

Abstract
Background. The cag pathogenicity island (cag-PAI) is one of the most important virulent determinants of Helicobacter pylori. An insertion sequence (IS) element of cag-PAI (IS605) has been found to generate H. pylori strains with varying virulence. Aim. To evaluate the impact of IS605 and cag-PAI on H. pylori strains isolated from Taiwanese patients with severity of gastric diseases. Methods. H. pylori isolates were cultured from gastric biopsies from 99 patients with peptic ulcer, chronic gastritis, and gastric carcinoma. Six distinct, well-separated colonies were isolated from each patient and analyzed by genotyping. Results. cagA, cagE, cagM, cagT, orf10, and orf13 were found to be present in 90.0%–100.0% of the H. pylori isolates. A total deletion of cagA, cagE, cagM, cagT, orf10, and orf13 was found in 1 isolate (1.0%). The IS605 element was found to be positive in 15.2% of the isolates. The presence of IS605 was higher in H. pylori isolated from patients with gastric carcinoma (25.0%) than in patients with duodenal ulcer (6.5%) or chronic gastritis (6.3%) (P < 0.001). Conclusions. The majority of the patients examined had intact cag-PAI. IS605 was present in 15.2% and was higher in H. pylori isolated from patients with gastric carcinoma than in those with peptic ulcer.
1. Introduction
Infection with Helicobacter pylori is one of the most common bacterial infections in humans [1]. It has been closely linked with chronic gastritis, peptic ulcer, gastric carcinoma, and mucosa-associated lymphoid tissue (MALT) lymphoma [2]. The cag pathogenicity island (cag-PAI), a major determinant of H. pylori virulence, has been well studied [3, 4]. cag-PAI is an ~40-kb region in the H. pylori genome that contains a cluster of around 30 genes, including a type IV secretion system (T4SS) and the cytotoxin-associated gene A (CagA) [5]. Translocation of CagA into gastric epithelial cells requires delivery by T4SS and subsequent induction of cell signaling, which contributes to the development of pathogenesis in gastric mucosa [6].
An insertion sequence (IS) element, IS605, found in some strains disrupts an otherwise-uninterrupted cag-PAI unit, thereby splitting it into 2 regions (Figure 1). These regions, termed cag I and cag II, contain at least 14 and 16 open reading frames (orfs), respectively [7]. Insertion of IS605 between cag I and cag II generates H. pylori strains with different levels of virulence [8, 9]. Censini et al. reported that the cagA gene is closely linked to the intact cag-PAI [4]. However, the presence of the cagA gene does not guarantee the existence of an intact cag-PAI [10–12].
Figure 1.
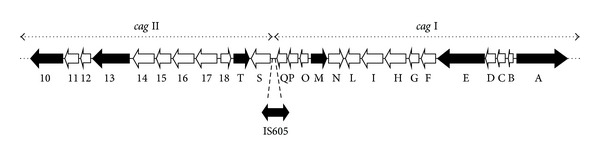
Structure of H. pylori cag pathogenicity island. The localizations of cag I, cag II, and IS605 are shown. The genes assessed in this study including cagA, cagE, cagM, cagT, orf10, and orf13 are indicated as close black.
Diversity within cag-PAI is noted between the people belonging to the Eastern and Western hemispheres [13]. Only one-half to two-thirds of the isolates from the western world carry cag-PAI. In contrast, nearly all East Asian isolates carry cag-PAI [14, 15]. Therefore, cagA and other genes comprising the cag-PAI should be individually investigated. Moreover, cag-PAI appears to be disrupted in the majority of isolates globally [14]. Partial deletion of cag-PAI has been reported in 4%–88% isolates [12–14, 16]. However, the clinical relevance of strains with an intact cag-PAI remains controversial [17]. The aim of this study was to evaluate the impact of IS605 and cag-PAI on H. pylori strains isolated from Taiwanese patients with respect to disease severity.
2. Materials and Methods
2.1. Study Subjects
The study between January 2001 and September 2009 comprised patients who were examined for gastric ulcer, duodenal ulcer, chronic gastritis, or gastric carcinoma, as well as those who were clinically diagnosed with upper gastrointestinal problems. All had completed a self-administered questionnaire prior to being enrolled in the study. Patients were excluded from the investigation if they presented with any of the following: inability to give written informed consent, bleeding tendency (platelet count <50,000/mm3, prothrombin time 3 seconds more than controls, if on anticoagulants), or having taken H2-receptor antagonists or proton pump inhibitors within 2 weeks of enrollment. The study was approved by the Clinical Research Committee of the Veterans General Hospital, Taipei, Taiwan.
2.2. H. pylori Strains and Bacterial Culture
H. pylori isolates were cultured from gastric biopsy specimens and were identified by their positive reactivity for catalase, urease, and oxidase activities [18]. The isolates were cultured at 37°C on brain heart infusion (BHI) agar plates supplemented with 7% horse blood (containing nalidixic acid 10 μg/mL; trimethoprim 5 μg/mL, vancomycin 3 μg/mL, and amphotericin 2 μg/mL) under 12% CO2 with high humidity as in our previously reported study [19]. The rapid urease test was performed using an in-house urease test at room temperature for color change up to 24 hours. The test was defined as positive if the color changed from yellow to red [19, 20].
2.3. Polymerase Chain Reaction
After obtaining positive cultures from the biopsy specimens, 6 isolated colonies from a single culture plate were tested for genotypes with polymerase chain reaction (PCR). These colonies were homogenized in guanidinium isothiocyanate, using a sterile micropestle. DNA were extracted, washed, and eluted in 100 μL of 10 mM Tris-Hcl (pH 8.3). Two microliters of the purified DNA was used for each PCR reaction. Eight primers were employed to assess the upstream and downstream of cag-PAI and IS605: cagA, cagE, cagM, cagT, orf10, orf13, and IS605 (tnpA and tnpB) (Table 1 and Figure 1). PCR was performed under the following conditions: 30 cycles at 94°C for 1 min, at 50.9°C–63°C for 2 min, and at 72°C for 1 min, and a final extension at 72°C for 5 min. PCR products were analyzed on 1.0% agarose gels. Mixed infection was defined if there was different cagA, cagE, cagT, cagM, and orf10, orf13 among the 6 isolates from one plate.
Table 1.
PCR primers used to detect CagA, CagE, CagM, CagT, orf10, orf13, TnpA, and TnpB in H. pylori isolates.
| Genes | Primers | Nucleotide sequences (5′–3′) | Length of PCR products | Location in genome of reference† |
|---|---|---|---|---|
| cagA | cagA-F | GATAACAGGCAAGCTTTTGAGG | 349 | |
| cagA-R | CTGCAAAAGATTGTTTGGCAGA | 14994–15342 | ||
| cagE | cagE-F | GTTACATCAAAAATAAAAGGAAGCG | 735 | |
| cagE-R | CAATAATTTTGAAGAGTTTCAAAGC | 12342–13076 | ||
| cagM | cagM-F | ACAAATACAAAAAAGAAAAAGAGGC | 587 | |
| cagM-R | ATTTTTCAACAAGTTAGAAAAAGCC | 4815–9264 | ||
| cagT | cagT-F | TCTAAAAAGATTACGCTCATAGGCG | 490 | |
| cagT-R | CTTTGGCTTGCATGTTCAAGTTGCC | 1182–1671 | ||
| orf10 | orf10-F | AATAGTGCTTTCTTTAGGATTAGCG | 658 | |
| orf10-R | CCGATTTAATCCTTTCGCTTATGTG | 8970–9627 | ||
| orf13 | orf13-F | CGTTCATGTTCCATACATCTTTGGC | 617 | |
| orf13-R | GATTTATAGCGATCTAAGAAACCGC | 704–1320 | ||
| tnpA | tnpA-F | ATCAGTCCAAAAAGTTTTTTCTTTCC | 338 | |
| tnpA-R | TAAGGGGGTATATTTCAACCAACCG | 154–541 | ||
| tnpB | tnpB-F | CGCTCTCCCTAAATTCAAAGAGGGC | 578 | |
| tnpB-R | AGCTAGGGAAAAATCTGTCTATGCC | 954–1531 |
†GenBank accession number. AF282853.1.
2.4. Statistical Analysis
Descriptive statistics are reported as the proportion for categorical variables with 95% confidence intervals (CI), and means ± standard deviation for continuous variables. The CIs for all proportions are calculated using the standard approximation of binomial. The Chi-square test with Fisher's exact test was used to compare the clinical variables and results. A P value of less than 0.01 was considered significant.
3. Results
Between January 2001 and September 2009, 449 patients (37 patients with chronic gastritis, 101 had duodenal ulcer, 140 with gastric ulcer, and 171 with gastric carcinoma) were enrolled in this study underwent endoscopic examination and H. pylori culture studies. Of the enrolled subjects, 234 patients were positive for H. pylori infection: 25 (68%) with chronic gastritis, 66 (65%) with duodenal ulcer, 77 (55%) with gastric ulcer, and 66 (39%) with gastric carcinoma. Among those patients, we randomly selected 99 subjects (16 patients with chronic gastritis, 31 with duodenal ulcer, 32 with gastric ulcer, and 20 with gastric carcinoma) for further analysis of their H. pylori isolates. Six distinct, well-separated colonies were obtained from each patient making a total of 594 isolates for H. pylori genotyping. In the 99 subjects, mixed infections of H. pylori strains were found in 11 (11.1%) patients.
Our data showed that cagA, cagE, cagM, cagT, orf10, and orf13 were present in between 90.0% and 100% of the isolates (Table 2). There were no differences in the cag-PAI status among isolates from patients with various clinical outcomes. Total deletion of cagA, cagE, cagM, cagT, orf10, and orf13 was found in 1 (1.0%) isolate from a patient with gastric carcinoma. There was only 1 isolate from patient with gastric ulcer had cagA deletion. Deletion of cagM, cagT, orf10, and orf13 was found in 2 (2.0%) isolates (1 isolate from patient with gastric carcinoma and 1 isolate with gastric ulcer), and cagE deletion was found in 4 (4.0%) isolates (each isolated from patient with gastric carcinoma, gastric ulcer, duodenal ulcer, or chronic gastritis, resp.).
Table 2.
Prevalence of H. pylori virulence factors among isolates from patients with gastric carcinoma, gastric ulcer, duodenal ulcer, and chronic gastritis.
| Diagnosis† | GC | GU | DU | CG | Total |
|---|---|---|---|---|---|
| Number of isolates | 20 | 32 | 31 | 16 | 99 |
| CagA | 20 (100.0%) | 31 (96.8%) | 31 (100.0%) | 16 (100.0%) | 98 (99.0%) |
| CagE | 19 (95.0%) | 31 (96.9%) | 30 (96.8%) | 15 (93.8%) | 95 (96.0%) |
| CagM | 19 (95.0%) | 31 (96.8%) | 31 (100.0%) | 15 (93.8%) | 96 (97.0%) |
| CagT | 19 (95.0%) | 31 (96.8%) | 31 (100.0%) | 16 (100.0%) | 97 (98.0%) |
| orf10 | 18 (90.0%) | 32 (100.0%) | 31 (100.0%) | 16 (100.0%) | 97 (98.0%) |
| orf13 | 18 (90.0%) | 32 (100.0%) | 31 (100.0%) | 16 (100.0%) | 97 (98.0%) |
†GC: gastric carcinoma; GU: gastric ulcer; DU: duodenal ulcer; CG: chronic gastritis.
IS605 was found to be present in 15.2% (15/99) of the isolates in this study (Table 3). The presence of IS605 was significantly higher in the H. pylori strains isolated from patients with gastric carcinoma (5/20, 25.0%) than in patients with duodenal ulcer (2/31, 6.5%) or chronic gastritis (1/16, 6.3%) (P < 0.001). It was also higher in isolates from patients with gastric ulcer (7/32, 21.9%) compared to those with duodenal ulcer and chronic gastritis (P < 0.01).
Table 3.
Prevalence of IS605 in H. pylori isolated from patients with gastric carcinoma, gastric ulcer, duodenal ulcer, and chronic gastritis.
| Diagnosis† | Number (%) of IS605 positive isolates |
|---|---|
| GC (n = 20) | 5 (25.0)* |
| GU (n = 32) | 7 (21.9)# |
| DU (n = 31) | 2 (6.5) |
| CG (n = 16) | 1 (6.3) |
| Total (n = 99) | 15 (15.2) |
†GC: gastric carcinoma; GU: gastric ulcer; DU: duodenal ulcer; CG: chronic gastritis.
*P < 0.001, GC versus DU; GC versus CG.
# P < 0.01, GU versus DU; GU versus CG.
4. Discussion
The total number of isolates used in this study was 594, and cag-PAI was found to be present in more than 96% of isolates, which is similar to that reported previously for eastern populations [12, 21–23]. Previous studies from the USA reported that cagA and cagE were detected in 66% and 62% H. pylori strains, respectively [24]. In other studies, cagA, cagE, cagM, and cagT were detected in 93%–100% of Korean and 80%–82% of the Colombian populations [13]. The presence of cagA and cagE in H. pylori isolated from Chinese, Indian, and Malay patients in Singapore ranged from 92.3% to 100% [12]. In southern Taiwan, cagA, cagE, and cagT were previously found to be present in 100% of the domestic strains [21]. Our present study of H. pylori isolates from patients in northern Taiwan yielded similar results, with cagA, cagE, cagM, and cagT being present in more than 96% of the isolates tested.
Patients with intact cag-PAI are thought to be associated with more severe clinical outcomes [10, 25]. Maeda et al. from Japan reported that strains with partial deletions within the cag-PAI were only derived from patients with nonulcer dyspepsia, whereas strains with an intact cag-PAI originated only from patients with gastric carcinoma [11]. Jenks et al. reported that intact cag-PAI was found in 85% and 53% of duodenal ulcer and nonulcer dyspepsia isolates, respectively, [10]. They indicated that the clinical outcome of H. pylori infection was not reliably predicted by any gene of the cag-PAI [10]. However, since the deletion of cag-PAI has been observed in patients with peptic ulcer and nonulcer dyspepsia, the pathogenicity of H. pylori may therefore not be determined by cag-PAI, as suggested by Kawamura et al. [25].
IS605 is located between cag I and cag II in the prototype H. pylori and is thought to be closely related to cag-PAI [4]. However, a recent study has suggested that IS605 is not related to the cag-PAI status [26]. In a previous study involving the Taiwanese population, all isolates were reported as positive for the cag-PAI, but only 36% of these isolates carried an IS605 insertion [21]. The previous controversial study [21] may have used pooled cultures for the analysis, and the actual positive rate of IS605 may be lower because of the possibility of mixed infections.
Owen et al. reported that the distribution of genomic IS605 inserts varied widely with respect to patient disease severity [27]. In this study, IS605 was detected at an overall frequency of 15.2% of isolates and in 25.0% of isolates from patients with gastric carcinoma (Table 3). Thus the presence of IS605 was found to be higher in H. pylori from patients with gastric carcinoma. It was also higher in isolates from patients with gastric ulcer (21.9%) compared to those with duodenal ulcer (6.5%) and chronic gastritis (6.3%). These results were consistent with previous findings by Deguchi et al. who indicated that the presence of IS605 was more frequently associated with the cag13 gene in gastric cancer patients [9]. Kersulyte et al. report that the IS605 element family has homology at the protein level with orfB (putative transposase gene) [28]; this gene is present in a Salmonella prophage and contributes to virulence during bacterial infection [29]. This evidence suggests that the presence of the IS605 element may have a possible modifying role with respect to strain pathogenicity [4]. However, the function of IS605 in relation to H. pylori cag-PAI and the severity of gastric diseases still remain unclear. Further studies are required to clarify the role of IS605 in the development of gastric carcinoma.
In conclusion, a majority of patients infected with H. pylori contained intact cag-PAI. However, the presence of the cagA gene does not guarantee the presence of intact cag-PAI. The presence of IS605 is significantly higher in isolates from patients with gastric carcinoma compared to those with duodenal ulcer and chronic gastritis.
Authors' Contribution
C.-L. Perng is the co-first author.
Conflict of Interests
All authors have no conflict of interests to declare.
Acknowledgments
This work was supported by the Tomorrow Medicine Foundation and the National Science Council, Taiwan, (NSC99-2314-B-038-034-MY2 and NSC101-2314-B-038-007). The authors thank Tzu-En Lin, Tze-Yu Tung, Austin Jen-Liang Lin, and Jen-Hao Lin for their expert technical assistance.
References
- 1.Blaser MJ. Ecology of Helicobacter pylori in the human stomach. The Journal of Clinical Investigation. 1997;100(4):759–762. doi: 10.1172/JCI119588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Dunn BE, Cohen H, Blaser MJ. Helicobacter pylori. Clinical Microbiology Reviews. 1997;10(4):720–741. doi: 10.1128/cmr.10.4.720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Yamaoka Y. Pathogenesis of Helicobacter pylori-related gastroduodenal diseases from molecular epidemiological studies. Gastroenterology Research and Practice. 2012;2012:9 pages. doi: 10.1155/2012/371503.371503 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Censini S, Lange C, Xiang Z, et al. cag, a pathogenicity island of Helicobacter pylori, encodes type I-specific and disease-associated virulence factors. Proceedings of the National Academy of Sciences of the United States of America. 1996;93(25):14648–14653. doi: 10.1073/pnas.93.25.14648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Taylor DE. Helicobacter pylori and its genome: lessons from the treasure map. Canadian Journal of Gastroenterology. 1999;13(3):218–223. doi: 10.1155/1999/964025. [DOI] [PubMed] [Google Scholar]
- 6.Hatakeyama M. Helicobacter pylori and gastric carcinogenesis. Journal of Gastroenterology. 2009;44(4):239–248. doi: 10.1007/s00535-009-0014-1. [DOI] [PubMed] [Google Scholar]
- 7.Slater E, Owen RJ, Williams M, Pounder RE. Conservation of the cag pathogenicity island of Helicobacter pylori: associations with vacuolating cytotoxin allele and IS605 diversity. Gastroenterology. 1999;117(6):1308–1315. doi: 10.1016/s0016-5085(99)70281-7. [DOI] [PubMed] [Google Scholar]
- 8.Akopyants NS, Clifton SW, Kersulyte D, et al. Analyses of the cag pathogenicity island of Helicobacter pylori . Molecular Microbiology. 1998;28(1):37–53. doi: 10.1046/j.1365-2958.1998.00770.x. [DOI] [PubMed] [Google Scholar]
- 9.Deguchi R, Igarashi M, Watanabe K, Takagi A. Analysis of the cag pathogenicity island and IS605 of Helicobacter pylori strains isolated from patients with gastric cancer in Japan. Alimentary Pharmacology and Therapeutics. 2004;20(supplement 1):13–16. doi: 10.1111/j.1365-2036.2004.01982.x. [DOI] [PubMed] [Google Scholar]
- 10.Jenks PJ, Mégraud F, Labigne A. Clinical outcome after infection with Helicobacter pylori does not appear to be reliably predicted by the presence of any of the genes of the cag pathogenicity island. Gut. 1998;43(6):752–758. doi: 10.1136/gut.43.6.752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Maeda S, Yoshida H, Ikenoue T, et al. Structure of cag pathogenicity island in Japanese Helicobacter pylori isolates. Gut. 1999;44(3):336–341. doi: 10.1136/gut.44.3.336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Schmidt HMA, Andres S, Nilsson C, et al. The cag PAI is intact and functional but HP0521 varies significantly in Helicobacter pylori isolates from Malaysia and Singapore. European Journal of Clinical Microbiology and Infectious Diseases. 2010;29(4):439–451. doi: 10.1007/s10096-010-0881-7. [DOI] [PubMed] [Google Scholar]
- 13.Hsu PI, Hwang IR, Cittelly D, et al. Clinical presentation in relation to diversity within the Helicobacter pylori cag pathogenicity island. American Journal of Gastroenterology. 2002;97(9):2231–2238. doi: 10.1111/j.1572-0241.2002.05977.x. [DOI] [PubMed] [Google Scholar]
- 14.Kauser F, Khan AA, Hussain MA, et al. The cag pathogenicity island of Helicobacter pylori is disrupted in the majority of patient isolates from different human populations. Journal of Clinical Microbiology. 2004;42(11):5302–5308. doi: 10.1128/JCM.42.11.5302-5308.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ali M, Khan A, Tiwari SK, Ahmed N, Rao LV, Habibullah CM. Association between cag-pathogenicity island in Helicobacter pylori isolates from peptic ulcer, gastric carcinoma, and non-ulcer dyspepsia subjects with histological changes. World Journal of Gastroenterology. 2005;11(43):6815–6822. doi: 10.3748/wjg.v11.i43.6815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Sozzi M, Tomasini ML, Vindigni C, et al. Heterogeneity of cag genotypes and clinical outcome of Helicobacter pylori infection. Journal of Laboratory and Clinical Medicine. 2005;146(5):262–270. doi: 10.1016/j.lab.2005.06.010. [DOI] [PubMed] [Google Scholar]
- 17.Lu H, Yamaoka Y, Graham DY. Helicobacter pylori virulence factors: facts and fantasies. Current Opinion in Gastroenterology. 2005;21(6):653–659. doi: 10.1097/01.mog.0000181711.04529.d5. [DOI] [PubMed] [Google Scholar]
- 18.Lai CH, Kuo CH, Chen YC, et al. High prevalence of cagA- and babA2-positive Helicobacter pylori clinical isolates in Taiwan. Journal of Clinical Microbiology. 2002;40(10):3860–3862. doi: 10.1128/JCM.40.10.3860-3862.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lin HJ, Lo WC, Perng CL, Tseng GY, Li AFY, Ou YH. Mucosal polymerase chain reaction for diagnosing Helicobacter pylori infection in patients with bleeding peptic ulcers. World Journal of Gastroenterology. 2005;11(3):382–385. doi: 10.3748/wjg.v11.i3.382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Redeen S, Petersson F, Tornkrantz E, et al. Reliability of diagnostic tests for Helicobacter pylori infection. Gastroenterology Research and Practice. 2011;2011:6 pages. doi: 10.1155/2011/940650.940650 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Sheu SM, Sheu BS, Yang HB, Li C, Chu TC, Wu JJ. Presence of iceA1 but not cagA, cagC, cagE, cagF, cagN, cagT, or orf13 genes of Helicobacter pylori is associated with more severe gastric inflammation in Taiwanese. Journal of the Formosan Medical Association. 2002;101(1):18–23. [PubMed] [Google Scholar]
- 22.Perng CL, Lin HJ, Lo WC, Tseng GY, Sun IC, Ou YH. Genotypes of Helicobacter pylori in patients with peptic ulcer bleeding. World Journal of Gastroenterology. 2004;10(4):602–605. doi: 10.3748/wjg.v10.i4.602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Chomvarin C, Namwat W, Chaicumpar K, et al. Prevalence of Helicobacter pylori vacA, cagA, cagE, iceA and babA2 genotypes in Thai dyspeptic patients. International Journal of Infectious Diseases. 2008;12(1):30–36. doi: 10.1016/j.ijid.2007.03.012. [DOI] [PubMed] [Google Scholar]
- 24.Podzorski RP, Podzorski DS, Wuerth A, Tolia V. Analysis of the vacA, cagA, cagE, iceA, and babA2 genes in Helicobacter pylori from sixty-one pediatric patients from the Midwestern United States. Diagnostic Microbiology and Infectious Disease. 2003;46(2):83–88. doi: 10.1016/s0732-8893(03)00034-8. [DOI] [PubMed] [Google Scholar]
- 25.Kawamura O, Murakami M, Araki O, et al. Relationship between gastric disease and deletion of cag pathogenicity island genes of Helicobacter pylori in gastric juice. Digestive Diseases and Sciences. 2003;48(1):47–53. doi: 10.1023/a:1021726213159. [DOI] [PubMed] [Google Scholar]
- 26.Antonio-Rincón F, López-Vidal Y, Castillo-Rojas G, et al. Pathogenicity island cag, vacA and IS605 genotypes in Mexican strains of Helicobacter pylori associated with peptic ulcers. Annals of Clinical Microbiology and Antimicrobials. 2011;10(article 18) doi: 10.1186/1476-0711-10-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Owen RJ, Peters TM, Varea R, Teare EL, Saverymuttu S. Molecular epidemiology of Helicobacter pylori in England: prevalence of cag pathogenicity island markers and IS605 presence in relation to patient age and severity of gastric disease. FEMS Immunology and Medical Microbiology. 2001;30(1):65–71. doi: 10.1111/j.1574-695X.2001.tb01551.x. [DOI] [PubMed] [Google Scholar]
- 28.Kersulyte D, Akopyants NS, Clifton SW, Roe BA, Berg DE. Novel sequence organization and insertion specificity of IS605 and IS606: chimaeric transposable elements of Helicobacter pylori . Gene. 1998;223(1-2):175–186. doi: 10.1016/s0378-1119(98)00164-4. [DOI] [PubMed] [Google Scholar]
- 29.Stanley TL, Ellermeier CD, Slauch JM. Tissue-specific gene expression identifies a gene in the lysogenic phage Gifsy-1 that affects Salmonella enterica serovar typhimurium survival in Peyer’s patches. Journal of Bacteriology. 2000;182(16):4406–4413. doi: 10.1128/jb.182.16.4406-4413.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]