder or urethra veins and re-establish the infection [66,67]. Since metrifonate is only effective against the urogenital disease, the drug is only recommended by the WHO to treat the infection of S. haematobium [68].
2.2.1.2. Oxamniquine
Oxamniquine (1,2,3,4-tetrahydro-2-[isopropylamino]methyl(−)7-nitro-6-nitro-quinoline methanol) was reported to be substrate of a sulfotransferase that produces an ester which is able to react with nucleic acids and thereby interferes with replication and transcription processes. This drug also causes an increased motility and tegument damage, which primarily affects male worms. S. mansoni is more susceptible to the drug than other species. Under the effect of the drug, the male worm migrates into the liver, where the cellular immune response eliminates the pathogen. Deleterious changes are also observed in females. However, they are mostly due to the lack of male stimulation rather than to a direct effect of the drug. The absence of a human homologue renders oxamniquine effective with a low cyto-toxicity profile to human cells [69]. However, the emergence of resistant strains combined with high production cost and its restricted use for S. mansoni, makes the drug unusable for control campaigns.
2.2.1.3. Praziquantel
Praziquantel (2-(cyclohexylearbonil-1,2,3,6,7,11-hexylhydro-4H-pyrazino[2,1-a]isoquinil-4-one) is the only drug presently available for the treatment of the three human pathogenic species of Schistosoma (S. mansoni, S. haematobium and S. japonicum). The mechanism of action is not yet well established. It was proposed that the drug induces membrane alterations, producing a Ca2+ entry in the muscle cells and a paralysis in the contracted state. The paralyzed parasites are then carried out by the bloodstream [70–72]. The drug also causes tegumental damage allowing the host-immune cells to reach the interior of the parasite and to eliminate it [73,74]. The drug is mainly active against the adult forms of the parasite, limiting its use against the early stages of the infection [74,75]. Praziquantel is a safe and low cost drug, and it has been used in the last 20 years for control strategies and patients’ treatment in countries such as Brazil, Cambodia, China, Egypt, Morocco and Saudi Arabia [76–78]. However, due to its use at large scale control programs, the emergency of resistance and diminished efficacy were reported [68,79–89].
In search for new lead compounds, the richness of chemical structures of metabolites available in nature is also being exploited. The identification of natural products (mainly derived from plants) with schistosomicidal activity is a valuable strategy for obtaining lead compounds to combat this health problem [90–92].
2.2.2. Natural Products for the Control of Schistosomiasis
Vegetable oil extracts probably constitute the first anthelmintic products in traditional medicine. The variety of their application modes evidenced the ethno-pharmacological potential of these extracts as sources of active compounds. However, there are only a few studies which are focusing on the isolation, the identification, and eventually the validation of the active molecules within the plant extracts [93–98]. More recently, an initiative of research groups all over the world has been founded to set up plant extract screening programs [99–103]. A variety of compounds that reveal schistosomicidal activity have been identified in in vitro assays (Table 2). However, the extrapolation of in vitro results to the “in vivo world” is far away from being trivial. A good example for this is Vernodalin (Table 2), a sesquiterpene lactone with a promising in vitro activity but no in vivo activity [104]. Even after promising results are obtained in in vivo studies, the application of this drug in an endemic region must take other factors into consideration, such as the presence of other infectious pathogens. These could cause co-infections and thereby interfere with the respective treatment. For example, the antimalarial activity of Artemisinin and some derivatives has been exploited for more than 20 years [105]. More recently, the discovery and in vivo validation of schistosomicidal activity of artemisinin was established by observing a reduction of the parasitaemia in experimentally S. japonicum infected mice and the susceptibility of schistosomula to the drug [106–108]. Later, these results were also confirmed for S. mansoni. Within this study, an increased sensitivity of the females to the drug was also observed [109]. Although these results were promising, the use of artemisinin in regions endemic for malaria should be carefully assessed, since the application of this drug to control schistosomiasis could have the serious adverse side effect of increasing artemisinin drug resistance in malaria [110,111].
Table 2.
List of medicinal plants, products and active compounds known to have anti-schistosomal properties.
| Compounds/Substances | Origin | Schistosomicidal activities | Assays | References | |||
|---|---|---|---|---|---|---|---|
| in vitro | in vivo | Observations | Toxicity | Clinical | |||
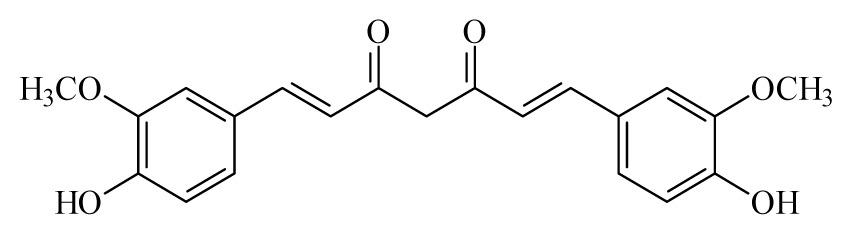 Curcumin |
Curcuma longa | S. mansoni adult—50 μM (100% mortality in male and female) | 400mg/Kg in mouse 1—43.5% mortality in male and 4.6% in female of S. mansoni) | Reduction in the oviposition Induced separation of males and females Reduction in the motor activity | nd | nd | [99,113] |
| Essencial oil (sesquiterpenes 57.20% and monoterpenes 42.13%) | Plectranthus neochilus | S. mansoni adult—LC50-value 89.65 mg/mL at 24 h LC50-value 58.18 mg/mL at 120 h | nd | Reduction in the oviposition Induced separation of males and females Reduction in the motor activity | Non-toxic in V79 cells2 (concentrations lower than 200 μg/mL) | nd | [101] |
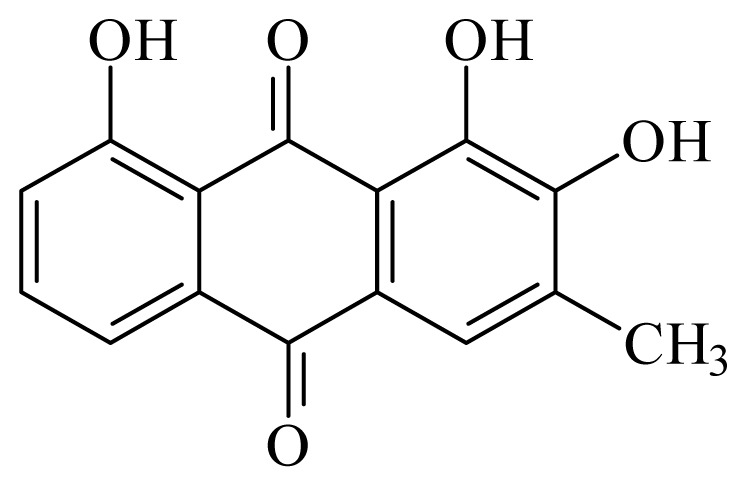 2-hydroxychrysophanol |
Hemerocallis fulva |
S. mansoni adult—50 μg/mL (35% mortality in male and female) Schistosomula—25 μg/mL (80% mortality) |
nd | nd | nd | nd | [114] |
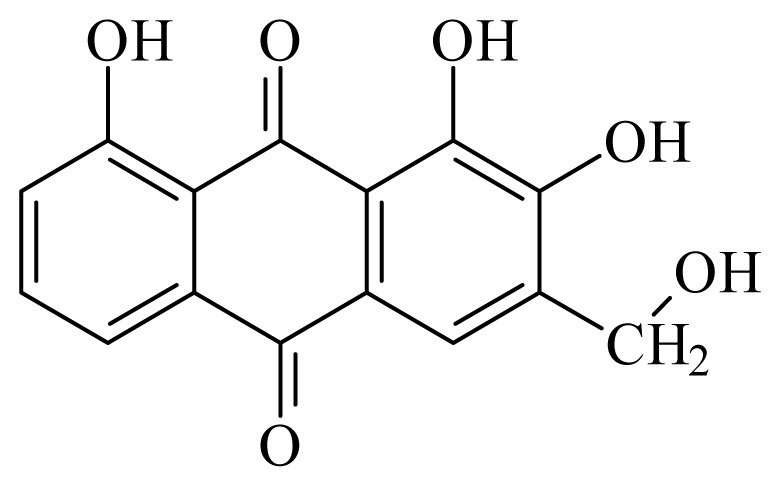 Kwanzoquinone E |
Hemerocallis fulva |
S. mansoni adult—50 μg/mL (55% mortality in male and female) Schistosomula—25 μg/mL (100% mortality) |
nd | nd | nd | nd | [114] |
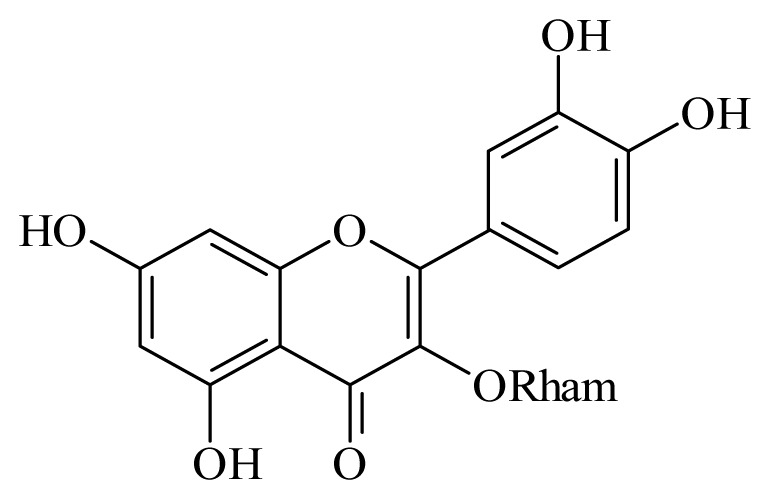 Quercetin 3-O-β-D-rhamnoside |
Schefflera vinosa | S. mansoni adult—100 μM (25% mortality in male and female) | nd | Reduction in the Motor activity | nd | nd | [98] |
| Methanol leaves extract | Cleome droserifolia | Nd | Doses of 0.31 g kg−1 for 21 days—32.46% reduction of S. mansoni in mice 3 | nd | nd | nd | [115] |
| Aqueous leaves extract | Clerodendrum umbellatum | Nd | 80 mg/kg in mice 4—100% mortality in S. mansoni | Reduction in the oviposition (75.49% released eggs in faeces) | nd | nd | [116] |
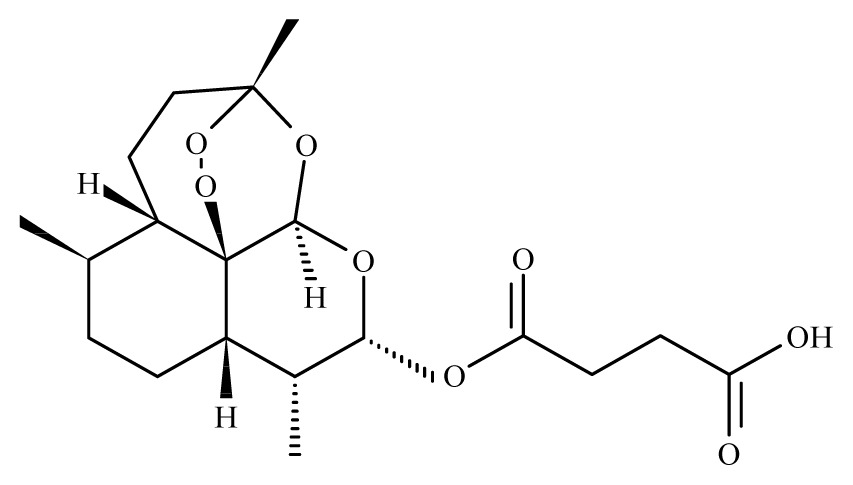 Artesunate |
Artemisia annua |
S. mekongi adult—40 μg/mL (100% mortality in male and female) S. mansoni adult—40 μg/mL (80% mortality in male and female) |
150 to 300 mg/kg in mice—67 and 77% mortality in male and female) | Reduction in the motor activity Reduction in the oviposition Tegumental disruption | nd | nd | [117,118] |
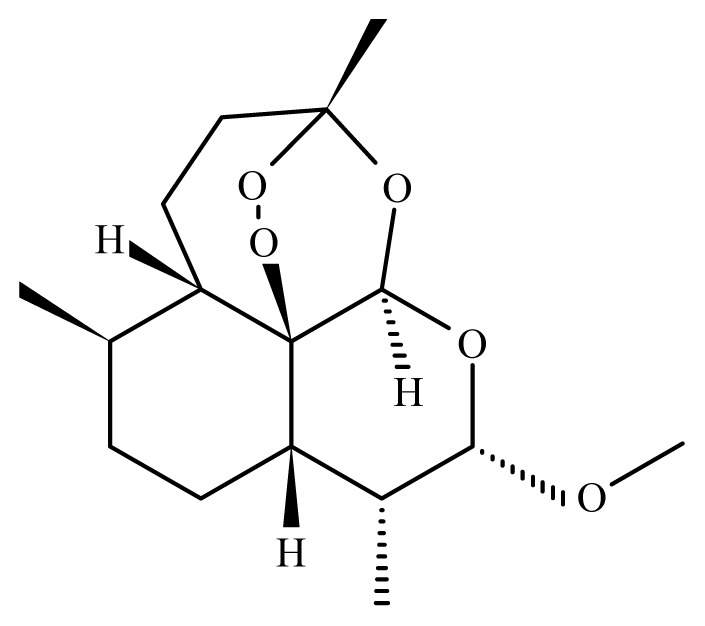 Artemether |
Artemisia annua | Nd | 50 mg/Kg in mice 5—reduction of S. mansoni female 100 mg/Kg in mice—61.5% mortality in S. mansoni female 150 to 300 mg/kg in mice—88 and 97% mortality in S. mansoni male and female 10 mg/Kg in rabbit 6—97% mortality in S. japonicum male and female 10 mg/Kg in dog 6—99.3% mortality in S. japonicum male and female |
Reduced liver and spleen weight of treated animals Reduction in the motor activity Tegumental disruption Alteration of the reproductive organs, ovarian volume reduction and depigmentation of the intestinal parasites portion |
nd | 30 mg/kg (2 oral doses) | [109,110, 118–120] |
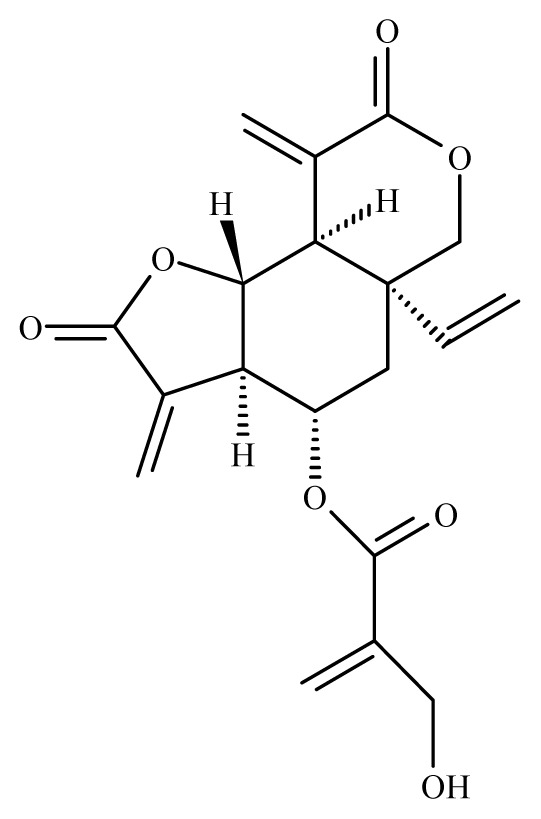 Vernodalin |
Vernonia amygdalina | S. japonicum adult —20 μg/mL (100% immobilization and oviposition) | 2.5 mg/kg in mice – no mortality S. japonicum | Inhibition of the oviposition Inhibition of the motor activity |
Toxic in KB, P-388 L-1210 cells7 at 120 mg/Kg |
nd | [104] |
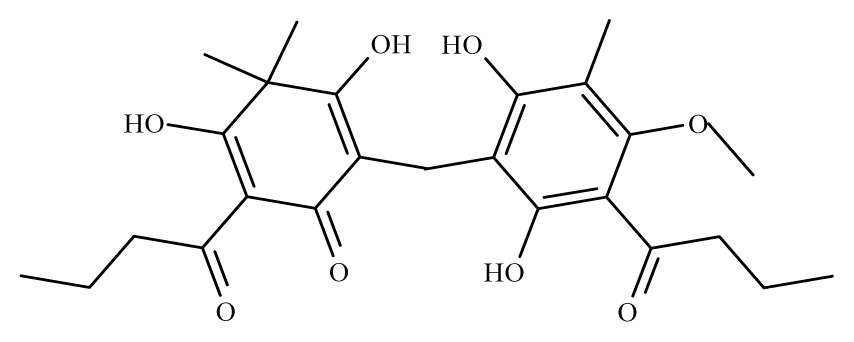 Aspidin |
Dryopteris spp. | S. mansoni adult—25 μM (100% mortality in male and female) | nd | Reduction in the motor activity Tegumental alterations |
nd | nd | [100] |
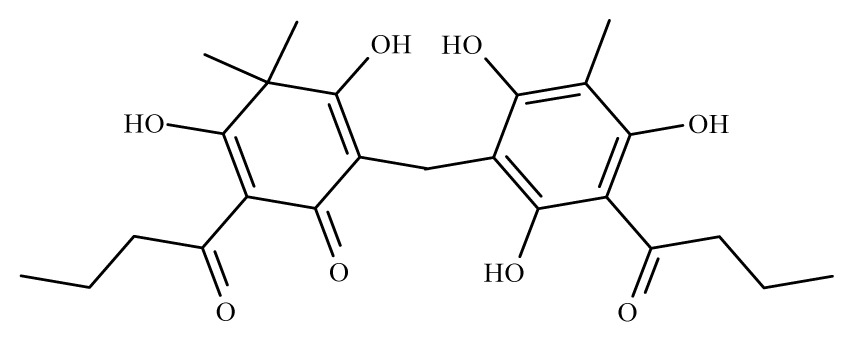 Favaspidic acid |
Dryopteris spp. | S. mansoni adult—50 μM (100% mortality in male and female) | nd | Reduction in motor the activity Tegumental alterations |
nd | nd | [100] |
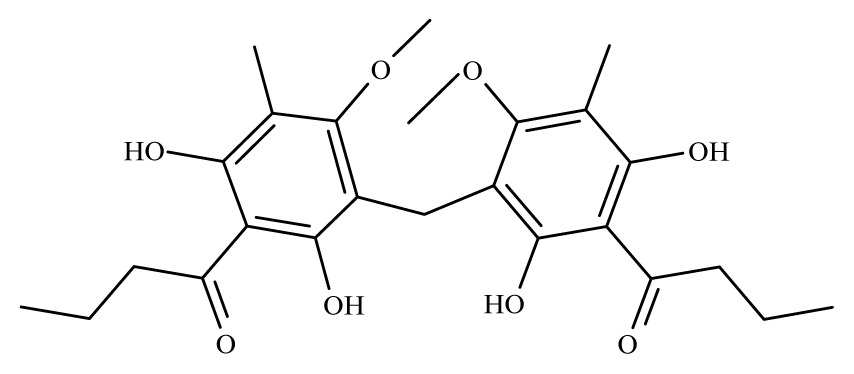 Methylene-bis-aspidinol |
Dryopteris spp. | S. mansoni adult—100 μM (100% mortality in male and female) | nd | Reduction in the motor activity | nd | nd | [100] |
 Desaspidin |
Dryopteris spp. | S. mansoni adult—25 μM (100% mortality in male and female) | nd | Reduction in the motor activity | nd | nd | [100] |
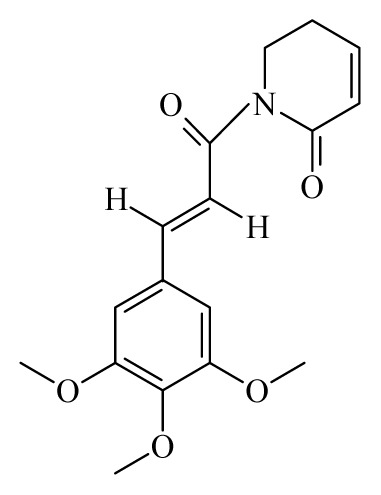 Piplartine |
Piper tuberculatum |
S. mansoni adult—15.8 μM (100% mortality in male and female) Schistosomula—5 μM (100% mortality) |
nd | Reduction in the motor activity Reduction in the oviposition |
Non-toxic in Vero cell 8 at 31.5 μM | nd | [92,103] |
| Methanol leaves extract | Baccharis trimera | S. mansoni—130 μg/mL (100% mortality in male, female and schistosomula | nd | Reduction in the motor activity Tegumental alterations |
Non-toxic in human keratinocytes cell line at 250 μg/mL | nd | [121] |
| Avocado/soybean unsaponifiables | Persea americana | Nd | 300 mg/kg—3 oral doses in mice—30% S. mansoni mortality | Reduction in the motor activity Tegumental alterations Reduction in the oviposition | nd | nd | [122] |
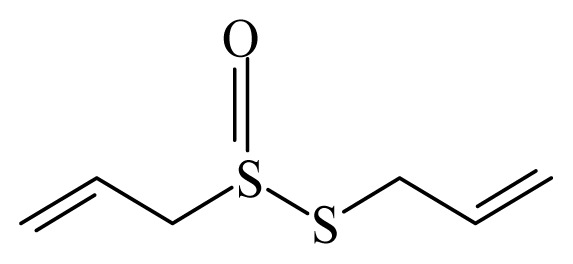 Allicin |
Allium sativum | S. mansoni adult—20 mg/mL—no mortality | nd | 5mg/mL tegumental alterations 10mg/mL changes in tubercles, loss or changes in the spines; 15 and 20mg/mL tegumental disruption (vesicle and ulceration) |
nd | nd | [123] |
Male CD1 albino—16 injections and total dose of 400 mg/kg;
Chinese hamster lung fibroblasts;
Swiss albino;
Male Balb/c—daily doses for 14 days;
Female Balb/c—doses administered at day 0, 20 and 60 post infection; Rats - intramuscular dose;
Rabbits and dogs treated 7, 14, 21, 28, 35 days post infection;
KB-Cell line of human oral carcinoma, P388-murine leukemia, L1210-Mouse lymphocytic leukemia cells;
African green monkey kidney fibroblast.
2.2.3. Mode of Action and Molecular Targets in Schistosoma
Synthetic drugs and natural compounds with schistosomicidal activity in general lead to alterations in their behavior (disruption of mating of males and females, reduction in reproductive fitness), in the morphology or constitution of the protective tegument, or in the muscle activity of the parasite.
2.2.3.1. Compounds that Disrupt Mating
Compounds such as curcumin, artesunate, artemeter, artemisina, vernodalin, piplartina, as well as avocado and soybean unsaponifiable oils, essential oils from Plectranthus neochilus and extracts from Calyptridium umbellatum cause the separation of males and females (Table 2). The mated state is a fundamental process of the parasite viability inside the human host and for establishing the infection. Only when the parasites are mated (the female is held in a groove within the male body, the gynecophoric canal) the sexual maturation and egg production occurs. The induction of separation of males and females reduces or even arrests the release of eggs, which is a relevant factor in the hepatic pathology and the transmission of the disease [112].
2.2.3.2. Compounds Acting on the Tegument Structure or Composition
The tegument in Schistosoma sp. is involved in several vital functions such as nutrient absorption, secretion and by acting as a physical barrier against components of the humoral and cellular host-immune system [124,125]. It is constantly being renewed. Due to these facts and the previous observations that drugs affecting the tegument make the parasites more sensitive to the host immune response, make it a relevant target for drug discovery. Thus, drugs leading to tegument alterations such as vacuolization or observable surface modifications, like peeling, loss of spines or changes in protuberances are considered to be suitable for in in vivo testing (Table 2) [88,120]. However, beside visible modifications of the tegument, additional criteria should be used to evaluate the potential schistosomicidal activity of a drug or extract, since these alterations not always affect the survival of the parasite [123]. Due to the morphological and biochemical differences between male and female parasites, the efficiency of the drug can be gender specific. For example, in in vivo studies with S. mansoni it was observed that at a concentration of 400 mg/kg curcumine was 10-fold more active against male than female worms [99,113]. On the other hand, it was reported that artemeter was more active against females of S. mansoni [119,126]. Compounds targeting Schistosoma sp. in a sex specific manner are effective for treating the infected individuals, improving their clinical outcome and their ability of transmit the disease through the diminution of the released eggs to the environment.
2.2.3.3. Compounds Acting on the Parasite Nervous System
The nervous system of Schistosoma sp., as well as those of other helminthes, has been considered a relevant target for drug discovery. Some observable changes such as parasite motility were associated to interfere with the activity of neurotransmitters or neuromodulators like serotonin, dopamine, acetylcholine, epinephrine, glutamate and a variety of neuropeptides [65,127–130]. In this context, Hillman et al. [131] used a fluorescent analogue of acetylcholine and validated the parasite cholinergic receptors as a drug target for S. mansoni. For these in vitro experiments the worms’ motor activity is a main parameter; however, although drugs causing significant motor alterations are usually evaluated as hits, their effect is not always related to the drug action.
3. Trypanosomatids
Human pathogens of the genus Leishmania and Trypanosoma are restricted to several species causing Chagas disease (Trypanosoma cruzi), sleeping sickness (T. brucei) and leishmaniosis (a group of disease caused by parasites of the genus Leishmania). The natural transmission of the first two infections is limited to America and Africa, respectively, while the leishmaniosis is distributed around the world’s tropical and subtropical belt. Plants have been used in the traditional medicine to treat infections caused by these human pathogens [132–137]. Pathogenic trypanosomatids are digenetic organisms with complex life cycles involving an insect vector and a mammalian host (including humans). To make it more complex—with exception of Trypanosoma brucei—all other pathogenic trypanosomatids have at least one intracellular stage in the mammalian host, which plays a major role in the establishment of the chronic infection and/or in the pathogenesis. Thus, successful therapeutic compounds should be accessible to the respective intracellular compartment in order to target the parasite. As mentioned, T. brucei does not have an intracellular cycle in the mammalian host. This fact does not make the challenge easier: in the advanced stages of the disease, the parasites reach the central nervous system (CNS) and as a consequence the successful drug should ideally be able to pass the blood-brain barrier.
As the insect stages of trypanosomatids can be easily cultured in axenic media, most of tests for trypanocidal and leishmanicidal activities have been carried out in these systems. However, the evaluation of compounds on insect stages presents severe limitations due to the fact that the effect cannot be extrapolated to the mammalian stages. Thus, last-generation high throughput screening platforms were developed to evaluate the effect of compounds (particularly those against T. cruzi and Leishmania spp.) in mammalian infected cells. Until today, most extracts and plant derived compounds have been evaluated in axenic systems, some of them in in vitro or ex vivo infections systems and only a limited number of them have been evaluated in vivo (Table 3). It is worth mentioning that—with some exceptions—only the mode of action of cubebin and its derivatives has thus far been elucidated [138].
Table 3.
List of medicinal plants, products and active compounds known to have anti-trypanosomal properties.
| Compounds/Substances | Origin | Trypanocidal activities | Assays | References | |||
|---|---|---|---|---|---|---|---|
| in vitro | in vivo | Observations | Toxicity | Clinical | |||
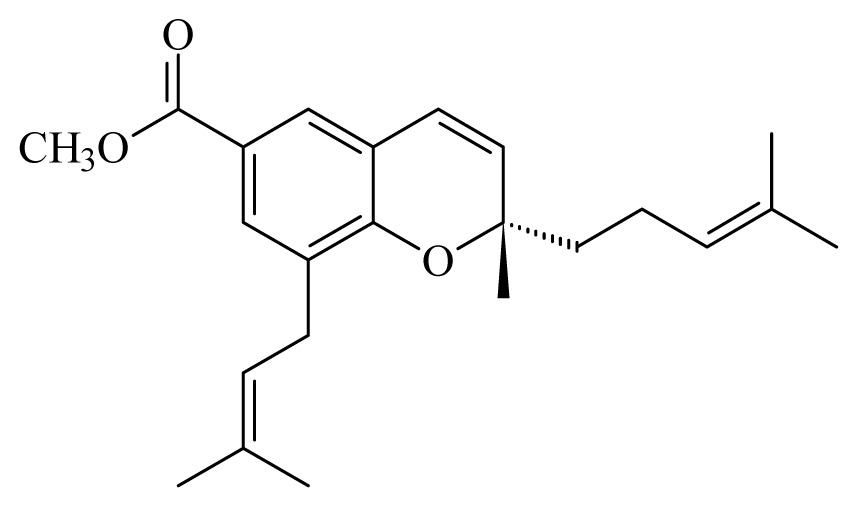 [(2S)-methyl-2-methyl-8-(3-methylbut-2′-enyl)-2-(4-methylpent-3-enyl)-2Hchromene-6-carboxylate] |
Piper1 | T. cruzi epimastigote—IC50-value 2.82 μM | nd | nd | nd | nd | [143] |
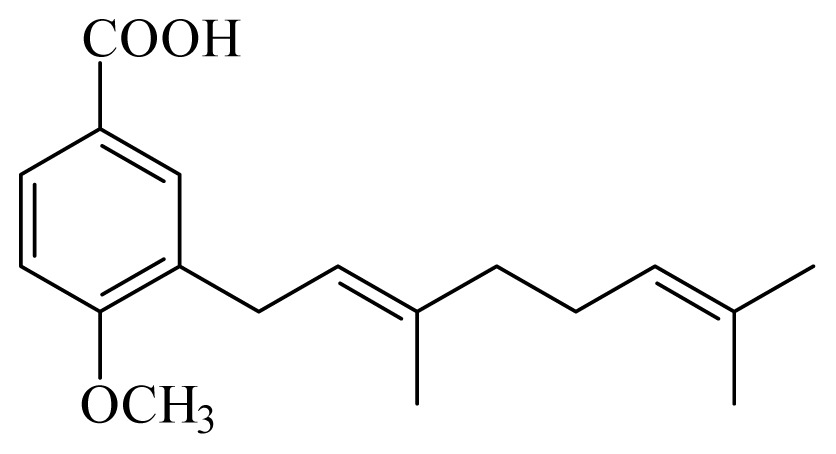 3-(3,7-dimethyl-2,6-octadienyl)-4-methoxy-benzoic acid |
Piper aduncum | L. braziliensis promastigote—IC50-value 6.5 μg/mL | nd | nd | nd | nd | [144] |
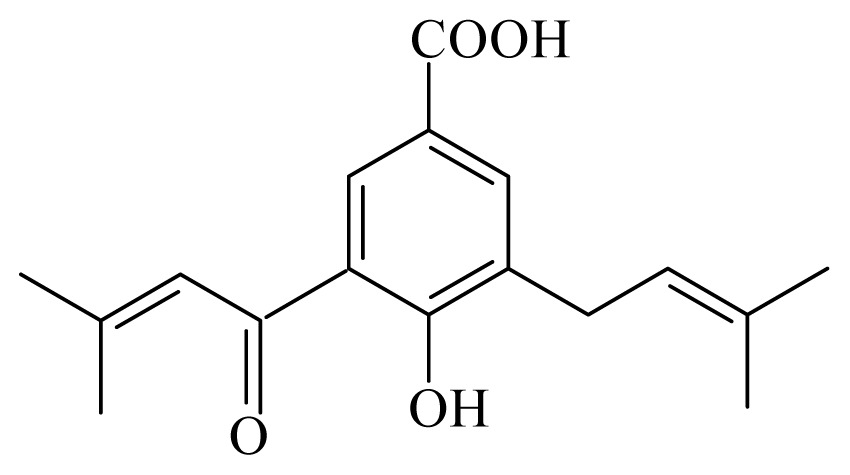 4-hydroxy-3-(3-methyl-1-oxo-2-butenyl)-5-(3-methyl-2-butenyl)benzoic acid |
Piper aduncum | T. cruzi epimastigote—IC50 16.5 μg/mL | nd | nd | nd | nd | [144] |
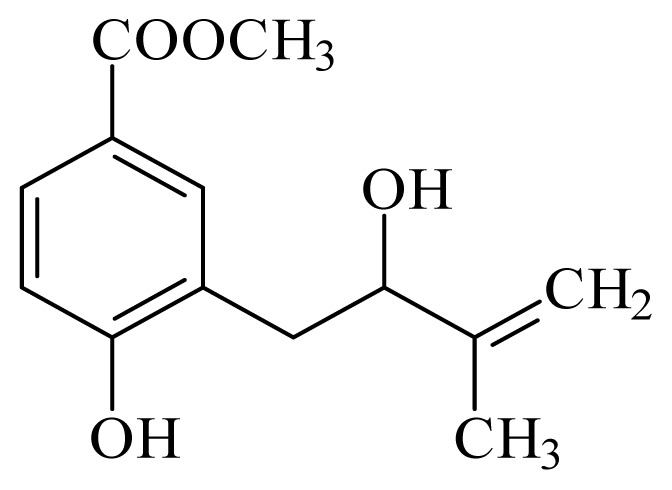 methyl 4-hydroxy-3-(2-hydroxy-3-methyl-3-butenyl)benzoate |
Piper glabratum | T. cruzi epimastigote—IC50-value 15.6 μg/mL | nd | nd | nd | nd | [145] |
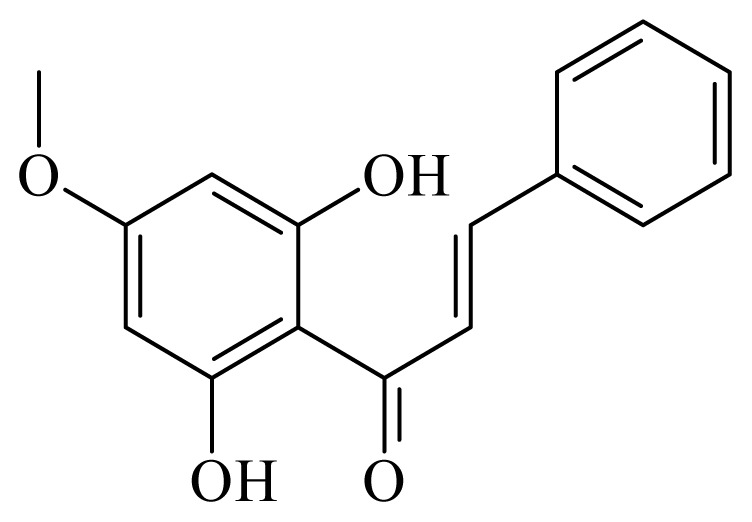 2,6-dihydroxi-4-metoxichalcone |
Piper aduncum | L. amazonensis promastigote—IC50-value 0.5 μg/mL amastigotes—IC50-value 24 μg/mL | nd | nd | nd | nd | [146] |
 dihydrochalcone |
Piper elongatum | L. braziliensis promastigote—IC50-value 2.98 μg/mL | nd | nd | nd | nd | [147] |
 Cubebin |
Piper cueba | L. donovani promastigotes—IC50-value 28 μg/mL | 100.0 mg/kg, i.p. in hamster infected with L. donovani | nd | nd | nd | [148] |
 Conocarpan |
Piper regnellii | T. cruzi epimastigote—IC50-value 8.0 μg/mL | nd | nd | nd | nd | [149] |
 Eupomatenoid |
Piper regnellii | T. cruzi epimastigote—IC50-value 7.0 μg/mL | nd | nd | Non-toxic in Vero cells (CC50 250 μg/mL) | nd | [149] |
 Grandisin |
Piper solmsianum | T. cruzi trypomastigote—IC50-value 8.74 μg/mL | nd | nd | nd | nd | [150] |
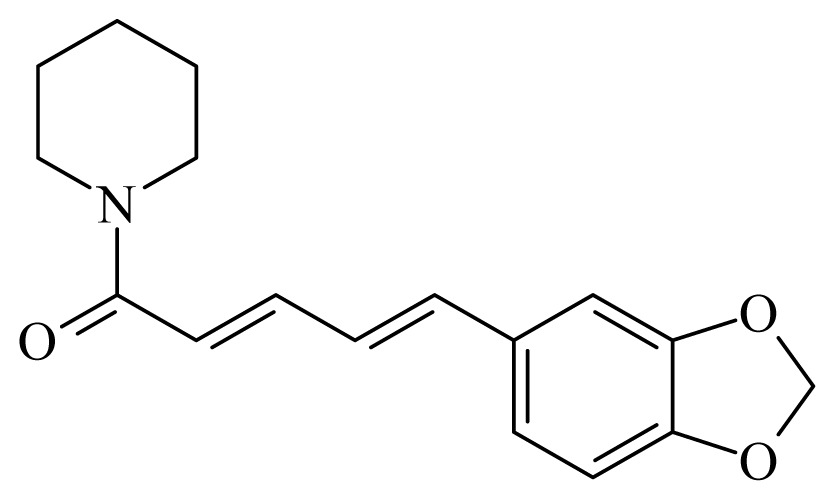 Piperine |
Piper | T. cruzi epimastigote—IC50-value 7.36 μM amastigote—IC50-value 4.91 μM | nd | nd | nd | nd | [151] |
 Piplartine |
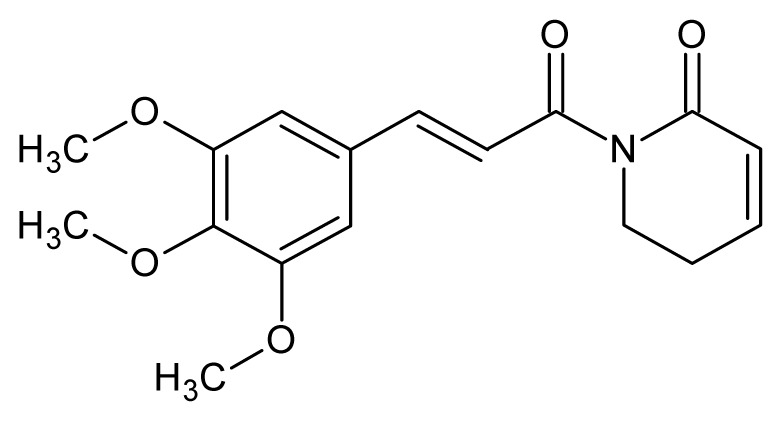
Piper tuberculatum Piper retrofractum | T. cruzi epimastigote—IC50-value 10.5 μM L. donovani promastigotes—IC50-value 7.5 μg/mL | 30 mg/kg ip. in hamster infected with L. donovani | nd | nd | nd | [148,152] |
 prenylated hydroquinone [1,4-dihydroxy-2-(3′,7′-dimethyl-1′-oxo-2′-E,6′-octadienyl)benzene |
Piper crassinervium | T. cruzi epimastigote—IC50-value 6.10 μg/mL | nd | nd | nd | nd | [153] |
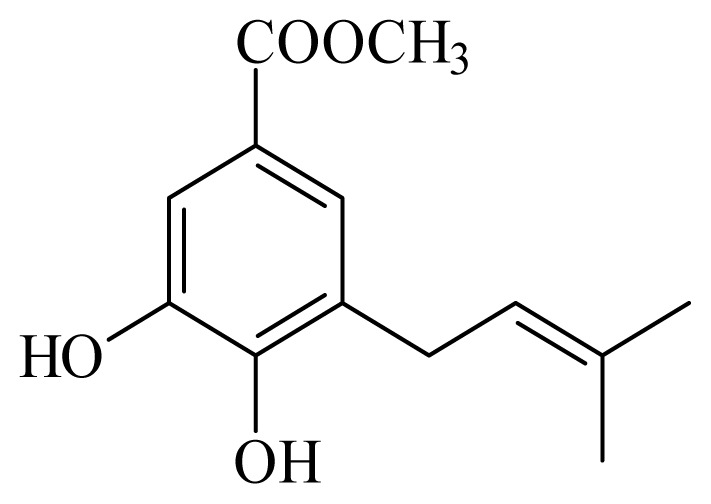 methyl 3,4-dihydroxy-5-(3′-methyl-2′-butenyl)benzoate |
Piper glabratum | L. braziliensis, L. amazonensis and L. donovani—IC50-value 13.8–18.5 μg/mL) | nd | nd | nd | nd | [145] |
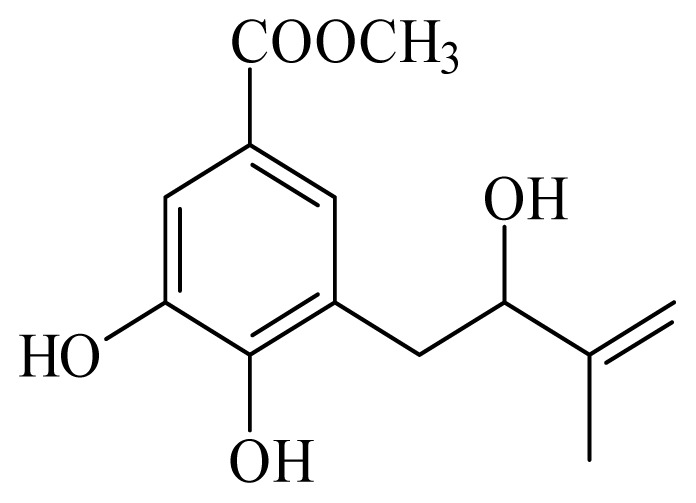 methyl 3,4-dihydroxy-5-(2-hydroxy-3-methylbutenyl)benzoate |
Piper glabratum | T cruzi epimestigote—IC50-value 16.4 μg/mL | nd | nd | nd | nd | [145] |
 methyl 4-hydroxy-3-(2-hydroxy-3-methyl-3-butenyl)benzoate |
Piper glabratum | T cruzi epimestigote—IC50-value 15.6 μg/mL | nd | nd | nd | nd | [145] |
 (15-acetoxy-8_-[(2-methylbutyryloxy)]-14-oxo-4,5-cis-acanthospermolide) |
Acanthospermum hispidum | T. brucei brucei—IC50-value 2.45 μM L. mexicana Mexicana —IC50-value 0.94 μM | nd | nd | nd | nd | [154] |
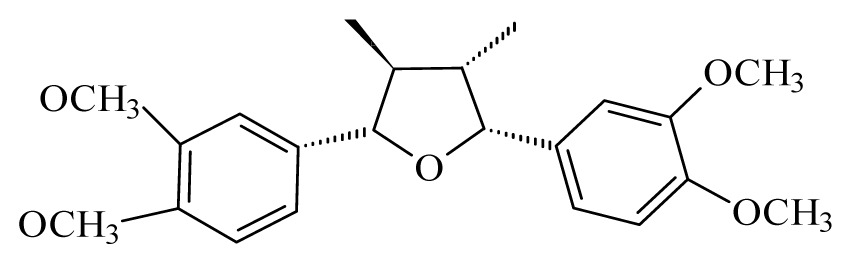 Veraguensin |
Nectandra megapotamica | L.donovani promastigotes—IC50-value 18 μg/mL and IC90-value 36 μg/mL | nd | nd | Non-toxic in Vero cells5 in 10 μg/ml | nd | [91] |
| Methanol extract from stem bark | Acacia nilotica | na | 200 mg/kg body weight in mice infected with T. brucei brucei | Clear the parasites from circulation within 6 days of treatment | na | na | [136] |
| Methanol stem bark extract | Bombax buonopozense | nd | 300 mg/kg body weight in mice infected with T. brucei brucei | Clear the parasites from circulation within 7 days of treatment | nd | nd | [136] |
| Dichloromethane bark extract | Warburgia salutaris | T. brucei brucei—IC50 -value 10.68 μg/mL | nd | nd | nd | nd | [155] |
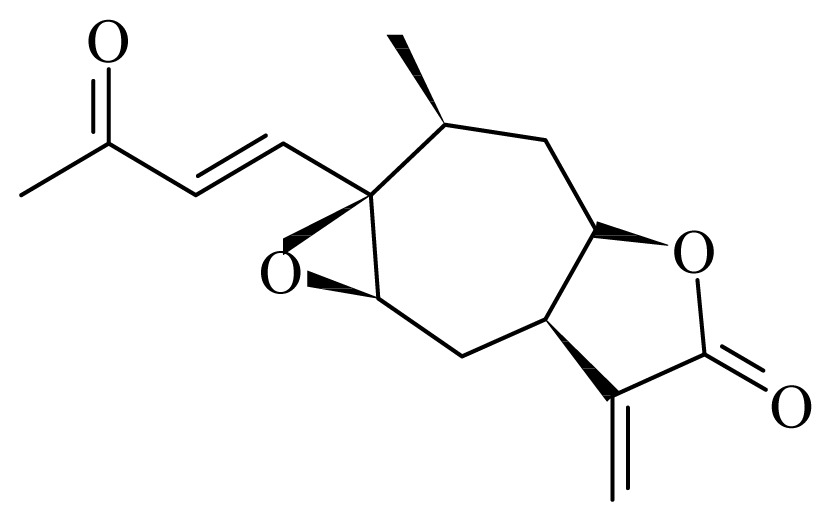 8-epixanthatin 1β,5β-epoxide |
Xanthium brasilicum Vell | T. brucei—IC50 0.09 μg/mL T. cruzi—IC50-value 2.95 μg/L L. donovani—IC50 0.16 μg/mL | nd | nd | nd | nd | [142] |
Synthetic compound derived from chromenes isolated from Piper aduncum and Piper gaudichaudianum;
Intraperitoneally, twice daily for 3 days;
per os, twice daily for 3 days;
Balb/c infected with L. amazonensis and treated for 28 days;
African green monkey kidney fibroblast.
A variety of compounds belonging to the alkaloid family presents trypanocidal activity. Most of them are phenols, therpenoids of quinines. However, none of them can be considered as promising drug candidates yet, due to the limited in vivo data (Table 3). Interestingly, most of these active compounds against Trypanosoma sp. and Leishmania spp. are derived from plant species of the genus Piper (belonging to the family Piperaceae). The variety of the metabolites derived from this species is well documented. Compounds of the primary and secondary metabolism of Piperaceae have been isolated and several of them present a well-documented biological activity within several organisms including S. mansoni as mentioned above [92,103,139–141]. Compounds of the plant families Asteraceae and Lauraceae reveal also activity against trypanosomatids. However, the quantity of these compounds reported up to now, is reduced in comparison to those found in Piperaceae [91,142]. The fact that most of anti-trypanosomatid activities were found in members of a single family suggests that the search for these activities was somehow restricted. As plants can be conceived as “natural organic synthesis laboratories”, it is highly emphasized to test and isolate more extracts from plants against trypanosomatids in order to obtain new molecules to fight these neglected diseases.
4. Summary
The most prevalent families revealing anti-filarial activity were Fabaceae represented by five species, followed by four species of Euphrobiaceae and finally Apiaceae, Rosaceae, Annonaceae and Lauraceae which were indicated by two species. The majority of the active compounds isolated from these species revealed an aromatic structure. Among these compounds, gallic, gentisic and ellagic acids were active against O. ochengi. Oleanonic, oleanolic acids, gedunin, photogedunin, coumarin and withanolide were interferring with B. malayi. It would be important to test these compounds on other filariae to establish a common and effective antifilaricide especially in areas where co-infection occurs.
Among the compounds revealing anti-schistosomal activity, a mode of action has been proposed only for artemisinin and its derivatives. The interference of these drugs with the hemoglobin metabolism is a common feature between Schistosoma and Plasmodium. Both organisms degrade the host hemoglobin into their constituting amino acids, and the free hem is eliminated by the formation of hemozoin, a less reactive iron containing molecule. Free hem is known to be involved in the generation of a variety of different reactive oxygen species and thereby causing oxidative stress. It was proposed that the hem reacts with artemisinin which affects the proliferation of the parasite [117,156].
In general, little is known about the mode of action of natural compounds leading to anti-schistosomal as well as anti trypanosomal activities. More information on the molecular mechanism involved in the action of natural compounds is necessary to predict side effects and analyze the probability of the emergence of resistant strains.
Presently, the set-up of high throughput screening facilities allows a more efficient and rapid identification of interesting schistosomicidal compounds. In this sense, previous studies on the characterization and inhibition of relevant parasite proteins like acetylcholinesterase [67,157], cysteine protease [158] serine protease [159] and glutamatergic receptors in adult S. mansoni [160] are relevant to the development of new possible methods towards the identification and validation of new targets. For example, Abdulla et al. [158] described the viability of applying a medium-throughput phenotypic screening of adult worms and schistosomula in vitro with a library of known drugs to identify putative schistosomicidal drugs. Interestingly, this screening could also be used to investigate the schistosomidal activity of plant derived extracts, products and active compounds.
Acknowledgements
The authors would like to thank the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP, grants 2011/50631-1 to A.M.S. and 2009/54325-2 to C.W.), the Instituto Nacional de Biologia Estrutural e Química Medicinal em Doenças Infecciosas (INBEQMeDI) and the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, grant 470272/2011-2 to A.M.S.) as well as the Deutsche Forschungsgemeinschaft (DFG, grant LI 793/5-1 to E.L.) within the framework of the Cameroonian-German Cooperation Project (cameroon.uni-muenster.de) for financial support. D.N. is a fellow of the Alexander von Humboldt Foundation.
References
- 1.Schmidt T.J., Khalid S.A., Romanha A.J., Alves T.M., Biavatti M.W., Brun R., Da Costa F.B., de Castro S.L., Ferreira V.F., de Lacerda M.V., et al. The potential of secondary metabolites from plants as drugs or leads against protozoan neglected diseases—Part II. Curr. Med. Chem. 2012;19:2176–2228. [PubMed] [Google Scholar]
- 2.Schmidt T.J., Khalid S.A., Romanha A.J., Alves T.M., Biavatti M.W., Brun R., Da Costa F.B., de Castro S.L., Ferreira V.F., de Lacerda M.V., et al. The potential of secondary metabolites from plants as drugs or leads against protozoan neglected diseases—Part I. Curr. Med. Chem. 2012;19:2128–2175. doi: 10.2174/092986712800229023. [DOI] [PubMed] [Google Scholar]
- 3.Christen P.C. Muriel plants as a source of therapeutic and health products. CHIMIA Int. J. Chem. 2012;66:320–323. doi: 10.2533/chimia.2012.320. [DOI] [PubMed] [Google Scholar]
- 4.Ioset J.R. Natural products for neglected diseases: A review. Curr. Organ. Chem. 2008;12:643–666. [Google Scholar]
- 5.Cavalier-Smith T. A revised six-kingdom system of life. Biol. Rev. Camb. Philos. Soc. 1998;73:203–266. doi: 10.1017/s0006323198005167. [DOI] [PubMed] [Google Scholar]
- 6.Lambshead P.J.D. Recent developments in marine benthic biodiversity research. Oceanis. 1993;19:5–24. [Google Scholar]
- 7.Hugot J.P., Baujard P., Morand S. Biodiversity in helminths and nematodes as a field of study: An overview. Nematology. 2001;3:199–208. [Google Scholar]
- 8.World Health Organization. Report of 512 a WHO Expert Committee on Onchocerciasis Control. WHO; Geneva, Switzerland: 1995. Onchocerciasis and Its Control; pp. 1–104. (Geneva; WHO Technical 513 Report Series). [PubMed] [Google Scholar]
- 9.Osei-Atweneboana M.Y., Eng J.K., Boakye D.A., Gyapong J.O., Prichard R.K. Prevalence and intensity of Onchocerca volvulus infection and efficacy of ivermectin in endemic communities in Ghana: A two-phase epidemiological study. Lancet. 2007;369:2021–2029. doi: 10.1016/S0140-6736(07)60942-8. [DOI] [PubMed] [Google Scholar]
- 10.Wagman R.J.E. The New Complete Medical and Health Encyclopedia. Vol. 4. Lexicon Publication Inc.; New York, NY, USA: 1982. p. 1310. [Google Scholar]
- 11.Navaratnam V. Recent Advances in the Chemotheraphy of Lymphatic Filariasis. Proceeding of 34th SEAMEO TROPMED Regional Seminar: Current Status of Filariasis in Southeast Asia; Kuala Lumpur, Malaysia. 26–27 June 1992. [Google Scholar]
- 12.World Health Organization. Wkly. Epidemiol. Rec. 2009. Global programme to eliminate lymphatic filariasis; pp. 437–444. [PubMed] [Google Scholar]
- 13.Gyapong J.O., Kumaraswami V., Biswas G., Ottesen E.A. Treatment strategies underpinning the global programme to eliminate lymphatic filariasis. Expert Opin. Pharmacother. 2005;6:179–200. doi: 10.1517/14656566.6.2.179. [DOI] [PubMed] [Google Scholar]
- 14.Bockarie M.J., Taylor M.J., Gyapong J.O. Current practices in the management of lymphatic filariasis. Expert Rev. Anti-Infect. Ther. 2009;7:595–605. doi: 10.1586/eri.09.36. [DOI] [PubMed] [Google Scholar]
- 15.Liang J.L., King J.D., Ichimori K., Handzel T., Pa'au M., Lammie P.J. Impact of five annual rounds of mass drug administration with diethylcarbamazine and albendazole on Wuchereria bancrofti infection in American Samoa. Am. J. Trop. Med. Hyg. 2008;78:924–928. [PubMed] [Google Scholar]
- 16.The Carter Center. [accessed on 29 December 2012]. Available online: http://www.cartercenter.org/health/lf/index.html.
- 17.The Center of Disease Control and Prevention. [accessed on 29 December 2012]. Available online: http://www.cartercenter.org/health/lf/index.html.
- 18.Winnen M., Plaisier A.P., Alley E.S., Nagelkerke N.J., van Oortmarssen G., Boatin B.A., Habbema J.D. Can ivermectin mass treatments eliminate onchocerciasis in Africa? Bull World Health Organ. 2002;80:384–391. [PMC free article] [PubMed] [Google Scholar]
- 19.Kaplan R.M. Drug resistance in nematodes of veterinary importance: A status report. Trends Parasitol. 2004;20:477–481. doi: 10.1016/j.pt.2004.08.001. [DOI] [PubMed] [Google Scholar]
- 20.Howell S.B., Burke J.M., Miller J.E., Terrill T.H., Valencia E., Williams M.J., Williamson L.H., Zajac A.M., Kaplan R.M. Prevalence of anthelmintic resistance on sheep and goat farms in the southeastern United States. J. Am. Vet. Med. Assoc. 2008;233:1913–1919. doi: 10.2460/javma.233.12.1913. [DOI] [PubMed] [Google Scholar]
- 21.Moussala M., Fobi G., Zogo O., Hiag B.L.A., Bengono G., McMoli T.E. Survenue d’hémorragies rétiniennes lors du traitement de l’onchocercose par l’invermectine chez une patiente co-infectée par la loase. J. Français d’Ophtalmologie. 2004;27:63–66. doi: 10.1016/s0181-5512(04)96094-9. [DOI] [PubMed] [Google Scholar]
- 22.Melo A.C.F.L., Reis I.F., Bevilaqua C.M.L., Vieira L.S., Echevarria F.M., Melo L.M. Nematodeos resistentes a anti-helminticos em rebanhos de ovinos ecaprinos do estado do Ceará, Brasil. Ciencia Rural. 2003;33:339–344. [Google Scholar]
- 23.Sullivan K., Shealy C.N. Complete Natural Home Remedies. Element Books Limited; Shafterburg, UK: 1997. [Google Scholar]
- 24.Robinson M.M., Zhang X. Traditional Medicines: Global Situation, Issues and Challenges. WHO; Geneva, Switzerland: 2011. [Google Scholar]
- 25.Achukwi M.D., Harnett W., Renz A. Onchocerca ochengi transmission dynamics and the correlation of O. ochengi microfilaria density in cattle with the transmission potential. Vet. Res. 2000;31:611–621. doi: 10.1051/vetres:2000144. [DOI] [PubMed] [Google Scholar]
- 26.Nyasse B., Ngantchou I., Nono J.J., Schneider B. Antifilarial activity in vitro of polycarpol and 3-O-acetyl aleuritolic acid from cameroonian medicinal plants against Onchocerca gutturosa. Nat. Prod. Res. 2006;20:391–397. doi: 10.1080/14786410600661377. [DOI] [PubMed] [Google Scholar]
- 27.Cho-Ngwa F., Abongwa M., Ngemenya M.N., Nyongbela K.D. Selective activity of extracts of Margaritaria discoidea and Homalium africanum on Onchocerca ochengi. BMC Complement. Altern. Med. 2010;10:62. doi: 10.1186/1472-6882-10-62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ndjonka D., Agyare C., Luersen K., Djafsia B., Achukwi D., Nukenine E.N., Hensel A., Liebau E. In vitro activity of Cameroonian and Ghanaian medicinal plants on parasitic (Onchocerca ochengi) and free-living (Caenorhabditis elegans) nematodes. J. Helminthol. 2011;85:304–312. doi: 10.1017/S0022149X10000635. [DOI] [PubMed] [Google Scholar]
- 29.Ndjonka D., Ajonina-Ekoti I., Djafsia B., Luersen K., Abladam E., Liebau E. Anogeissus leiocarpus extract on the parasite nematode Onchocerca ochengi and on drug resistant mutant strains of the free-living nematode Caenorhabditis elegans. Vet. Parasitol. 2012;190:136–142. doi: 10.1016/j.vetpar.2012.05.027. [DOI] [PubMed] [Google Scholar]
- 30.Smith R.A., Pontiggia L., Waterman C., Lichtenwalner M., Wasserman J. Comparison of motility, recovery, and methyl-thiazolyl-tetrazolium reduction assays for use in screening plant products for anthelmintic activity. Parasitol. Res. 2009;105:1339–1343. doi: 10.1007/s00436-009-1560-4. [DOI] [PubMed] [Google Scholar]
- 31.Hoste H., Brunet S., Paolini V., Bahuaud D., Chauveau S., Fouraste I., Lefrileux Y. Compared in vitro anthelmintic effects of eight tannin-rich plants browsed by goats in the southern part of France. Option Méditerrenéennes. 2009:431–436. [Google Scholar]
- 32.Thomsen H., Reider K., Franke K., Wessjohann L.A., Keiser J., Dagne E., Arnold N. Characterization of constituents and anthelmintic properties of Hagenia abyssinica. Sci. Pharm. 2012;80:433–446. doi: 10.3797/scipharm.1109-04. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Katiki L.M., Ferreira J.F., Gonzalez J.M., Zajac A.M., Lindsay D.S., Chagas A.C., Amarante A.F. Anthelmintic effect of plant extracts containing condensed and hydrolyzable tannins on Caenorhabditis elegans, and their antioxidant capacity. Vet. Parasitol. 2012;18:218–227. doi: 10.1016/j.vetpar.2012.09.030. [DOI] [PubMed] [Google Scholar]
- 34.Lakshmi V., Kumar R., Gupta P., Varshney V., Srivastava M.N., Dikshit M., Murthy P.K., Misra-Bhattacharya S. The antifilarial activity of a marine red alga, Botryocladia leptopoda, against experimental infections with animal and human filariae. Parasitol. Res. 2004;93:468–474. doi: 10.1007/s00436-004-1159-8. [DOI] [PubMed] [Google Scholar]
- 35.Fujimaki Y., Kamachi T., Yanagi T., Caceres A., Maki J., Aoki Y. Macrofilaricidal and microfilaricidal effects of Neurolaena lobata, a Guatemalan medicinal plant, on Brugia pahangi. J. Helminthol. 2005;79:23–28. doi: 10.1079/joh2004262. [DOI] [PubMed] [Google Scholar]
- 36.Misra N., Sharma M., Raj K., Dangi A., Srivastava S., Misra-Bhattacharya S. Chemical constituents and antifilarial activity of Lantana camara against human lymphatic filariid Brugia malayi and rodent filariid Acanthocheilonema viteae maintained in rodent hosts. Parasitol. Res. 2007;100:439–448. doi: 10.1007/s00436-006-0312-y. [DOI] [PubMed] [Google Scholar]
- 37.Sahare K.N., Anandhraman V., Meshram V.G., Meshram S.U., Reddy M.V., Tumane P.M., Goswami K. Anti-microfilarial activity of methanolic extract of Vitex negundo and Aegle marmelos and their phytochemical analysis. Indian J. Exp. Biol. 2008;46:128–131. [PubMed] [Google Scholar]
- 38.Sharma R.D., Veerpathran A.R., Dakshinamoorthy G., Sahare K.N., Goswami K., Reddy M.V. Possible implication of oxidative stress in anti filarial effect of certain traditionally used medicinal plants in vitro against Brugia malayi microfilariae. Pharmacogn. Res. 2010;2:350–354. doi: 10.4103/0974-8490.75453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Gaur R.L., Sahoo M.K., Dixit S., Fatma N., Rastogi S., Kulshreshtha D.K., Chatterjee R.K., Murthy P.K. Antifilarial activity of Caesalpinia bonducella against experimental filarial infections. Indian J. Med. Res. 2008;128:65–70. [PubMed] [Google Scholar]
- 40.Mathew N., Misra-Bhattacharya S., Perumal V., Muthuswamy K. Antifilarial lead molecules isolated from Trachyspermum ammi. Molecules. 2008;13:2156–2168. doi: 10.3390/molecules13092156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Singh M., Shakya S., Soni V.K., Dangi A., Kumar N., Bhattacharya S.M. The n-hexane and chloroform fractions of Piper betle L. trigger different arms of immune responses in BALB/c mice and exhibit antifilarial activity against human lymphatic filarid Brugia malayi. Int. Immunopharmacol. 2009;9:716–728. doi: 10.1016/j.intimp.2009.02.012. [DOI] [PubMed] [Google Scholar]
- 42.Misra S., Verma M., Mishra S.K., Srivastava S., Lakshmi V., Misra-Bhattacharya S. Gedunin and photogedunin of Xylocarpus granatum possess antifilarial activity against human lymphatic filarial parasite Brugia malayi in experimental rodent host. Parasitol. Res. 2011;109:1351–1360. doi: 10.1007/s00436-011-2380-x. [DOI] [PubMed] [Google Scholar]
- 43.Zaridah M.Z., Idid S.Z., Omar A.W., Khozirah S. In vitro antifilarial effects of three plant species against adult worms of subperiodic Brugia malayi. J. Ethnopharmacol. 2001;78:79–84. doi: 10.1016/s0378-8741(01)00286-0. [DOI] [PubMed] [Google Scholar]
- 44.Sashidhara K.V., Singh S.P., Misra S., Gupta J., Misra-Bhattacharya S. Galactolipids from Bauhinia racemosa as a new class of antifilarial agents against human lymphatic filarial parasite, Brugia malayi. Eur. J. Med. Chem. 2012;50:230–235. doi: 10.1016/j.ejmech.2012.01.057. [DOI] [PubMed] [Google Scholar]
- 45.Azeez S., Babu R.O., Aykkal R., Narayanan R. Virtual screening and in vitro assay of potential drug like inhibitors from spices against glutathione-S-transferase of filarial nematodes. J. Mol. Model. 2012;18:151–163. doi: 10.1007/s00894-011-1035-2. [DOI] [PubMed] [Google Scholar]
- 46.Kushwaha S., Roy S., Maity R., Mallick A., Soni V.K., Singh P.K., Chaurasiya N.D., Sangwan R.S., Misra-Bhattacharya S., Mandal C. Chemotypical variations in Withania somnifera lead to differentially modulated immune response in BALB/c mice. Vaccine. 2012;30:1083–1093. doi: 10.1016/j.vaccine.2011.12.031. [DOI] [PubMed] [Google Scholar]
- 47.Kushwaha S., Soni V.K., Singh P.K., Bano N., Kumar A., Sangwan R.S., Misra-Bhattacharya S. Withania somnifera chemotypes NMITLI 101R, NMITLI 118R, NMITLI 128R and withaferin A protect Mastomys coucha from Brugia malayi infection. Parasite Immunol. 2012;34:199–209. doi: 10.1111/j.1365-3024.2012.01352.x. [DOI] [PubMed] [Google Scholar]
- 48.Salem M.L. Immunomodulatory and therapeutic properties of the Nigella sativa L. seed. Int. Immunopharmacol. 2005;5:1749–1770. doi: 10.1016/j.intimp.2005.06.008. [DOI] [PubMed] [Google Scholar]
- 49.Hoerauf A., Mand S., Fischer K., Kruppa T., Marfo-Debrekyei Y., Debrah A.Y., Pfarr K.M., Adjei O., Buttner D.W. Doxycycline as a novel strategy against bancroftian filariasis-depletion of Wolbachia endosymbionts from Wuchereria bancrofti and stop of microfilaria production. Med. Microbiol. Immunol. 2003;192:211–216. doi: 10.1007/s00430-002-0174-6. [DOI] [PubMed] [Google Scholar]
- 50.Taylor M.J., Makunde W.H., McGarry H.F., Turner J.D., Mand S., Hoerauf A. Macrofilaricidal activity after doxycycline treatment of Wuchereria bancrofti: A double-blind, randomised placebo-controlled trial. Lancet. 2005;365:2116–2121. doi: 10.1016/S0140-6736(05)66591-9. [DOI] [PubMed] [Google Scholar]
- 51.Sahare K.N., Anandharaman V., Meshram V.G., Meshram S.U., Gajalakshmi D., Goswami K., Reddy M.V. In vitro effect of four herbal plants on the motility of Brugia malayi microfilariae. Indian J. Med. Res. 2008;127:467–471. [PubMed] [Google Scholar]
- 52.World Health Organization. Schistosomiasis: Population requiring preventive chemotherapy and number of people treated in 2010. Wkly Epidemiol. Rec. 2012;4:37–44. [PubMed] [Google Scholar]
- 53.King C.H. Parasites and poverty: The case of schistosomiasis. Acta Trop. 2010;113:95–104. doi: 10.1016/j.actatropica.2009.11.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Gray D.J., McManus D.P., Li Y., Williams G.M., Bergquist R., Ross A.G. Schistosomiasis elimination: Lessons from the past guide the future. Lancet Infect. Dis. 2010;10:733–736. doi: 10.1016/S1473-3099(10)70099-2. [DOI] [PubMed] [Google Scholar]
- 55.Siddiqui A.A., Siddiqui B.A., Ganley-Leal L. Schistosomiasis vaccines. Hum. Vaccin. 2011;7:1192–1197. doi: 10.4161/hv.7.11.17017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Gryseels B., Polman K., Clerinx J., Kestens L. Human schistosomiasis. Lancet. 2006;368:1106–1118. doi: 10.1016/S0140-6736(06)69440-3. [DOI] [PubMed] [Google Scholar]
- 57.Rollinson D., Knopp S., Levitz S., Stothard J.R., Tchuente L.A., Garba A., Mohammed K.A., Schur N., Person B., Colley D.G., et al. Time to set the agenda for schistosomiasis elimination. Acta Trop. 2012 doi: 10.1016/j.actatropica.2012.04.013. in press. [DOI] [PubMed] [Google Scholar]
- 58.Fenwick A.W., J.P. Schistosomiasis: Challenges for control, treatment and drug resistance. Curr. Opin. Infect. Dis. 2006;19:577–582. doi: 10.1097/01.qco.0000247591.13671.6a. [DOI] [PubMed] [Google Scholar]
- 59.Steinmann P., Keiser J., Bos R., Tanner M., Utzinger J. Schistosomiasis and water resources development: Systematic review, meta-analysis, and estimates of people at risk. Lancet Infect. Dis. 2006;6:411–425. doi: 10.1016/S1473-3099(06)70521-7. [DOI] [PubMed] [Google Scholar]
- 60.Fenwick A., Savioli L. Schistosomiasis elimination. Lancet Infect. Dis. 2011;11:346–347. doi: 10.1016/S1473-3099(11)70110-4. [DOI] [PubMed] [Google Scholar]
- 61.Rapado L.N., Nakano E., Ohlweiler F.P., Kato M.J., Yamaguchi L.F., Pereira C.A., Kawano T. Molluscicidal and ovicidal activities of plant extracts of the Piperaceae on Biomphalaria glabrata (Say, 1818) J. Helminthol. 2011;85:66–72. doi: 10.1017/S0022149X10000258. [DOI] [PubMed] [Google Scholar]
- 62.Chimbari M.J. Enhancing schistosomiasis control strategy for zimbabwe: Building on past experiences. J. Parasitol. Res. 2012;2012 doi: 10.1155/2012/353768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.World Health Organization. Working to Overcome the Global Impact of Neglected Tropical Diseases—First WHO Report on Neglected Tropical Diseases. WHO Press; Geneva, Switzerland: 2010. p. 184. [Google Scholar]
- 64.Bueding E., Fisher J. Factors affecting the inhibition of phosphofructokinase activity of Schistosoma mansoni by trivalent organic antimonials. Biochem. Pharmacol. 1966;15:1197–1211. doi: 10.1016/0006-2952(66)90285-1. [DOI] [PubMed] [Google Scholar]
- 65.Cioli D., Pica-Mattoccia L., Archer S. Antischistosomal drugs: Past, present and future? Pharmacol. Ther. 1995;68:35–85. doi: 10.1016/0163-7258(95)00026-7. [DOI] [PubMed] [Google Scholar]
- 66.Denham DA, H.R. The effect of metrifonate in vitro on Schistosoma haematobium and S. mansoni adults. Trans. R. Soc. Trop. Med. Hyg. 1971;65:695–696. doi: 10.1016/0035-9203(71)90070-8. [DOI] [PubMed] [Google Scholar]
- 67.Holmstedt B., Nordgren I., Sandoz M., Sundwall A. Metrifonate. Summary of toxicological and pharmacological information available. Arch. Toxicol. 1978;41:3–29. doi: 10.1007/BF00351766. [DOI] [PubMed] [Google Scholar]
- 68.Cioli D. Chemotherapy of schistosomiasis: An update. Parasitol. Today. 1998;14:418–422. doi: 10.1016/s0169-4758(98)01323-4. [DOI] [PubMed] [Google Scholar]
- 69.Pica-Mattoccia L., Cioli D. Studies on the mode of action of oxamniquine and related schistosomicidal drugs. Am. J. Trop. Med. Hyg. 1985;34:112–118. doi: 10.4269/ajtmh.1985.34.112. [DOI] [PubMed] [Google Scholar]
- 70.Kohn A.B., Anderson P.A., Roberts-Misterly J.M., Greenberg R.M. Schistosome calcium channel beta subunits. Unusual modulatory effects and potential role in the action of the antischistosomal drug praziquantel. J. Biol. Chem. 2001;276:36873–36876. doi: 10.1074/jbc.C100273200. [DOI] [PubMed] [Google Scholar]
- 71.Jeziorski M.C., Greenberg R.M. Voltage-gated calcium channel subunits from platyhelminths: Potential role in praziquantel action. Int. J. Parasitol. 2006;36:625–632. doi: 10.1016/j.ijpara.2006.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Salvador-Recatala V., Greenberg R.M. Calcium channels of schistosomes: Unresolved questions and unexpected answers. Wiley Interdiscip. Rev. 2012;1:85–93. doi: 10.1002/wmts.19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Day T.A.B.J., Pax R.A. Praziquantel: The enigmatic antiparasitic. Parasitol. Today. 1992;8:342–344. doi: 10.1016/0169-4758(92)90070-i. [DOI] [PubMed] [Google Scholar]
- 74.Utzinger J., Keiser J., Shuhua X., Tanner M., Singer B.H. Combination chemotherapy of schistosomiasis in laboratory studies and clinical trials. Antimicrob. Agents Chemother. 2003;47:1487–1495. doi: 10.1128/AAC.47.5.1487-1495.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Sabah A.A., Fletcher C., Webbe G., Doenhoff M.J. Schistosoma mansoni: Chemotherapy of infections of different ages. Exp. Parasitol. 1986;61:294–303. doi: 10.1016/0014-4894(86)90184-0. [DOI] [PubMed] [Google Scholar]
- 76.Fenwick A., Savioli L., Engels D., Robert Bergquist N., Todd M.H. Drugs for the control of parasitic diseases: Current status and development in schistosomiasis. Trends Parasitol. 2003;19:509–515. doi: 10.1016/j.pt.2003.09.005. [DOI] [PubMed] [Google Scholar]
- 77.Doenhoff M.J., Hagan P., Cioli D., Southgate V., Pica-Mattoccia L., Botros S., Coles G., Tchuem Tchuente L.A., Mbaye A., Engels D. Praziquantel: Its use in control of schistosomiasis in sub-Saharan Africa and current research needs. Parasitology. 2009;136:1825–1835. doi: 10.1017/S0031182009000493. [DOI] [PubMed] [Google Scholar]
- 78.Utzinger J., N’Goran E.K., Caffrey C.R., Keiser J. From innovation to application: Social-ecological context, diagnostics, drugs and integrated control of schistosomiasis. Acta Trop. 2011;120:S121–S137. doi: 10.1016/j.actatropica.2010.08.020. [DOI] [PubMed] [Google Scholar]
- 79.Bennett J.L., Day T., Liang F.T., Ismail M., Farghaly A. The development of resistance to anthelmintics: A perspective with an emphasis on the antischistosomal drug praziquantel. Exp. Parasitol. 1997;87:260–267. doi: 10.1006/expr.1997.4229. [DOI] [PubMed] [Google Scholar]
- 80.Kusel J., Hagan P. Praziquantel—its use, cost and possible development of resistance. Parasitol. Today. 1999;15:352–354. doi: 10.1016/s0169-4758(99)01510-0. [DOI] [PubMed] [Google Scholar]
- 81.Ismail M., Botros S., Metwally A., William S., Farghally A., Tao L.F., Day T.A., Bennett J.L. Resistance to praziquantel: Direct evidence from Schistosoma mansoni isolated from Egyptian villagers. Am. J. Trop. Med. Hyg. 1999;60:932–935. doi: 10.4269/ajtmh.1999.60.932. [DOI] [PubMed] [Google Scholar]
- 82.Cioli D. Praziquantel: Is there real resistance and are there alternatives? Curr. Opin. Infect. Dis. 2000;13:659–663. doi: 10.1097/00001432-200012000-00014. [DOI] [PubMed] [Google Scholar]
- 83.Doenhoff M.J., Kusel J.R., Coles G.C., Cioli D. Resistance of Schistosoma mansoni to praziquantel: Is there a problem? Trans. R. Soc. Trop. Med. Hyg. 2002;96:465–469. doi: 10.1016/s0035-9203(02)90405-0. [DOI] [PubMed] [Google Scholar]
- 84.Doenhoff M., Kimani G., Cioli D. Praziquantel and the control of schistosomiasis. Parasitol. Today. 2000;16:364–366. doi: 10.1016/s0169-4758(00)01749-x. [DOI] [PubMed] [Google Scholar]
- 85.Gryseels B., Mbaye A., de Vlas S.J., Stelma F.F., Guisse F., van Lieshout L., Faye D., Diop M., Ly A., Tchuem-Tchuente L.A., et al. Are poor responses to praziquantel for the treatment of Schistosoma mansoni infections in Senegal due to resistance? An overview of the evidence. Trop. Med. Int. Health. 2001;6:864–873. doi: 10.1046/j.1365-3156.2001.00811.x. [DOI] [PubMed] [Google Scholar]
- 86.Caffrey C.R. Chemotherapy of schistosomiasis: Present and future. Curr. Opin. Chem. Biol. 2007;11:433–439. doi: 10.1016/j.cbpa.2007.05.031. [DOI] [PubMed] [Google Scholar]
- 87.Sabra A.N., Botros S.S. Response of Schistosoma mansoni isolates having different drug sensitivity to praziquantel over several life cycle passages with and without therapeutic pressure. J. Parasitol. 2008;94:537–541. doi: 10.1645/GE-1297.1. [DOI] [PubMed] [Google Scholar]
- 88.Doenhoff M.J., Cioli D., Utzinger J. Praziquantel: Mechanisms of action, resistance and new derivatives for schistosomiasis. Curr. Opin. Infect. Dis. 2008;21:659–667. doi: 10.1097/QCO.0b013e328318978f. [DOI] [PubMed] [Google Scholar]
- 89.Barakat R., El Morshedy H. Efficacy of two praziquantel treatments among primary school children in an area of high Schistosoma mansoni endemicity, Nile Delta, Egypt. Parasitology. 2011;138:440–446. doi: 10.1017/S003118201000154X. [DOI] [PubMed] [Google Scholar]
- 90.Kayser O., Kiderlen A.F., Croft S.L. Natural products as antiparasitic drugs. Parasitol. Res. 2003;90:S55–S62. doi: 10.1007/s00436-002-0768-3. [DOI] [PubMed] [Google Scholar]
- 91.Da Silva Filho A.A., Costa E.S., Cunha W.R., Silva M.L., Nanayakkara N.P., Bastos J.K. In vitro antileishmanial and antimalarial activities of tetrahydrofuran lignans isolated from Nectandra megapotamica (Lauraceae) Phytother. Res. 2008;22:1307–1310. doi: 10.1002/ptr.2486. [DOI] [PubMed] [Google Scholar]
- 92.Moraes J., Nascimento C., Lopes P.O., Nakano E., Yamaguchi L.F., Kato M.J., Kawano T. Schistosoma mansoni: In vitro schistosomicidal activity of piplartine. Exp. Parasitol. 2011;127:357–364. doi: 10.1016/j.exppara.2010.08.021. [DOI] [PubMed] [Google Scholar]
- 93.Ndamba J., Chidimu M.G., Zimba M., Gomo E., Munjoma M. An investigation of the schistosomiasis transmission status in Harare. Cent. Afr. J. Med. 1994;40:337–342. [PubMed] [Google Scholar]
- 94.Clark T.E., Appleton C.C., Kvalsvig J.D. Schistosomiasis and the use of indigenous plant molluscicides: A rural South African perspective. Acta Trop. 1997;66:93–107. doi: 10.1016/s0001-706x(97)00672-4. [DOI] [PubMed] [Google Scholar]
- 95.Sparg S.G., van Staden J., Jager A.K. Efficiency of traditionally used South African plants against schistosomiasis. J. Ethnopharmacol. 2000;73:209–214. doi: 10.1016/s0378-8741(00)00310-x. [DOI] [PubMed] [Google Scholar]
- 96.Molgaard P., Nielsen S.B., Rasmussen D.E., Drummond R.B., Makaza N., Andreassen J. Anthelmintic screening of Zimbabwean plants traditionally used against schistosomiasis. J. Ethnopharmacol. 2001;74:257–264. doi: 10.1016/s0378-8741(00)00377-9. [DOI] [PubMed] [Google Scholar]
- 97.Bah S., Diallo D., Dembele S., Paulsen B.S. Ethnopharmacological survey of plants used for the treatment of schistosomiasis in Niono District, Mali. J. Ethnopharmacol. 2006;105:387–399. doi: 10.1016/j.jep.2005.11.026. [DOI] [PubMed] [Google Scholar]
- 98.Cunha N.L.U.C., Cintra L.S., Souza H.C., Peixoto J.A., Silva C.P., Magalhães L.G., Groppo M., Rodrigues V., Filho A.A.S., Silva M.L.A., et al. In Vitro schistosomicidal activity of some brazilian cerrado species and their isolated compounds. Evidence-Based Complement. Alter. Med. 2012;2012 doi: 10.1155/2012/173614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Magalhaes L.G., Machado C.B., Morais E.R., Moreira E.B., Soares C.S., da Silva S.H., da Silva Filho A.A., Rodrigues V. In vitro schistosomicidal activity of curcumin against Schistosoma mansoni adult worms. Parasitol. Res. 2009;104:1197–1201. doi: 10.1007/s00436-008-1311-y. [DOI] [PubMed] [Google Scholar]
- 100.Magalhaes L.G., Kapadia G.J., da Silva Tonuci L.R., Caixeta S.C., Parreira N.A., Rodrigues V., da Silva Filho A.A. In vitro schistosomicidal effects of some phloroglucinol derivatives from Dryopteris species against Schistosoma mansoni adult worms. Parasitol. Res. 2010;106:395–401. doi: 10.1007/s00436-009-1674-8. [DOI] [PubMed] [Google Scholar]
- 101.Caixeta S.C., Melo N.I., Wakabayashi K.A.L., Aguiar G.P., Aguiar D.P., Mantovani A.L.L., Alves J.M., Oliveira P.F., Tavares D.C., Groppo M.G., et al. Chemical composition and in vitro schistosomicidal activity of the essential oil of Plectranthus neochilus grown in southeast Brazil. Chem. Biodivers. 2011;8:2147–2157. doi: 10.1002/cbdv.201100167. [DOI] [PubMed] [Google Scholar]
- 102.Pereira A.C., Magalhaes L.G., Goncalves U.O., Luz P.P., Moraes A.C., Rodrigues V., da Matta Guedes P.M., da Silva Filho A.A., Cunha W.R., Bastos J.K., et al. Schistosomicidal and trypanocidal structure-activity relationships for (+/−)-licarin A and its (−)- and (+)-enantiomers. Phytochemistry. 2011;72:1424–1430. doi: 10.1016/j.phytochem.2011.04.007. [DOI] [PubMed] [Google Scholar]
- 103.Moraes J., Nascimento C., Yamaguchi L.F., Kato M.J., Nakano E. Schistosoma mansoni: In vitro schistosomicidal activity and tegumental alterations induced by piplartine on schistosomula. Exp. Parasitol. 2012;132:222–227. doi: 10.1016/j.exppara.2012.07.004. [DOI] [PubMed] [Google Scholar]
- 104.Jisaka M., Kawanaka M., Sugiyama H., Takegawa K., Huffman M.A., Ohigashi H., Koshimizu K. Antischistosomal activities of sesquiterpene lactones and steroid glucosides from Vernonia amygdalina, possibly used by wild chimpanzees against parasite-related diseases. Biosci. Biotechnol. Biochem. 1992;56:845–846. doi: 10.1271/bbb.56.845. [DOI] [PubMed] [Google Scholar]
- 105.McIntosh H.M., Olliaro P. Artemisinin derivatives for treating uncomplicated malaria. Cochrane Database Syst. Rev. 2000:CD000256. doi: 10.1002/14651858.CD000256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Le W.J., You J.Q., Yang Y.Q., Mei J.Y., Guo H.F., Yang H.Z., Zhang C.W. Studies on the efficacy of artemether in experimental schistosomiasis (author’s transl) Yao Xue Xue Bao. 1982;17:187–193. [PubMed] [Google Scholar]
- 107.Yue W.J., You J.Q., Mei J.Y. Effects of artemether on Schistosoma japonicum adult worms and ova. Zhongguo Yao Li Xue Bao. 1984;5:60–63. [PubMed] [Google Scholar]
- 108.Utzinger J., Xiao S., N’Goran E.K., Bergquist R., Tanner M. The potential of artemether for the control of schistosomiasis. Int. J. Parasitol. 2001;31:1549–1562. doi: 10.1016/s0020-7519(01)00297-1. [DOI] [PubMed] [Google Scholar]
- 109.Araujo N., Kohn A., Katz N. Activity of the artemether in experimental schistosomiasis mansoni. Mem. Inst. Oswaldo Cruz. 1991;86:185–188. doi: 10.1590/s0074-02761991000600042. [DOI] [PubMed] [Google Scholar]
- 110.Utzinger J., N’Goran E.K., N’Dri A., Lengeler C., Xiao S., Tanner M. Oral artemether for prevention of Schistosoma mansoni infection: Randomised controlled trial. Lancet. 2000;355:1320–1325. doi: 10.1016/s0140-6736(00)02114-0. [DOI] [PubMed] [Google Scholar]
- 111.Keiser J., Chollet J., Xiao S.H., Mei J.Y., Jiao P.Y., Utzinger J., Tanner M. Mefloquine—An aminoalcohol with promising antischistosomal properties in mice. PLoS Negl. Trop. Dis. 2009;3:e350. doi: 10.1371/journal.pntd.0000350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Erasmus D.A. A comparative study of the reproductive system of mature, immature and “unisexual” female Schistosoma mansoni. Parasitology. 1973;67:165–183. doi: 10.1017/s0031182000046394. [DOI] [PubMed] [Google Scholar]
- 113.Allam G. Immunomodulatory effects of curcumin treatment on murine schistosomiasis mansoni. Immunobiology. 2009;214:712–727. doi: 10.1016/j.imbio.2008.11.017. [DOI] [PubMed] [Google Scholar]
- 114.Cichewicz R.H.L., McKerrow JH, Nair MG. Kwanzoquinones A–G and other constituents of Hemerocallis fulva “Kwanzo” roots and their activity against the human pathogenic trematode Schistosoma mansoni. Tetrahedron. 2002;58:8597–8606. [Google Scholar]
- 115.El-Shenawy N.S., Soliman M.F., Abdel-Nabi I.M. Does Cleome droserifolia have anti-schistosomiasis mansoni activity? Rev. Inst. Med. Trop. Sao Paulo. 2006;48:223–228. doi: 10.1590/s0036-46652006000400010. [DOI] [PubMed] [Google Scholar]
- 116.Jatsa H.B., Ngo Sock E.T., Tchuem Tchuente L.A., Kamtchouing P. Evaluation of the in vivo activity of different concentrations of Clerodendrum umbellatum Poir against Schistosoma mansoni infection in mice. Afr. J. Tradit. Complement. Altern. Med. 2009;6:216–221. [PMC free article] [PubMed] [Google Scholar]
- 117.Jiraungkoorskul W., Sahaphong S., Sobhon P., Riengrojpitak S., Kangwanrangsan N. Schistosoma mekongi: The in vitro effect of praziquantel and artesunate on the adult fluke. Exp. Parasitol. 2006;113:16–23. doi: 10.1016/j.exppara.2005.12.001. [DOI] [PubMed] [Google Scholar]
- 118.Utzinger J., Chollet J., Tu Z., Xiao S., Tanner M. Comparative study of the effects of artemether and artesunate on juvenile and adult Schistosoma mansoni in experimentally infected mice. Trans. R. Soc. Trop. Med. Hyg. 2002;96:318–323. doi: 10.1016/s0035-9203(02)90110-0. [DOI] [PubMed] [Google Scholar]
- 119.Lescano S.Z., Chieffi P.P., Canhassi R.R., Boulos M., Amato Neto V. Antischistosomal activity of artemether in experimental Schistosomiasis mansoni. Rev. Saude Publica. 2004;38:71–75. doi: 10.1590/s0034-89102004000100010. [DOI] [PubMed] [Google Scholar]
- 120.Xiao S., Shen B., Chollet J., Utzinger J., Tanner M. Tegumental changes in adult Schistosoma mansoni harbored in mice treated with artemether. J. Parasitol. 2000;86:1125–1132. doi: 10.1645/0022-3395(2000)086[1125:TCIASM]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- 121.De Oliveira C.B., Comunello L.N., Lunardelli A., Amaral R.H., Pires M.G., da Silva G.L., Manfredini V., Vargas C.R., Gnoatto S.C., de Oliveira J.R., Gosmann G. Phenolic enriched extract of Baccharis trimera presents anti-inflammatory and antioxidant activities. Molecules. 2012;17:1113–1123. doi: 10.3390/molecules17011113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Soliman M.F. Evaluation of avocado/soybean unsaponifiable alone or concurrently with praziquantel in murine schistosomiasis. Acta Trop. 2012;122:261–266. doi: 10.1016/j.actatropica.2012.02.002. [DOI] [PubMed] [Google Scholar]
- 123.Lima C.M., Freitas F.I., Morais L.C., Cavalcanti M.G., Silva L.F., Padilha R.J., Barbosa C.G., Santos F.A., Alves L.C., de Diniz M.F. Ultrastructural study on the morphological changes to male worms of Schistosoma mansoni after in vitro exposure to allicin. Rev. Soc. Bras. Med. Trop. 2011;44:327–330. doi: 10.1590/s0037-86822011005000023. [DOI] [PubMed] [Google Scholar]
- 124.Race G.J., Martin J.H., Moore D.V., Larsh J.E., Jr Scanning and transmission electronmicroscopy of Schistosoma mansoni eggs, cercariae, and and adults. Am. J. Trop. Med. Hyg. 1971;20:914–924. doi: 10.4269/ajtmh.1971.20.914. [DOI] [PubMed] [Google Scholar]
- 125.Hockley D.J. Ultrastructure of the tegument of Schistosoma. Adv Parasitol. 1973;11:233–305. doi: 10.1016/s0065-308x(08)60188-8. [DOI] [PubMed] [Google Scholar]
- 126.Mitsui Y., Miura M., Aoki Y. In vitro effects of artesunate on the survival of worm pairs and egg production of Schistosoma mansoni. J Helminthol. 2009;83:7–11. doi: 10.1017/S0022149X08070235. [DOI] [PubMed] [Google Scholar]
- 127.Thompson D.P., Klein R.D., Geary T.G. Prospects for rational approaches to anthelmintic discovery. Parasitology. 1996;113:S217–S238. doi: 10.1017/s0031182000077994. [DOI] [PubMed] [Google Scholar]
- 128.Sangster N.C., Song J., Demeler J. Resistance as a tool for discovering and understanding targets in parasite neuromusculature. Parasitology. 2005;131:S179–S190. doi: 10.1017/S0031182005008656. [DOI] [PubMed] [Google Scholar]
- 129.Marks N.J., Maule A.G. Neuropeptides in helminths: Occurrence and distribution. Adv. Exp. Med. Biol. 2010;692:49–77. doi: 10.1007/978-1-4419-6902-6_4. [DOI] [PubMed] [Google Scholar]
- 130.Taman A., Ribeiro P. Glutamate-mediated signaling in Schistosoma mansoni: A novel glutamate receptor is expressed in neurons and the female reproductive tract. Mol. Biochem. Parasitol. 2011;176:42–50. doi: 10.1016/j.molbiopara.2010.12.001. [DOI] [PubMed] [Google Scholar]
- 131.Hillman G.R., Gibler W.B., Chu S.H. Fluorescent probes of acetylcholine binding sites-indicators of drug action in Schistosoma mansoni. Biochem. Pharmacol. 1976;25:2529–2535. doi: 10.1016/0006-2952(76)90460-3. [DOI] [PubMed] [Google Scholar]
- 132.Bizimana N., Tietjen U., Zessin K.H., Diallo D., Djibril C., Melzig M.F., Clausen P.H. Evaluation of medicinal plants from Mali for their in vitro and in vivo trypanocidal activity. J. Ethnopharmacol. 2006;103:350–356. doi: 10.1016/j.jep.2005.08.023. [DOI] [PubMed] [Google Scholar]
- 133.Freiburghaus F., Kaminsky R., Nkunya M.H., Brun R. Evaluation of African medicinal plants for their in vitro trypanocidal activity. J. Ethnopharmacol. 1996;55:1–11. doi: 10.1016/s0378-8741(96)01463-8. [DOI] [PubMed] [Google Scholar]
- 134.Youan B.B., Coulibaly S., Miezan T.B., Doua F., Bamba M. In vivo evaluation of sixteen plant extracts on mice inoculated with Trypanosoma brucei gambiense. Bull. World Health Organ. 1997;75:343–348. [PMC free article] [PubMed] [Google Scholar]
- 135.Dua V.K., Verma G., Agarwal D.D., Kaiser M., Brun R. Antiprotozoal activities of traditional medicinal plants from the Garhwal region of North West Himalaya, India. J. Ethnopharmacol. 2011;136:123–128. doi: 10.1016/j.jep.2011.04.024. [DOI] [PubMed] [Google Scholar]
- 136.Mann A., Ifarajimi O.R., Adewoye A.T., Ukam C., Udeme E.E., Okorie II, Sakpe M.S., Ibrahim D.R., Yahaya Y.A., Kabir A.Y., et al. In vivo antitrypanosomal effects of some ethnomedicinal plants from Nupeland of north central Nigeria. Afr. J. Tradit. Complement. Altern. Med. 2011;8:15–21. doi: 10.4314/ajtcam.v8i1.60486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Buckner F.S.N.N. Advances in Chagas disease drug development: 2009–2010. Curr. Opin. Infect. Dis. 2010;23:609–616. doi: 10.1097/QCO.0b013e3283402956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Andrade e Silva M.L., Cicarelli R.M., Pauletti P.M., Luz P.P., Rezende K.C., Januario A.H., da Silva R., Pereira A.C., Bastos J.K., de Albuquerque S., et al. Trypanosoma cruzi: Evaluation of (−)-cubebin derivatives activity in the messenger RNAs processing. Parasitol. Res. 2011;109:445–451. doi: 10.1007/s00436-011-2275-x. [DOI] [PubMed] [Google Scholar]
- 139.Parmar V.S.J., Gupta S., Talwar S., Rajwanshi V.K.K., Azim A., Malhotra S.K., Jain R., Sharma N.K., Tyagi O.D.L., Errington W., et al. Polyphenols and alkaloids from Piper species. Phytochemistry. 1998;49:1069–1078. [Google Scholar]
- 140.Baldoqui D.C., Kato M.J., Cavalheiro A.J., Bolzani V.S., Young M.C., Furlan M.A. chromene and prenylated benzoic acid from Piper aduncum. Phytochemistry. 1999;51:899–902. doi: 10.1016/s0031-9422(99)00132-6. [DOI] [PubMed] [Google Scholar]
- 141.Kato M.J., Furlan M. Chemistry and evolution of the Piperaceae. Pure Appl. Chem. 2007;79:529–538. [Google Scholar]
- 142.Nour A.M., Khalid S.A., Kaiser M., Brun R., Abdallah W.E., Schmidt T.J. The antiprotozoal activity of sixteen asteraceae species native to Sudan and bioactivity-guided isolation of xanthanolides from Xanthium brasilicum. Planta Med. 2009;75:1363–1368. doi: 10.1055/s-0029-1185676. [DOI] [PubMed] [Google Scholar]
- 143.Batista J.M., Jr, Lopes A.A., Ambrosio D.L., Regasini L.O., Kato M.J., Bolzani V.S., Cicarelli R.M., Furlan M. Natural chromenes and chromene derivatives as potential anti-trypanosomal agents. Biol. Pharm. Bull. 2008;31:538–540. doi: 10.1248/bpb.31.538. [DOI] [PubMed] [Google Scholar]
- 144.Flores N., Jimenez I.A., Gimenez A., Ruiz G., Gutierrez D., Bourdy G., Bazzocchi I.L. Antiparasitic activity of prenylated benzoic acid derivatives from Piper species. Phytochemistry. 2009;70:621–627. doi: 10.1016/j.phytochem.2009.03.010. [DOI] [PubMed] [Google Scholar]
- 145.Flores N., Jimenez I.A., Gimenez A., Ruiz G., Gutierrez D., Bourdy G., Bazzocchi I.L. Benzoic acid derivatives from Piper species and their antiparasitic activity. J. Nat. Prod. 2008;71:1538–1543. doi: 10.1021/np800104p. [DOI] [PubMed] [Google Scholar]
- 146.Torres-Santos E.C., Moreira D.L., Kaplan M.A., Meirelles M.N., Rossi-Bergmann B. Selective effect of 2′,6′-dihydroxy-4′-methoxychalcone isolated from Piper aduncum on Leishmania amazonensis. Antimicrob Agents Chemother. 1999;43:1234–1241. doi: 10.1128/aac.43.5.1234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Hermoso A., Jimenez I.A., Mamani Z.A., Bazzocchi I.L., Pinero J.E., Ravelo A.G., Valladares B. Antileishmanial activities of dihydrochalcones from Piper elongatum and synthetic related compounds. Structural requirements for activity. Bioorg. Med. Chem. 2003;11:3975–3980. doi: 10.1016/s0968-0896(03)00406-1. [DOI] [PubMed] [Google Scholar]
- 148.Bodiwala H.S.S.G., Singh R., Dey C.S., Sharma S.S., Bhutani K.K., Singh I.P. Antileishmanial amides and lignans from Piper cubeba and Piper retrofractum. J. Nat. Med. 2007;61:418–421. [Google Scholar]
- 149.Luize P.S., Ueda-Nakamura T., Dias Filho B.P., Cortez D.A., Nakamura C.V. Activity of neolignans isolated from Piper regnellii (MIQ.) C.DC. var. pallescens against Trypanosoma cruzi. Biol. Pharm. Bull. 2006;29:2126–2130. doi: 10.1248/bpb.29.2126. [DOI] [PubMed] [Google Scholar]
- 150.Martins R.C., Lago J.H., Albuquerque S., Kato M.J. Trypanocidal tetrahydrofuran lignans from inflorescences of Piper solmsianum. Phytochemistry. 2003;64:667–670. doi: 10.1016/s0031-9422(03)00356-x. [DOI] [PubMed] [Google Scholar]
- 151.Ribeiro T.S., Freire-de-Lima L., Previato J.O., Mendonca-Previato L., Heise N., de Lima M.E. Toxic effects of natural piperine and its derivatives on epimastigotes and amastigotes of Trypanosoma cruzi. Bioorg. Med. Chem. Lett. 2004;14:3555–3558. doi: 10.1016/j.bmcl.2004.04.019. [DOI] [PubMed] [Google Scholar]
- 152.Cotinguiba F.R., Bolzani V.S., Debonsi H.M., Passerini G.D., Cicarelli R.M.B., Kato M.J., Furlan M. Piperamides and their derivatives as potential anti-trypanosomal agents. Med. Chem. Res. 2009;18:703–711. [Google Scholar]
- 153.Lopes A.A., Lopez S.N., Regasini L.O., Junior J.M., Ambrosio D.L., Kato M.J., da Silva Bolzani V., Cicarelli R.M., Furlan M. In vitro activity of compounds isolated from Piper crassinervium against Trypanosoma cruzi. Nat. Prod. Res. 2008;22:1040–1046. doi: 10.1080/14786410802243271. [DOI] [PubMed] [Google Scholar]
- 154.Ganfon H., Bero J., Tchinda A.T., Gbaguidi F., Gbenou J., Moudachirou M., Frederich M., Quetin-Leclercq J. Antiparasitic activities of two sesquiterpenic lactones isolated from Acanthospermum hispidum D.C. J. Ethnopharmacol. 2012;141:411–417. doi: 10.1016/j.jep.2012.03.002. [DOI] [PubMed] [Google Scholar]
- 155.Nibret E., Ashour M.L., Rubanza C.D., Wink M. Screening of some Tanzanian medicinal plants for their trypanocidal and cytotoxic activities. Phytother. Res. 2010;24:945–947. doi: 10.1002/ptr.3066. [DOI] [PubMed] [Google Scholar]
- 156.Oliveira M.F., d’Avila J.C., Tempone A.J., Soares J.B., Rumjanek F.D., Ferreira-Pereira A., Ferreira S.T., Oliveira P.L. Inhibition of heme aggregation by chloroquine reduces Schistosoma mansoni infection. J. Infect. Dis. 2004;190:843–852. doi: 10.1086/422759. [DOI] [PubMed] [Google Scholar]
- 157.Jones A.K., Bentley G.N., Oliveros Parra W.G., Agnew A. Molecular characterization of an acetylcholinesterase implicated in the regulation of glucose scavenging by the parasite Schistosoma. FASEB J. 2002;16:441–443. doi: 10.1096/fj.01-0683fje. [DOI] [PubMed] [Google Scholar]
- 158.Abdulla M.H., Ruelas D.S., Wolff B., Snedecor J., Lim K.C., Xu F., Renslo A.R., Williams J., McKerrow J.H., Caffrey C.R. Drug discovery for schistosomiasis: Hit and lead compounds identified in a library of known drugs by medium-throughput phenotypic screening. PLoS Negl. Trop. Dis. 2009;3:e478. doi: 10.1371/journal.pntd.0000478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Quezada L.A., McKerrow J.H. Schistosome serine protease inhibitors: Parasite defense or homeostasis? An. Acad. Bras. Cienc. 2011;83:663–672. doi: 10.1590/s0001-37652011000200025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Mendonca-Silva D.L., Pessoa R.F., Noel F. Evidence for the presence of glutamatergic receptors in adult Schistosoma mansoni. Biochem. Pharmacol. 2002;64:1337–1344. doi: 10.1016/s0006-2952(02)01358-8. [DOI] [PubMed] [Google Scholar]