

Abstract
Functional serotonin 3 (5-HT3) receptors are transiently expressed by cerebellar granule cells during early postnatal development, where they modulate short-term synaptic plasticity at the parallel fibre–Purkinje cell synapse. Here, we show that serotonin controls maturation of Purkinje cells in the mouse cerebellum. The 5-HT3 receptors regulate morphological maturation of Purkinje cells during early postnatal development, and this effect is mediated by the glycoprotein reelin. Using whole-cell patch-clamp recordings we also investigated physiological development of Purkinje cells in 5-HT3A receptor knockout mice during early postnatal development, and found abnormal physiological maturation, characterized by a more depolarized resting membrane potential, an increased input resistance and the ability to fire action potentials upon injection of a depolarizing current at an earlier age. Furthermore, short-term synaptic plasticity was impaired at both the parallel fibre–Purkinje cell and the climbing fibre–Purkinje cell synapses, and both the amplitude and the frequency of spontaneous miniature events recorded from Purkinje cells were increased. The expedited morphological and physiological maturation affects the whole cerebellar cortical network, as indicated by delayed climbing fibre elimination in 5-HT3A receptor knockout mice. There was no difference between wild-type and 5-HT3A receptor knockout mice in any of the morphological or physiological properties described above at later ages, indicating a specific time window during which serotonin regulates postnatal development of the cerebellum via 5-HT3 receptors expressed by granule cells.
Key points
Serotonin 3 (5-HT3) receptors are expressed by excitatory granule cells in the cerebellum during early postnatal development.
Here we show a novel role for serotonin in the regulation of cerebellar postnatal development via 5-HT3 receptors.
Using 5-HT3A receptor knockout mice we show that 5-HT3 receptors expressed by granule cells, via the glycoprotein reelin, regulate the morphological maturation of Purkinje cells.
The 5-HT3A receptor knockout mice show abnormal physiological maturation of Purkinje cells and impaired short-term plasticity at the parallel fibre–Purkinje cell synapse, resulting in delayed climbing fibre elimination.
With these results, we provide a better understanding of the role of serotonin in the developing brain, the control it has on the postnatal maturation of the cerebellum, and the cerebellum as a highly adaptive system during early postnatal development.
Introduction
Both the anatomical and the functional development of the rodent cerebellum occurs for a substantial part postnatally (Altman & Bayer, 1996). At birth, no cerebellum-dependent behaviour can be detected, and cells display an immature phenotype. During the first 3 weeks after birth, granule cells migrate from the external to the internal granule cell layer, and Purkinje cells develop their dendritic tree fully. Furthermore, connections between parallel fibres and Purkinje cells and between climbing fibres and Purkinje cells are formed into functional synapses during this time. The rodent cerebellum is physiologically mature by ∼4 weeks after birth (Altman, 1972a; van Welie et al. 2011).
Purkinje cells are the sole output of the cerebellar cortex to the deep cerebellar nuclei. The morphological and physiological maturation of Purkinje cells is therefore of special interest (Kapfhammer, 2004). McKay & Turner (2005) described the following three stages of Purkinje cell maturation in the rat: an initial stable immature stage of minimal change from postnatal day (P) 0 to P9; a transitional stage in which the Purkinje cells undergo major morphological and physiological maturation; and from P18, a stable adult stage with only minor refinements. Functional parallel fibre–Purkinje cell synapses are formed at the end of the first postnatal week (Altman, 1972b). Parallel fibres form synapses at the distal dendrites of Purkinje cells. Climbing fibres wrap around the soma and proximal dendrites of Purkinje cells and form their synapses there. In newborn rodents, a Purkinje cell is innervated by multiple climbing fibres, of which all but one will be removed during the first few postnatal weeks (Crepel et al. 1976). Parallel fibre input has a highly dominant role in climbing fibre elimination during development. In polyinnervated Purkinje cells, competition between different climbing fibres appears between P3 and P7 and continues during the second postnatal week (Scelfo & Strata, 2005).
Serotonin 3 (5-HT3) receptors are involved in postnatal maturation of pyramidal neurons in the cortex. Glutamatergic Cajal–Retzius cells express 5-HT3 receptors until the first two postnatal weeks (Chameau et al. 2009), during which they also synthesize and secrete the glycoprotein reelin (D’Arcangelo et al. 1999). We have shown that reelin regulates the maturation of apical, but not basal, dendrites of layer II/III pyramidal neurons in the somatosensory cortex in a 5-HT3 receptor-dependent manner. Specifically, the dendritic complexity of these neurons in the 5-HT3A receptor knockout (KO) mouse is increased, and the hypertrophy of dendritic arborization can be rescued by addition of recombinant reelin (Chameau et al. 2009).
Recently, we have shown that 5-HT3 receptors are transiently expressed on glutamatergic granule cells in the cerebellum during the first 3 weeks after birth (Oostland et al. 2011). Interestingly, this coincides with the time window during which Purkinje cell dendrites develop (Altman & Bayer, 1996). In addition, it has been shown that granule cells synthesize and secrete reelin (Sinagra et al. 2008). We therefore hypothesized that serotonin modulates morphological and physiological maturation of Purkinje cells via 5-HT3 receptors expressed on glutamatergic granule cells.
Methods
Ethical approval
Wild-type (WT) C57/Bl6 (Harlan Laboratories, Venray, the Netherlands) and 5-HT3A receptor knockout mice (Zeitz et al. 2002), both males and females, were used for this study between the age of P5 and P72. In total, 62 animals were used for this study. All experiments were performed in accordance with the committee on animal bioethics of the University of Amsterdam.
Electrophysiological recordings
For both single-cell electroporation and whole-cell patch-clamp recordings, animals were killed by decapitation between P5 and P72. Sagittal brain slices were cut using a vibrating blade microtome (Leica VT1200S) at a thickness of 300 μm. During slicing, the brains were kept in cooled (4°C) oxygenated artificial cerebrospinal fluid, which was composed of the following (mm): NaCl, 120; KCl, 3.5; CaCl2, 2.5; MgSO4, 1.3; NaH2PO4, 1.25; NaHCO3, 25; and glucose, 25; continuously bubbled with 95% O2 and 5% CO2 (pH 7.4). Brains from mice older than 3 weeks were sliced in modified artificial cerebrospinal fluid, composed of the following (mm): choline chloride, 120; KCl, 3.5; CaCl2, 0.5; MgSO4, 6.0; NaH2PO4, 1.25; glucose, 25; and NaHCO3, 25. During the experiments slices were kept submerged at room temperature and continuously superfused with artificial cerebrospinal fluid. Electroporation pipettes were pulled from boroscilicate glass and had a resistance between 7 and 12 MΩ. Pipettes were filled with biocytin hydrochloride (2–4 mg ml−1; Sigma) dissolved in internal solution containing (mm): potassium gluconate, 105; KCl, 30; EGTA, 5; CaCl2, 0.5; Hepes, 10; Mg-ATP, 5; (pH adjusted to 7.3 with KOH). The pipette was positioned in close proximity to the membrane of the Purkinje cell, and the membrane was permeabilized by electrical stimulation with an Axoporator 800A (Molecular Devices, Sunnyvale, CA, USA). Patch pipettes were pulled from borosilicate glass with a resistance of 2–3 MΩ and filled with internal solution as described above. Whole-cell recordings from Purkinje cells were made using an EPC9 patch-clamp amplifier and PULSE software (HEKA Electronic, Lambrecht, Germany). Signals were filtered at 1–5 kHz and sampled at 10 kHz. Series resistance ranged from 2 to 11 MΩ and was compensated for at least 70%. Cells were voltage clamped at −70 mV, corrected for liquid junction potential.
Action potential firing and input resistance were determined from whole-cell current-clamp recordings from Purkinje cells, during which a range of 12 currents was injected, with a duration of 1 s each and ranging from −100 to +175 pA. The number of Purkinje cells that showed more than one action potential during any of these current injection steps was scored. Input resistance was calculated from the change in membrane potential with a current injection of −25 pA, during which Purkinje cells did not fire action potentials. Resting membrane potential was calculated from the input resistance of the cell and the amount of current necessary to inject in order to maintain the cell at a holding potential of −70 mV using Ohm's law.
Miniature postsynaptic currents (mPSCs) from Purkinje cells in both WT and 5-HT3A receptor knockout mice were recorded in the voltage-clamp configuration at P7–P9 in the presence of 0.5 μm TTX (Latoxan, Valence, France) and analysed as described before (van Hooft, 2002). Per cell, one or two traces of 5 min each were used for analysis, with at least 100 miniature synaptic events per cell, which were visually verified. The distributions, average amplitudes and interevent intervals were analysed per cell. The distributions and average values for each cell were then averaged over all recorded cells to obtain the final distributions and values as shown in Fig. 5.
Figure 5. Increased frequency and amplitude of spontaneous miniature postsynaptic currents in Purkinje cells from 1-week-old 5-HT3A receptor knockout mice.
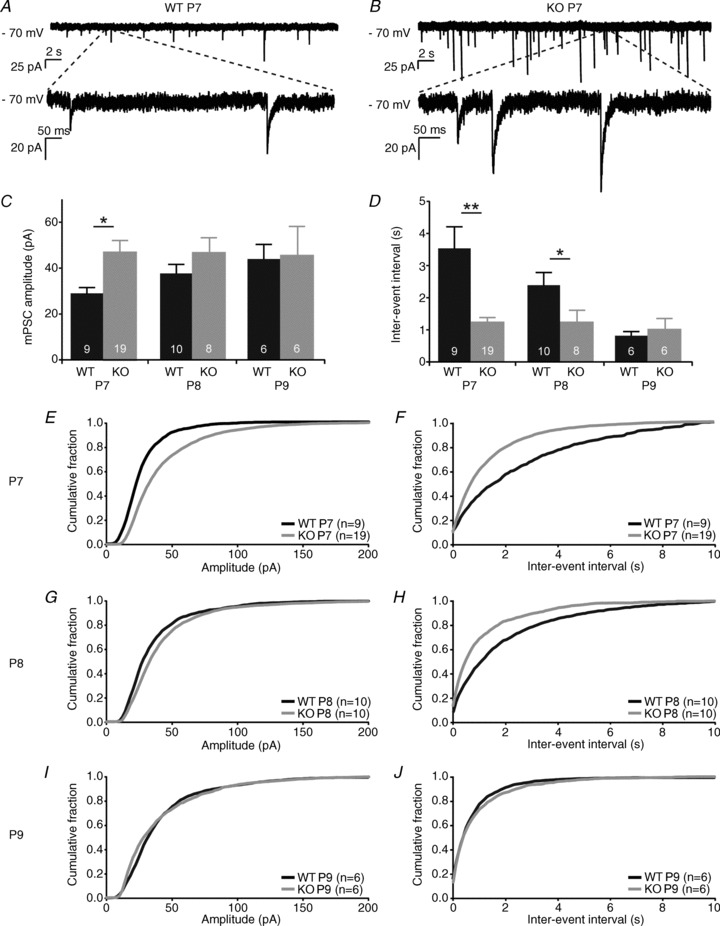
A, example trace of spontaneous miniature postsynaptic currents (mPSCs) recorded at −70 mV from Purkinje cells in P7 WT mice. B, similar example trace in P7 5-HT3A receptor knockout mice. C, amplitudes of mPSCs in Purkinje cells from WT and 5-HT3A receptor knockout mice indicate a significant difference at P7, but not at P8. D, average interevent intervals of the mPSCs recorded from Purkinje cells in WT and 5-HT3A receptor knockout mice indicate a difference at P7 and P8, but not at P9. E, G and I, amplitude distributions of mPSCs recorded from Purkinje cells show a significant difference between WT and 5-HT3A receptor knockout mice at P7. F, H and J, interevent interval distributions of mPSCs recorded from Purkinje cells show a significant difference between WT and 5-HT3A receptor knockout mice at P7 and P8. The numbers in the bars of the graphs indicate the number of cells used for analysis.
Glutamatergic synaptic currents in Purkinje cells were evoked by stimulation of either the parallel fibres or the climbing fibres using a glass electrode filled with artificial cerebrospinal fluid. Paired stimuli (100–400 μA, 0.2–1.0 ms duration, interstimulus interval 50 ms) were delivered either to the molecular layer or the internal granule cell layer using a custom-made isolated bipolar current stimulator. Paired-pulse stimulations were delivered with a 20 s interval, and only recordings which were stable for at least 15 min were used to analyse the paired-pulse ratio (PPR). The PPR was defined as the amplitude of the second EPSC divided by the amplitude of the first EPSC.
Organotypic slice cultures
Cell culture inserts (1 μm pore size; BD Falcon, Breda, the Netherlands) were coated with 66.7 μg ml−1 poly-l-lysine (Sigma) overnight at room temperature. Before use, inserts were rinsed three times with sterile H2O and wells were filled with 1.8 ml culture medium, consisting of Neurobasal-A medium containing 100 μg ml−1 penicillin–streptomycin and supplemented with either the serum-free supplements B27 (1:50 v/v), N2 (1:100 v/v) and 2 mm l-glutamine, or supplemented with 25% horse serum, 30 mm glucose and 2 mm glutamax (all from Invitrogen, Bleiswijk, the Netherlands). Cerebellar slice cultures treated with the N-terminal specific anti-reelin antibody G10 (Abcam, Cambridge, UK) at P8 during 2 days in vitro (Div) in culture medium with 25% horse serum gave similar results to those for the cultures without serum, indicating that addition of the serum did not change the reelin-mediated maturation process of Purkinje cells. Western blot analysis showed that horse serum did not contain detectable amounts of reelin (data not shown). Eight-day-old mice were killed by decapitation, after which brains were quickly dissected out and sagittal slices made as described above (see ‘Electrophysiological recordings’). Slices were washed three times in sterile culture medium and placed on inserts, and excessive medium was removed. Slices were cultured for 1–2 Div at 37°C in a humidified chamber containing air supplemented with 5% CO2. Culture medium from slices in the experimental conditions was supplemented G10 at a dilution of 1:1000 or the selective 5-HT3 antagonist, granisetron (100 nm; Sigma).
Morphological analysis
In order to reveal Purkinje cell morphology, Purkinje cells from lobules III–IX in the cerebellar vermis were filled with biocytin (Sigma; 2–4 mg ml−1 dissolved in internal solution as described above, see ‘Electrophysiological recordings’) during the whole-cell patch-clamp recordings or with biocytin hydrochloride (Sigma; 4 mg ml−1 dissolved in internal solution as described above, see ‘Electrophysiological recordings’) during single-cell electroporation. Slices were fixed overnight in paraformaldehyde (4% in 0.05 m PBS, pH 7.4) at 4°C and visualized using immunohistochemical methods. Slices were washed five times in PBS for 8 min, and endogenous peroxidase was inhibited by a 30 min incubation in H2O2 (3% in PBS). After 60 min permeabilization in 2% Triton X-100 (2% in PBS), slices were incubated for 2 h in avidin–biotin–peroxidase complex (Vectastain ABC Elite kit; Vector Laboratories, Peterborough, UK). Biocytin was visualized as a dark brown substrate using a DAB (3,3′-diaminobenzidine-tetrahydrochloride; Sigma) reaction. Slices were washed three times, each for 8 min, before being mounted on a glass slide with mowiol 4-88 (Sigma; dissolved in 0.2 m Tris–HCl and glycerol, pH 8.5). Images were made using a confocal microscope (Zeiss LSM 510) equipped with a ×20/0.75 objective and using the 543 nm line of an ArKr laser. These images were then used to make a three-dimensional reconstruction of the Purkinje cells for morphological analysis. For the reconstruction, ImageJ (National Institutes of Health, Bethesda, MD, USA; http://rsb.info.nih.gov/ij/) was used in combination with the NeuroMorpho plug-in. The different parameters were analysed using LMeasure software (Scorcioni et al. 2008). With these parameters, the dendritic complexity index (DCI) was calculated using the formula shown below, in which the branch tip order was defined for every branch tip as the number of branch points between the primary dendrite and the tip.
 |
Sholl analysis was performed on the reconstructed Purkinje cells by plotting the sum of the length of the dendrites in bins of 10 μm at P9 or 20 μm at P35 as a function of the distance from the soma using NeuronStudio software (http://research.mssm.edu/cnic/tools-ns.html).
Western blot
For quantification of reelin levels, we dissected the cerebellum from five WT and five 5-HT3A receptor knockout mice at P9 and homogenized these individually with a glass douncer in lysis buffer consisting of 320 mm sucrose, 10 mm Hepes and protease inhibitor mixture (Complete; pH 7.4; Roche, Almere, the Netherlands). The homogenates were centrifuged at 510g for 5 min at 4°C, and the supernatant was centrifuged again at 10,000g for 10 min at 4°C to discard undisrupted tissue. Samples were mixed with sample buffer, and proteins were denaturalized at 70°C during 10 min. In total, 20 μg of protein, quantified using a NanoDrop 2000 Spectrophotometer (Isogen Life Science, IJsselstein, the Netherlands), was loaded in triplicate and separated by SDS-PAGE in a 6% Tris–glycine gel and transferred onto a nitrocellulose membrane. The membranes were allowed to dry overnight and then processed for immunodetection of reelin. Non-specific binding sites on the nitrocellulose were blocked by immersion of the membranes in 4% non-fat dry milk (BioRad Laboratories, Veenendaal, the Netherlands) in Tris-buffered saline (20 mm Tris–HCl and 150 mm NaCl, pH 7.4) containing 0.1% (v/v) Tween-20 (TBST). Membranes were then incubated for 2 h at room temperature with mouse anti-reelin G10 antibody (Abcam; diluted 1:1000 in the blocking solution) and mouse anti-β-actin (Sigma; 1:4000). Blots were then rinsed three times in TBST and incubated for 1 h at room temperature with horseradish peroxidase-conjugated goat anti-mouse IgG (diluted 1:3000 in blocking solution). The protein bands were detected using the ECL Prime Western Blotting detection reagents (Amersham, Diegem, Belgium) and were visualized with the Odyssey 2800 (Li-Cor, Leusden, the Netherlands). Optical density (OD) levels of protein bands were quantified using ImageJ software, after which the reelin OD levels were compared with the OD levels of the β-actin bands within the same lane to obtain the OD ratio. At least two replicates were performed for each sample.
Statistical analysis
Values are expressed as means ± SEM. Comparisons were made using Student's unpaired t test unless stated otherwise. Fisher's exact test was used to compare frequency distributions between WT and 5-HT3A receptor knockout mice. Frequency and amplitude distributions of the miniature synaptic events were compared with a two-sample Kolmogorov–Smirnov test. A value of P < 0.05 was used to indicate a significant difference. Asterisks indicate P < 0.05 (*), P < 0.01 (**) and P < 0.001 (***).
Results
Serotonin 5-HT3 receptors control morphological maturation of Purkinje cells via reelin
Purkinje cells of both WT and 5-HT3A receptor knockout mice were filled with biocytin during whole-cell patch-clamp recordings to reveal their morphology. In 9-day-old mice, the DCI of Purkinje cells was 77 ± 24% (n = 20) higher in 5-HT3A receptor knockout mice than in WT mice (n = 15; P < 0.05; Fig. 1A and B). The total dendritic length of Purkinje cells in 5-HT3A receptor knockout mice was 35 ± 10% higher than in WT mice (P < 0.05). Sholl analysis indicated a significant difference in total dendritic length per concentric circle of 10 μm between WT and 5-HT3A receptor knockout mice in the area between 40 and 60 μm from the soma (P < 0.05; Fig. 1C). In 5-week-old mice, both the DCI (105 ± 15%; n = 13; Fig. 1D and E) and total dendritic length (103 ± 8%; n = 13) of Purkinje cells were not different from those in WT mice (n = 8), and Sholl analysis did not reveal topological differences (Fig. 1F).
Figure 1. Increased morphological complexity of Purkinje cells in 5-HT3A receptor knockout mice at postnatal day (P) 9 but not at P35.
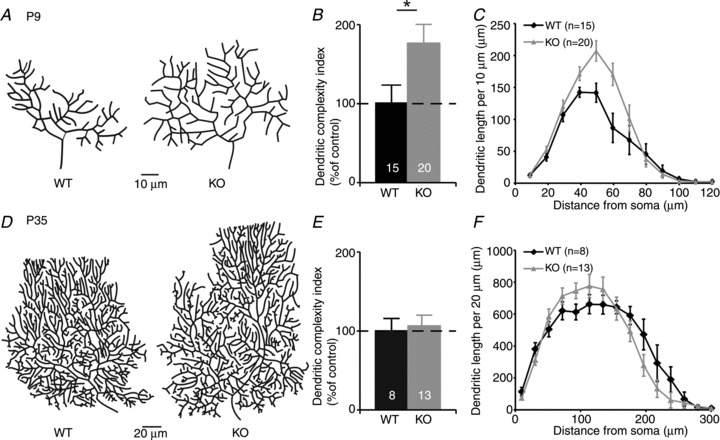
A, reconstructed Purkinje cells from wild-type (WT) and 5-HT3A receptor knockout (KO) mice at P9. B, dendritic complexity index of Purkinje cells at P9 indicates an increased dendritic complexity in 5-HT3A receptor knockout mice in comparison to WT mice. C, Sholl analysis indicates an increased dendritic length, specifically at 30–60 μm from the soma, in 5-HT3A receptor knockout mice at P9. D, reconstructed Purkinje cells from WT and 5-HT3A receptor knockout mice at P35. E, the dendritic complexity index of WT and 5-HT3A receptor knockout mice does not reveal any difference at P35. F, Sholl analysis does not show any topological difference between Purkinje cells from WT and 5-HT3A receptor knockout mice at P35.
The involvement of 5-HT3 receptors in controlling the maturation of Purkinje cells was corroborated in organotypic slice cultures. Blocking 5-HT3 receptors with 100 nm granisetron at P8 for 2 days in culture resulted in an increase of the DCI to 228 ± 42% (n = 6; P < 0.05; Fig. 2B and D). The total dendritic length was increased to 187 ± 14% (P < 0.01) after treatment with granisetron. Sholl analysis revealed a topological difference at 60–130 μm from the soma (P < 0.05; Fig. 2E).
Figure 2. Organotypic slice cultures show that morphological maturation of Purkinje cells is regulated by 5-HT3 receptors via the glycoprotein reelin.
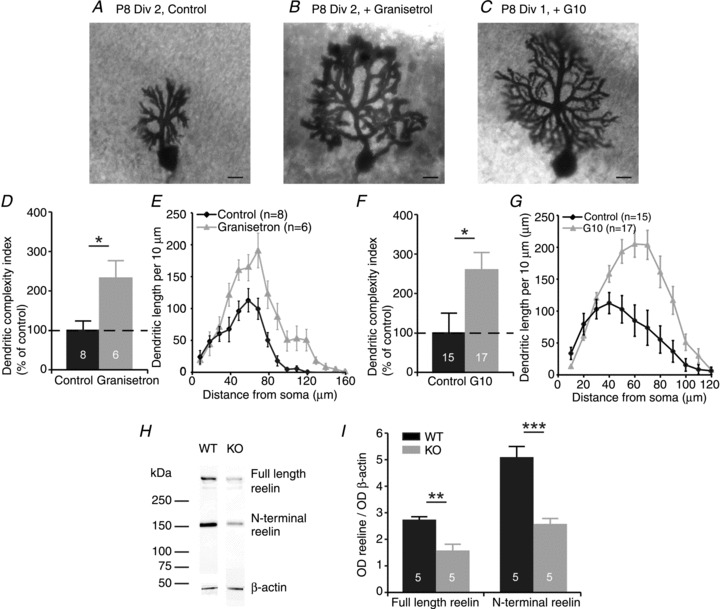
A, example of a P8 Purkinje cell in control conditions after 2 days in culture (Div). B, example of a P8 Purkinje cell after 2 days in culture with 100 nm granisetron, a selective 5-HT3 receptor antagonist. C, example of a P8 Purkinje cell after 2 days in culture with G10, an antibody against the N-terminal of reelin. Scale bars in A–C indicate 10 μm. D, the dendritic complexity index of Purkinje cells from cerebellar slices cultured at P8 after 2 days in culture shows an increased complexity of the Purkinje cells treated during the culture period with granisetron. E, Sholl analysis indicates an increase in dendritic length in granisetron-treated Purkinje cells in the area 60–130 μm from the soma. F, the dendritic complexity index from Purkinje cells from cerebellar slices cultured at P8 after 1 day in culture shows an increased complexity of the Purkinje cells treated during the culture period with G10. G, Sholl analysis shows an increase in dendritic length between 40 and 100 μm from the soma in the culture conditions with G10. H, Western blots for whole-cerebellum homogenates from P9 WT and 5-HT3A receptor knockout mice showing protein bands for full-length reelin, the N-terminal of reelin, and β-actin. I, quantification of Western blot analysis indicates that the level of optical density (OD) of reelin is reduced in 5-HT3A receptor knockout mice at P9. The numbers in the bars of the graphs indicate the number of cells used for analysis.
Given the fact that 5-HT3 receptors are expressed by reelin-producing granule cells, we next investigated whether reelin is involved in the dendritic maturation of Purkinje cells. After blocking the N-terminal region of reelin at P8 during 1 day in vitro in serum-free culture medium by treatment with anti-reelin antibody G10, the DCI was increased to 260 ± 44% (n = 17; P < 0.05; Fig. 2C and F) and the total dendritic length to 185 ± 16% (P < 0.01). Sholl analysis indicated a significant difference in dendritic length 40–100 μm from the soma (P < 0.01; Fig. 2G). Western blots were used to detect reelin protein levels in WT (n = 5) and 5-HT3A receptor knockout mice (n = 5) at P9 and revealed that the levels of both full-length reelin (P < 0.01) and the N-terminal of reelin (P < 0.001) were reduced in 5-HT3A receptor knockout mice (Fig. 2H and I).
Four different stages of morphological development were defined in 1-week-old Purkinje cells (Fig. 3A), comparable to stages II–V as described by Altman (1972a). Stage I is defined as the most immature stage, in which Purkinje cells have multiple primary dendrites in all directions. Stage II is a stage in which Purkinje cells still have multiple primary dendrites, but which are already polarized into one direction. Purkinje cells in stage III have only one primary dendrite, with an immature dendritic tree. Stage IV is the last stage before full maturation, in which Purkinje cells have one primary dendrite with an almost fully developed dendritic tree. Purkinje cells from 5-HT3A receptor knockout mice were in a more mature morphological stage than Purkinje cells from WT mice (Fisher's exact test: at P7, P < 0.001; P8, P < 0.001; and P9, P < 0.001; Fig. 3B). Taken together, these results indicate that there is a specific time window in 5-HT3A receptor knockout mice during which Purkinje cell dendritic tree morphology is altered.
Figure 3. Expedited morphological development of Purkinje cells in 5-HT3A receptor knockout mice.
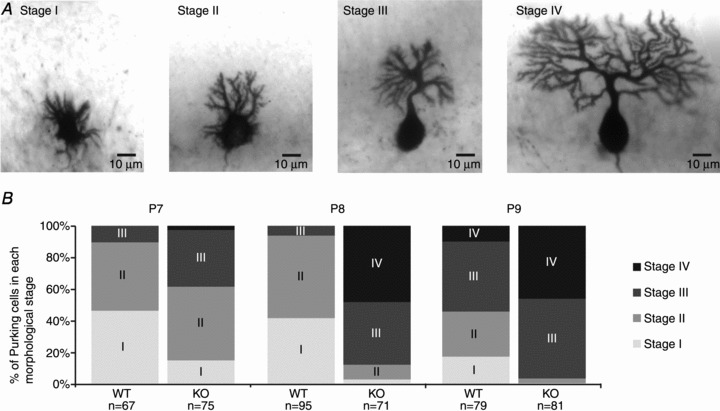
A, Purkinje cells from P8 mice show the following four stages of morphological maturation: stage I is the most immature stage, in which Purkinje cells have multiple primary dendrites in all directions; stage II is a stage in which Purkinje cells still have multiple primary dendrites, but which are already polarized into one direction; stage III are Purkinje cells which have one primary dendrite with a very immature dendritic tree; and stage IV is the last stage before the final maturation, in which Purkinje cells have one primary dendrite with an almost fully developed dendritic tree. B, distribution of Purkinje cells in each morphological maturation stage in both WT and 5-HT3A receptor knockout mice at P7, P8 and P9 shows that Purkinje cells from 5-HT3A receptor knockout mice display a more mature morphology at an earlier age than Purkinje cells from WT mice. The numbers underneath the bars of the graphs indicate the number of cells used for analysis.
Abnormal physiological maturation of Purkinje cells in 5-HT3A receptor knockout mice
Given the expedited morphological maturation of Purkinje cells in 5-HT3A receptor knockout mice, we hypothesized that the physiological properties of Purkinje cells show a concurrent increase in speed of maturation. At P6–P9, Purkinje cells from 5-HT3A receptor knockout mice had a more depolarized resting membrane potential (−46.6 ± 2.4 mV) than Purkinje cells in WT mice (−60.7 ± 1.7 mV; P < 0.001; Fig. 4A and B) and a higher input resistance (KO, 760.7 ± 95.9 MΩ; and WT, 136.0 ± 16.1 MΩ; P < 0.0001; Fig. 4C). In addition, at this age, the percentage of Purkinje cells firing action potentials upon a depolarizing current injection was significantly higher in 5-HT3A receptor knockout mice (80%; n = 25) than in WT mice (23%; n = 22; Fisher's exact test, P < 0.001; Fig. 4D). At P10–P12, there was no difference between Purkinje cells in WT (n = 29) and 5-HT3A receptor knockout mice (n = 20) with respect to resting membrane potential or the percentage of cells firing repetitive action potentials upon a depolarizing current injection (Fig. 4E–H), which was comparable to previous studies (Crepel et al. 1987; Doughty et al. 1999; McKay & Turner, 2005; Fry, 2006). However, the input resistance in 5-HT3A receptor knockout mice (271.0 ± 41.4 MΩ) was still significantly higher than that in WT mice (170.7 ± 18.8 MΩ; P < 0.05; Fig. 4G). Both the resting membrane potential (−69.0 ± 1.9 mV) and the input resistance (182.8 ± 32.1 MΩ) in 8- to 10-week-old 5-HT3A receptor knockout mice (n = 18) were not different from those in WT (−69.2 ± 0.5 mV and 164.9 ± 21.8 MΩ, respectively; n = 17).
Figure 4. Purkinje cells in 5-HT3A receptor knockout mice display more advanced physiological maturation.
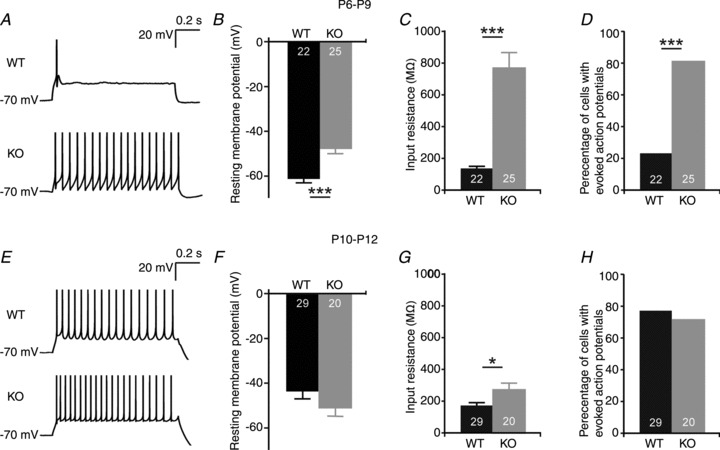
A, example traces showing action potential firing upon a depolarizing current injection of +175 pA in current-clamp recordings from Purkinje cells in WT and 5-HT3A receptor knockout mice at P6–P9. B, Purkinje cells from 5-HT3A receptor knockout mice show a more depolarized resting membrane potential than Purkinje cells from WT mice at P6–P9. C, input resistance of Purkinje cells from 5-HT3A receptor knockout mice at P6–P9 is increased when compared with WT mice. D, percentages of Purkinje cells with evoked action potentials in response to a depolarizing current injection of +175 pA show that more Purkinje cells from 5-HT3A receptor knockout mice display this physiological feature than Purkinje cells from WT mice at P6–P9. E, example traces showing action potential firing upon a depolarizing current injection of +175 pA in current-clamp recordings from Purkinje cells in WT and 5-HT3A receptor knockout mice at P10–P12. At P10–P12, there is no difference between Purkinje cells from WT and Purkinje cells from 5-HT3A receptor knockout mice in the resting membrane potential (F) or in the percentage of cells firing action potentials upon a depolarizing current injection of +175 pA (H), indicating an advanced physiological maturation of Purkinje cells in 5-HT3A receptor knockout mice only during a specific period of postnatal development. Input resistance of Purkinje cells was further decreased in 5-HT3A receptor knockout mice at P10–P12 but still significantly different from the input resistance of Purkinje cells in WT mice (G). The numbers in the bars of the graphs indicate the number of cells used for analysis.
In order to assess the maturation of the input of Purkinje cells, mPSCs from whole-cell voltage-clamped Purkinje cells were recorded in WT (Fig. 5A) and 5-HT3A receptor knockout mice (Fig. 5B) between P7 and P9. Average amplitudes and interevent intervals of the mPSCs were compared using the Mann–Whitney U test. In WT mice, the average amplitude of the mPSCs increased with age, showing a developmental pattern. In 5-HT3A receptor knockout mice, the amplitude of the mPSCs was already high at P7 (P < 0.05 compared with WT P7) and did not show a further increase with age (Fig. 5C). The interevent interval of the mPSCs recorded from Purkinje cells in WT mice showed a developmental pattern between P7 and P9, revealing a decrease in the interevent interval and thus an increase in frequency of the mPSCs at a later age. The average interevent interval of the mPSCs in 5-HT3A receptor knockout mice was already low at P7 (P < 0.01 compared with WT P7) and remained low at P8 (P < 0.05 compared with WT P8) and P9 (n.s.; Fig. 5D). Amplitude distributions were significantly different between WT and 5-HT3A receptor knockout mice at P7 (Fig. 5E; P < 0.001), but not at P8 (Fig. 5G) and P9 (Fig. 5I). Interevent interval distributions were significantly different between WT and 5-HT3A receptor knockout mice at P7 and P8 (Fig. 5F and H; both P < 0.001), but not at P9 (Fig. 5J).
Delayed climbing fibre elimination in 5-HT3A receptor knockout mice
Given that the frequency of mPSCs in Purkinje cells from 5-HT3A receptor knockout mice is increased, we wondered whether this was due to an increase in parallel fibre inputs, climbing fibre inputs, or both. We have previously shown that acute block of presynaptic 5-HT3 receptors at the parallel fibre–Purkinje cell synapse converts this synapse from a facilitating into a depressing one (Oostland et al. 2011). This finding was corroborated in 5-HT3A receptor knockout mice at P8, reflected by a decreased PPR upon parallel fibre stimulation in comparison to WT mice (P < 0.05; Fig. 6Aa, Ab and C). At P21, the PPR in WT mice (Fig. 6Ba and C) and in 5-HT3A receptor knockout mice (Fig. 6Bb and C) did not differ and showed a facilitating ratio, which is a known feature of the mature parallel fibre–Purkinje cell synapse.
Figure 6. Short-term synaptic plasticity at both the parallel fibre–Purkinje cell synapse and at the climbing fibre–Purkinje cell synapse is impaired in 5-HT3A receptor knockout mice.
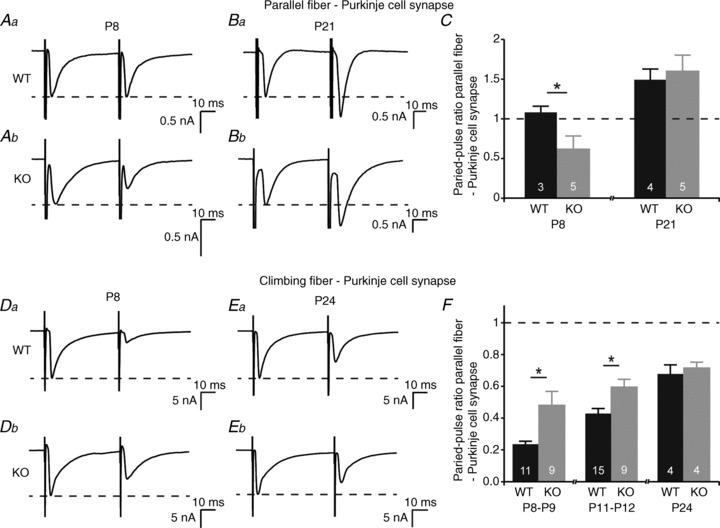
A–C, paired-pulse recordings from parallel fibre–Purkinje cell synapses at P8 in WT mice (Aa) and in 5-HT3A receptor knockout mice (Ab) and at P21 in WT mice (Ba) and in 5-HT3A receptor knockout mice (Bb). C, the paired-pulse ratio at the parallel fibre–Purkinje cell is reduced in 5-HT3A receptor knockout mice at P8, but not at P21. D–F, paired-pulse recordings from climbing fibre–Purkinje cell synapses at P8 in WT mice (Da) and in 5-HT3A receptor knockout mice (Db) and at P24 in WT mice (Ea) and in 5-HT3A receptor knockout mice (Eb). F, the paired-pulse ratio at the climbing fibre–Purkinje cell synapse is increased in 5-HT3A receptor knockout mice at P8–P9 and at P11–P12, but there is no difference at P24. The numbers in the bars of the graphs indicate the number of cells used for analysis.
We next investigated the short-term synaptic transmission at the climbing fibre–Purkinje cell synapse by means of paired-pulse stimulation. The PPR at the climbing fibre–Purkinje cell synapse in WT mice increased with age, indicating maturation of the climbing fibre–Purkinje cell synapse (Fig. 6Da, Ea and F). Compared with WT mice, in 5-HT3A receptor knockout mice the PPR was higher at P8–P9 (P < 0.05; Fig. 6Db and F) and at P11–P12 (P < 0.05; Fig. 6F) and showed a similar level to the PPR in WT mice at P24 (Fig. 6Eb and F).
During early postnatal development there is a competition between parallel fibres and climbing fibres for the available postsynaptic sites on Purkinje cells. As the parallel fibre–Purkinje cell synapse is impaired in 5-HT3A receptor knockout mice, we hypothesized that this might favour the total number of climbing fibres. Whole-cell patch-clamp recordings of Purkinje cells were made, and climbing fibres were activated with electrical stimulations in the internal granule cell layer. A climbing fibre EPSC (Fig. 7A) was defined as an EPSC which showed a stepwise change in amplitude in response to a gradual increase in stimulus intensity (Fig. 7B), in combination with paired-pulse depression (as seen in Fig. 6D–F). We found that the number of climbing fibres by which a Purkinje cell is innervated decreases over time in both WT and 5-HT3A receptor knockout mice (Fig. 7C). At P5 and P6 there is an average of four climbing fibres per Purkinje cell in both WT and 5-HT3A receptor knockout mice. In WT mice, there is a transitional stage between P7 and P12 during which the number of climbing fibres gradually decreases to just over one climbing fibre per Purkinje cell. This number is reduced to exactly one climbing fibre per Purkinje cell at P24 in all cells recorded. In 5-HT3A receptor knockout mice, this transitional stage of climbing fibre elimination lasts longer, with multiple climbing fibres still innervating one Purkinje cell at P24 (P < 0.001, Mann–Whitney U test). In 10-week-old (P68–P72) 5-HT3A receptor knockout mice, the number of climbing fibres innervating a single Purkinje cell was eliminated to 1.18 ± 0.10 (n = 17). Thus, climbing fibre elimination is delayed in 5-HT3A receptor knockout mice.
Figure 7. Climbing fibre elimination is delayed in 5-HT3A receptor knockout mice.
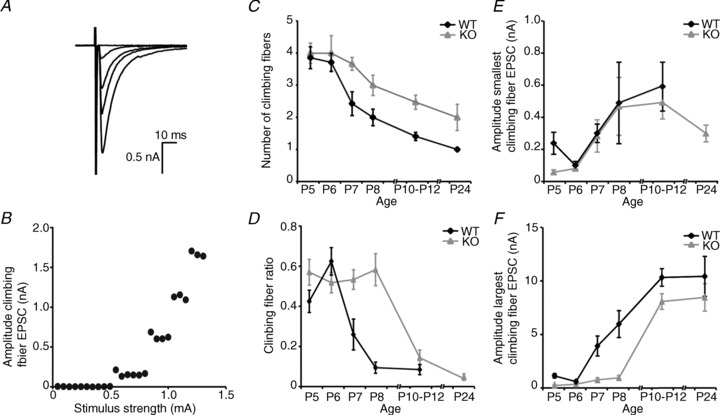
A, example of a recording from a Purkinje cell with inputs from four different climbing fibres. B, input–output graph of the same recording as in A, which shows a stepwise increase in amplitude of the climbing fibre EPSC upon a gradual increase in stimulus strength, indicating four climbing fibres innervating the recorded Purkinje cell. C, number of climbing fibres in WT and 5-HT3A receptor knockout mice between P5 and P24 shows a delay in climbing fibre elimination in 5-HT3A receptor knockout mice. D, the climbing fibre ratio, defined as the amplitude of one climbing fibre EPSC divided by the amplitude of the largest climbing fibre EPSC within the same Purkinje cell, is increased in 5-HT3A receptor knockout mice at P7 and P8. E, there is no difference in the amplitude of the smallest climbing fibre EPSC between WT and 5-HT3A receptor knockout mice at P6–P12. Only at P5 is the amplitude of the smallest climbing fibre EPSC smaller in 5-HT3A receptor knockout mice. At P24 there was only one climbing fibre innervating each Purkinje cell in WT mice, which we counted as the climbing fibre with the largest climbing fibre EPSC and thus there is no value for the amplitude of the smallest climbing fibre EPSC in P24 WT mice. F, the amplitude of the largest climbing fibre EPSC is decreased at P5, P7 and P8 in 5-HT3A receptor knockout mice, accounting for the increase in climbing fibre ratio as shown in C.
We additionally analysed the amplitudes of the smallest and the largest climbing fibre EPSC and the climbing fibre ratio. The amplitude of a single climbing fibre EPSC was calculated as the difference in amplitude between any EPSC and the previous recorded EPSCs. The climbing fibre ratio was defined as the amplitude of any given climbing fibre EPSC divided by the amplitude of the largest climbing fibre EPSC synapsing onto the same Purkinje cell. The number of climbing fibres (n) for which the climbing fibre ratio can be determined is n – 1 for each Purkinje cell. Wild-type mice at P24 had already completed climbing fibre elimination, and therefore there is no climbing fibre ratio or amplitude of the minimal climbing fibre EPSC. The climbing fibre ratio remained higher for a prolonged time in 5-HT3A receptor knockout mice compared with WT mice (Fig. 7D). The minimal amplitude of the climbing fibre EPSC (Fig. 7E) was significantly different at P5, but not at other ages. The maximal amplitude of the climbing fibre EPSC (Fig. 7F) was different at young ages, but not in older animals.
It thus seems that in 5-HT3A receptor knockout mice there is not only expedited morphological and physiological maturation of Purkinje cells, but also the two glutamatergic inputs to the Purkinje cells are affected, as shown by impairment of both the parallel fibre and the climbing fibre input, and delayed climbing fibre elimination.
Discussion
The results in this study show a novel role for serotonin in the regulation of cerebellar postnatal development. This is mediated via 5-HT3 receptors expressed by excitatory granule cells, which regulate the morphological and physiological maturation of Purkinje cells. The serotonergic modulation via 5-HT3 receptors also affects the surrounding cerebellar cortical microcircuit, as indicated by impaired short-term synaptic plasticity at the climbing fibre–Purkinje cell synapse and delayed climbing fibre elimination in 5-HT3A receptor knockout mice.
The role of 5-HT3 receptors in spontaneous miniature events recorded from Purkinje cells
Recently, we have shown that acute pharmacological blockade of presynaptic 5-HT3 receptors leads to a reduction in frequency of mPSCs recorded from Purkinje cells from P9 WT mice (Oostland et al. 2011). In the present study, we show that the frequency of mPSCs recorded from Purkinje cells from P7 5-HT3A receptor knockout mice was increased in comparison to P7 WT mice. Thus, recordings from Purkinje cells in WT mice during pharmacological blockade of 5-HT3 receptors for a short amount of time and recordings from Purkinje cells from 5-HT3A receptor knockout mice give contradictory results. The increase in mPSC frequency reflects changes in both the parallel fibre–Purkinje cell synapse and the climbing fibre–Purkinje cell synapse. We have shown before that the parallel fibre–Purkinje cell synapse is impaired and turns into a depressing synapse after pharmacological blockade of 5-HT3 receptors (Oostland et al. 2011). In the present study, we show a similar effect on the parallel fibre–Purkinje cell synapse in 5-HT3A receptor knockout mice. Morphological and electrophysiological data demonstrate that the development of parallel fibre synapses is significantly correlated with the time course of the climbing fibre elimination (Scelfo & Strata, 2005). Given that the increase in mPSC frequency in 5-HT3A receptor knockout mice may reflect a change in both parallel fibre and climbing fibre input, that the parallel fibre input is impaired in 5-HT3A receptor knockout mice and that the parallel fibre input is necessary for climbing fibre elimination because of the competitive nature of this process, we hypothesized that the increase in mPSC frequency recorded from Purkinje cells from 5-HT3A receptor knockout mice results from an increase in climbing fibre input owing to delayed climbing fibre elimination. This can explain the contradictory results described above, because pharmacologically blocking 5-HT3 receptors for a short amount of time during recording does not affect climbing fibre elimination but only affects functioning of the parallel fibre–Purkinje cell synapse, leading to a decrease in mPSC frequency recorded from Purkinje cells (Oostland et al. 2011). Alternatively, an increase in parallel fibre–Purkinje cell synapse formation in 5-HT3A receptor knockout mice could account for the increase in mPSC frequency, although this would be accompanied by an increase in climbing fibre elimination, opposite to what we have observed.
Mechanisms of delayed climbing fibre elimination in 5-HT3A receptor knockout mice
In this study, we show that climbing fibre elimination is delayed in 5-HT3A receptor knockout mice until at least 3 weeks after birth, and that this process is normalized between P24 and 10 weeks postnatally. The course of climbing fibre elimination in the WT mice we have found in the present study is comparable to what has been described before (reviewed by Watanabe & Kano, 2011). Climbing fibre elimination is a developmental process that occurs in two stages. The early stage is characterized by homosynaptic competition between climbing fibres, and lasts up to P8 in rats (Crepel et al. 1981). The late phase consists of heterosynaptic competition between parallel fibres and climbing fibres, and typically occurs between P9 and P17 in rats (Crepel et al. 1981). In the present study, we find no difference in the number of climbing fibres innervating one Purkinje cell between WT and 5-HT3A receptor knockout mice at P5 and P6. Only from P7 onwards does climbing fibre elimination start to be delayed in the 5-HT3A receptor knockout mice, resulting in multiple climbing fibre innervation in 3-week-old 5-HT3A receptor knockout mice. Given that our study was carried out in mice, whose developmental pattern is up to 2 days earlier than in rats, this period of delayed climbing fibre elimination is in concordance with the late phase as described by Crepel et al. (1981) in rats. Thus, the delayed climbing fibre elimination we find in the 5-HT3A receptor knockout mice occurs during the period of heterosynaptic competition between parallel fibres and climbing fibres. This supports our hypothesis that presynaptic 5-HT3 receptors at the parallel fibre terminal affect the climbing fibre elimination. However, we cannot exclude the possibility that a 5-HT3 receptor knockout-unrelated change in climbing fibre elimination is involved.
Parallel fibre inputs to Purkinje cells can activate the mGluR1 receptor, which via the Gαq (Offermans et al. 1997), phsopholipase Cβ4 (Kano et al. 1998) and the γ isoform of protein kinase C (Kano et al. 1995) signalling cascade in Purkinje cells plays a central role in elimination of surplus climbing fibre synapses, through still unknown mechanisms (Kano et al. 2008). The effects on behaviour of an increased number of climbing fibres during early postnatal development are unclear. It is known that persistent polyinnervation of Purkinje cells by climbing fibres leads to problems in motor learning in (young) adult mice (i.e. Kimpo & Raymond, 2007). Other mouse models with delayed but not completely impaired climbing fibre elimination, such as the αCaMKII knockout mouse line, do show motor learning deficits in adult mice (Hansel et al. 2006). In these αCaMKII knockout mice, 49% of Purkinje cells were innervated by two climbing fibres at P21–P28, while climbing fibre elimination was fully completed in adult mutants. However, these motor learning deficits can most probably be explained by a specific impairment of induction of long-term depression in Purkinje cells. As 5-HT3 receptors in the cerebellum are not expressed after 3 weeks postnatally, we do not expect physiological or behavioural deficits in the 5-HT3A receptor knockout mice, and cerebellum-specific motor tests cannot be performed in mice younger than 3 weeks old.
Involvement of 5-HT3 receptors in Purkinje cell development
Physiological and morphological postnatal development of Purkinje cells has been closely examined in the past decade (reviewed by Kapfhammer, 2004). Purkinje cells from 5-HT3A receptor knockout mice have different electrophysiological properties from Purkinje cells in WT mice. They are able to fire action potentials upon a depolarizing current injection of +175 pA at an earlier age and have a more depolarized resting membrane potential than in WT mice. Furthermore, input resistance is increased in 5-HT3A receptor knockout mice. Data on the input resistance of Purkinje cells during postnatal development are inconsistent. McKay & Turner (2005) found an input resistance of Purkinje cells that decreased with age between P0 and P18, recorded in slices from rats. On the contrary, Fry (2006) found in dissociated Purkinje cells from mice a decrease in input resistance from P5 to P7, followed by an increase between P11 and P18.
In addition to the role in electrophysiological maturation, we showed that 5-HT3 receptors regulate dendritic maturation of Purkinje cells via the glycoprotein reelin, and we investigated this further by looking at the stage of morphological maturation between P7 and P9 in more detail. We defined four different stages of Purkinje cell dendritic development (see also Altman, 1972a), and the results from the present study show that in 5-HT3A receptor knockout mice the Purkinje cells have a more mature morphology at an earlier age than the Purkinje cells in WT mice, indicating advanced morphological maturation. However, we were unable to detect any morphological differences between Purkinje cells in WT and 5-HT3A receptor knockout mice at 5 weeks of age, showing that the dendritic morphology was normalized by that age. Thus, the serotonergic system, modulated by 5-HT3 receptors and reelin, controls the time course of maturation in Purkinje cells.
In this study, we have used 5-HT3A receptor knockout mice, in which transgenic modifications are not restricted to one particular cell type or time window. The effects we have shown in this study could be affected by compensatory mechanisms. However, to the best of our knowledge the granule cell is the only cell type in the cerebellum to express 5-HT3 receptors (Oostland et al. 2011). In order to investigate the morphology of Purkinje cells in a more time-specific manner, we used organotypic slice cultures, in which we blocked 5-HT3 receptors or reelin at P8 for 1 or 2 days. Using this time-specific method, we found similar effects as in 5-HT3A receptor knockout mice at P9.
5-HT3 receptor-dependent regulation of dendritic morphology in cerebral cortex and cerebellum
Our group has shown before that 5-HT3 receptors affect morphology of layer II/III pyramidal neurons in the somatosensory cortex both during early postnatal development and at P90 (Chameau et al. 2009). Furthermore, in P14–P21 mice, pyramidal neurons with a more complex dendritic tree were found to have different electrophysiological properties, i.e. reduced spike frequency adaptation (van der Velden et al. 2012). Thus, the serotonergic system has a sustained effect on morphological properties of pyramidal neurons in the somatosensory cortex, while in the cerebellum the serotonergic system more specifically affects the time course of both morphological and physiological maturation. Interestingly, in both brain areas the 5-HT3 receptors on glutamatergic cells are present only during the first 3 weeks postnatally, and in both cases the dendritic maturation is regulated via reelin (Chameau et al. 2009; Oostland et al. 2011). However, it is only in the cerebellum that the 5-HT3 receptor-dependent regulation of dendritic morphology is limited to the early postnatal stages of development.
Concluding remarks
The role of the cerebellum has in recent years been extended from motor co-ordination and motor learning to involvement in neurodevelopmental disorders, such as autism and schizophrenia. This cognitive function of the cerebellum is mediated by amongst others reelin and serotonin. In the cerebellum, decreased levels of reelin mRNA and protein have been found in brains from patients with autism (reviewed by Fatemi et al. 2005). Alterations in expression and distribution of serotonin receptors in the cerebellum have been implicated in schizophrenia (Slater et al. 1998; Eastwood et al. 2001). In order to gain a better understanding of the development of autism and schizophrenia and the role of the cerebellum therein, it is of importance to understand the role of reelin and serotonin in postnatal development of the cerebellum and its projections to the cerebral cortex. Efferent projections from Purkinje cells to the cerebellar and vestibular nuclei occur during late embryogenesis, and synaptic contacts are established around birth (Eisenman et al. 1991). The exact timing of the development of projections from the cerebellar nuclei to the thalamus is not known, but it is likely that these connections are made before P21, because it is during this neonatal and juvenile period that systematic changes take place and the cerebellar network is formed (Altman & Bayer, 1996). Thus, the involvement of serotonin and reelin via the transiently expressed 5-HT3 receptors in postnatal Purkinje cell development during this critical period, even though we show in the present study normalization of climbing fibre input in young adult 5-HT3A receptor knockout mice, could be functionally relevant in neurodevelopmental disorders by changing Purkinje cell output. Future studies focused on the development of efferent pathways from Purkinje cells to the cerebral cortex and the role of serotonin therein could shed more light on this topic.
Acknowledgments
This research was supported in part by the Research Council for Earth and Life Sciences (ALW) with financial aid from The Netherlands Organization for Scientific Research (NWO). We thank David Julius (University of San Fransisco, CA, USA) for providing the 5-HT3A receptor knockout mice, and Ronald Breedijk from the van Leeuwenhoek Centre for Advanced Microscopy (University of Amsterdam, The Netherlands) for use of the confocal microscope. We thank Wytse Wadman and Pascal Chameau for their support and comments on the manuscript.
Glossary
- DAB
3,3′-diaminobenzidine-tetrahydrochloride
- DCI
dendritic complexity index
- Div
days in vitro
- KO
knockout
- mPSC
miniature postsynaptic current
- OD
optical density
- P
postnatal day
- PPR
paired-pulse ratio
- WT
wild-type
Author contributions
M.O. and M.R.B. performed the experiments and analysed the data with help of J.A.v.H.; M.O. and J.A.v.H. wrote the manuscript. All authors approved the final version of the manuscript.
References
- Altman J. Postnatal development of the cerebellar cortex in the rat. II. Phases in the maturation of Purkinje cells and of the molecular layer. J Comp Neurol. 1972a;145:399–463. doi: 10.1002/cne.901450402. [DOI] [PubMed] [Google Scholar]
- Altman J. Postnatal development of the cerebellar cortex in the rat. III. Maturation of the components of the granular layer. J Comp Neurol. 1972b;145:465–513. doi: 10.1002/cne.901450403. [DOI] [PubMed] [Google Scholar]
- Altman J, Bayer SA. Development of the Cerebellar System. New York: CRC Press; 1996. [Google Scholar]
- Chameau P, Inta D, Vitalis T, Monyer H, Wadman WJ, van Hooft JA. The N-terminal region of reelin regulates postnatal dendritic maturation of cortical pyramidal neurons. Proc Natl Acad Sci U S A. 2009;106:7227–7232. doi: 10.1073/pnas.0810764106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crepel F, Debono M, Flores R. α-Adrenergic inhibition of rat cerebellar Purkinje cells in vitro: a voltage-clamp study. J Physiol. 1987;383:487–498. doi: 10.1113/jphysiol.1987.sp016423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crepel F, Delhaye-Bouchaud N, Dupont JL. Fate of the multiple innervation of cerebellar Purkinje cells by climbing fibers in immature control, X-irradiated and hypothyroid rats. Brain Res. 1981;227:59–71. doi: 10.1016/0165-3806(81)90094-8. [DOI] [PubMed] [Google Scholar]
- Crepel F, Mariani J, Delhaye-Bouchaud N. Evidence for a multiple innervation of Purkinje cells by climbing fibres in the immature rat cerebellum. J Neurobiol. 1976;7:567–578. doi: 10.1002/neu.480070609. [DOI] [PubMed] [Google Scholar]
- D’Arcangelo G, Homayouni R, Keshvara L, Rice DS, Sheldon M, Curran T. Reelin is a ligand for lipoprotein receptors. Neuron. 1999;24:471–479. doi: 10.1016/s0896-6273(00)80860-0. [DOI] [PubMed] [Google Scholar]
- Doughty ML, Lohof A, Selimi F, Delhaye-Bouchaud N, Mariani J. Afferent–target cell interactions in the cerebellum: negative effect of granule cells on Purkinje cell development in lurcher mice. J Neurosci. 1999;19:3448–3456. doi: 10.1523/JNEUROSCI.19-09-03448.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eastwood SL, Burnet PW, Gittins R, Baker K, Harrison PJ. Expression of serotonin 5-HT2A receptors in the human cerebellum and alterations in schizophrenia. Synapse. 2001;42:104–114. doi: 10.1002/syn.1106. [DOI] [PubMed] [Google Scholar]
- Eisenman LM, Schalekamp MP, Voogd J. Development of the cerebellar cortical efferent projection: an in-vitro anterograde tracing study in rat brain slices. Brain Res Dev Brain Res. 1991;60:261–266. doi: 10.1016/0165-3806(91)90055-n. [DOI] [PubMed] [Google Scholar]
- Fatemi SH, Snow AV, Stary JM, Araghi-Niknam M, Reutiman TJ, Lee S, Brooks AI, Pearce DA. Reelin signaling is impaired in autism. Biol Psychiatry. 2005;57:777–787. doi: 10.1016/j.biopsych.2004.12.018. [DOI] [PubMed] [Google Scholar]
- Fry M. Developmental expression of Na+ currents in mouse Purkinje neurons. Eur J Neurosci. 2006;24:2557–2566. doi: 10.1111/j.1460-9568.2006.05139.x. [DOI] [PubMed] [Google Scholar]
- Hansel C, de Jeu M, Belmeguenai A, Houtman SH, Buitendijk GHS, Andreev D, De Zeew C, Elgersma Y. αCaMKII is essential for cerebellar LTD and motor learning. Neuron. 2006;51:835–843. doi: 10.1016/j.neuron.2006.08.013. [DOI] [PubMed] [Google Scholar]
- Kano M, Hashimoto K, Chen C, Abeliovich A, Aiba A, Kurihara H, Watanabe M, Inoue Y, Tonegawa S. Impaired synapse elimination during cerebellar development in PKCγ mutant mice. Cell. 1995;83:1223–1231. doi: 10.1016/0092-8674(95)90147-7. [DOI] [PubMed] [Google Scholar]
- Kano M, Hashimoto K, Tabata T. Type-1 metabotropic glutamate receptor in cerebellar Purkinje cells: a key molecule responsible for long-term depression, endocannabinoid signalling and synapse elimination. Philos Trans R Soc Lond B Biol Sci. 2008;363:2173–2186. doi: 10.1098/rstb.2008.2270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kano M, Hashimoto K, Watanabe M, Kurihara H, Offermans S, Jiang H, Wy Y, Jun K, Shin H-S, Inoue Y, Simon MI, Wu D. Phospholipase Cβ4 is specifically involved in climbing fibre synapse elimination in the developing cerebellum. Proc Natl Acad Sci U S A. 1998;95:15724–15729. doi: 10.1073/pnas.95.26.15724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kapfhammer JP. Cellular and molecular control of dendritic growth and development of cerebellar Purkinje cells. Prog Histochem Cytochem. 2004;39:131–182. doi: 10.1016/j.proghi.2004.07.002. [DOI] [PubMed] [Google Scholar]
- Kimpo RR, Raymond JL. Impaired motor learning in the vestibulo-ocular reflex in mice with multiple climbing fibre input to cerebellar Purkinje cells. J Neurosci. 2007;27:5672–5682. doi: 10.1523/JNEUROSCI.0801-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKay BE, Turner RW. Physiological and morphological development of the rat cerebellar Purkinje cell. J Physiol. 2005;567:829–850. doi: 10.1113/jphysiol.2005.089383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Offermans S, Hashimoto K, Watanabe M, Sun W, Kurihara H, Thompson RF, Inoue Y, Kano M, Simon MI. Impaired motor coordination and persistent multiple climbing fibre innervation of cerebellar Purkinje cells in mice lacking Gαq. Proc Natl Acad Sci U S A. 1997;94:14089–14094. doi: 10.1073/pnas.94.25.14089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oostland M, Sellmeijer J, van Hooft JA. Transient expression of functional 5-HT3 receptors by glutamatergic granule cells in the early postnatal mouse cerebellum. J Physiol. 2011;589:4837–4846. doi: 10.1113/jphysiol.2011.217307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scelfo B, Strata P. Correlation between multiple climbing fibre regression and parallel fibre response development in the postnatal mouse cerebellum. Eur J Neurosci. 2005;21:971–978. doi: 10.1111/j.1460-9568.2005.03933.x. [DOI] [PubMed] [Google Scholar]
- Scorcioni R, Polavaram S, Ascoli GA. L-Measure: a web-accessible tool for the analysis, comparison and search of digital reconstructions of neuronal morphologies. Nat Protoc. 2008;3:866–876. doi: 10.1038/nprot.2008.51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sinagra M, Gonzalez Campo C, Verrier D, Moustié O, Manzoni OJ, Chavis P. Glutamatergic cerebellar granule neurons synthesize and secrete reelin in vitro. Neuron Glia Biol. 2008;4:189–196. doi: 10.1017/S1740925X09990214. [DOI] [PubMed] [Google Scholar]
- Slater P, Doyle CA, Deakin JF. Abnormal persistence of cerebellar serotonin-1A receptors in schizophrenia suggests failure to regress in neonates. J Neural Transm. 1998;105:305–315. doi: 10.1007/s007020050060. [DOI] [PubMed] [Google Scholar]
- van der Velden L, van Hooft JA, Chameau P. Altered dendritic complexity affects firing properties of cortical layer 2/3 pyramidal neurons in mice lacking the 5-HT3A receptor. J Neurophysiol. 2012;108:1521–1528. doi: 10.1152/jn.00829.2011. [DOI] [PubMed] [Google Scholar]
- van Hooft JA. Fast Green FCF (Food Green 3) inhibits synaptic activity in rat hippocampal interneurons. Neurosci Lett. 2002;318:163–165. doi: 10.1016/s0304-3940(01)02452-1. [DOI] [PubMed] [Google Scholar]
- van Welie I, Smith IT, Watt AJ. The metamorphosis of the developing cerebellar microcircuit. Curr Opin Neurobiol. 2011;21:245–253. doi: 10.1016/j.conb.2011.01.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watanabe M, Kano M. Climbing fiber synapse elimination in cerebellar Purkinje cells. Eur J Neurosci. 2011;34:1697–1710. doi: 10.1111/j.1460-9568.2011.07894.x. [DOI] [PubMed] [Google Scholar]
- Zeitz KP, Guy N, Malmberg AB, Dirajlal S, Martin WJ, Sun L, Bonhaus DW, Stucky CL, Julius D, Basbaum AI. The 5-HT3 subtype of serotonin receptor contributes to nociceptive processing via a novel subset of myelinated and unmyelinated nociceptors. J Neurosci. 2002;22:1010–1019. doi: 10.1523/JNEUROSCI.22-03-01010.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]