

SUMMARY
Infection with Helicobacter pylori is a major risk factor for the development of gastric cancer, and infection with strains carrying the virulence factor CagA significantly increases this risk. To investigate the mechanisms by which CagA promotes carcinogenesis, we generated transgenic zebrafish expressing CagA ubiquitously or in the anterior intestine. Transgenic zebrafish expressing either the wild-type or a phosphorylation-resistant form of CagA exhibited significantly increased rates of intestinal epithelial cell proliferation and showed significant upregulation of the Wnt target genes cyclinD1, axin2 and the zebrafish c-myc ortholog myca. Coexpression of CagA with a loss-of-function allele encoding the β-catenin destruction complex protein Axin1 resulted in a further increase in intestinal proliferation. Coexpression of CagA with a null allele of the key β-catenin transcriptional cofactor Tcf4 restored intestinal proliferation to wild-type levels. These results provide in vivo evidence of Wnt pathway activation by CagA downstream of or in parallel to the β-catenin destruction complex and upstream of Tcf4. Long-term transgenic expression of wild-type CagA, but not the phosphorylation-resistant form, resulted in significant hyperplasia of the adult intestinal epithelium. We further utilized this model to demonstrate that oncogenic cooperation between CagA and a loss-of-function allele of p53 is sufficient to induce high rates of intestinal small cell carcinoma and adenocarcinoma, establishing the utility of our transgenic zebrafish model in the study of CagA-associated gastrointestinal cancers.
INTRODUCTION
Helicobacter pylori is a pathogenic Gram-negative bacterium that colonizes over 50% of the world’s human population. Colonization with H. pylori is linked to numerous gastric disorders including gastritis, peptic ulcer disease and gastric adenocarcinoma (Blaser and Atherton, 2004). Although gastric cancer occurs in fewer than 1% of people colonized by H. pylori (Amieva and El-Omar, 2008), it is still the second-most common cause of cancer mortality worldwide (Peek and Blaser, 2002), and more than 50% of gastric adenocarcinomas can be attributed to infection with H. pylori (Asghar and Parsonnet, 2001). Most people infected with H. pylori, however, do not develop gastric cancer and the molecular mechanisms underlying this disparity have yet to be fully elucidated.
Although there are many factors that appear to contribute to the carcinogenicity of H. pylori, strains that translocate the CagA protein into host cells are significantly more likely to cause gastric cancer than strains lacking this ability. CagA is one of 28 gene products encoded by the cag pathogenicity island (cag PAI), a 40-kb stretch of DNA shown to be present in most strains isolated from patients with severe gastric pathology (Censini et al., 1996). During infection with H. pylori, CagA is translocated into host cells via a type IV secretion system (TFSS), where it interacts with a multitude of host cell proteins. These interactions have been shown to affect signal transduction pathways, the cytoskeleton and cell junctions (Bourzac and Guillemin, 2005).
After translocation into host cells by the H. pylori TFSS, CagA can be phosphorylated by Src family kinases on tyrosine residues within conserved Glu-Pro-Ile-Tyr-Ala (EPIYA) motifs (Selbach et al., 2002; Stein et al., 2002). After phosphorylation, CagA has been shown to induce morphological changes in cultured epithelial cells through interaction with a variety of host-cell proteins such as SHP-2, Met, Csk, Grb2 and ZO-1 (Amieva et al., 2003; Churin et al., 2003; Higashi et al., 2002; Tsutsumi et al., 2003; Mimuro et al., 2002). In addition to its phosphorylation-dependent effects, CagA has also been shown to interact in a phosphorylation-independent manner with pathways associated with proliferation and inflammation (Suzuki et al., 2009). Although it is not yet clear which of these myriad interactions are required for the development of gastric cancer in persons colonized by H. pylori, the ability of CagA to interact with components of the canonical Wnt signaling pathway provides a potential link between the observed oncogenic effects of CagA and a host signaling pathway frequently deregulated in gastrointestinal cancers (Franco et al., 2005).
In addition to its role in early embryogenesis, the canonical Wnt signaling pathway plays a crucial role in regulating the proliferation and homeostasis of gastrointestinal epithelia. In normal stomach and intestinal epithelia, Wnt signaling has been shown to be important for proliferation, stem cell maintenance, and tissue renewal (Barker et al., 2010; Sato et al., 2011; Pinto et al., 2003; Ootani et al., 2009; Sato et al., 2004). On the other hand, activation of Wnt signaling has been shown to result in cancers of the stomach and colon (Oshima et al., 2006; Powell et al., 1992; Fearon and Vogelstein, 1990). Wnt pathway activity is tightly controlled via regulation of the primary Wnt effector protein, β-catenin. β-catenin complexes with E-cadherin to form adherens junctions between epithelial cells, and in the absence of Wnt ligand, is also bound by Axin/APC/Gsk3β in the so-called ‘β-catenin destruction complex’, where it is targeted for proteosomal degradation. Upon binding of Wnt by the co-receptors Frizzled and LRP, Axin1 is sequestered at the membrane, preventing assembly of the β-catenin destruction complex. This results in cytoplasmic accumulation of β-catenin and subsequent translocation of β-catenin into the nucleus. Upon nuclear translocation, β-catenin binds the essential transcriptional cofactor TCF and initiates transcription of Wnt target genes, including axin, myc and cyclin genes.
TRANSLATIONAL IMPACT.
Clinical issue
Infection with the bacterium Helicobacter pylori represents a major global health burden that has been implicated in a wide range of gastric disorders, from acute gastric inflammation to stomach cancer. H. pylori strains that are capable of translocating the bacterial effector protein CagA into host epithelial cells are associated with the highest risk of gastric cancer development. CagA-induced pathogenesis is multifactorial, and in vitro studies have reported different effects in diverse cell lines. Furthermore, CagA-induced oncogenesis is strongly associated with variations in host genotype, highlighting the need for a genetically tractable in vivo model that faithfully recapitulates the mechanisms underlying carcinogenesis in humans.
Results
In this manuscript, the authors report the development of a novel transgenic zebrafish system for the investigation of the H. pylori virulence factor CagA. This system recapitulates the major hallmarks of CagA pathogenesis observed in cell culture and murine models, while providing distinct advantages over previous models. Using the zebrafish model, the authors demonstrate that the early effects of CagA on intestinal epithelial proliferation require the function of the canonical Wnt signaling component Tcf4, a β-catenin cofactor, but not Axin1, a component of the β-catenin destruction complex. They further report that co-expression of CagA with a loss-of-function allele of the tumor suppressor p53 results in high rates of neoplastic transformation, thereby providing the first direct in vivo evidence for oncogenic cooperation between CagA and p53.
Implications and future directions
The CagA transgenic zebrafish model described herein presents several key advantages over current in vivo models. First, the rapid development of the zebrafish digestive tract makes it an ideal system for the study of CagA-associated gastrointestinal disease. Second, the ease of transgenesis via Tol2 transposition enables the rapid introduction of additional alleles, facilitating structure-function studies that are difficult to perform in other vertebrate models. Finally, the microbiota of CagA transgenic zebrafish is readily manipulated or ablated, paving the way for future CagA gnotobiotic studies.
Non-phosphorylated CagA has been previously shown to disrupt the β-catenin–E-cadherin complex in cultured epithelial cells, causing cytoplasmic and nuclear accumulation of β-catenin, and subsequent activation of the Wnt pathway (El-Etr et al., 2004; Suzuki et al., 2005; Murata-Kamiya et al., 2007). Additionally, CagA has been shown to increase signaling through β-catenin via activation of phosphatidylinositol 3-kinase and/or Akt (Suzuki et al., 2009). Although the mechanisms of interactions of CagA with the Wnt pathway have yet to be fully elucidated, it is clear both that CagA is capable of activating Wnt signaling through β-catenin and that inappropriate activation of Wnt signaling is potentially oncogenic. Understanding the wide variety of host cell interactions required for H. pylori-induced pathogenesis has necessitated the use of animal models, and to date numerous primate and rodent models have been developed (Wirth et al., 1998; Lee et al., 1997; Ohnishi et al., 2008; Solnick et al., 1999). Although previously unexploited in the study of H. pylori pathogenesis, the teleost fish Danio rerio (zebrafish) has emerged as a model organism for the study of various human diseases, including leukemia (Feng et al., 2010), melanoma (Ceol et al., 2011; White et al., 2011) and intestinal neoplasia (Haramis et al., 2006). In lieu of a stomach, zebrafish possess an anterior digestive compartment known as the intestinal bulb. The zebrafish intestinal bulb epithelium is columnar and non-ciliated like that of the mammalian stomach and expresses sox2 and barx1 (Muncan et al., 2007), two mammalian stomach markers (Tsukamoto et al., 2005; Tissier-Seta et al., 1995; Kim et al., 2005). Unlike the mammalian stomach, however, it lacks the chief and parietal cell types. Nonetheless, the zebrafish intestinal bulb has been proposed to share a common ontogeny with the mammalian stomach and its renewal is regulated by similar molecular pathways, including the Notch and Wnt pathways (Crosnier et al., 2005; Cheesman et al., 2011). Finally, the rapid development of the zebrafish intestinal tract makes it an ideal model for the study of gastrointestinal development and disease (Faro et al., 2009).
Here, we describe the development of a novel transgenic model system that simplifies the complexity of H. pylori infection to study the effects of a single bacterial protein, CagA, on host cell biology in the zebrafish intestine. We report that proliferation in the zebrafish larval intestinal epithelium is increased by transgenic expression of CagA and that this increase occurs independently of CagA phosphorylation. We demonstrate that expression of CagA induces cytoplasmic and nuclear accumulation of the Wnt effector β-catenin, as well as activation of known Wnt target genes. The genetic tractability of the zebrafish system allowed us to explore genetic interactions between CagA and a number of host signaling pathways. We show that CagA causes proliferation of the zebrafish intestinal epithelium via activation of the canonical Wnt signaling pathway downstream of or in parallel to the β-catenin destruction complex and upstream of the β-catenin transcriptional cofactor Tcf4. Additionally, we demonstrate that long-term expression of wild-type CagA, but not the phosphorylation-resistant form, is sufficient to induce pathologic intestinal hyperplasia in adults and that oncogenic cooperation between the cagA transgene and a loss-of-function allele of p53 results in high rates of intestinal adenocarcinoma and small cell carcinoma.
RESULTS
Generation of CagA-expressing transgenic zebrafish
In order to generate cagA transgenic animals, we cloned the cagA gene from H. pylori strain G27. Strain G27 was originally isolated from Grossetto Hospital (Tuscany, Italy), and has been used extensively in research on the CagA virulence factor (Amieva et al., 2003; El-Etr et al., 2004; Guillemin et al., 2002; Segal et al., 1999). The cloned gene was then 3′-tagged with EGFP to facilitate in vivo visualization of CagA expression. To express CagA ubiquitously in zebrafish, the cagA/EGFP fusion construct was connected downstream of the 5.3 kb beta-actin (b−) (Higashijima et al., 1997) promoter (supplementary material Fig. S1A). To facilitate intestine-specific expression of the fusion construct, we connected cagA/EGFP downstream of a 1.6-kb fragment of the zebrafish intestinal fatty acid binding protein (i−) (Her et al., 2004) promoter (supplementary material Fig. S1B). By 6 days post-fertilization (dpf) b-cagA/EGFP transgenic zebrafish exhibited ubiquitous fluorescence, whereas i-cagA/EGFP transgenic larvae exhibited fluorescence in the distal esophagus and anterior intestine (Fig. 1A,B). The phosphorylation state of CagA has been previously shown to have significant effects on the type and severity of CagA-induced pathologies, so in order to determine the role of CagA phosphorylation in the intestinal epithelium, we fused the previously described phosphorylation-resistant cagAEPISA allele (Stein et al., 2002) (supplementary material Fig. S1C) to EGFP and connected it downstream of the b-actin promoter (supplementary material Fig. S1D). b-cagAEPISA/EGFP transgenics exhibited ubiquitous fluorescence and were indistinct from b-cagA/EGFP fish (Fig. 1C). Expression of cagA mRNAs was verified in transgenic animals by RT-PCR (Fig. 1D), and analysis of relative intestinal cagA transcript level in the transgenic lines via quantitative real-time PCR revealed significantly elevated expression of the cagA transgene when driven by the b-actin promoter rather than the i-fabp promoter (Fig. 1E).
Fig. 1.

Development of CagA+ transgenic zebrafish. (A) Ubiquitous CagA-EGFP fusion protein expression driven by the b-actin promoter. (B) Ubiquitous CagAEPISA-EGFP fusion protein expression driven by the b-actin promoter. (C) Intestinal CagA-EGFP fusion protein expression driven by the i-fabp promoter. (D) RT-PCR of dissected larval intestine showing expression of cagA and the housekeeping control gene RPL13 at 6 dpf. (E) Quantitative RT-PCR of dissected adult intestines showing relative expression levels of cagA transcript in transgenic lines at 1 year of age. Expression levels were normalized to SDHA and β-actin; bars indicate mean ± s.d. of biological triplicates. Scale bars: 500 μm.
CagA expression causes overproliferation of the intestinal epithelium
To determine the effects of CagA expression on the larval zebrafish intestine, we examined wild-type and CagA transgenic animals at 6 dpf, by which time autonomous feeding has begun, and at 15 dpf, by which time intestinal folding is complete (Ng et al., 2005). CagA-expressing zebrafish larvae showed normal intestinal development (Fig. 2A,B) and were histologically indiscernible from wild-type clutch-mates (Fig. 2C,D). In addition, the CagA-expressing larvae exhibited no gross abnormalities in cell junctions, as assessed by staining with a pan-cadherin antibody (supplementary material Fig. S2). We next sought to establish the effects of CagA on larval intestinal proliferation, because CagA had been previously shown to increase epithelial cell proliferation in vitro and in vivo (Mimuro et al., 2002; Nagy et al., 2011). To determine the proliferation state of CagA-expressing intestines, we analyzed animals at 6 and 15 dpf that had been exposed to the nucleotide analog 5-ethynyl-2′-deoxyuridine (EdU) for ∼10 hours and counted S-phase nuclei in 30 serial sections of the intestinal bulb. Expression of CagA resulted in a significant increase in EdU-labeled cells in all transgenic lines at 6 and 15 dpf (Fig. 2E,F). To determine whether this increase in proliferation had an effect on the cell census, we quantified total epithelial cell number in single hematoxylin and eosin (H&E)-stained sagittal sections along the length of the intestine. We did not observe any significant difference in total cell counts between CagA transgenics and wild-type animals at 6 and 15 dpf (Fig. 2G,H), indicating that expression of CagA caused increased turnover of intestinal epithelial cells. Increased intestinal cell turnover would require an increase in cell death; however, consistent with previous reports and due to the transient nature of extruded apoptotic cells (Crosnier et al., 2005), we observed very few TUNEL-positive cells in the intestines of wild-type and CagA-expressing animals (Fig. 2I,J), with no significant difference observed between the two groups. Finally, the intestinal epithelia of b-cagA animals did not display an increased number of local neutrophils at 8 dpf, indicating a lack of CagA-induced intestinal inflammation at this stage (supplementary material Fig. S3).
Fig. 2.

CagA expression causes overproliferation of the intestinal epithelium. (A,B) H&E stained sagittal sections of wild-type (A) and b-cagA transgenic (B) zebrafish intestine at 6 dpf. (C,D) H&E stained sagittal sections of wild-type (C) and b-cagA transgenic (D) zebrafish intestine at 15 dpf. (E,F) Intestinal epithelial cell proliferation at 6 dpf (E) and 15 dpf (F). Bars represent proliferation (mean ± s.e.m.) as a percentage of wild-type; n=10, *P<0.05 using one-way ANOVA with Tukey’s test. (G,H) Total intestinal epithelial cell counts of single H&E stained midline sagittal sections at 6 dpf (G) and 15 dpf (H). (I,J) TUNEL-positive cells in the intestinal epithelium at 6 dpf (I) and 15 dpf (J). Scale bars: 10 μm.
CagA expression activates the Wnt pathway downstream of the β-catenin destruction complex
We had previously shown that epithelial cell proliferation in the zebrafish intestine is regulated by the Wnt pathway (Cheesman et al., 2011). In addition, previous studies had shown that CagA can induce cytoplasmic and nuclear accumulation of the Wnt effector protein β-catenin and can activate transcription of canonical Wnt target genes (Franco et al., 2005; Suzuki et al., 2009; Nagy et al., 2011). Accordingly, we examined whether CagA expression was capable of activating the Wnt signaling pathway in the zebrafish intestine at different developmental stages. We first utilized quantitative real-time PCR to assess the relative expression levels of known Wnt target genes in dissected adult intestines. Transcript levels of the Wnt target genes c-myc (myca) (He et al., 1998), axin2 (Yan et al., 2001) and cyclinD1 (Tetsu and McCormick, 1999) were modestly increased in all CagA-expressing lines relative to the wild-type strain (Fig. 3A–C). We next asked whether CagA was capable of inducing β-catenin accumulation in epithelial cells of the larval intestine, indicating activation of the Wnt pathway. CagA expression caused a significant increase in the number of intestinal epithelial cells with cytoplasmic and nuclear accumulation of β-catenin as compared with wild-type animals (Fig. 3D,E). The fact that EdU labeling was not usually coincident with cytoplasmic and nuclear accumulation of β-catenin is probably due to the fact that, whereas relocalization of β-catenin is a transient event, the EdU-labeled cells had undergone S-phase any time during the 12-hour labeling period.
Fig. 3.

CagA activates canonical Wnt signaling in the intestinal epithelium. (A–C) Quantitative RT-PCR data showing relative expression levels of the Wnt target genes mycA (A), cyclinD1 (B) and axin2 (C). Expression levels were assayed in dissected adult intestines and normalized to SDHA and β-actin; bars indicate mean ± s.d. of biological triplicates. (D–G) Immunofluorescence micrographs showing proliferating cells (EdU, green, 10-hour label) and cells with nuclear and cytoplasmic accumulation of β-catenin (red staining and white arrowheads) in intestinal cross-sections of wild-type (D), b-cagA (E), axin1tm213 (F) and b-cagA; axin1tm213 (G) animals at 6 dpf. (H) Quantification of proliferating (EdU+) cells. (I) Quantification of cells with nuclear and cytoplasmic accumulation of β-catenin.
In order to assess the significance of CagA-induced β-catenin accumulation, we next compared the intestinal β-catenin accumulation observed in CagA-expressing animals to that of a known Wnt signaling mutant, axin1tm213. The axin1tm213 homozygotes exhibit deregulated Wnt signaling as a result of a missense mutation in the Gsk3β binding domain of Axin1, which prevents assembly of the β-catenin destruction complex. These mutants die as a result of craniofacial defects, but are viable until 8 dpf, allowing study of the juvenile intestine (Heisenberg et al., 2001; van de Water et al., 2001). As expected, we observed increases over wild-type and CagA-expressing animals in both the number of proliferating cells and the number of cells featuring cytoplasmic and/or nuclear accumulation of β-catenin in the intestinal epithelia of axin1tm213/tm213 mutants, consistent with constitutively activated Wnt signaling (Fig. 3E).
We reasoned that if CagA were capable of activating Wnt signaling upstream of the β-catenin destruction complex, then axin1tm213 homozygotes should be refractory to CagA-induced accumulation of β-catenin and levels of β-catenin accumulation in b-cagA; axin1tm213/tm213 double mutants should resemble those of axin1tm213 homozygotes. Instead, when we generated b-cagA, axin1tm213/tm213 fish, we found that expression of CagA in axin1 homozygous mutants resulted in a dramatic increase in cell proliferation and β-catenin accumulation (Fig. 3F). Taken together, these data indicate that CagA is capable of causing sustained activation of canonical Wnt signaling in the intestinal epithelium and that it does so either downstream of, or in parallel to, the β-catenin destruction complex. Furthermore, CagA-induced accumulation of β-catenin was strongly correlated with increased epithelial proliferation (Fig. 3G,H), suggesting that CagA stimulates proliferation through activation of the Wnt pathway.
CagA-dependent overproliferation of the intestinal epithelium requires tcf4
To determine whether CagA-induced overproliferation of the intestinal epithelium is dependent on canonical Wnt signaling downstream of the β-catenin destruction complex, we utilized a null allele of the essential β-catenin transcriptional cofactor, Tcf4 (Muncan et al., 2007). We reasoned that if the pro-proliferative effects of CagA were acting upstream of Tcf4, rates of intestinal proliferation in i-cagA; tcf4exl double mutants should be identical to those observed in tcfnull animals. As previously observed, i-cagA animals showed a significant increase in proliferation over the wild type, whereas tcf4exl/exl mutants showed levels of intestinal proliferation similar to wild-type animals (Fig. 4). Rates of intestinal proliferation in i-cagA; tcf4exl/exl larvae were statistically indistinguishable from wild-type and tcf4exl/exl mutants, indicating that CagA requires Tcf4 function to increase intestinal epithelial proliferation. This result places activation of the Wnt signaling pathway by CagA downstream of or in parallel to Axin1 and upstream of Tcf4 (supplementary material Fig. S4).
Fig. 4.
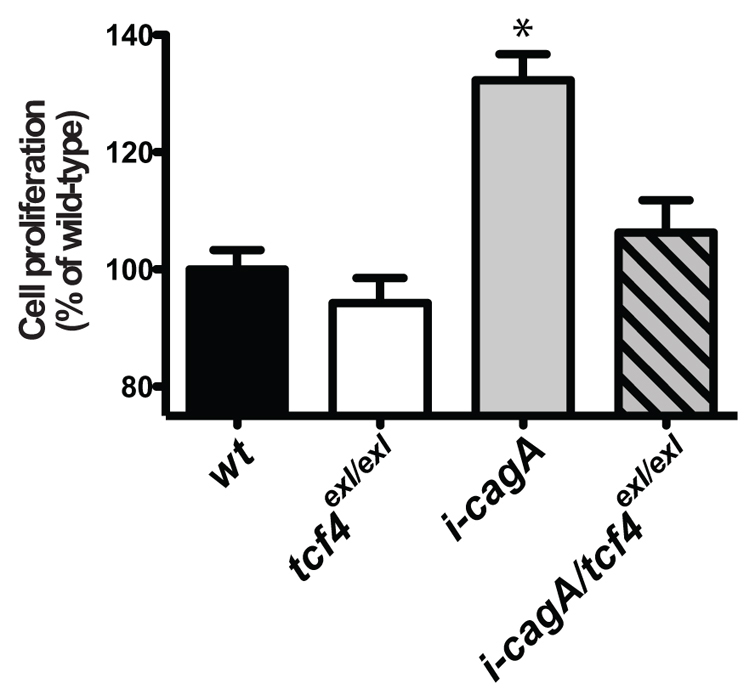
CagA-dependent overproliferation of the intestinal epithelium requires tcf4. Intestinal epithelial cell proliferation at 15 dpf. Bars represent proliferation (mean ± s.e.m.) as a percentage of wild-type; n=10, *P<0.05 using one-way ANOVA with Tukey’s test.
CagA expression causes phosphorylation-dependent intestinal hyperplasia in adult zebrafish
H. pylori-associated gastric adenocarcinoma occurs as a result of lifelong exposure to the bacterium, with CagA+ strains posing a significantly greater cancer risk (Asghar and Parsonnet, 2001). In order to study the long-term effects of CagA exposure in our model, we performed histological analysis of adult b-cagA, i-caga and b-cagaEPISA animals at 1 year of age. Wild-type adults (18 months post-fertilization) served as controls. Upon examination, no hyperplastic or neoplastic lesions were found in any of the wild-type controls (Fig. 5A,G; supplementary material Table S1). A proportion of the b-cagA and i-cagA individuals exhibited significant intestinal epithelial hyperplasia at 1 year of age (Fig. 5B,C). Surprisingly, despite the significant increases in proliferation and Wnt activation observed in younger b-cagaEPISA animals, no hyperplasia was observed in age-matched adults of this genotype (Fig. 5D). These data suggest that although the phosphorylation-independent activation of Wnt signaling by CagA is sufficient to induce sustained overproliferation of the larval intestinal epithelium, it is not sufficient to induce significant hyperplastic changes in the adult intestinal epithelium, as seen in the groups expressing the non-mutant CagA, either ubiquitously or in an intestine-specific manner.
Fig. 5.
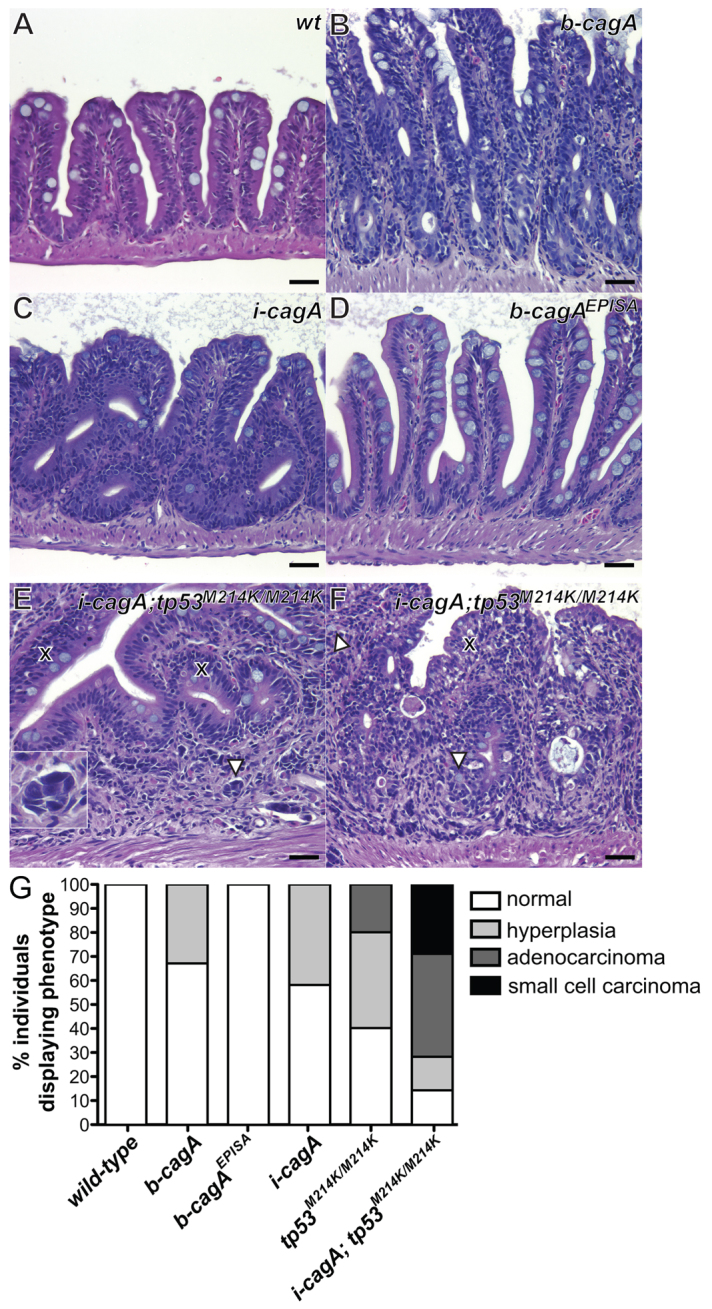
CagA expression causes phosphorylation-dependent intestinal epithelial hyperplasia and induces adenocarcinoma formation in combination with p53 loss. (A–F) H&E stained sagittal sections of adult zebrafish intestine (A) Wild-type intestine at 18 months post-fertilization (mpf) showing normal intestinal architecture, with a single layer of epithelial cells lining the mucosal folds. (B,D) b-cagA (B) and i-cagA (D) intestines at 12 mpf, displaying mucosal fold epithelial hyperplasia, dysplasia within mucosal sulci and mucosal fold fusion. (C) b-cagAEPISA intestine at 12 mpf showing normal intestinal architecture, identical to wild-type. (E) i-cagA; tp53M214K/M214K small cell carcinoma with small nests of neoplastic cells in the lamina propria (arrowheads). Inset depicts higher magnification of tumor cells; the cross marks the epithelium. (F) i-cagA; tp53M214K/M214K adenocarcinoma, poorly differentiated, invade into the lamina propria with complete disorganization of the epithelium, as shown by goblet cells randomly scattered throughout (arrowheads). (G) Summary of intestinal histological abnormalities observed in adult CagA-expressing animals as a result of a blinded histological analysis of H&E stained sections (wild-type, n=22; b-cagA, n=24; b-cagAEPISA, n=18; i-cagA, n=19; tp53M214K/M214K, n=5; i-cagA/tp53M214K/M214K, n=7). Scale bars: 25 μm.
Coexpression of the cagA transgene with a p53 loss-of-function allele results in high rates of intestinal adenocarcinoma
The tumor suppressor gene p53 is frequently mutated in diffuse-and intestinal-type gastric cancers (Nobili et al., 2011; Ranzani et al., 1995), and gastric adenocarcinomas isolated from CagA+ H. pylori-infected patients exhibit frequent mutation in p53 (Shibata et al., 2002). Additionally, CagA has been shown to subvert the tumor suppressor function of the apoptosis-stimulating protein ASPP2 in cultured cells, leading to enhanced degradation of p53 (Buti et al., 2011). In order to examine the potential for oncogenic cooperation between the cagA transgene and p53 we bred b-cagA and i-cagA animals to animals homozygous for a loss-of-function allele of p53 (tp53M214K) to obtain b-cagA; tp53M214K/M214K or i-cagA; tp53M214K/M214K animals. The zebrafish ortholog of p53 is highly conserved in both structure and function and the tp53M214K DNA-binding domain mutation is orthologous to methionine 246 missense mutations previously identified in human tumors (Storer and Zon, 2010). At 1 year post-fertilization, all of the tp53M214K/M214K fish failed to thrive and exhibited high rates of ocular malignant peripheral nerve sheath tumors, recapitulating previous studies using this p53 allele (Berghmans et al., 2005). An insufficient number of b-cagA; tp53M214K/M214K individuals survived to this time point for analysis, but we were able to examine small numbers of both tp53M214K/M214K and i-cagA; tp53M214K/M214K (Fig. 5E,F) lines. In both lines, we observed examples of intestinal epithelial hyperplasia and definitive neoplasia (Fig. 5G).
In the affected genotypes displaying hyperplastic changes, the intestinal mucosa was thrown into irregular and haphazard folds lined by a ragged and thickened epithelium often 2–6 cells deep with pseudostratification of nuclei, which was most prominent within invaginations between the mucosal villi (mucosal sulci). Infolding of the hyperplastic epithelium frequently resulted in formation of mucosal pseudocrypts, with the most severely affected intestines also displaying frequent epithelial fusion between adjacent mucosal folds. In addition, numerous aponecrotic intestinal epithelial cells were observed and directly reflected rapid epithelial cell proliferation and turnover. Small numbers of a chronic inflammatory cell infiltrate, composed mostly of lymphocytes and few eosinophilic granule cells, were seen percolating through the hyperplastic epithelium in many areas. Foci of dysplastic intestinal epithelial cells were often identified in hyperplastic areas, usually within mucosal sulci. Dysplastic cells demonstrated progressive disorganization including ‘piling-up’ of cells and loss of nuclear polarity, nuclear and cytologic pleomorphism, hyperchromatic elongated nuclei and inconspicuous nucleoli with sparse cytoplasm (increased nuclear to cytoplasm ratio) and occasional bizarre mitotic figures. In all cases where dysplastic cells were observed, there was no invasion through the basement membrane (i.e. carcinoma in situ) except for one fish in the tp53M214K/M214K group, which had a solitary maxillary (upper jaw) focus of carcinoma in situ within the oropharyngeal cavity. When definitive intestinal neoplasia was seen, adenocarcinoma was most often found in the anterior intestine and small cell carcinoma in the anterior or mid-intestine.
Adenocarcinomas displayed variable degrees of differentiation, ranging from well to poorly differentiated, with a tendency to form disorganized and cribrose acinar-like pseudocrypts that penetrated deep into the lamina propria, in the absence of an interceding basement membrane. Individual tumor cells had hyperchromatic, ovoid to elongated nuclei with granular chromatin, multiple small nucleoli and sparse basophilic cytoplasm. In less differentiated adenocarcinomas, bizarre mitotic figures were occasionally seen. Locally extensive fibrogenesis within the lamina propria (intraproprial desmoplasia) and variable numbers of chronic inflammatory cell infiltrates, comprised of intermingled lymphocytes and eosinophilic granule cells, were often associated with the adenocarcinomas. The two small cell carcinomas identified in the i-cagA; tp53M214K/M214K group were composed of densely cellular nests of polygonal to fusiform cells, lacking an organoid pattern, which infiltrated deep into the lamina propria and were not associated with pseudocrypts. Individual neoplastic cells within nests had pleomorphic, deeply basophilic nuclei with dense granular chromatin, inconspicuous nucleoli and minimal cytoplasm. Solitary necrotic tumor cells were seen in some of the nests, accompanied by small aggregates of lymphocytes. Lymphovascular invasion and distant metastasis was not observed in either of the tumor types. Incidence and overall severity of lesions within the expression domain of the cagA transgene were higher in i-cagA; tp53M214K/M214K animals than in the corresponding anatomical region of tp53M214K/M214K animals (Fig. 5G; supplementary material Table S1). These data indicate that expression of CagA with concomitant p53 loss is sufficient to induce high rates of adenocarcinoma and small cell carcinoma in the zebrafish intestine and demonstrate the utility of our model for the study of CagA-associated gastrointestinal cancers.
DISCUSSION
Here, we describe the development of a novel in vivo model of CagA-induced intestinal pathology in zebrafish that recapitulates major hallmarks of CagA pathogenesis observed in cell culture and murine models such as increased epithelial proliferation, cellular accumulation of β-catenin and intestinal hyperplasia (Ohnishi et al., 2008; Mimuro et al., 2002; El-Etr et al., 2004; Suzuki et al., 2005; Murata-Kamiya et al., 2007; Nagy et al., 2011). We utilized transgenic expression of CagA to investigate how the H. pylori virulence factor CagA is able to disrupt normal programs of intestinal epithelial renewal via activation of an important host signaling pathway, the Wnt pathway, to cause significant overproliferation of an intact epithelium in vivo. We show that activation of canonical Wnt signaling upstream of the essential β-catenin cofactor Tcf4 and downstream of the β-catenin destruction complex is required for the early effects of CagA on intestinal epithelial proliferation.
We further utilized our novel transgenic zebrafish system to demonstrate that long-term expression of CagA is sufficient to cause intestinal hyperplasia in adult zebrafish. Notably, although expression of the phosphorylation-resistant b-cagAEPISA allele is capable of inducing significant sustained overproliferation of the larval intestinal epithelium coupled with increased Wnt activation, it failed to induce significant intestinal hyperplasia in adult animals. These data corroborate a previous study using a CagA transgenic mouse model, which demonstrated that the ability of CagA to induce severe epithelial hyperplasia in vivo is correlated with its capacity to be phosphorylated by host kinases (Ohnishi et al., 2008). It is possible that the activation of Wnt signaling by CagA and subsequent induction of proliferation act in concert with further oncogenic stimuli, which might occur in the form of previously observed phosphorylation-dependent events such as epithelial depolarization (Amieva et al., 2003) or ERK activation by CagA (Higashi et al., 2004). These data illustrate the utility of long-term in vivo modeling of CagA pathogenesis because the cumulative effects of CagA expression cannot be predicted from the transient cellular responses it elicits.
Host genetics play a significant role in the development of H. pylori-associated gastric cancer. For example, certain alleles of the host genes p53, IL-1β and IL-10 are strongly correlated with the development of gastric adenocarcinoma in H. pylori-infected humans (Shibata et al., 2002; El-Omar et al., 2003). Transgenic expression of CagA in mice was sufficient to cause gastric and intestinal carcinomas, but these only developed in less than 5% of the animals (Ohnishi et al., 2008). We observed high rates of intestinal neoplasia in our CagA transgenic zebrafish model when expressed with a mutant allele of the tumor suppressor p53. These data provide the first direct in vivo evidence for oncogenic cooperation between CagA and p53 and provide a robust model of CagA-induced carcinoma. Our results are consistent with previous findings of increased p53 mutational frequency in H. pylori-associated gastric cancer cases (Shibata et al., 2002) and corroborate a previous study establishing CagA as a bona-fide oncoprotein (Ohnishi et al., 2008). More importantly, these data support the use of our model in the screening of putative gastric cancer susceptibility loci for oncogenic cooperation with CagA.
MATERIALS AND METHODS
Ethics
All zebrafish experiments were carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. The University of Oregon Animal Care Service is fully accredited by the Association for Assessment and Accreditation of Laboratory Animal Care and complies with all United States Department of Agriculture, Public Health Service, Oregon State and local area animal welfare regulations. All activities were approved by the University of Oregon Institutional Animal Care and Use Committee (Animal Welfare Assurance number A-3009-01).
Animals
Transgenic zebrafish were developed using the Tol2kit as previously described (Kwan et al., 2007). tp53M214K (Berghmans et al., 2005), and axin1tm213 (Heisenberg et al., 2001) animals were obtained from Monte Westerfield (University of Oregon) and tcf4exI (Muncan et al., 2007) from Tatjana Piotrowski (University of Utah). All zebrafish experiments were performed using protocols approved by the University of Oregon Institutional Care and Use Committee, and following standard protocols (Westerfield, 2007). CagA transgenics can be obtained by contacting the corresponding author.
EdU labeling and detection
Zebrafish larvae were immersed in 100 μg/ml EdU (A10044; Invitrogen) with 0.5% DMSO for 8–12 hours, fixed overnight at 4°C (4% paraformaldehyde in PBS) with gentle shaking, processed for paraffin embedding and cut into 7-μM sections. Slides were then processed using the Click-iT EdU Imaging Kit (C10337, Invitrogen). EdU-labeled nuclei within the intestinal epithelium were counted over 30 serial sections, beginning at the intestinal-esophageal junction and proceeding caudally into the intestinal bulb.
TUNEL staining
Staining was carried out using the Click-iT TUNEL Imaging Assay (C10245, Invitrogen). TUNEL-positive cells within the intestinal epithelium were counted over 30 serial sections, beginning at the intestinal-esophageal junction and proceeding caudally into the intestinal bulb.
Immunohistochemistry
Immunohistochemistry was carried out on paraffin sections as previously described using anti-β-catenin (1:1000; C2206 rabbit polyclonal, Sigma) (Cheesman et al., 2011).
Histopathology
Histopathological analysis of H&E stained sections was performed by pathologists with expertise in laboratory fish (T.S.P. and M.L.K.) in a blinded manner. For each adult zebrafish genotype, four consecutive sagittal serial sections of the entire intestinal tract, anterior to posterior, were evaluated for epithelial hyperplasia, dysplasia and the presence of neoplasia. Classification of intestinal epithelial hyperplasia included two or more of the following criteria: epithelial cell nuclear pseudostratification, multi-layering of mucosal fold epithelial cells and formation of pseudocrypts, which indicated extensive infolding of hyperplastic epithelium lining the intestinal mucosal folds. Dysplastic changes of the intestinal epithelial cells, observed in several fish within the hyperplastic intestinal epithelium, were classified as an increased nuclear to cytoplasm ratio, nuclear hyperchromatism with indiscernible nucleoli, ‘piling-up’ of epithelial cells, loss of nuclear polarity (i.e. loss of basally oriented epithelial cell nuclei) and abnormal mitotic figures. Classification of intestinal adenocarcinoma included the following criteria: invasive cribriform pseudocrypts that interfaced directly with the lamina propria in the absence of an interceding basement membrane, disorganized histoarchitectural patterns of the pseudocrypts, loss of differentiation from well-defined pseudocrypts to complete absence of acinar-like structures and a desmoplastic response to the neoplastic cells. Small cell carcinoma was classified as densely cellular and discrete small sheets and nests of tumor cells within the lamina propria, with minimal cytoplasm, that lacked an organoid growth pattern. Intratumoral inflammatory infiltrates were also accounted for and classified by chronicity and cell type. Other proliferative lesions, which occurred in only one fish, are described in the Results section.
Quantitative RT-PCR
Reference gene testing was performed using the geNorm reference gene selection kit (Primerdesign) and qBasePLUS software (Biogazelle). Baseline, threshold and efficiency calculations were performed using LinRegPCR software (Ruijter et al., 2009). Quantitative RT-PCR reactions were performed using the SYBR FAST qPCR kit (Kapa Biosystems) on a StepOnePlus Real-Time PCR System (Applied Biosystems) using primers listed in supplementary material Table S2. Expression data were normalized to the geometric mean of the reference genes using StepOne (ABI) software.
Myeloperoxidase staining
Myeloperoxidase (Mpo) staining was carried out using the Leukocyte Peroxidase (Myeloperoxidase) Staining Kit (Sigma-Aldrich). Mpo-positive cells within the intestinal epithelium were counted over 30 serial sections, beginning at the intestinal-esophageal junction and proceeding caudally into the intestinal bulb.
Statistical analysis
All statistical analyses were performed with Graph-Pad Prism software.
Supplementary Material
Acknowledgments
We thank Erika Mittge for technical assistance, Rose Gaudreau and the staff of the University of Oregon Zebrafish Facility for excellent fish husbandry, Poh Kheng Loi and the staffs of the University of Oregon and Oregon State University histology facilities for histology services.
Footnotes
COMPETING INTERESTS
The authors declare that they do not have any competing or financial interests.
AUTHOR CONTRIBUTIONS
J.T.N. and K.G. designed experiments. J.T.N. performed experiments. J.T.N., T.S.P., M.L.K. and K.G. analyzed data. J.T.N., T.S.P., M.L.K. and K.G. wrote the paper.
FUNDING
This research was supported by the National Institutes of Health (NIH) [grant number 1R01DK075667 to K.G.] and a Burroughs Wellcome Fund Investigator in the Pathogenesis of Infectious Disease Award (to K.G.). The NIH [grant number HD22486] provided support for the Oregon Zebrafish Facility.
SUPPLEMENTARY MATERIAL
Supplementary material for this article is available at http://dmm.biologists.org/lookup/suppl/doi:10.1242/dmm.011163/-/DC1
REFERENCES
- Amieva M. R., El-Omar E. M. (2008). Host-bacterial interactions in Helicobacter pylori infection. Gastroenterology 134, 306–323 [DOI] [PubMed] [Google Scholar]
- Amieva M. R., Vogelmann R., Covacci A., Tompkins L. S., Nelson W. J., Falkow S. (2003). Disruption of the epithelial apical-junctional complex by Helicobacter pylori CagA. Science 300, 1430–1434 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Asghar R. J., Parsonnet J. (2001). Helicobacter pylori and risk for gastric adenocarcinoma. Semin. Gastrointest. Dis. 12, 203–208 [PubMed] [Google Scholar]
- Barker N., Huch M., Kujala P., van de Wetering M., Snippert H. J., van Es J. H., Sato T., Stange D. E., Begthel H., van den Born M., et al. (2010). Lgr5(+ve) stem cells drive self-renewal in the stomach and build long-lived gastric units in vitro. Cell Stem Cell 6, 25–36 [DOI] [PubMed] [Google Scholar]
- Berghmans S., Murphey R. D., Wienholds E., Neuberg D., Kutok J. L., Fletcher C. D., Morris J. P., Liu T. X., Schulte-Merker S., Kanki J. P., et al. (2005). tp53 mutant zebrafish develop malignant peripheral nerve sheath tumors. Proc. Natl. Acad. Sci. USA 102, 407–412 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blaser M. J., Atherton J. C. (2004). Helicobacter pylori persistence: biology and disease. J. Clin. Invest. 113, 321–333 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bourzac K. M., Guillemin K. (2005). Helicobacter pylori-host cell interactions mediated by type IV secretion. Cell. Microbiol. 7, 911–919 [DOI] [PubMed] [Google Scholar]
- Buti L., Spooner E., Van der Veen A. G., Rappuoli R., Covacci A., Ploegh H. L. (2011). Helicobacter pylori cytotoxin-associated gene A (CagA) subverts the apoptosis-stimulating protein of p53 (ASPP2) tumor suppressor pathway of the host. Proc. Natl. Acad. Sci. USA 108, 9238–9243 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Censini S., Lange C., Xiang Z., Crabtree J. E., Ghiara P., Borodovsky M., Rappuoli R., Covacci A. (1996). cag, a pathogenicity island of Helicobacter pylori, encodes type I-specific and disease-associated virulence factors. Proc. Natl. Acad. Sci. USA 93, 14648–14653 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ceol C. J., Houvras Y., Jane-Valbuena J., Bilodeau S., Orlando D. A., Battisti V., Fritsch L., Lin W. M., Hollmann T. J., Ferré F., et al. (2011). The histone methyltransferase SETDB1 is recurrently amplified in melanoma and accelerates its onset. Nature 471, 513–517 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheesman S. E., Neal J. T., Mittge E., Seredick B. M., Guillemin K. (2011). Epithelial cell proliferation in the developing zebrafish intestine is regulated by the Wnt pathway and microbial signaling via Myd88. Proc. Natl. Acad. Sci. USA 108 Suppl. 1, 4570–4577 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Churin Y., Al-Ghoul L., Kepp O., Meyer T. F., Birchmeier W., Naumann M. (2003). Helicobacter pylori CagA protein targets the c-Met receptor and enhances the motogenic response. J. Cell Biol. 161, 249–255 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crosnier C., Vargesson N., Gschmeissner S., Ariza-McNaughton L., Morrison A., Lewis J. (2005). Delta-Notch signalling controls commitment to a secretory fate in the zebrafish intestine. Development 132, 1093–1104 [DOI] [PubMed] [Google Scholar]
- El-Etr S. H., Mueller A., Tompkins L. S., Falkow S., Merrell D. S. (2004). Phosphorylation-independent effects of CagA during interaction between Helicobacter pylori and T84 polarized monolayers. J. Infect. Dis. 190, 1516–1523 [DOI] [PubMed] [Google Scholar]
- El-Omar E. M., Rabkin C. S., Gammon M. D., Vaughan T. L., Risch H. A., Schoenberg J. B., Stanford J. L., Mayne S. T., Goedert J., Blot W. J., et al. (2003). Increased risk of noncardia gastric cancer associated with proinflammatory cytokine gene polymorphisms. Gastroenterology 124, 1193–1201 [DOI] [PubMed] [Google Scholar]
- Faro A., Boj S. F., Clevers H. (2009). Fishing for intestinal cancer models: unraveling gastrointestinal homeostasis and tumorigenesis in zebrafish. Zebrafish 6, 361–376 [DOI] [PubMed] [Google Scholar]
- Fearon E. R., Vogelstein B. (1990). A genetic model for colorectal tumorigenesis. Cell 61, 759–767 [DOI] [PubMed] [Google Scholar]
- Feng H., Stachura D. L., White R. M., Gutierrez A., Zhang L., Sanda T., Jette C. A., Testa J. R., Neuberg D. S., Langenau D. M., et al. (2010). T-lymphoblastic lymphoma cells express high levels of BCL2, S1P1, and ICAM1, leading to a blockade of tumor cell intravasation. Cancer Cell 18, 353–366 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Franco A. T., Israel D. A., Washington M. K., Krishna U., Fox J. G., Rogers A. B., Neish A. S., Collier-Hyams L., Perez-Perez G. I., Hatakeyama M., et al. (2005). Activation of beta-catenin by carcinogenic Helicobacter pylori. Proc. Natl. Acad. Sci. USA 102, 10646–10651 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guillemin K., Salama N. R., Tompkins L. S., Falkow S. (2002). Cag pathogenicity island-specific responses of gastric epithelial cells to Helicobacter pylori infection. Proc. Natl. Acad. Sci. USA 99, 15136–15141 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haramis A. P., Hurlstone A., van der Velden Y., Begthel H., van den Born M., Offerhaus G. J., Clevers H. C. (2006). Adenomatous polyposis coli-deficient zebrafish are susceptible to digestive tract neoplasia. EMBO Rep. 7, 444–449 [DOI] [PMC free article] [PubMed] [Google Scholar]
- He T. C., Sparks A. B., Rago C., Hermeking H., Zawel L., da Costa L. T., Morin P. J., Vogelstein B., Kinzler K. W. (1998). Identification of c-MYC as a target of the APC pathway. Science 281, 1509–1512 [DOI] [PubMed] [Google Scholar]
- Heisenberg C. P., Houart C., Take-Uchi M., Rauch G. J., Young N., Coutinho P., Masai I., Caneparo L., Concha M. L., Geisler R., et al. (2001). A mutation in the Gsk3-binding domain of zebrafish Masterblind/Axin1 leads to a fate transformation of telencephalon and eyes to diencephalon. Genes Dev. 15, 1427–1434 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Her G. M., Chiang C. C., Wu J. L. (2004). Zebrafish intestinal fatty acid binding protein (I-FABP) gene promoter drives gut-specific expression in stable transgenic fish. Genesis 38, 26–31 [DOI] [PubMed] [Google Scholar]
- Higashi H., Tsutsumi R., Muto S., Sugiyama T., Azuma T., Asaka M., Hatakeyama M. (2002). SHP-2 tyrosine phosphatase as an intracellular target of Helicobacter pylori Cag. Science 295, 683–686 [DOI] [PubMed] [Google Scholar]
- Higashi H., Nakaya A., Tsutsumi R., Yokoyama K., Fujii Y., Ishikawa S., Higuchi M., Takahashi A., Kurashima Y., Teishikata Y., et al. (2004). Helicobacter pylori CagA induces Ras-independent morphogenetic response through SHP-2 recruitment and activation. J. Biol. Chem. 279, 17205–17216 [DOI] [PubMed] [Google Scholar]
- Higashijima S., Okamoto H., Ueno N., Hotta Y., Eguchi G. (1997). High-frequency generation of transgenic zebrafish which reliably express GFP in whole muscles or the whole body by using promoters of zebrafish origin. Dev. Biol. 192, 289–299 [DOI] [PubMed] [Google Scholar]
- Kim B. M., Buchner G., Miletich I., Sharpe P. T., Shivdasani R. A. (2005). The stomach mesenchymal transcription factor Barx1 specifies gastric epithelial identity through inhibition of transient Wnt signaling. Dev. Cell 8, 611–622 [DOI] [PubMed] [Google Scholar]
- Kwan K. M., Fujimoto E., Grabher C., Mangum B. D., Hardy M. E., Campbell D. S., Parant J. M., Yost H. J., Kanki J. P., Chien C. B. (2007). The Tol2kit: a multisite gateway-based construction kit for Tol2 transposon transgenesis constructs. Dev. Dyn. 236, 3088–3099 [DOI] [PubMed] [Google Scholar]
- Lee A., O’Rourke J., De Ungria M. C., Robertson B., Daskalopoulos G., Dixon M. F. (1997). A standardized mouse model of Helicobacter pylori infection: introducing the Sydney strain. Gastroenterology 112, 1386–1397 [DOI] [PubMed] [Google Scholar]
- Mimuro H., Suzuki T., Tanaka J., Asahi M., Haas R., Sasakawa C. (2002). Grb2 is a key mediator of helicobacter pylori CagA protein activities. Mol. Cell 10, 745–755 [DOI] [PubMed] [Google Scholar]
- Muncan V., Faro A., Haramis A. P., Hurlstone A. F., Wienholds E., van Es J., Korving J., Begthel H., Zivkovic D., Clevers H. (2007). T-cell factor 4 (Tcf7l2) maintains proliferative compartments in zebrafish intestine. EMBO Rep. 8, 966–973 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murata-Kamiya N., Kurashima Y., Teishikata Y., Yamahashi Y., Saito Y., Higashi H., Aburatani H., Akiyama T., Peek R. M., Jr, Azuma T., et al. (2007). Helicobacter pylori CagA interacts with E-cadherin and deregulates the beta-catenin signal that promotes intestinal transdifferentiation in gastric epithelial cells. Oncogene 26, 4617–4626 [DOI] [PubMed] [Google Scholar]
- Nagy T. A., Wroblewski L. E., Wang D., Piazuelo M. B., Delgado A., Romero-Gallo J., Noto J., Israel D. A., Ogden S. R., Correa P., et al. (2011). β-Catenin and p120 mediate PPARδ-dependent proliferation induced by Helicobacter pylori in human and rodent epithelia. Gastroenterology 141, 553–564 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ng A. N., de Jong-Curtain T. A., Mawdsley D. J., White S. J., Shin J., Appel B., Dong P. D., Stainier D. Y., Heath J. K. (2005). Formation of the digestive system in zebrafish: III. Intestinal epithelium morphogenesis. Dev. Biol. 286, 114–135 [DOI] [PubMed] [Google Scholar]
- Nobili S., Bruno L., Landini I., Napoli C., Bechi P., Tonelli F., Rubio C. A., Mini E., Nesi G. (2011). Genomic and genetic alterations influence the progression of gastric cancer. World J. Gastroenterol. 17, 290–299 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohnishi N., Yuasa H., Tanaka S., Sawa H., Miura M., Matsui A., Higashi H., Musashi M., Iwabuchi K., Suzuki M., et al. (2008). Transgenic expression of Helicobacter pylori CagA induces gastrointestinal and hematopoietic neoplasms in mouse. Proc. Natl. Acad. Sci. USA 105, 1003–1008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ootani A., Li X., Sangiorgi E., Ho Q. T., Ueno H., Toda S., Sugihara H., Fujimoto K., Weissman I. L., Capecchi M. R., et al. (2009). Sustained in vitro intestinal epithelial culture within a Wnt-dependent stem cell niche. Nat. Med. 15, 701–706 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oshima H., Matsunaga A., Fujimura T., Tsukamoto T., Taketo M. M., Oshima M. (2006). Carcinogenesis in mouse stomach by simultaneous activation of the Wnt signaling and prostaglandin E2 pathway. Gastroenterology 131, 1086–1095 [DOI] [PubMed] [Google Scholar]
- Peek R. M., Jr, Blaser M. J. (2002). Helicobacter pylori and gastrointestinal tract adenocarcinomas. Nat. Rev. Cancer 2, 28–37 [DOI] [PubMed] [Google Scholar]
- Pinto D., Gregorieff A., Begthel H., Clevers H. (2003). Canonical Wnt signals are essential for homeostasis of the intestinal epithelium. Genes Dev. 17, 1709–1713 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Powell S. M., Zilz N., Beazer-Barclay Y., Bryan T. M., Hamilton S. R., Thibodeau S. N., Vogelstein B., Kinzler K. W. (1992). APC mutations occur early during colorectal tumorigenesis. Nature 359, 235–237 [DOI] [PubMed] [Google Scholar]
- Ranzani G. N., Luinetti O., Padovan L. S., Calistri D., Renault B., Burrel M., Amadori D., Fiocca R., Solcia E. (1995). p53 gene mutations and protein nuclear accumulation are early events in intestinal type gastric cancer but late events in diffuse type. Cancer Epidemiol. Biomarkers Prev. 4, 223–231 [PubMed] [Google Scholar]
- Ruijter J. M., Ramakers C., Hoogaars W. M., Karlen Y., Bakker O., van den Hoff M. J., Moorman A. F. (2009). Amplification efficiency: linking baseline and bias in the analysis of quantitative PCR data. Nucleic Acids Res. 37, e45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sato N., Meijer L., Skaltsounis L., Greengard P., Brivanlou A. H. (2004). Maintenance of pluripotency in human and mouse embryonic stem cells through activation of Wnt signaling by a pharmacological GSK-3-specific inhibitor. Nat. Med. 10, 55–63 [DOI] [PubMed] [Google Scholar]
- Sato T., van Es J. H., Snippert H. J., Stange D. E., Vries R. G., van den Born M., Barker N., Shroyer N. F., van de Wetering M., Clevers H. (2011). Paneth cells constitute the niche for Lgr5 stem cells in intestinal crypts. Nature 469, 415–418 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Segal E. D., Cha J., Lo J., Falkow S., Tompkins L. S. (1999). Altered states: involvement of phosphorylated CagA in the induction of host cellular growth changes by Helicobacter pylori. Proc. Natl. Acad. Sci. USA 96, 14559–14564 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Selbach M., Moese S., Hauck C. R., Meyer T. F., Backert S. (2002). Src is the kinase of the Helicobacter pylori CagA protein in vitro and in vivo. J. Biol. Chem. 277, 6775–6778 [DOI] [PubMed] [Google Scholar]
- Shibata A., Parsonnet J., Longacre T. A., Garcia M. I., Puligandla B., Davis R. E., Vogelman J. H., Orentreich N., Habel L. A. (2002). CagA status of Helicobacter pylori infection and p53 gene mutations in gastric adenocarcinoma. Carcinogenesis 23, 419–424 [DOI] [PubMed] [Google Scholar]
- Solnick J. V., Canfield D. R., Yang S., Parsonnet J. (1999). Rhesus monkey (Macaca mulatta) model of Helicobacter pylori: noninvasive detection and derivation of specific-pathogen-free monkeys. Lab. Anim. Sci. 49, 197–201 [PubMed] [Google Scholar]
- Stein M., Bagnoli F., Halenbeck R., Rappuoli R., Fantl W. J., Covacci A. (2002). c-Src/Lyn kinases activate Helicobacter pylori CagA through tyrosine phosphorylation of the EPIYA motifs. Mol. Microbiol. 43, 971–980 [DOI] [PubMed] [Google Scholar]
- Storer N. Y., Zon L. I. (2010). Zebrafish models of p53 functions. Cold Spring Harb. Perspect. Biol. 2, a001123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suzuki M., Mimuro H., Suzuki T., Park M., Yamamoto T., Sasakawa C. (2005). Interaction of CagA with Crk plays an important role in Helicobacter pylori-induced loss of gastric epithelial cell adhesion. J. Exp. Med. 202, 1235–1247 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suzuki M., Mimuro H., Kiga K., Fukumatsu M., Ishijima N., Morikawa H., Nagai S., Koyasu S., Gilman R. H., Kersulyte D., et al. (2009). Helicobacter pylori CagA phosphorylation-independent function in epithelial proliferation and inflammation. Cell Host Microbe 5, 23–34 [DOI] [PubMed] [Google Scholar]
- Tetsu O., McCormick F. (1999). Beta-catenin regulates expression of cyclin D1 in colon carcinoma cells. Nature 398, 422–426 [DOI] [PubMed] [Google Scholar]
- Tissier-Seta J. P., Mucchielli M. L., Mark M., Mattei M. G., Goridis C., Brunet J. F. (1995). Barx1, a new mouse homeodomain transcription factor expressed in cranio-facial ectomesenchyme and the stomach. Mech. Dev. 51, 3–15 [DOI] [PubMed] [Google Scholar]
- Tsukamoto T., Mizoshita T., Mihara M., Tanaka H., Takenaka Y., Yamamura Y., Nakamura S., Ushijima T., Tatematsu M. (2005). Sox2 expression in human stomach adenocarcinomas with gastric and gastric-and-intestinal-mixed phenotypes. Histopathology 46, 649–658 [DOI] [PubMed] [Google Scholar]
- Tsutsumi R., Higashi H., Higuchi M., Okada M., Hatakeyama M. (2003). Attenuation of Helicobacter pylori CagA x SHP-2 signaling by interaction between CagA and C-terminal Src kinase. J. Biol. Chem. 278, 3664–3670 [DOI] [PubMed] [Google Scholar]
- van de Water S., van de Wetering M., Joore J., Esseling J., Bink R., Clevers H., Zivkovic D. (2001). Ectopic Wnt signal determines the eyeless phenotype of zebrafish masterblind mutant. Development 128, 3877–3888 [DOI] [PubMed] [Google Scholar]
- Westerfield M. (2007). The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Danio rerio). Eugene, OR: University of Oregon Press [Google Scholar]
- White R. M., Cech J., Ratanasirintrawoot S., Lin C. Y., Rahl P. B., Burke C. J., Langdon E., Tomlinson M. L., Mosher J., Kaufman C., et al. (2011). DHODH modulates transcriptional elongation in the neural crest and melanoma. Nature 471, 518–522 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wirth H. P., Beins M. H., Yang M., Tham K. T., Blaser M. J. (1998). Experimental infection of Mongolian gerbils with wild-type and mutant Helicobacter pylori strains. Infect. Immun. 66, 4856–4866 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan D., Wiesmann M., Rohan M., Chan V., Jefferson A. B., Guo L., Sakamoto D., Caothien R. H., Fuller J. H., Reinhard C., et al. (2001). Elevated expression of axin2 and hnkd mRNA provides evidence that Wnt/beta-catenin signaling is activated in human colon tumors. Proc. Natl. Acad. Sci. USA 98, 14973–14978 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.