

Abstract
Small-world networks, according to Watts and Strogatz, are a class of networks that are “highly clustered, like regular lattices, yet have small characteristic path lengths, like random graphs.” These characteristics result in networks with unique properties of regional specialization with efficient information transfer. Social networks are intuitive examples of this organization, in which cliques or clusters of friends being interconnected but each person is really only five or six people away from anyone else. Although this qualitative definition has prevailed in network science theory, in application, the standard quantitative application is to compare path length (a surrogate measure of distributed processing) and clustering (a surrogate measure of regional specialization) to an equivalent random network. It is demonstrated here that comparing network clustering to that of a random network can result in aberrant findings and that networks once thought to exhibit small-world properties may not. We propose a new small-world metric, ω (omega), which compares network clustering to an equivalent lattice network and path length to a random network, as Watts and Strogatz originally described. Example networks are presented that would be interpreted as small-world when clustering is compared to a random network but are not small-world according to ω. These findings have important implications in network science because small-world networks have unique topological properties, and it is critical to accurately distinguish them from networks without simultaneous high clustering and short path length.
Key words: brain networks, graph theory
Introduction
The discovery of small-world networks has revolutionized research in network science. In their 1998 landmark paper, Watts and Strogatz described networks that are “highly clustered, like regular lattices, yet have small characteristic path lengths, like random graphs” (Watts and Strogatz, 1998). In other words, small-world networks have the unique ability to have specialized nodes or regions within a network (e.g., a computer network with a group of machines dedicated to a certain task) while simultaneously exhibiting shared or distributed processing across all of the communicating nodes within a network (e.g., all computers sharing the work load). Since that original paper, numerous networks have been described as exhibiting small-world properties, including systems as diverse as the Internet, social groups, and biochemical pathways. Given the unique processing or information transfer capabilities of small-world networks, it is vital to determine whether this is a universal property of naturally occurring networks or whether small-world properties are restricted to specialized networks. An overly liberal definition of small-worldness might miss the specific benefits of these networks—high clustering and low path length—and obscure them with networks more closely associated with regular lattices and random networks.
The current accepted definition of a small-world network is that it has clustering similar to a regular lattice and path length similar to a random network. However, in practice, networks are typically defined as small-world by comparing clustering and path length to those of a comparable random network (Humphries et al., 2006). Unfortunately, this means that networks with very low clustering can be, and indeed are, defined as small-world. We propose that such a method is unable to distinguish true small-world networks from those that are more closely aligned with random or lattice structures and overestimates the occurrence of small-world networks. Networks that are more similar to random or lattice structures are interesting in their own right, but they do not behave like small-world networks. Having a metric to accurately characterize networks as small-world, random, or lattice, or at least tendencies toward one of these types of networks, is an extremely important factor in the study of network science. In this paper, we define a new metric is defined that quantifies small-world properties and places the network in question along a continuum from lattice to small-world to random. This new metric clearly demonstrates that small-world networks are not as ubiquitous as reported and suggests that many systems originally thought to have small-world processing capabilities may in fact not.
Identifying small-world networks
Small-world networks are distinguished from other networks by two specific properties, the first being high clustering (C) among nodes. Mathematically, C is the proportion of edges ei that exist between the neighbors of a particular node (i) relative to the total number of possible edges between neighbors (Bullmore and Sporns, 2009). The equation for C at an individual node of degree ki is:
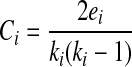 |
[1] |
The overall clustering in a network can be determined by averaging the clustering across all individual nodes. High clustering supports specialization as local collections of strongly interconnected nodes readily share information or resources. Conceptually, clustering is quite straightforward to comprehend. In a real-world analogy, clustering represents the probability that one's friends are also friends of each other.
Small-world networks also have short path lengths (L) as is commonly observed in random networks. Path length is a measure of the distance between nodes in the network, calculated as the mean of the shortest geodesic distances between all possible node pairs:
 |
[2] |
where dij is the shortest geodesic distance between nodes i and j.
Small values of L ensure that information or resources easily spreads throughout the network. This property makes distributed information processing possible on technological networks and supports the six degrees of separation often reported in social networks.
Decades of research has established that short path length is a characteristic of random graphs, while high clustering is a property of lattice networks. Watts and Strogatz developed a network model (WS model) that resulted in the first-ever networks with clustering close to that of a lattice and path lengths similar to those of random networks. The WS model demonstrates that random rewiring of a small percentage of the edges in a lattice results in a precipitous decrease in the path length, but only trivial reductions in the clustering (Fig. 1). Across this rewiring probability, there is a range where the discrepancy between clustering and path length is very large, and it is in this area that the benefits of small-world networks are realized.
FIG. 1.

Watts and Strogatz's small-world model. A simulated 1000-node lattice with average degree k=10 was rewired at varying probabilities p, ranging from 0 to 1. At small values p, network small-worldness is seen in a network with simultaneous high clustering (squares) and low path length (circles).
In the investigation of small-world properties, it has become common to compare both clustering and path length of a network of interest to those same metrics from an equivalent random network. In 2006, Humphries and colleagues introduced a quantitative metric, small-world coefficient σ, that uses a ratio of network clustering and path length compared to its random network equivalent (Humphries et al., 2006). This metric has since gained considerable popularity (Bassett et al., 2008; Guye et al. 2010; Liu et al., 2008), particularly among neuroscientists (Bullmore and Sporns 2009), and has been more extensively evaluated by Humphries and Gurney (2008). In this quantitative approach, C and L are measured against those of their equivalent derived random networks (Crand and Lrand, respectively) to generate the ratios γ=C/Crand and λ=L/Lrand. These ratios are then used to calculate the small-coefficient as:
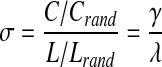 |
[3] |
The conditions that must be met for a network to be classified as small-world are C≫Crand and L≈Lrand, which results in σ>1. The small-world coefficient has been used to ascribe small-world properties to numerous networks ranging from the power grid to the actors network (Humphries and Gurney, 2008).
Comparing path length to that of a random network makes sense—the path length of a small-world network should be short, like that of a random network. However, the comparison of clustering to an equivalent random network does not properly capture small-world behavior because clustering in a small-world network more closely mimics that of a lattice network. Furthermore, it is generally accepted that clustering in the original network is much greater than that of a random network. But how much greater must this clustering be to resemble that of a lattice? If a random network has a Crand of 0.001, on average, only 1 of every 1000 possible connections are present between the neighbors of a node. If the original network has a C five to 10 times greater than Crand, there still exists only five to 10 links out of the 1000 possible connections present. This low level of clustering has been shown in networks (Humphries and Gurney, 2008; Newman 2003) and exemplifies a level of clustering not typically considered for a lattice network. Nonetheless, the magnitude difference between C and Crand shows that the original network is not completely a random network. However, given the low clustering, the extent to which this network is considered small-world is also unclear. It is interesting to note that even the earliest evaluation of small-world properties in real networks compared the clustering to a random network (Watts and Strogatz, 1998).
As it turns out, a major issue with σ is that the clustering coefficient of the equivalent random network greatly influences the small-world coefficient. In the small-world coefficient equation (3), σ uses the relationship between C and Crand to determine the value of γ. Because clustering in a random network is typically extremely low (Humphries and Gurney, 2008; Watts and Strogatz, 1998) the value of γ can be unduly influenced by only small changes in Crand. Consider two networks, A and B, with similar path lengths yet disparate clustering coefficients of 0.5 and 0.05, respectively. If the clustering coefficients of the equivalent random networks are both 0.01, then network A clearly has greater small-world properties. However, if the clustering of the random networks for A and B are 0.01 and 0.001, respectively, then the two networks will have similar values of σ. Interpretation of σ suggests that both networks have the same small-world characteristics even though network B has considerably lower clustering. Although it is true that relative to the comparable random networks, these two networks have the same values of σ, they clearly have different levels of clustering, and network A appears to be more closely aligned with a lattice than does network B. This finding occurs because Crand is in the denominator of the equation for γ; thus, small changes in Crand ultimately drive the value of σ.
The comparison of clustering to a random network presents several limitations to the use of σ. For example, values of σ range from 0 to ∞ and depend on the size of the network in question (Humphries and Gurney, 2008). Larger networks with similar clustering and path length tend to have higher values of σ than do their smaller counterparts. Finally, it may be valuable to know whether a network has properties that tend to be more lattice-like or random-like. It is not possible to determine these properties if clustering and path length are compared to a random equivalent. It is important to determine whether networks exhibit specific behaviors, such as specialization (lattices) or ability to effectively transmit information (random networks). We explored a new metric to quantify small-world properties, ω, that addresses each of the limitations described in the preceding paragraphs and is more in keeping with the original description of small-world networks as defined by Watts and Strogatz.
Novel small-world measurement: ω
Given a graph with characteristic path length, L, and clustering, C, the small-world measurement, ω, is defined by comparing the clustering of the network to that of an equivalent lattice network, Clatt, and comparing path length to that of an equivalent random network, Lrand; the relationship is simply the difference of two ratios defined as:
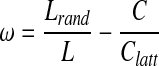 |
[4] |
In using the clustering of an equivalent lattice network rather than a random network, this metric is less susceptible to the fluctuations seen with Crand. Moreover, values of ω are restricted to the interval −1 to 1 regardless of network size. Values close to zero are considered small world: near zero, L≈Lrand and C≈Clatt. Positive values indicate a graph with more random characteristics: L≈Lrand, and C≪Clatt. Negative values indicate a graph with more regular, or lattice-like, characteristics: L≫Lrand, and C≈Clatt.
Methods
Well-known networks datasets
Biological, social, and technological networks were obtained from various sources. All networks in this study were analyzed as binary matrices with unweighted and undirected edges. For disconnected graphs, network analysis was done on the largest component of the network. The e-mail (Guimerà et al., 2003), and Caenorhabditis elegans. metabolic network (Duch and Arenas, 2005) were obtained from Alex Arenas's network datasets (http://deim.urv.cat/∼aarenas/data/welcome.htm). The karate (Zachary 1977), word adjacency (Newman 2006), football (Girvan and Newman, 2002), and Internet networks were obtained from Mark Newman's network data sets (www-personal.umich.edu/∼mejn/netdata/). The Internet network is from unpublished data by the University of Oregon Route Views Project (http://routeviews.org/).
Brain imaging data collection
Brain imaging data were collected from 11 healthy older adults as part of a separate study evaluating an exercise program (Burdette et al., 2010). All data reported here are from post-treatment scans obtained from participants in the control or the treatment group. Whole-brain functional connectivity was evaluated by using graph theory methods on a voxel-by-voxel basis (Eguíluz et al., 2005; Fox et al., 2005; Hayasaka and Laurienti 2010; van den Heuvel et al., 2008). A correlation matrix was produced by computing the Pearson correlation coefficient between all voxel pairs within function magnetic resonance imaging time series. A threshold was applied to the correlation matrix, whereby voxel pairs above the threshold were considered connected and assigned a value of 1, and voxel pairs below the threshold were considered not connected and assigned a value of 0. This binary matrix produces an undirected, unweighted adjacency matrix (Aij) representing the whole-brain functional connectivity for each subject. Mean graph metrics for both groups were compared for degree (k), clustering coefficient (C), and minimum path length (L). For large scale networks, L was calculated by using the harmonic mean of geodesic distances in case of fragmentation (Hayasaka and Laurienti 2010; Latora and Marchiori, 2001). Scanning protocol and network analysis specifics can be found in the supplementary materials.
Random and lattice network construction
To calculate σ for a given network, an equivalent random graph was created by assigning an edge to a node pair with uniform probability while maintaining the degree distribution of the original graph (Maslov and Sneppen, 2002). In this study, edges were rewired at random an average of 10 times for the entire network. Network randomization was performed in 50 networks, with the clustering coefficient and path length calculated for each network. The mean for both graph metrics was calculated and served as the Crand and Lrand of the equivalent random network. It is important to note that ω is valid only if the comparable random network preserves the degree distribution of the original network.
The lattice network was generated by using a modified version of the “latticization” algorithm (Sporns and Zwi, 2004) found in the brain connectivity toolbox (www.brain-connectivity-toolbox.net) (Rubinov and Sporns, 2010). This algorithm was previously used to scale clustering and path length between random and lattice networks (Sporns and Zwi, 2004) and to evaluate the efficiency of networks over varying cost (Achard and Bullmore, 2007). The procedure is based on a Markov-chain algorithm that maintains node degree and swaps edges with uniform probability; however, swaps are carried out only if the resulting matrix has entries that are closer to the main diagonal. To optimize the clustering coefficient of the lattice network, the latticization procedure is performed over several user-defined repetitions. Storing the initial adjacency matrix and its clustering coefficient, the latticization procedure is performed on the matrix. If the clustering coefficient of the resulting matrix is lower, the initial matrix is kept and latticization is performed again on the same matrix; if the clustering coefficient is higher, then the initial adjacency matrix is replaced. This latticization process is repeated until clustering is maximized. This process results in a highly clustered network with long path length approximating a lattice topology. To decrease the processing time in larger networks, a “sliding window” procedure was developed. Smaller sections of the matrix are sampled along the main diagonal, latticized, and reinserted into the larger matrix in a step-wise fashion.
Results
Simulated networks
A 1000-node lattice with average degree k=10 was simulated by creating a 1000×1000 adjacency matrix and assigning links along the five sub- and super-diagonals. The lattice was rewired at varying probabilities p, ranging from 0 to 1, where p=1 resulted in a completely random graph. The rewiring regime used was previously developed by Humphries and Gurney (2008) and is described in detail in the Methods section. The clustering coefficient, path length, and ω were calculated at each rewiring probability (Fig. 2A). The small-world regime was defined qualitatively by Watts and Strogatz (1998) as the region in which clustering (squares) is high and path length (circles) is low. When the plot for ω is overlaid on the Watts and Strogatz plot, three crossing points are of particular interest. When ω crosses the zero point, the network is balanced such that path length is as close to random as clustering is to a lattice. This would be the point where the network exhibits ideal small-world properties, with a balance between normalized clustering and normalized path length. As the ω curve moves toward −1, the trade-off between C and L favors C. When the ω curves crosses the L curve, the trade-off is such that the network falls out of the small-world regime and becomes more like a lattice network. Similarly, as the curve moves toward +1, the balance favors L, and when the ω curves crosses the C curve, the network becomes more like a random network. Recall that ω defines a continuum from lattice to small-world to random. No precise cut-off points define small-worldness, but the proximity of ω to 0 indicates small-world tendencies. However, should one decide that a cut-off is desirable, in this example, the small-world region approximately spans the range −0.5 ≤ω ≤0.5. The small-world coefficient, σ, is shown in a similar fashion (Fig. 2B). According to the definition of the small-world coefficient, a network is considered small-world if σ>1.However, it appears that the network is considered small-world across all rewiring probabilities except for p=1. Thus, one might question the meaning of a network that is almost completely rewired to the level of a random network yet has σ>1.
FIG. 2.

Dynamics of ω and σ. A 1000-node lattice network was simulated and rewired randomly at varying rewiring probabilities (p). (A) Clustering (squares), path length (circles), and the resulting ω curve (dashed line) are shown. (B) Similarly, the resulting σ curve (dashed line) is shown. The axis on the left in both plots indicates the ω and σ values, respectively. The axis on the right in both plots indicates the level of clustering or path length with respect to the C and L of the lattice at p=0.
The simulated network analysis shown in Figure 1 was repeated across networks having N=100, 500, 1000, 5000, and 10,000 nodes. It is important to note the consistency of ω in the range −0.5 ≤ω ≤0.8 across different network sizes (Fig. 3A). This suggests that networks with different sizes, yet similar properties, will have similar ω values, thus making it easier to compare these networks. As stated earlier, values of σ trend higher as network size increases, as seen in the peak σ value range from 3 to 223 (Fig. 3B). In addition, as seen in Figure 2B, networks are classified as small-world across most rewiring probabilities.
FIG. 3.

Demonstration of validity of ω by comparing networks of differing sizes. Networks of varying sizes were simulated and rewired over a range of probabilities as discussed previously. For each network, an ω curve (A) and a σ curve (B) are shown. Although σ is highly dependent on the size of the network, ω curves demonstrate greater consistency over a range of network sizes. This consistency demonstrates the utility of ω in making cross-subject or cross-network comparisons.
Note that as the size of the network is reduced, the tails of the ω curve deviate from the bounds of the [−1,1] interval, particularly from −1. This finding is due to the fact that in a very small network the real path length never becomes very large relative to the path length of the random network. In the equation for ω a “pure lattice” approaches −1 because the real path length is so much larger than the random path length that the Lrand/L of equation (4) approaches zero. If the network is truly small, the path length never exceeds that of the random network by orders of magnitude, as observed for large networks. Nevertheless, the middle portion of the curve for all network sizes overlaps. The tails of the ω curve can also be affected by the density of connections as presented in the Supplementary Materials (Fig. S1). It is important to note that similar curve alterations were observed when the normalized clustering and path length were evaluated by using the plotting method of Watts and Strogatz (Fig. 1). To account for these deviations from predicted behavior, the evaluation of the small-world regime for ω should use plots of C and L for networks of equivalent size and edge density. This method was used to evaluate several real-world networks, presented as follows.
Real-world networks
Comparing the σ values obtained for several biological, social, and technological networks (Table 1) reveals that most networks evaluated here have σ>1 and are, therefore, considered small-world. However, is the most highly clustered network, the C. elegans metabolic network, less small-world than the e-mail network? Clearly the size of these networks is different, and it is not clear how to take this difference into account when evaluating the small-world properties. Similarly, should one interpret that the karate and protein interactions networks share similar properties because their σ values are close to each other? By using the small-world measurement ω with a small-world criterion defined specifically for each system based on the number of nodes and the average degree, conclusions and classification of these networks are quite different. For example, the e-mail network achieves the largest σ value, but the ω value indicates that it is closer to a random network than to a small-world network. Likewise, although the karate and protein interactions networks have σ values that are similar, the ω values indicate that both networks are small-world, but the protein network trends closer to a random topology. It is worth noting that both metrics were able to determine that the word adjacency network is not small-world. However, whereas σ can only say the network is not small-world, ω indicates whether this network is more like a lattice or random network; in this case, the word adjacency network trends toward a random topology.
Table 1.
Network Statistics of Several Well-Known Biological, Social, and Technological Networks
| Network | N | K | C | Crand | Clatt | L | Lrand | σ | ω |
|---|---|---|---|---|---|---|---|---|---|
| Karate (Zachary 1977) | 35 | 4.46 | 0.55 | 0.31 | 0.65 | 2.41 | 2.24 | 1.66 | 0.08 |
| Word adjacency (Newman 2006) | 112 | 7.59 | 0.17 | 0.19 | 0.69 | 2.54 | 2.49 | 0.89 | 0.73 |
| Football (Girvan and Newman, 2002) | 115 | 10.66 | 0.40 | 0.08 | 0.67 | 2.51 | 2.24 | 4.67 | 0.29 |
| Caenorhabditis elegans (metabolic) (Duch and Arenas, 2005) | 453 | 8.94 | 0.65 | 0.28 | 0.80 | 2.66 | 2.50 | 2.18 | 0.12 |
| E-mail (Guimerà et al., 2003) | 1133 | 9.62 | 0.22 | 0.09 | 0.55 | 3.60 | 3.27 | 8.14 | 0.56 |
| Protein interactions (Jeong et al., 2001) | 1539 | 2.67 | 0.07 | 0.04 | 0.19 | 6.81 | 5.69 | 1.47 | 0.47 |
| Internet | 22,963 | 4.22 | 0.23 | 0.09 | 0.68 | 3.84 | 3.58 | 2.28 | 0.51 |
The networks were each obtained from previous work as referenced. N, network size; K, degree; C, clustering coefficient; Crand, clustering of an equivalent random network; Clatt, clustering of an equivalent lattice network; L, path length; Lrand, path length of an equivalent random network; σ and ω, small-worldness metrics.
Another illustration that demonstrates the advantage of ω can be seen in the football network (Fig. 4). The original network is presented here flanked by an equivalent lattice and random network using the Kamada–Kawai model, an algorithm used to optimize the spatial representation of a graph based on the connections (Kamada and Kawai, 1989). The small-world coefficient for each network is σ=3.49, 4.67, and 0.96 for lattice, real, and random networks, respectively. Although the real network is classified as small-world, the equivalent lattice network is also categorized as small-world according to σ. The aberration seen here clearly demonstrates the drawback of comparing the network to an equivalent random network alone. As described earlier, the clustering in a random network, Crand, has a great influence on the value of γ, thus ultimately affecting σ. This occurs because the magnitude difference between C and Crand is often large compared to that of L and Lrand; thus, most lattice networks will be classified as small-world because σ will always be greater than 1 unless the network is completely random. Such influence is not seen with ω, where ω equals −0.60, 0.29, and 0.89, for lattice, real, and random networks, respectively. It can be seen here that ω provides a quantitative measure of the small-world properties over a spectrum of network topologies.
FIG. 4.

Comparison of the football network (B) to its equivalent lattice (A) and random (C) networks. Each network is represented using the Kamada–Kawai algorithm. Although the network itself is considered small-world (σ=4.67), the latticized network is also considered small-world (σ=3.49). Values of ω were −0.60, 0.29, and 0.89 for the lattice, real, and random networks, respectively.
Brain networks
In this section, the utility of ω is demonstrated by using data from a study comparing the effects of an exercise program in older adults previously completed by our group (Burdette et al., 2010). There was no significant difference for the graph metrics C and L between the control and exercise group (Table 2). However, when the mean small-world coefficients for both groups were compared, they were found to be significantly different. Such a group difference was not seen with ω, for −which there was no significant difference between the groups. From an intuitive standpoint, if clustering in a group of networks were to increase, then the small-world coefficient should also increase. However, although the groups had similarly sized networks, the σ value in the exercise group was lower. Given the similar levels of clustering and path length for both groups, the significant difference seen in σ suggests that another factor might influence the calculated value.
Table 2.
Mean Statistics and Standard Deviations for Real Brain Data Comparing Network Changes Due to an Exercise Regimen
| Group | N | C | L | K | σ | ω |
|---|---|---|---|---|---|---|
| Control group | 13,488±327 | 0.32±0.02 | 4.14±0.14 | 45.18±0.46 | 10.28±1.22 | 0.28±0.05 |
| Exercise group | 12,950±662 | 0.34±0.03 | 4.62±0.49 | 46.25±0.59 | 5.62±2.37 | 0.19±0.10 |
The exercise group showed an increase, but nonsignificant increase for both clustering and path length. An investigation of small-world properties revealed a significant difference for σ. In contrast, a nonsignificant difference was found for ω, matching the findings for clustering and path length more closely. N, network size; C, clustering coefficient; L, path length; K, degree; σ and ω, small-worldness metrics.
Plotting the small-world values against the clustering of the network provides some explanation for the differing results for σ and ω (Fig. 5A). A comparison of σ with network clustering reveals a weak relationship (R2=0.4135). In contrast, the value for σ and the equivalent random network is almost perfect (R2=0.9968). A similar relationship was found in real-world networks, where σ was more highly correlated with random clustering, Crand (Fig. 5B). These results suggest that the main factor driving changes in σ is the clustering of the random network. In contrast, when ω is compared to network clustering, its value more accurately reflects clustering in the real networks without undue influence of clustering in the lattice network.
FIG. 5.

Correlations between small-world coefficients and clustering coefficient in brain networks (A) and real-world networks (B). The clustering coefficient of the original networks and clustering of equivalent random network (C and Crand, respectively) were compared with σ. In addition, ω was compared to C and clustering of the equivalent lattice networks (Clatt). R2 values indicate goodness of fit of the trendline (solid line). In both groups σ shows a stronger association with Crand compared to C. In contrast, ω shows a strong association with C without undue influence by Clatt.
Discussion
We have introduced a small-world measurement, ω, that more accurately quantifies network small-world properties as originally defined by Watts and Strogatz. In our exploration of networks, the traditionally used small-world metric σ appears to have high sensitivity in classifying small-world networks. However, it appears that this high sensitivity is coupled with low specificity, resulting in networks being classified as small-world when they are essentially random, with only minor clustering present. In fact, the small-world coefficient has been found to classify nearly all networks as small-world (σ>1) unless the network is a completely random graph. Another drawback of σ is that its value gives no sense of where along the spectrum of lattice to random it falls. Greater σ values are associated with higher levels of small-worldness; however, this may not be the case because σ values do not increase monotonically over varying rewiring probabilities. As seen in Figure 3B, with the exception of the peak, a particular σ value can represent networks with radically different topological properties, thus making network comparisons unreliable; σ can determine whether a network is a random or not, but it cannot effectively determine small-worldness in a network. In our investigation, ω was found to increase monotonically across all networks used in this study, thus facilitating its use as a tool for comparing networks. Unlike the calculation of σ, the calculation of ω involves comparing the clustering coefficient of the network to a lattice network, thereby allowing one to determine how similar the original network is to its lattice equivalent. Using this approach provides inherent scaling and determines how much the original network is like its lattice or random equivalents.
Limitations
One possible reason why comparisons with network lattices have not been used in the literature up to this point is the length of time it takes to generate lattice networks, particularly for large networks. One appeal of comparing the original network to only a random network is the rather fast processing time to generate the random network. Although latticization is fast in smaller networks, large networks such as functional brain networks and the Internet can take several hours to generate and optimize. The latticization procedure described here uses an algorithm developed by Sporns and Zwi in 2004, but the algorithm was used on much smaller datasets. On the basis of some relatively minor modifications (see the Methods section), we were able to use this latticization method on brain networks with more than 15,000 nodes. As processor speed increases, or perhaps with the development of a more computationally efficient algorithm, it will become possible to perform latticization on very large datasets.
Observations of ω show that as network size decreases, the range of ω tends to decrease. As seen in the 100-node network in Figure 2, the minimum ω value is greater than −1. This occurs because in smaller networks, the equivalent lattice tends to have shorter path length. Because the path length in the lattice network is closer to that of the random network, the Lrand/L term in Equation 4 deviates from 0; thus, ω does not approach −1. ω is also limited by networks that have very low clustering that cannot be appreciably increased, such as networks with “super hubs” or hierarchical networks. In hierarchical networks, the nodes are often configured in branches that contain little to no clustering. In networks with “super hubs,” the network may contain a hub that has a node with a degree that is several times in magnitude greater than the next most connected hub. In both these networks, there are fewer configurations to increase the clustering of the network. Moreover, in a targeted assault of these networks, the topology is easily destroyed (Albert et al., 2000). Such vulnerability to attack signifies a network that may not be small-world. However, because of the normalization of clustering in Equation 4, the closeness of C and Clatt can introduce a bias that makes the network appear as if it has high clustering. Similar to the WS model, ω may become unreliable for very sparse networks; thus, it is crucial to examine properties such as edge density to determine whether ω is applicable to the network of interest. In our investigation, one network that may fall into this category is the protein interactions network due to its low edge density and 60% of its edges forming trees. Thus, one should use caution when trying to attribute small-world properties to such networks.
The Watts and Strogatz model is based on a regular lattice with nodes of equal degree. Observed in it are C and L curves in relation to a specific network size and degree distribution. Changing the number of nodes and degree distribution will certainly change these curves, thus affecting the extent of the small-world criterion. For this study, the small-world range was defined to be [−0.5,0.5], but this interval may not be static across differently sized networks. In an ideal network, ω ranges from −1 to 1; a value of 0 indicates a network that is in perfect balance between normalized values of high clustering and low path length, whereas the extremes at −1 and 1 represent a pure lattice and random network, respectively. It is important to note that although ω shows a perfect balance between these normalized values, they may not necessarily be equivalent. Values for ω fall along a continuum between an equivalent lattice and random network, thus values closer to zero have more small-world tendencies. There is no particular range or cut-off for small-worldness; however, a network with an ω of 0.6 shows more tendencies toward a random network than a network with an ω value of 0.4. If one chooses a cut-off range along this continuum, wider ranges will be more sensitive to classifying networks as small-world while a more narrow range will narrow increase specificity. The use of a cut-off depends on the goal of the study, but it is important to recognize that ω should be used as a guideline for classifying small-worldness and has no explicit cut-off range.
Future investigation of network topology, particularly in relation to extreme hubs and the heterogeneity of node degree, is warranted. However, such exploration is beyond the scope of this work. The intention of ω is to indicate the level of small-worldness along a continuum and whether a network exhibits more lattice-like or more random-like behavior. It is worth noting that ω acts as a summary statistic and estimates the overall small-world properties in a network. Mean graph metrics and summary statistics should not serve as an endpoint for network analysis. Instead, these metrics should be used in conjunction with other analyses that can provide further information about the complex organization of the network. Nonetheless, ω is an appealing metric because networks with similar properties share the same value regardless of network size, and in most cases, researchers compare networks of similar size and topology, as seen with brain imaging data. In this regard, ω may be a better choice because it is sensitive to changes in these networks.
Conclusion
We have introduced a new network metric, ω, and have shown that small-world networks may be less ubiquitous than suggested in the current literature. We have demonstrated that this metric more accurately identifies small-world networks and it is able to determine whether a network has more lattice- or random-like properties. In addition, this metric is less sensitive to the size of a network and benefits from inherent scaling, which provides a powerful tool for comparing and ranking small-world properties in various systems. However, the strength of this metric truly lies in its ability to compare similarly sized networks because it allows for a more direct comparison of network properties. The capacity to describe a network as more random- or lattice-like provides considerable benefits to studies investigating dynamic changes in a network. Moreover, direct comparison of small-world properties is useful for group studies, such as brain networks, where understanding the network topology in a particular population may provide further insight into a disease or pathology. This metric provides a useful tool for studying complex systems, but, more important, it truly characterizes where a network falls in the Watts and Strogatz small-world model.
Supplementary Material
Acknowledgments
This work was sponsored by the Wake Forest University General Clinical Research Center (RR07122), National Institutes on Aging (AG029285), National Institute of Neurological Disorders and Stroke (NS070917), and National Institute on Alcohol Abuse and Alcoholism (AA017710, AA019893).
Author Disclosure Statement
No competing financial interests exist.
References
- Achard S. Bullmore E. Efficiency and cost of economical brain functional networks. PLoS Comput Biol. 2007;3:e17. doi: 10.1371/journal.pcbi.0030017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Albert R. Jeong H. Barabási AL. Error and attack tolerance of complex networks. Nature. 2000;406:378–382. doi: 10.1038/35019019. [DOI] [PubMed] [Google Scholar]
- Bassett DS. Bullmore E. Verchinski BA, et al. Hierarchical organization of human cortical networks in health and schizophrenia. J Neurosci. 2008;28:9239–9248. doi: 10.1523/JNEUROSCI.1929-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bullmore E. Sporns O. Complex brain networks: Graph theoretical analysis of structural and functional systems. Nat Rev Neurosci. 2009;10:186–198. doi: 10.1038/nrn2575. [DOI] [PubMed] [Google Scholar]
- Burdette JH. Laurienti PJ. Espeland MA, et al. Using network science to evaluate exercise-associated brain changes in older adults. Front Aging Neurosci. 2010;2:23. doi: 10.3389/fnagi.2010.00023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duch J. Arenas A. Community detection in complex networks using extremal optimization. Phys Rev E Stat Nonlin Soft Matter Phys. 2005;72:027104. doi: 10.1103/PhysRevE.72.027104. [DOI] [PubMed] [Google Scholar]
- Eguíluz VM. Chialvo DR. Cecchi GA. Baliki M. Apkarian AV. Scale-free brain functional networks. Phys Rev Lett. 2005;94:018102. doi: 10.1103/PhysRevLett.94.018102. [DOI] [PubMed] [Google Scholar]
- Fox MD. Snyder AZ. Vincent JL. Corbetta M. Van Essen DC. Raichle ME. The human brain is intrinsically organized into dynamic, anticorrelated functional networks. Proc Natl Acad Sci U S A. 2005;102:9673–9678. doi: 10.1073/pnas.0504136102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Girvan M. Newman MEJ. Community structure in social and biological networks. Proc Natl Acad Sci U S A. 2002;99:7821–7826. doi: 10.1073/pnas.122653799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guimerà R. Danon L. Díaz-Guilera A. Giralt F. Arenas A. Self-similar community structure in a network of human interactions. Phys Rev E Stat Nonlin Soft Matter Phys. 2003;68:065103. doi: 10.1103/PhysRevE.68.065103. [DOI] [PubMed] [Google Scholar]
- Guye M. Bettus G. Bartolomei F. Cozzone P. Graph theoretical analysis of structural and functional connectivity MRI in normal and pathological brain networks. MAGMA. 2010;23:409–421. doi: 10.1007/s10334-010-0205-z. [DOI] [PubMed] [Google Scholar]
- Hayasaka S. Laurienti PJ. Comparison of characteristics between region- and voxel-based network analyses in resting-state fMRI data. Neuroimage. 2010;50:499–508. doi: 10.1016/j.neuroimage.2009.12.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Humphries MD. Gurney K. Network 'small-world-ness': a quantitative method for determining canonical network equivalence. PLoS One. 2008;3:e0002051. doi: 10.1371/journal.pone.0002051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Humphries MD. Gurney K. Prescott TJ. The brainstem reticular formation is a small-world, not scale-free, network. Proc R Soc Lond [Biol] 2006;273:503–511. doi: 10.1098/rspb.2005.3354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jeong H. Mason SP. Barabási A-L. Oltvai ZN. Lethality and centrality in protein networks. Nature. 2001;411:41–42. doi: 10.1038/35075138. [DOI] [PubMed] [Google Scholar]
- Kamada T. Kawai S. An algorithm for drawing general undirected graphs. Inf Process Lett. 1989;31:7–15. [Google Scholar]
- Latora V. Marchiori M. Efficient behavior of small-world networks. Phys Rev Lett. 2001;87:198701. doi: 10.1103/PhysRevLett.87.198701. [DOI] [PubMed] [Google Scholar]
- Liu Y. Liang M. Zhou Y, et al. Disrupted small-world networks in schizophrenia. Brain. 2008;131:945–961. doi: 10.1093/brain/awn018. [DOI] [PubMed] [Google Scholar]
- Maslov S. Sneppen K. Specificity and stability in topology of protein networks. Science. 2002;296:910–913. doi: 10.1126/science.1065103. [DOI] [PubMed] [Google Scholar]
- Newman MEJ. The structure and function of complex networks. SIAM Rev Soc Ind Appl Math. 2003;45:167–256. [Google Scholar]
- Newman MEJ. Finding community structure in networks using the eigenvectors of matrices. Phys Rev E Stat Nonlin Soft Matter Phys. 2006;74:036104. doi: 10.1103/PhysRevE.74.036104. [DOI] [PubMed] [Google Scholar]
- Rubinov M. Sporns O. Complex network measures of brain connectivity: uses and interpretations. Neuroimage. 2010;52:1059–1069. doi: 10.1016/j.neuroimage.2009.10.003. [DOI] [PubMed] [Google Scholar]
- Sporns O. Zwi J. The small world of the cerebral cortex. Neuroinformatics. 2004;2:145–162. doi: 10.1385/NI:2:2:145. [DOI] [PubMed] [Google Scholar]
- van den Heuvel MP. Stam CJ. Boersma M. Hulshoff Pol HE. Small-world and scale-free organization of voxel-based resting-state functional connectivity in the human brain. Neuroimage. 2008;43:528–539. doi: 10.1016/j.neuroimage.2008.08.010. [DOI] [PubMed] [Google Scholar]
- Watts DJ. Strogatz SH. Collective dynamics of 'small-world' networks. Nature. 1998;393:440–442. doi: 10.1038/30918. [DOI] [PubMed] [Google Scholar]
- Zachary WW. An information flow model for conflict and fission in small groups. J Anthropol Res. 1977;33:452–473. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.