

Abstract
Understanding the host genetics of the immune response in retrovirus infection models could provide insights for basic HIV vaccine discovery. In Friend retrovirus (FV) infection of mice, Fv1 differentially inhibits N-tropic versus B-tropic FV infection by mediating a capsid-dependent post-entry block, Fv2 susceptibility governs splenomegaly induction, and Rfv3 resistance primes a stronger neutralizing antibody response due to more potent Apobec3 activity. Apobec3 polymorphisms in inbred mouse strains correlate with Rfv3 resistance and susceptibility, with one unresolved exception. The 129/OlaHsd (129P2) mouse strain is Fv2 and Rfv3 susceptible based on genotyping, but infection of 129P2 mice with B-tropic FV resulted in strong neutralizing antibody responses and no splenomegaly. Here we confirm that 129P2 mice are Fv1nr/nr, explaining its resistance to B-tropic FV. Infection of 129P2 mice with NB-tropic FV, which can efficiently infect mice independent of Fv1 genotype, resulted in severe splenomegaly, high levels of viremia and weak neutralizing antibody responses regardless of Apobec3 status. Notably, high-dose B-tropic FV infection of 129P2 Apobec3-deficient mice induced significant adaptive immune responses and conferred high levels of protection following challenge with pathogenic NB-tropic FV. This immunological protection complemented previous studies that N-tropic FV can act as a live-attenuated vaccine in Fv1 b/b mice. Altogether, the results obtained in 129P2 mice strengthen the conclusion that Rfv3 is encoded by Apobec3, and highlight Fv1 incompatibility as a retroviral vaccine paradigm in mice. Due to its susceptibility to disease that allows for pathogenic challenge studies, B-tropic FV infection of 129P2 mice may be a useful model to study the immunological pathways induced by retroviral capsid restriction.
Introduction
The innate arm of the immune system could critically shape the adaptive immune response against pathogens. Major efforts to understand these innate immune mechanisms against HIV-1 resulted in the identification of restriction factors such as TRIM5α [1] and APOBEC3G [2], but how these factors shape adaptive immune responses against HIV-1 is difficult to study in humans in vivo. In contrast, the interplay between innate and adaptive immunity has been studied extensively in the Friend retrovirus (FV) infection model [3]–[5]. FV is a complex of a replication-competent Friend Murine Leukemia Virus (F-MuLV) and a replication-defective Spleen Focus Forming Virus (SFFV), that cause severe splenomegaly and erythroleukemia in mice [3]–[5]. Different inbred mouse strains exhibited distinct resistance and susceptibility to FV infection and disease that mapped to a handful of genes that include Fv1, Fv2, and Rfv3 [3]–[5] (Table 1). Interestingly, Fv1 and Rfv3 are the functional counterparts of human TRIM5α and APOBEC3G, respectively [6]–[8]. Thus, understanding the impact of Fv1 and mouse Apobec3 (or mA3) on FV adaptive immunity may provide insights for basic HIV vaccine discovery.
Table 1. FV genotype status of various inbred mouse strains.
| Genotypea | ||||
| Host strain | Fv1 | Fv2 | Rfv3 | References |
| C57BL/6 (B6) | b/b | r/r | r/r | [7], [12], [13] |
| C57BL/10 (B10) | b/b | r/r | r/r | [13] |
| BALB/c | b/b | s/s | s/s | [7], [12], [13] |
| A.BYb | b/b | s/s | s/s | [7], [12] |
| A/WySnb | b/b | s/s | s/s | [11], [12], [13] |
| 129/OlaHsd (129P2) | nr/nr | s/s | s/s | This study |
Fv1 controls capsid-dependent tropism; Fv2 is a dominant susceptibility (s) gene governing splenomegaly; Rfv3 resistance (r) is associated with a stronger NAb response. bA/WySn mice also have a defective B-cell activating receptor gene, BAFF-R, that maps near the Rfv3 locus [11], [12], but this was not observed in the related A.BY mice [12].
Rfv3 is a classical gene that influences recovery from FV viremia by promoting a strong neutralizing antibody (NAb) response [9], [10]. Its molecular identification as Apobec3 [7], [11], [12] raised intriguing implications for HIV-1 immunity, since the human homologue APOBEC3G is counteracted by the HIV-1 protein Vif [2]. The case for Apobec3 as the gene encoded by Rfv3 was primarily built on evidence from F1 transcomplementation studies [7], [11], [12]. However, this evidence was also supported by the strong correlation between Apobec3 polymorphisms and the Rfv3 genotype of the inbred strains used to identify and map the Rfv3 gene. Compared to the Apobec3 alleles of Rfv3 susceptible mice such as BALB/c, A/WySn and A.BY strains, Rfv3 resistant C57BL mice (B6 or B10) mice exhibit: (1) high Apobec3 mRNA expression levels [11]–[16] that was linked to a 530 bp Xenotropic Murine Leukemia Virus Long Terminal Repeat (X-MLV) insertion at the Apobec3 exon 2 splice site [12], [16]; (2) splicing of Apobec3 exon 5, resulting in increased translation of an isoform with more potent antiretroviral activity [13], [17]–[19]; and (3) amino acid changes potentially flanking the putative polynucleotide-accommodating groove [16]. These differences could all account for why the Rfv3 resistant allele of Apobec3 is more potent at restricting FV than the Rfv3 susceptible allele in vivo. However, it remains unknown whether the Rfv3 susceptible Apobec3 allele could promote recovery from FV viremia and NAb responses compared to Apobec3-deficient mice.
The Apobec3 polymorphisms in the inbred mouse strains used to define Rfv3 are highly concordant. However, the Apobec3/Rfv3 status of a more recently studied mouse strain in the FV infection model, 129/OlaHsd (129P2 [20]; cited previously as 129/Ola [7]), remains unclear. 129P2 is a substrain of a diverse family of inbred mice under the generic 129 background ([20]–[22]; Fig. 1A), and was used extensively for gene-targeting studies, including Apobec3 [7]. In a previous study, 129P2 mice were classified as Rfv3 resistant because high-dose infection with B-tropic FV resulted in undetectable viremia and potent NAb responses by 28 days post-infection (dpi), similar to B6 mice [7]. However, quantitative PCR data showing that 129P2 mice had relatively high Apobec3 mRNA levels were incorrect [12]. The Fv2 genotype of 129P2 mice also needed to be clarified. Fv2 is a dominant susceptibility gene that dictates splenomegaly induction and is encoded by the Stk gene [23], [24]. In Fv2 susceptible mice, a 3 nt (GGA) insertion in the Stk intron 10 of Fv2 susceptible strains results in an alternative promoter that drives the transcription of a short-form of the Stk kinase (sf-Stk) [24]. Sf-Stk interacts with the erythropoietin receptor in conjunction with the SFFV gp55P protein, resulting in the uncontrolled proliferation of erythroblast precursors that leads to severe splenomegaly [25], [26]. Since 129P2 mice did not develop splenomegaly following B-tropic FV infection, 129P2 mice were classified as Fv2 resistant [7]. However, a different 129 substrain, 129X1 (formerly 129/SvJ [20]), was genotyped as Fv2 susceptible [24]. Since 129 mice were long separated from the Fv2 resistant C57BL lineage [20]–[22], [27], it is unlikely that 129P2 mice are Fv2 resistant. However, due to the complex genealogy of the 129 lineage (Fig. 1A) [20]–[22], [27], direct confirmation of the Fv2 genotype of 129P2 mice would be ideal. Recently, infection of 129P2 mice with an N-tropic MuLV strain, CasFrKP, resulted in high infection levels [28]. These results contrasted from the lack of FV viremia following B-tropic FV infection of 129P2 mice [7]. Thus, the use of a B-tropic [7] versus an N-tropic [28] MuLV strain resulted in divergent infection outcomes in 129P2 mice.
Figure 1. Resolving the Rfv3 status of the 129P2 strain.
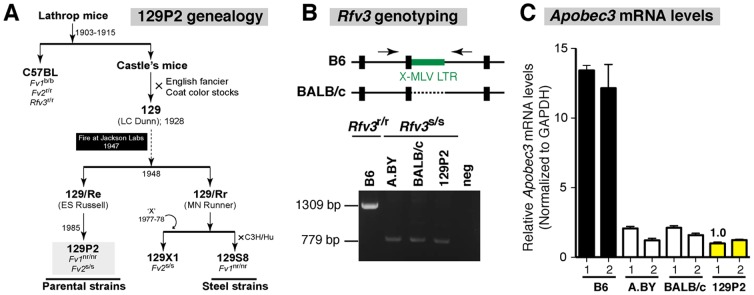
(A) Genealogy of 129P2 mice. Two sublines, 129/Re and 129/Rr, gave rise to contemporary Parental and Steel strains, respectively. 129X1 was genetically contaminated by an unknown strain ‘X’ [21], [22], and genotyped later as Fv2 s/s [24]. The Steel substrain 129S8 was previously genotyped as Fv1 nr/nr [33], [36]. As expected, 129P2 mice had the same Fv1 and Fv2 genotype, suggesting that the entire 129 lineage are Fv1 nr/nr and Fv2 s/s. (B) Rfv3 genotyping. Primers flanking a 530-bp X-MLV LTR insertion in B6 Apobec3 were used. PCR products visualized by agarose electrophoresis revealed that 129P2 mice lacked the insert and are therefore Rfv3 susceptible. (C) Apobec3 mRNA levels. Spleen mRNA were extracted and subjected to quantitative PCR using Apobec3-specific primers and normalized to GAPDH levels [12]. Relative to B6 mice, 129P2 mice had >10-fold lower Apobec3 mRNA levels. Two mice from each strain were tested. Error bars correspond to standard deviations from triplicate determinations.
Fv1 is the gene that dictates N- versus B-tropism [29]. Mouse strains classified as Fv1 n/n support the replication of N-tropic FV strains, whereas Fv1 b/b mice support B-tropic FV replication [30], [31]. Some FV strains can infect both Fv1 n/n and Fv1 b/b mouse strains efficiently and are considered as dual- or NB-tropic. Fv1 incompatibility, such as N-tropic FV infection of Fv1 b/b mice, results in a potent post-entry block [30], [31] that map to amino acid differences in Fv1 and the F-MuLV capsid [32]–[35]. Interestingly, another 129 substrain, 129S8 (formerly 129/SvEv [20]) was genotyped as Fv1 nr, a variant of Fv1 n [33], [36]. Fv1nr restricts B-tropic as well as some N-tropic strains. The Fv1 data on 129S8 [33], [36] strongly suggest that 129P2 mice should also be Fv1 nr/nr. However, the 129 sublines 129/Re and 129/Rr that gave rise to 129P2 and 129S8, respectively, were separated and independently maintained since 1948 (Fig. 1A). Thus, confirmation that 129P2 mice are Fv1 nr/nr may be warranted. We therefore re-evaluated the Fv1, Fv2 and Rfv3 genotype status of 129P2 mice and confirmed these genotypes with in vivo infection with a dual-tropic FV strain. The results demonstrate that the previous classification of 129P2 mice as Fv2 and Rfv3 resistant based on phenotypic data [7] was confounded by Fv1 incompatibility. Importantly, this endeavor revealed new insights on the virological and immunological impact of Apobec3 and Fv1 restriction in 129P2 mice, a mouse genetic background that may be useful for basic retroviral vaccine studies.
Materials and Methods
Mice
Wild-type (WT) 129P2 mice were purchased from Harlan Laboratories, Incorporated. Apobec3 knock-out (KO) mice were generated from the XN150 embryonic stem cell line (BayGenomics) that was made in the 129P2 background. The chimeric mouse was backcrossed once to 129P2 to generate a 100% congenic 129P2 Apobec3 KO strain [7]. B6, A.BY and BALB/c mice were obtained from The Jackson Laboratory. Experimental protocols in mice specifically for this study were approved by the Institutional Animal Care and Use Committee at the University of Colorado Denver [Permit Number B-89709(10)1E]. Infections were performed under isoflurane anesthesia. At indicated timepoints, mice were euthanized using a double-procedure consisting of carbon dioxide inhalation followed by cervical dislocation. All efforts were made to minimize suffering.
Cell culture
Mus dunni and 293T cells were maintained in DMEM (Mediatech) with 2% penicillin/streptomycin/glutamine (Mediatech) and 10% fetal bovine serum (FBS; Gemini).
Genotyping
Tail DNA from 129P2, B6 (Fv1b/b Fv2r/r Rfv3r/r) and BALB/c (Fv1b/b Fv2s/s Rfv3s/s) mice were extracted using the DNAEasy Kit (Qiagen). To genotype Fv1, the full-length gene was amplified using a forward primer Fv1.F (5′-AAGCTTGCGGCCGCGAATTTCCCACGTGCGCTTGCT) and a reverse primer that was either Fv1 b-specific, Fv1b.R (5′-GAATCCTCTAGACTATTAACTGTTGCTTTGATGTTTC), or Fv1 n-specific (5′GAATCCTCTAGACTATCAGAGTTTTGTAGCTGCTGT). PCR products were directly purified (Qiagen), sequenced, and compared to published Fv1 alleles.
To genotype Fv2, we designed primers flanking a 3 nt (GGA) indel in the Stk intron 10 of Fv2 susceptible strains. This indel results in an alternative promoter that drives the transcription of a short-form of the Stk kinase (sf-Stk) [24]. The primers were Stk.Fv2.F (5′-CAGTCCCCTGATGTCCAACT) and Stk.Fv2.R (5′-CCACGGTCATGTTCACAGTC). DNA (100 ng) was subjected to PCR amplification in a 50 μl reaction that contained 1× Phusion HF buffer (Thermo Scientific), 0.2 μM primers, 20 μM dNTP (New England Biolabs), and 1 U of Phusion High-fidelity DNA polymerase (New England Biolabs). The cycling conditions consisted of a 98°C initial denaturation step for 30 s followed by 36 cycles of 98°C for 10 s, 59°C for 15 s, and 72°C for 15 s, and a final extension step of 72°C for 7 min. The 450-bp PCR products were purified (Qiagen) and directly sequenced.
To genotype Rfv3, a primer pair flanking the X-MLV LTR insertion in the exon 2 splice site of Apobec3 was used for PCR [12], [16]. PCR amplicons were sized by agarose gel electrophoresis.
Quantification of Apobec3 mRNA levels
Spleen RNA was extracted using the RNAEasy kit and Apobec3 mRNA levels were quantified by real-time PCR as described [12] and normalized against GAPDH. Apobec3 mRNA levels from spleens of 2 mice of each strain were quantified in triplicate. Quantitative real-time PCR was performed in a Biorad CFX6 machine.
Virus stocks and FV infection
B-tropic and NB-tropic FV stocks, previously obtained from Kim Hasenkrug, Leonard Evans and Bruce Chesebro at the Rocky Mountain Laboratories, National Institutes of Allergy and Infectious Diseases, were prepared and titered in BALB/c mice [7], [37]. Both stocks contain F-MuLV, SFFV and lactate-dehydrogenase elevating virus (LDV). LDV is an RNA virus endemic in wild mice [38] and was present in the classical FV stocks used to define Fv1, Fv2, H-2 and Rfv3 [39]. Since this study builds on historical FV studies that defined Fv1, Fv2, H-2 and Rfv3, FV/LDV stocks were required. Infections were performed by intravenous injection through the retro-orbital route of 140 to 7,500 Spleen Focus Forming Units (SFFU) of FV. Mock infections were performed using DMEM without FBS. Mice were sacrificed at indicated timepoints. In some experiments, 129P2 Apobec3 KO mice that were previously infected with B-tropic FV were challenged with an NB-tropic strain at 28 dpi.
Infectious viremia
Infectious viremia was determined using a focal infectivity assay involving incubations of infected Mus dunni cells with FV gp70-specific MAb 720 [7], [40]. Briefly, serial dilutions of plasma were added into Mus dunni cells seeded the previous day with polybrene, fixed after 2–3 days with ethanol, then MAb 720 supernatant was added for 3 h at 37°C. The fixed cells were washed 3× with TNE buffer (10 mM Tris, 200 mM NaCl, 1 mM EDTA, pH = 7.4) with 0.25% Tween-20 (TBS-T), then 1∶500 sheep anti-mouse IgG conjugated to horseradish peroxidase (HRP; Amersham) was added for 1 h. The cells were washed 3× with TNE then developed with aminoethylcarbazole (Sigma) substrate in the presence of H2O2 (Sigma). Foci were counted and expressed as focal infectivity units (FFU) per ml of plasma.
Spleen infectious center assay
A day before harvesting spleens, Mus dunni cells were seeded at 8,000 cells per well in 24-well plates and 32,000 cells per well in 6-well plates in media with polybrene. The next day, spleens were pre-weighed, and no more than 400 mg were disaggregated and diluted to 10 ml with complete media. For the 6-well plates, 1 ml of spleen suspension was added, while for 24-well plates, the splenocytes were serially diluted 10-fold, adding 100 µl of cell suspension. Infectious centers were detected using the same procedure for the infectious viremia assay described above, using MAb 720 supernatant as the detecting antibody. Foci were counted, multiplied by the dilution factor for the entire spleen, and expressed as spleen infectious centers per spleen.
Plasma viral load
Viral RNA copy numbers were quantified from 10 µl plasma by quantitative real-time PCR [12], [41]. T7-transcribed RNA was used as a standard for absolute quantification. The assay has a limit of detection of 103 copies/ml and >95% efficiency.
Endpoint FV-specific IgG titer
Endpoint ELISAs were performed as described [41]. Native FV virions were coated (500 ng/well) into Immulon-4 plates overnight at 4°C then blocked for 2 h with Superblock (Pierce). Serial 2-fold dilutions of plasma were added onto the plate, incubated at 37°C for 1 h, then washed 6× with PBS with 0.25% Tween-20 (PBS-T). Biotinylated goat anti-mouse IgG (Southern Biotechnology) was added at 1:4000 and incubated at 37°C for 1 h then washed 6× with PBS-T. Streptavidin conjugated to HRP was added at 1:4000 and incubated at 37°C for 30 min then washed 6× with PBS-T, followed by the addition of 3,3′,5,5′-tetramethylbenzidine (TMB) substrate (BioFX). Reactions were stopped with 0.3N H2SO4, then read in a Victor X5 (Perkin Elmer) plate reader at 405 nm. Endpoint titers were calculated by interpolating 2× the average background per plate from a best-fit nonlinear regression curve calculated using Prism 5.0 software (GraphPad).
NAb assay
NAb titers were determined by incubating serial dilutions of plasma with 150 FFU of N-tropic F-MuLV for 1 h at 37°C and adding a third of the mixture onto plated Mus dunni cells. FV infection foci were counted [7], and 80% inhibitory concentrations were calculated by interpolation from a best-fit nonlinear regression curve using Prism 5.0 (GraphPad).
Flow cytometry
To assess cell-mediated immune responses, splenocytes (106 cells) were stimulated for 5 h with PMA (25 ng/ml) and ionomycin (0.7 ng/ml) (Sigma) at 37°C and simultaneously stained with anti-CD107a-PE-Cy7 (clone ID4B) (BD Biosciences). Brefeldin A (100 µg/ml) (Sigma) was added after 1 h. Cells were washed 2× in FACS buffer (PBS + 1% FBS), stained with anti-CD4-PE-CF594 (RM4-5), CD8-FITC (53-6.7) and CD49b-APC (DX5) (BD Biosciences) for 30 min at 4°C, washed 2×, then permeabilized and fixed in Perm/fix buffer (BD Biosciences) prior to staining with anti-IFNγ-PE (XMG1.2) (BD Biosciences). To assess FV infection levels, BM and splenocytes were stained for 1 h at 4°C with MAb34, a monoclonal antibody against the FV Glyco-Gag protein, as previously described [42]. The cells were washed once in FACS buffer, then costained with goat anti-mouse IgG2b-APC (Columbia Biosciences). Cells were also stained with antibodies to CD3-AlexaFluor700 (17A2), CD19-APC-H7 (1D3), CD11b-PE-CF594 (MI/70) (BD Biosciences); Ter119-FITC (TER-119) and CD11c-PE-Cy7 (N418) (eBioscience). To monitor B cell subpopulations, cells were stained with anti-B220-PerCP (RA3-6B2), CD138-PE (281-2), GL7-FITC (GL7) (BD Biosciences) and IgD-eFlour450 (11-26c) (eBioscience) for 30 min at 4°C. For all stainings, the cells were washed then fixed in 1% paraformaldehyde in PBS. Isotype controls were used for gate construction. Samples were analyzed on an LSR II (BD Biosciences) flow cytometer, collecting 200,000–500,000 events per sample. Data were analyzed using FlowJo (Tree Star).
Statistical analysis
Datasets with a normal distribution based on the Kolmogorov-Smirnov normality test were analyzed using a 2-tailed Student’s t test. Otherwise, a 2-tailed Mann-Whitney U test was used. Statistical analyses were performed using the Prism 5.0c (GraphPad). P values less than 0.05 were considered statistically significant.
Results
Fv1, Fv2 and Rfv3 genotypes of 129P2 mice
The 129 mouse lineage exhibits a complex genealogy resulting in multiple substrains (Fig. 1A). Thus, the Fv1, Fv2 and Rfv3 genotypes of 129P2 mice, a representative of the ‘Parental’ 129 substrains, were re-evaluated. As expected, sequencing of the entire Fv1 gene of 129P2 mice revealed 100% nucleotide and amino acid identity to the previously reported Fv1nr sequence from 129S8 [36]. The C-terminal residues were distinct from Fv1b as previously reported. In contrast to Fv1n and Fv1b which encode a Ser at position 352, the 129P2 Fv1 encoded a Phe, consistent with its designation as Fv1nr [36]. To genotype Fv2, a segment of the Stk gene encompassing a 3-nt indel (GGA) that dictates alternative transcription of sf-Stk [24] was amplified from DNA of 129P2, B6 (Fv2 r/r), BALB (Fv2 s/s) and A.BY (Fv2 s/s) strains. 129P2 mice encoded the GGA insertion, similar to BALB and A.BY, but in contrast to B6. To genotype Rfv3, a primer set encompassing a 530 bp X-MLV LTR insertion in the Apobec3 exon 2 splice site [12], [16] was used for PCR. These analyses revealed that 129P2 mice lacked this X-MLV LTR insertion in Apobec3 (Fig. 1B), similar to Rfv3 s/s BALB and A.BY mice. In addition, 129P2 mice had >10-fold lower Apobec3 mRNA levels in the spleen compared to B6 mice (Fig. 1C). Thus, based on genotyping, 129P2 mice are Fv1 nr/nr Fv2 s/s Rfv3 s/s (Table 1).
Infection of 129P2 WT and Apobec3 KO mice with B-tropic versus NB-tropic FV
High-dose (7500 SFFU) infection of 129P2 mice with B-tropic FV did not induce splenomegaly and resulted in potent NAb responses, prompting the earlier designation of 129P2 mice as Fv2 r/r and Rfv3 r/r [7]. However, these results were likely due to Fv1 incompatibility, since 129P2 mice are Fv1 nr/nr (Table 1). To overcome a potential Fv1 restriction block, 129P2 mice were infected with NB-tropic FV, which should infect mice regardless of Fv1 genotype [37]. The mice were sacrificed at 28 dpi (Fig. 2A), because recovery from viremia and NAb responses could be reproducibly detected by this timepoint [9], [10].
Figure 2. 129P2 Apobec3 does not promote recovery from viremia and disease following NB-tropic FV infection.
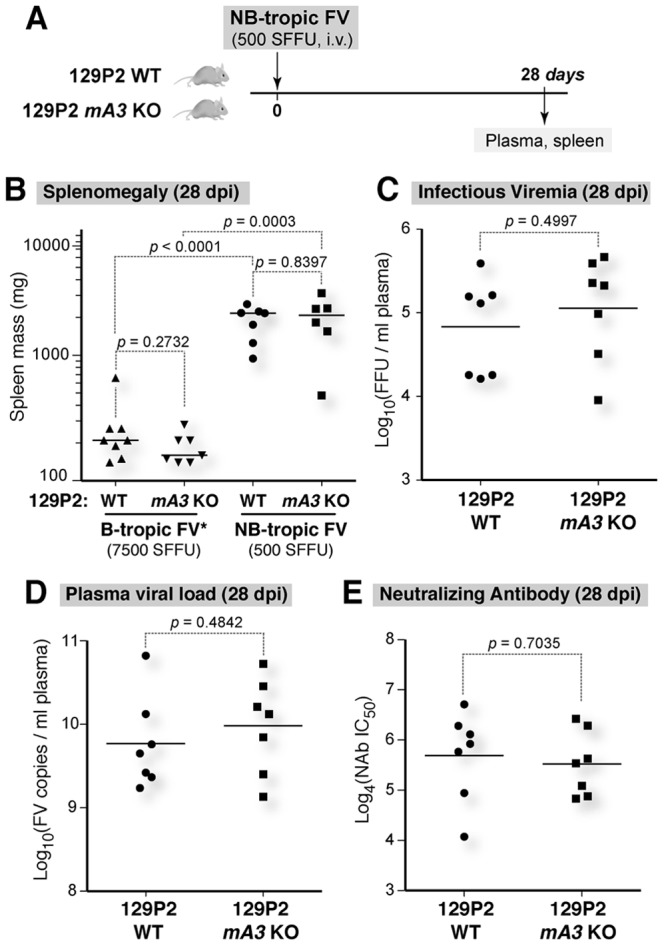
(A) Infection schedule. Mice were sacrificed at 28 dpi for subsequent analyses. (B) Spleen mass. A normal mouse spleen would have ∼100 mg of wet spleen weight. NB-tropic FV infection induced severe splenomegaly but not B-tropic FV. (C) Plasma infectious viremia, based on a focal infectivity assay in Mus dunni cells. (D) Plasma viral load, based on quantitative PCR. (E) NAb titer, calculated based on neutralization curves following incubation of serial dilutions of heat-inactivated plasma with FV in Mus dunni cells. For panels B to E, black bars represent the mean, and each dot corresponds to an individual mouse. Statistical analyses were performed using a 2-tailed Student’s t-test, and p values <0.05 were considered statistically significant.
Consistent with previous data [7], high-dose B-tropic FV infection did not elicit severe splenomegaly in 129P2 mice (median spleen mass = 0.21 g) at 28 dpi. In sharp contrast, NB-tropic FV infection resulted in 10-fold higher splenomegaly (median spleen mass = 2.2 g), despite a 15-fold lower inoculum dose compared to B-tropic FV infection (Fig. 2B). This result confirms that 129P2 mice are Fv2 s/s.
In a previous study, CasFrKP MuLV that lacks the putative Apobec3 antagonist, Glyco-Gag, replicated to higher levels in 129P2 Apobec3 KO versus WT mice [28]. These results, coupled with data showing that 129P2 Apobec3 can significantly inhibit F-MuLV in vitro [7], suggested that 129P2 Apobec3 is a functional antiretroviral restriction factor. However, the virological and immunological impact of 129P2 Apobec3 in pathogenic FV infection remains unknown. We therefore infected 129P2 WT and Apobec3 KO mice with NB-tropic FV and evaluated infection parameters at 28 dpi. There was no significant difference in splenomegaly between 129P2 WT and Apobec3 KO mice (Fig. 2B). Importantly, plasma viral load (Fig. 2C), infectious viremia (Fig. 2D) and NAb titer (Fig. 2E) at 28 dpi were not significantly different between 129P2 WT and Apobec3 KO mice. Thus, the 129P2 Apobec3 allele did not promote recovery from viremia and NAb responses during pathogenic FV infection, consistent with the genotype status of 129P2 mice as Rfv3 s/s.
B-tropic FV infection of Apobec3-deficient 129P2 mice induced significant cell-mediated and humoral immune responses
The finding that B-tropic FV did not induce splenomegaly in Fv1 nr/nr 129P2 mice suggested that FV replication was blocked due to Fv1 incompatibility. This observation mirrored previous results with N-tropic FV infection of Fv1 b mice, where N-tropic FV replicated at near-undetectable levels [43]–[47]. Interestingly, N-tropic FV infection of Fv1 b mice resulted in substantial protection from pathogenic B-tropic FV challenge [43]–[46] (Table 2). N-tropic FV induced potent cellular and humoral immune responses that can be adoptively transferred into naïve hosts [45], [47]. Thus, N-tropic FV functioned as a live-attenuated vaccine in an Fv1-incompatible host. However, the reverse scenario with B-tropic FV had not previously been tested.
Table 2. Retrovirus vaccine studies based on Fv1 incompatibility.
| Genotype | |||||||||
| Year | Host strain | Fv1 | Fv2 | H2 a | Rfv3 | Vaccine strainb | Challenge Strainb | Results | Ref. |
| 1986 | (B10.A × A.BY)F1 | b/b | r/s | a/b | r/s | N-tropic FV complex | B-tropic FV complex | Protection from challenge independent of H2 | [43] |
| (B10.A × A/WySn)F1 | b/b | r/s | a/a | r/s | N-tropic FV complex | B-tropic FV complex | Protection from challenge independent of H2 | [43] | |
| 1998 | (B10.A × A/WySn)F1 | b/b | r/s | a/a | r/s | N-tropic F-MuLV helperc | B-tropic FV complex | Protection due to immunity, not receptor interference | [44] |
| 1999 | (B10.A × A.BY)F1 | b/b | r/s | a/b | r/s | N-tropic F-MuLV helperc | B-tropic FV complex | T-cell protection transferrable to naïve host | [45], [46] |
| 2004 | B6 (adoptive transfer) | b/b | r/r | b/b | r/r | N-tropic F-MuLV helperc | B-tropic FV complex | Virus-specific antibodies are critical for protection | [47] |
| 2013 | 129P2 Apobec3 KO | nr/nr | s/s | b/b | null | B-tropic FV complex | NB-tropic FV complex | Protection from challenge independent of Apobec3 | This study |
H2 dictates cell-mediated immune responses, with the b haplotype being more protective than a. bEntries designated as FV complex correspond to the classical FV stocks that contain F-MuLV helper virus, SFFV and LDV. cWithout SFFV, F-MuLV is nonpathogenic and does not cause splenomegaly.
We therefore investigated if B-tropic FV can function as a live-attenuated vaccine in 129P2 mice. We utilized 129P2 Apobec3 KO mice to ensure that Apobec3 will have no impact on the results. Although the 129P2 Apobec3 allele did not influence NAb responses and recovery from pathogenic FV infection (Fig. 2), 129P2 Apobec3 may still restrict less pathogenic or attenuated MuLV strains [28]. 129P2 Apobec3 KO mice were infected with either mock (DMEM) or high-dose (7500 SFFU) B-tropic FV. At 28 dpi, the mice were evaluated for FV infection and immune responses (Fig. 3A). As expected, B-tropic FV infection of 129P2 Apobec3 KO mice did not induce severe splenomegaly (Fig. 3B) and infectious titers were below the limit of detection (≤600 FFU/ml) in the plasma (Fig. 3C). In contrast, infection of BALB/c mice with 53-fold lower dose (140 SFFU) of the same B-tropic FV stock induced 11-fold higher splenomegaly (Fig. 3B). Moreover, assuming that the infectious titer in B-tropic FV infected 129P2 mice is the limit of detection (600 FFU/ml), the same virus stock induced at least an 800-fold higher infectious viremia in BALB/c mice (Fig. 3C). The relatively mild splenomegaly induced by B-tropic FV infection of 129P2 Apobec3 KO mice (Fig. 3B) suggested that B-tropic FV replicated below the detection limit of the plasma infectious viremia assay (Fig. 3C). We therefore measured FV infection levels using the more sensitive spleen infectious center assay, which involves co-incubating splenocytes with Mus dunni cells. As expected, this assay revealed very high (>108) spleen infectious centers in BALB/c mice, whereas only 2 of 5 129P2 Apobec3 KO mice had detectable signals (Fig. 3D). We also evaluated 129P2 wild-type mice infected with B-tropic FV at 28 dpi and found only 2 of 4 infected mice had detectable spleen infectious centers (data not shown). The mild splenomegaly, undetectable infectious viremia and sporadic detection of spleen infectious centers suggested that B-tropic FV replicated at very low levels in 129P2 Apobec3 mice by 28 dpi, and was therefore ‘live-attenuated’.
Figure 3. B-tropic FV replication in 129P2 Apobec3 KO versus BALB/c mice.

(A) Infection schedule. At 28 dpi, mice were sacrificed for analyses. (B) Spleen mass. Severe splenomegaly was observed in BALB/c, but not 129P2 Apobec3 KO mice following B-tropic FV infection. Data were analyzed using a 2-tailed Student’s t test. (C) Infectious viremia. B-tropic FV infection of 129P2 Apobec3 KO mice resulted in undetectable infectious viremia using the focal infectivity assay in Mus dunni cells. The limit of detection is 600 FFU/ml plasma. (D) Spleen infectious centers. Titrated amounts of 28 dpi splenocytes were co-incubated with Mus dunni cells and developed using the focal infectivity assay procedure. The data in panels C and D were analyzed using a 2-tailed Mann-Whitney U test. For panels B to D, p values <0.05 were considered statistically significant. Black bars represent the mean, and each dot corresponds to an individual mouse. Data are representative of two independent experiments.
We next evaluated the induction of immune responses by flow cytometry (Fig. 4A). Compared to uninfected controls, significant induction of IFN-γ in stimulated CD4+ T cells, CD8+ T cells and DX5+ NK cells were observed (Fig. 4B). Significant induction of CD107a, a degranulation marker, was observed in CD8+ T cells but not in CD4+ T cells and DX5+ cells (Fig. 4C). In addition, splenic germinal center (GL7+ IgD-) B cells were significantly induced in mice infected with B-tropic FV (Fig. 4D). Using 28 dpi plasma from 129P2 Apobec3 KO mice infected with pathogenic NB-tropic FV as a reference, we observed similar FV-specific endpoint IgG titers from 28 dpi plasma of B-tropic FV infected 129P2 Apobec3 KO mice (Fig. 4E). However, significantly higher NAb responses (Fig. 4F) were observed in B-tropic FV infected 129P2 Apobec3 KO mice. These findings demonstrate that B-tropic FV infection of 129P2 Apobec3 KO mice induced significant cell-mediated and humoral immune responses.
Figure 4. Fv1 incompatibility induces significant cell-mediated and humoral immune responses.

(A–D) Apobec3-deficient 129P2 mice were infected with 7500 SFFU B-tropic FV (FV-B) or DMEM (mock). Mouse splenocytes were analyzed by flow cytometry at 28 dpi. (A–C) Cell mediated immune responses. Splenocytes were stimulated with PMA/ionomycin for 5 h prior to flow cytometric analysis. (A) Representative pseudo-color plots and histograms. Flow cytometry plots show the gating strategy for CD4+ and CD8+ lymphocytes. (B) Intracellular IFN-γ and (C) Surface CD107a in lymphocyte subsets. (D) Splenic germinal center (GL7+ IgD-) B cells. (E-F) Analysis of FV-specific antibody responses in 28 dpi plasma of 129P2 Apobec3 KO mice following infection with B-tropic FV (7500 SFFU) or NB-tropic FV (500 SFFU). (E) FV-specific IgG endpoint titers as measured by ELISA. (F) NAb titers. For panels B to F, the black bars represent the mean, and each dot corresponds to an individual mouse. Data were analyzed using a 2-tailed Student’s t test with p<0.05 considered as significant. Data are representative of two independent experiments.
B-tropic FV infection of Apobec3-deficient 129P2 mice protects from pathogenic NB-tropic FV challenge
We next evaluated whether B-tropic FV infection of 129P2 Apobec3 KO mice (Fig. 5) will protect from subsequent infection with pathogenic NB-tropic FV. Mice previously inoculated with either DMEM or B-tropic FV were challenged with pathogenic NB-tropic FV at 28 dpi (Fig. 5A). FV infection levels were determined 7 days later by flow cytometry (Fig. 5B). Significantly lower cellular FV infection in BM cells as well as major BM target cells that include Ter119+ erythroblasts and CD11b+ myeloid cells were observed in B-tropic FV ‘vaccinated’ mice (Fig. 5C). Significant protection was also observed in the spleen, including erythroblasts and B cells (Fig. 5D). Mice previously inoculated with B-tropic FV had undetectable infectious plasma viremia at 7 days post-challenge with NB-tropic FV, whereas mock-infected mice showed high levels of infectious plasma viremia (Fig. 5E). More sensitive plasma viral load assays showed that mice previously inoculated with B-tropic FV had >2500 fold lower viral RNA load in the plasma compared to mock (Fig. 5F). Thus, B-tropic FV acted as a live-attenuated vaccine in Fv1-incompatible 129P2 mice.
Figure 5. B-tropic FV protected 129P2 Apobec3 KO mice from pathogenic NB-tropic FV challenge.
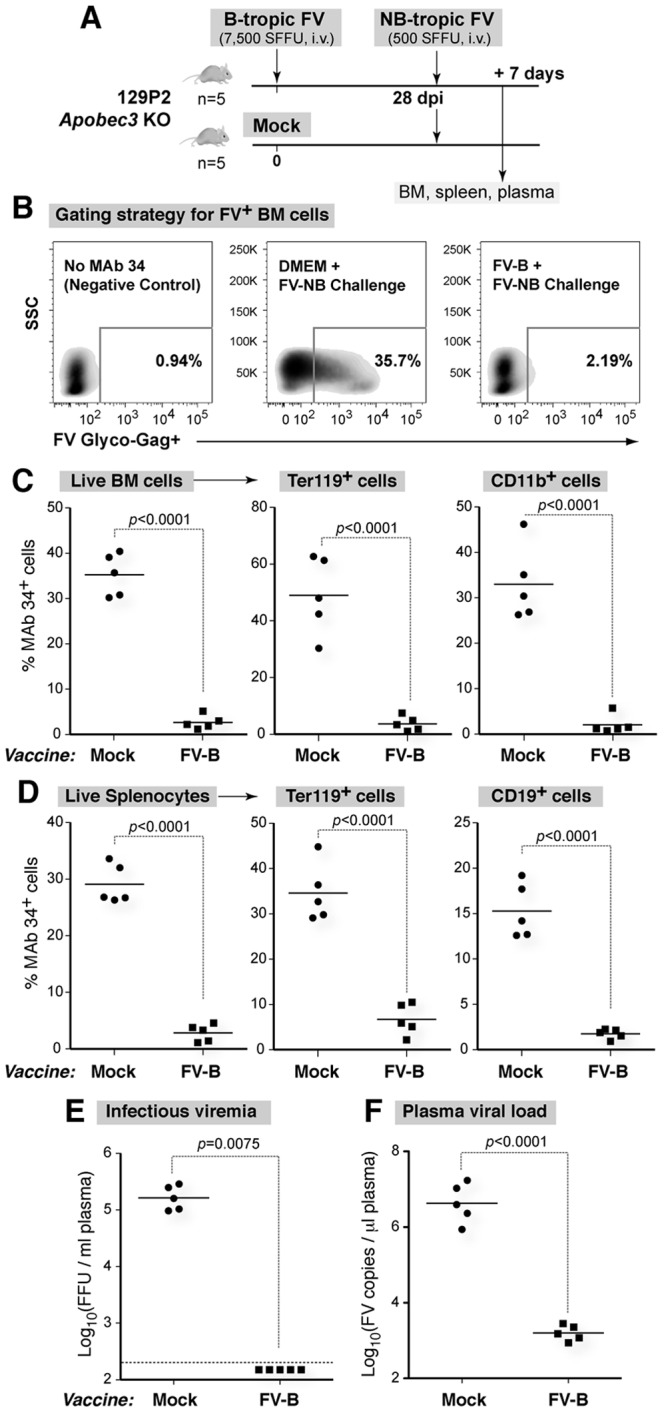
(A) Infection schedule. 129P2 Apobec3 KO mice were infected with B-tropic FV (FV-B) or mock (DMEM). At 28 dpi, the mice were challenged with NB-tropic FV (FV-NB) and evaluated for infection levels by flow cytometry 7 days later. (B) Gating strategy for FV+ cells. BM cells and splenocytes (not shown) were incubated with an FV Glyco-Gag-specific IgG2b antibody, MAb 34, then stained with a goat anti-mouse IgG2b antibody conjugated to APC. Representative flow plots demonstrating the gating strategy for MAb 34+ cells are shown. Cellular infection levels at 7 days post-challenge are shown for (C) BM and (D) Splenocytes. Ter119+ erythroblasts, CD11b+ myeloid cells and/or CD19+ B cells were gated from the live population. Plasma samples were analyzed for (E) Infectious viremia based on a focal infectivity assay in Mus dunni cells and (F) plasma viral load based on viral RNA copies detected by quantitative RT-PCR. Black bars represent the mean, and each dot corresponds to an individual mouse. Flow cytometry and plasma viral load data were analyzed using a 2-tailed Student’s t test. Infectious viremia data were analyzed using a 2-tailed Mann-Whitney U test. Data were considered statistically significant with p<0.05. Data are representative of two independent experiments.
Discussion
The Friend retrovirus (FV) infection model was instrumental in the identification of host genes that can influence the retrovirus-specific immune response. One classical gene, Rfv3, influences recovery from FV viremia by modulating the NAb response, and was recently found to be encoded by the innate restriction factor Apobec3. However, the correlation between Apobec3 polymorphisms and Rfv3 genotype is challenged by uncertainties in the Apobec3/Rfv3 genotype of an inbred strain known as 129P2. We therefore resolved the FV resistance genotype and phenotype of 129P2 mice. The results revealed that 129P2 are resistant to B-tropic FV (Fv1 nr/nr), are susceptible to splenomegaly (Fv2 s/s), and encode an Rfv3-susceptible allele of Apobec3 (Table 1). The latter result provided a unique opportunity to evaluate the Rfv3-susceptible allele of Apobec3. Most studies on Apobec3/Rfv3 have focused on the B6 Apobec3 allele, which exhibit properties such as higher levels of expression that make it more potent in vivo [11]-[16]. Here we show that the 129P2 Apobec3 allele did not promote recovery from viremia and NAb responses relative to Apobec3-null mice, consistent with Rfv3 susceptibility. Thus, the results resolve issues on the concordance between Apobec3 polymorphisms and Rfv3 genotype status with respect to the 129P2 strain, and further strengthen the case for Apobec3 as the gene encoded by Rfv3.
The lack of replication of B-tropic FV in Fv1 nr/nr 129P2 mice mirrored earlier studies involving N-tropic FV infection of Fv1 b/b mice (Table 2). These studies showed that N-tropic FV could function as a live-attenuated vaccine in an Fv1-incompatible host, resulting in the best immunological protection so far against FV infection [5]. Thus, Fv1 incompatibility may be an important mechanism to augment retrovirus-specific adaptive immune responses. However, these prior Fv1-restricted vaccine studies utilized mice that encode the B6 Apobec3 allele and are therefore Rfv3 resistant (Table 2). In other words, the resulting protective adaptive immune response may not be exclusively due to Fv1 restriction, as B6 Apobec3 is known to prime a stronger NAb response [7], [11], [12]. In addition, the reverse scenario involving B-tropic FV ‘vaccination’ of an Fv1 n/n host had not yet been performed. Thus, the potential contribution of B6 Apobec3 in prior Fv1 vaccine studies, and the lack of data on B-tropic FV as a live-attenuated vaccine, raise uncertainties on whether Fv1 restriction is a general retrovirus vaccine concept in mice.
We therefore performed a vaccination study to test if B-tropic FV can function as a live-attenuated vaccine. We utilized 129P2 Apobec3 KO mice to ensure that 129P2 Apobec3 will not contribute to the vaccine-elicited immune response. Even though the 129P2 Apobec3 allele did not promote NAb responses and recovery from FV infection, these phenotypes were observed in the context of pathogenic FV infection. Apobec3 restriction is saturable in vivo [41], and by 28 dpi, high replication levels may have overwhelmed a less potent 129P2 Apobec3 restriction phenotype [28]. Thus, 129P2 Apobec3 KO mice were ‘vaccinated’ with B-tropic FV and after 28 days, challenged with pathogenic, NB-tropic FV. Our results revealed that B-tropic FV replicated at near-undetectable levels by 28 dpi, induced significant cell-mediated and humoral immune responses and protected mice from pathogenic NB-tropic FV infection independent of Apobec3. Thus, Fv1 restriction was sufficient to induce immunological protection, strengthening the case for Fv1 incompatibility as a retrovirus vaccine paradigm in mice. Since Fv1 restriction prevented splenomegaly and promoted NAb responses in Fv2 and Rfv3 susceptible mice, the data also demonstrate that in 129P2 mice, Fv1 restriction is dominant over Fv2 resistance and Apobec3 restriction. This finding is consistent with a proposed ‘restriction factor hierarchy’ [37], whereby dominant resistance mechanisms may mask the impact of other restriction mechanisms in vivo.
The mechanism for how Fv1 incompatibility elicited potent adaptive immunity could readily be explained by live-attenuation: FV replicated at low levels, thereby inducing a protective immune response [43]–[47]. However, the quality of the immunological response induced by Fv1 incompatibility raise the question of whether signaling pathways that potentiate and/or amplify the adaptive immune response were also induced. Recently, HIV-1 capsid recognition by TRIM5α induced downstream AP-1 and NF-κB regulated genes that may orchestrate the adaptive immune response [48]. TRIM5α exhibited all the attributes of a pattern-recognition protein or innate sensor [48]. Nanotube reconstitution studies revealed that Fv1 recognizes MLV capsids in a lattice arrangement [49], analogous to TRIM5α. Thus, we speculate that Fv1 incompatibility may also trigger a similar cascade of immunity genes. A vaccine model involving B-tropic FV infection of 129P2 mice could prove useful in testing the importance of specific immunological pathways induced by capsid-dependent restriction in vivo. 129P2 mice offer advantages that include disease susceptibility that allows for straightforward pathogenic retrovirus challenge studies, and a genetic background that is extensively used in KO/transgenic technologies. It is conceivable that these Fv1 restriction studies in 129P2 mice may help direct approaches for harnessing TRIM5α biology for human retroviral vaccines.
Acknowledgments
We thank Warner Greene (J. David Gladstone Institutes) for the 129P2 Apobec3 KO mice, Kim Hasenkrug, Bruce Chesebro and Leonard Evans for the FV stocks, detection reagents and protocols, Brent Palmer and Kejun Guo (University of Colorado Denver) for flow cytometry and quantitative PCR advice, respectively, and the University of Colorado Denver Animal Core Facility for routine mouse colony maintenance.
Funding Statement
This work was supported by the National Institutes of Health R01 AI090795 (M.L.S), the Robert D. Watkins Predoctoral Fellowship of the American Society for Microbiology (K.H.), the Colorado Clinical and Translational Sciences Institute TL1 TR000155 (S.X.L.), the Tim Gill Foundation (M.S.H.), and the UCD Early Career Scholar Program (M.L.S.).The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Stremlau M, Owens CM, Perron MJ, Kiessling M, Autissier P, et al. (2004) The cytoplasmic body component TRIM5alpha restricts HIV-1 infection in Old World monkeys. Nature 427: 848–853. [DOI] [PubMed] [Google Scholar]
- 2. Sheehy AM, Gaddis NC, Choi JD, Malim MH (2002) Isolation of a human gene that inhibits HIV-1 infection and is suppressed by the viral Vif protein. Nature 418: 646–650. [DOI] [PubMed] [Google Scholar]
- 3. Friend C (1957) Cell-free transmission in adult Swiss mice of a disease having the character of a leukemia. J Exp Med 105: 307–318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Hasenkrug KJ, Chesebro B (1997) Immunity to retroviral infection: the Friend virus model. Proc Natl Acad Sci U S A 94: 7811–7816. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Halemano K, Harper MS, Guo K, Li SX, Heilman KJ, et al. (2012) Humoral immunity in the Friend retrovirus infection model. Immunol Res 55: 249–260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Luban J (2007) Cyclophilin A, TRIM5, and resistance to human immunodeficiency virus type 1 infection. J Virol 81: 1054–1061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Santiago ML, Montano M, Benitez R, Messer RJ, Yonemoto W, et al. (2008) Apobec3 encodes Rfv3, a gene influencing neutralizing antibody control of retrovirus infection. Science 321: 1343–1346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Yap MW, Mortuza GB, Taylor IA, Stoye JP (2007) The design of artificial retroviral restriction factors. Virology 365: 302–314. [DOI] [PubMed] [Google Scholar]
- 9. Chesebro B, Wehrly K (1979) Identification of a non-H-2 gene (Rfv-3) influencing recovery from viremia and leukemia induced by Friend virus complex. Proc Natl Acad Sci U S A 76: 425–429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Doig D, Chesebro B (1979) Anti-Friend virus antibody is associated with recovery from viremia and loss of viral leukemia cell-surface antigens in leukemic mice. Identification of Rfv-3 as a gene locus influencing antibody production. J Exp Med 150: 10–19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Tsuji-Kawahara S, Chikaishi T, Takeda E, Kato M, Kinoshita S, et al. (2010) Persistence of viremia and production of neutralizing antibodies differentially regulated by polymorphic APOBEC3 and BAFF-R loci in Friend virus-infected mice. J Virol 84: 6082–6095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Santiago ML, Smith DS, Barrett BS, Montano M, Benitez RL, et al. (2011) Persistent Friend virus replication and disease in Apobec3-deficient mice expressing functional B-cell-activating factor receptor. J Virol 85: 189–199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Takeda E, Tsuji-Kawahara S, Sakamoto M, Langlois MA, Neuberger MS, et al. (2008) Mouse APOBEC3 restricts friend leukemia virus infection and pathogenesis in vivo . J Virol 82: 10998–11008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Okeoma CM, Petersen J, Ross SR (2009) Expression of murine APOBEC3 alleles in different mouse strains and their effect on mouse mammary tumor virus infection. J Virol 83: 3029–3038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Langlois MA, Kemmerich K, Rada C, Neuberger MS (2009) The AKV murine leukemia virus is restricted and hypermutated by mouse APOBEC3 . J Virol 83: 11550–11559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Sanville B, Dolan MA, Wollenberg K, Yan Y, Martin C, et al. (2010) Adaptive evolution of Mus Apobec3 includes retroviral insertion and positive selection at two clusters of residues flanking the substrate groove. PLoS Pathog 6: e1000974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Abudu A, Takaori-Kondo A, Izumi T, Shirakawa K, Kobayashi M, et al. (2006) Murine retrovirus escapes from murine APOBEC3 via two distinct novel mechanisms. Curr Biol 16: 1565–1570. [DOI] [PubMed] [Google Scholar]
- 18. Browne EP, Littman DR (2008) Species-specific restriction of Apobec3-mediated hypermutation. J Virol 82: 1305–1313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Li J, Hakata Y, Takeda E, Liu Q, Iwatani Y, et al. (2012) Two genetic determinants acquired late in Mus evolution regulate the inclusion of exon 5, which alters mouse APOBEC3 translation efficiency. PLoS Pathog 8: e1002478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Eppig JT, Blake JA, Bult CJ, Kadin JA, Richardson JE (2012) The Mouse Genome Database (MGD): comprehensive resource for genetics and genomics of the laboratory mouse. Nucleic Acids Res 40: D881–886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Simpson EM, Linder CC, Sargent EE, Davisson MT, Mobraaten LE, et al. (1997) Genetic variation among 129 substrains and its importance for targeted mutagenesis in mice. Nat Genet 16: 19–27. [DOI] [PubMed] [Google Scholar]
- 22. Threadgill DW, Yee D, Matin A, Nadeau JH, Magnuson T (1997) Genealogy of the 129 inbred strains: 129/SvJ is a contaminated inbred strain. Mamm Genome 8: 390–393. [DOI] [PubMed] [Google Scholar]
- 23. Lilly F (1970) Fv-2: identification and location of a second gene governing the spleen focus response to Friend leukemia virus in mice. J Natl Cancer Inst 45: 163–169. [PubMed] [Google Scholar]
- 24. Persons DA, Paulson RF, Loyd MR, Herley MT, Bodner SM, et al. (1999) Fv2 encodes a truncated form of the Stk receptor tyrosine kinase. Nat Genet 23: 159–165. [DOI] [PubMed] [Google Scholar]
- 25. Nishigaki K, Hanson C, Jelacic T, Thompson D, Ruscetti S (2005) Friend spleen focus-forming virus transforms rodent fibroblasts in cooperation with a short form of the receptor tyrosine kinase Stk. Proc Natl Acad Sci U S A 102: 15488–15493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Jelacic TM, Thompson D, Hanson C, Cmarik JL, Nishigaki K, et al. (2008) The tyrosine kinase sf-Stk and its downstream signals are required for maintenance of Friend spleen focus-forming virus-induced fibroblast transformation. J Virol 82: 419–427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Beck JA, Lloyd S, Hafezparast M, Lennon-Pierce M, Eppig JT, et al. (2000) Genealogies of mouse inbred strains. Nat Genet 24: 23–25. [DOI] [PubMed] [Google Scholar]
- 28. Kolokithas A, Rosenke K, Malik F, Hendrick D, Swanson L, et al. (2010) The glycosylated Gag protein of a murine leukemia virus inhibits the antiretroviral function of APOBEC3. J Virol 84: 10933–10936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Best S, Le Tissier P, Towers G, Stoye JP (1996) Positional cloning of the mouse retrovirus restriction gene Fv1 . Nature 382: 826–829. [DOI] [PubMed] [Google Scholar]
- 30. Hartley JW, Rowe WP, Huebner RJ (1970) Host-range restrictions of murine leukemia viruses in mouse embryo cell cultures. J Virol 5: 221–225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Pincus T, Rowe WP, Lilly F (1971) A major genetic locus affecting resistance to infection with murine leukemia viruses. II. Apparent identity to a major locus described for resistance to Friend murine leukemia virus. J Exp Med 133: 1234–1241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Kozak CA, Chakraborti A (1996) Single amino acid changes in the murine leukemia virus capsid protein gene define the target of Fv1 resistance. Virology 225: 300–305. [DOI] [PubMed] [Google Scholar]
- 33. Jung YT, Kozak CA (2000) A single amino acid change in the murine leukemia virus capsid gene responsible for the Fv1(nr) phenotype. J Virol 74: 5385–5387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Rommelaere J, Donis-Keller H, Hopkins N (1979) RNA sequencing provides evidence for allelism of determinants of the N-, B- or NB-tropism of murine leukemia viruses. Cell 16: 43–50. [DOI] [PubMed] [Google Scholar]
- 35. Ou CY, Boone LR, Koh CK, Tennant RW, Yang WK (1983) Nucleotide sequences of gag-pol regions that determine the Fv-1 host range property of BALB/c N-tropic and B-tropic murine leukemia viruses. J Virol 48: 779–784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Stevens A, Bock M, Ellis S, LeTissier P, Bishop KN, et al. (2004) Retroviral capsid determinants of Fv1 NB and NR tropism. J Virol 78: 9592–9598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Barrett BS, Smith DS, Li SX, Guo K, Hasenkrug KJ, et al. (2012) A single nucleotide polymorphism in Tetherin promotes retrovirus restriction in vivo . PLoS Pathog 8: e1002596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Li K, Schuler T, Chen Z, Glass GE, Childs JE, et al. (2000) Isolation of Lactate dehydrogenase-elevating viruses from wild house mice and their biological and molecular characterization. Virus Res 67: 153–162. [DOI] [PubMed] [Google Scholar]
- 39. Robertson SJ, Ammann CG, Messer RJ, Carmody AB, Myers L, et al. (2008) Suppression of acute anti-Friend virus CD8+ T-cell responses by coinfection with Lactate dehydrogenase-elevating virus. J Virol 82: 408–418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Robertson MN, Miyazawa M, Mori S, Caughey B, Evans LH, et al. (1991) Production of monoclonal antibodies reactive with a denatured form of the Friend murine leukemia virus gp70 envelope protein: use in a focal infectivity assay, immunohistochemical studies, electron microscopy and western blotting. J Virol Methods 34: 255–271. [DOI] [PubMed] [Google Scholar]
- 41. Smith DS, Guo K, Barrett BS, Heilman KJ, Evans LH, et al. (2011) Noninfectious retrovirus particles drive the Apobec3/Rfv3 dependent neutralizing antibody response. PLoS Pathog 7: e1002284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Santiago ML, Benitez RL, Montano M, Hasenkrug KJ, Greene WC (2010) Innate retroviral restriction by Apobec3 promotes antibody affinity maturation in vivo . J Immunol 185: 1114–1123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Earl PL, Moss B, Morrison RP, Wehrly K, Nishio J, et al. (1986) T-lymphocyte priming and protection against Friend leukemia by vaccinia-retrovirus env gene recombinant. Science 234: 728–731. [DOI] [PubMed] [Google Scholar]
- 44. Dittmer U, Brooks DM, Hasenkrug KJ (1998) Characterization of a live-attenuated retroviral vaccine demonstrates protection via immune mechanisms. J Virol 72: 6554–6558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Dittmer U, Brooks DM, Hasenkrug KJ (1999) Requirement for multiple lymphocyte subsets in protection by a live attenuated vaccine against retroviral infection. Nat Med 5: 189–193. [DOI] [PubMed] [Google Scholar]
- 46. Dittmer U, Brooks DM, Hasenkrug KJ (1999) Protection against establishment of retroviral persistence by vaccination with a live attenuated virus. J Virol 73: 3753–3757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Messer RJ, Dittmer U, Peterson KE, Hasenkrug KJ (2004) Essential role for virus-neutralizing antibodies in sterilizing immunity against Friend retrovirus infection. Proc Natl Acad Sci U S A 101: 12260–12265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Pertel T, Hausmann S, Morger D, Zuger S, Guerra J, et al. (2011) TRIM5 is an innate immune sensor for the retrovirus capsid lattice. Nature 472: 361–365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Hilditch L, Matadeen R, Goldstone DC, Rosenthal PB, Taylor IA, et al. (2011) Ordered assembly of murine leukemia virus capsid protein on lipid nanotubes directs specific binding by the restriction factor, Fv1. Proc Natl Acad Sci U S A 108: 5771–5776. [DOI] [PMC free article] [PubMed] [Google Scholar]