

Abstract
Meninges are a three tissue membrane primarily known as coverings of the brain. More in depth studies on meningeal function and ultrastructure have recently changed the view of meninges as a merely protective membrane. Accurate evaluation of the anatomical distribution in the CNS reveals that meninges largely penetrate inside the neural tissue. Meninges enter the CNS by projecting between structures, in the stroma of choroid plexus and form the perivascular space (Virchow-Robin) of every parenchymal vessel. Thus, meninges may modulate most of the physiological and pathological events of the CNS throughout the life. Meninges are present since the very early embryonic stages of cortical development and appear to be necessary for normal corticogenesis and brain structures formation. In adulthood meninges contribute to neural tissue homeostasis by secreting several trophic factors including FGF2 and SDF-1. Recently, for the first time, we have identified the presence of a stem cell population with neural differentiation potential in meninges. In addition, we and other groups have further described the presence in meninges of injury responsive neural precursors. In this review we will give a comprehensive view of meninges and their multiple roles in the context of a functional network with the neural tissue. We will highlight the current literature on the developmental feature of meninges and their role in cortical development. Moreover, we will elucidate the anatomical distribution of the meninges and their trophic properties in adult CNS. Finally, we will emphasize recent evidences suggesting the potential role of meninges as stem cell niche harbouring endogenous precursors that can be activated by injury and are able to contribute to CNS parenchymal reaction.
Keywords: Meninges, leptomeninges, arachnoid mater, pia mater, corticogenesis, neurogenesis, neural stem cells, neural progenitors, stem cell niche
Introduction
Meninges are formed by three tissue membranes that are primarily known as wrappers of the brain. They consist of dura mater, arachnoid and pia mater. The dura mater or pachymeninx (pachy-thick) is the outer membrane and forms a sac that envelops the other meningeal layers. It surrounds and supports the dural venous sinuses and it reflects in three infoldings, the first separating the two hemispheres of the cortex (falx cerebri), the second between the cerebellum and the occipital lobe (tentorium cerebelli and falx cerebelli) and the third covering the pituitary gland and the sella turcica [1]. The leptomeninges (lepto-, thin) are the inner membranes formed by two layers: the outer is named arachnoid (arachn-, spider) and the inner pia mater (pia-, tender) [1]. The arachnoid is linked to the pia by arachnoid trabeculae that span the subarachnoid space filled with cerebrospinal fluid (CSF) produced by choroid plexi [1].
The first known anatomical descriptions of the meninges and of the cerebrospinal fluid have been found in the Edwin Smith Papyrus, an Ancient Egyptian medical text on surgical trauma (ca. 1600 BCE). The text seems to be related to previous manuscripts attributed to Imhotep, an Egyptian physician (ca. 3000-2500 BCE) [2].
The primary function commonly attributed to meninges and CSF is to protect the central nervous system (CNS). This is mainly because meninges provide a tight anchoring of the CNS to the surrounding bones able to prevent side-to-side movement and providing stability. In the head, the external part of the dura, the periosteal layer, is tightly fixed to the skull. In the spinal cord, in addition to the periosteal layer, 21 pairs of denticulate ligaments pass from pia through arachnoid attaching to dura mater and vertebral bones. Moreover, meninges are filled up with the CSF letting the CNS “float” in it and thus cushioning hurting events [1].
In this review we will give a comprehensive view of meninges in the context of a functional network with the neural tissue. We will revise the literature highlighting the development of meninges, their distribution in adult CNS, their role in cortical development and in CNS homeostasis. Moreover, we will analyse new data suggesting the potential role of meninges as a stem cell niche harbouring endogenous injury-activated neural precursors.
Meninges development and distribution
In the early embryo, the neural tube is enveloped by a layer of mesenchymal cells that will result in the primary meninx. Both mesenchymal and neural crest-derived cells appear to be involved in the formation of the primary meninx that will differentiate during the embryo development by forming two different layers: the dura mater and the leptomeninges respectively [3]. Embryologic and anatomic differences appear to exist between the meninges of the brain and the spinal cord. Encephalic meninges have been described to originate from both the mesenchyme and the encephalic neural crest, while the meninges of the spine and of the caudal regions of the head originate from the paraxial mesenchyme [4,5]. The three meningeal layers are formed starting from a single pial meshwork structure composed of cells that progressively generates next to the pia mater two other arachnoid layers: an inner layer of cells with round nuclei and an outer layer of cells with oval nuclei; at the same time the dura mater layer forming next to the arachnoid shows spindle shaped cells with collagen deposition [6]. Similar developmental morphogenesis is observed in rat and human embryos [5,7].
In adult CNS the meninges cover and penetrate the brain deeply at every level of its organization: as large projections between major brain structures, as sheaths of blood vessels and as stroma of the choroid plexus [1]. Meninges also project between substructures. A major meningeal projection is located underneath the hippocampal formation [8,9]. This meningeal projection is continuous with the choroid plexus stroma. The cranial pia mater actually envelops the cerebrum and cerebellum and extends into the sulci and fissures. It also forms the non-neural roof of the third ventricle, the lateral ventricle and the fourth ventricle [10]. The reconsideration of the distribution of meninges in the CNS (Figure 1) set the stage for a more complex consideration of meningeal functions as modulator of CNS in homeostasis and disease.
Figure 1.
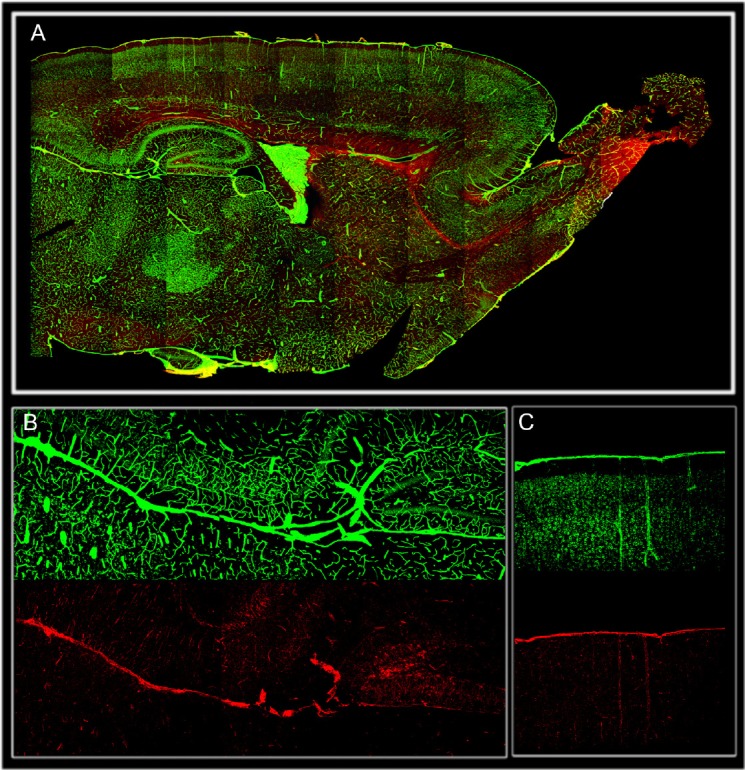
Distribution of brain meninges (laminin green) and nestin positive cells (red) in 15 days postnatal rat brain transversal section. (A). CNS brain meninges meninges cover and penetrate the brain deeply at every level of its organization including sheaths of blood vessels (perivascular space) and projections located underneath the hippocampal formation that continue with the choroid plexus. High magnification showing nestin positive cells associated with meningeal projection underneath the hippocampus (B), and penetrating the cortex as sheath of blood vessels (C). Meningeal stem cells (nestin) appear to be largely diffuse inside the parenchyma.
Meninges and the brain vasculature
The primitive meninx is also the site of origin of a complex vascular plexus that evolves to give rise to the brain vasculature [11-15]. The extracerebral vascular (pial) meningeal compartment is the first to develop by forming the major venous sinuses, the arachnoidal arteries and the veins that cover the surface of the developing and adult cerebral cortex [15]. Subsequently, sprouting vascular elements from pial capillaries pierce the brain external glial limiting membrane and penetrate the cortex establishing the intracerebral microvascular compartment. Every perforating vessel is associated to extroflexions of the meninges; a perivascular space is thus formed limited by the basal laminae of the vasculature on the inner side and of the glia on the external side. The pial vascular plexus vessels play an essential role in the brain intracerebral microvascularization and retain, throughout life, a remarkable activity, with continuous remodeling and readaptation to local functional needs [15].
The anatomical basis for the extension of the subarachnoid space within the perivascular spaces has been established in several mammalian species, including man [16-21,22]. It has been shown that the pia mater on the surface of the brain and spinal cord reflects onto the surface of blood vessels in the subarachnoid space, thus separating the perivascular and subpial spaces from the subarachnoid space [23-27]. Together with basal lamina, the perivascular space is endowed with a thin sheath of leptomeningeal cells that surrounds arterioles and arteries [16]. Perivascular space represents a perilymphatic drainage channel connected to the meningeal interstitial spaces, thus permitting the exchange of fluids and cells between brain and meninges [15]. Leptomeningeal cells forming the perivascular sheath have been characterized by light and electron microscopy and mostly identified as meningeal fibroblasts and meningeal macrophages [21]. Evidence of perivascular nestin-positive cells [28-30], (Figure 1) and nestin-positive proliferating endothelial cells [31], have also been reported. Meningeal fibroblasts were observed in the perivascular space of vessels upstream of the capillaries and forming a multilayer around the larger arteries [10,32]. Perivascular fibroblasts and macrophages form a network by contacting each other with no discontinuity along meninges and the longitudinal axis of the blood vessels [33]. Of note is that the blood-brain-barrier (BBB) is mainly located at the level of the endothelium, thus the extraparenchymal cells residing in the perivascular space are located beyond the BBB [34].
Role of meninges in corticogenesis
Several studies have shown that meninges are essential for the correct development of the CNS. However, all the molecular mechanisms by which meninges and the meningeal cells participate in this process are still to be elucidated.
Meningeal tissue has been shown to be required for encephalon development [4,35]. The mesodermal components of the forebrain meninges provide the endothelial walls of blood vessels that penetrate the neuroepithelium, while the neural crest originating from the posterior neural folds yields pericytes and connective tissue. The removal of the posterior diencephalic and mesencephalic neural folds prevents meninges development and causes apoptosis of the neuroepithelium of the entire forebrain [4]. Etchevers et al. have shown that the presence of primitive leptomeninges is needed for the survival and subsequent growth of the developing proencephalon. When the neural folds are ablated, paraxial mesoderm can replace the neural crest cells generating primitive leptomeninges that allow encephalon development [4].
Later in development, destruction of the meninges overlying the cerebellum leads to cerebellar hypoplasia, formation of neuronal ectopia and gliosis in the subarachnoid space and reduction of the total number of granular cells [36-40]. In the hippocampus, destruction of the meninges may induce secondary malformations of the dentate gyrus [41].
Due to their strategic position within the parenchyma and the connection with the vasculature, meninges have the potential to provide short-range factors to neural cells of the developing brain structures [42]. An example is the signal that involves the stromal-derived factor (SDF1)- and CXC chemokine receptor 4 (CXCR4)- dependent pathway. This signal is involved in the homing of various kinds of stem cells as well as in forebrain development [43,44]. Meninges secrete SDF1 that guides the tangential migration of Cajal-Retzius cells and cortical interneurons along the cortical marginal zone, ensuring their correct distribution during corticogenesis [45-48].
Other morphogens, including retinoic acid, are active at the meningeal level. Indeed, meninges express high levels of retinoic acid, which appears to be critical for early cortical neuron generation [49] and for anterior hindbrain development [50]. Recently, meninges have been shown to be involved in the generation of a cascade of morphogenic signals regulating corpus callosum development; in this case, meninges are involved by producing BMP7, an inhibitor of callosal axon outgrowth. This activity is overcome by the induction of expression of Wnt3 by the callosal pathfinding neurons, which antagonizes the inhibitory effects of BMP7 [51].
Due to the ability to bind several morphogenetic and trophic factors, a special role for corticogenesis is reserved to the extracellular matrix forming the basal membrane. Meningeal cells actively participate in the formation of the extracellular matrix (ECM). Collagens are the most abundant ECM structures in meninges, where the most represented types are I, III, and IV. In addition to collagen, meningeal cells synthesize non-collagen proteins including fibronectin, laminin and tenascin [52]. The pial basal membrane is an important anchor for the endfeet of radial processes that originate from neural progenitors cells residing in the VZ; moreover, it is a physical barrier for the migrating neurons. During cortical and cerebellar development, the radial processes provide a migratory scaffold for neurons that ensures cortical cellular layering. Genetic ablation of components of the pial basal membrane (i.e laminin components alpha1, alpha2 and alpha5) or of the proteins that mediate extracellular matrix attachment leads to a loss of pial integrity, radial endfeet detachment and disruption of cortical and cerebellar histogenesis [53,54]. Premature detachment of radial endfeet also leads to increased neural progenitor cell death and, eventually, reduced production of cortical neurons [53,54]. Moreover, mice lacking laminin subtypes show alterations in the distribution of connexin (Cx) 43, a gap junction protein that is expressed in the arachnoid and pia mater of the meninges [55]. It is noteworthy that ablation of this protein, like in the glia-specific Cx43 knockout mice, results in reduction in size of the cerebellum, with appearance of granule cell ectopies and dislamination of Purkinje cells, granule cells, and Bergmann glia [56]. It can be hypothesized that laminins have a role in the functional localization of connexins. Targeted deletion of focal adhesion kinase in meninges elicited altered cortical histogenesis similar to type II cobblestone lissencephaly, with clusters of neurons invading the marginal zone with retraction of radial glial endfeet, midline fusion of brain hemispheres, and gliosis, as seen in congenital muscular dystrophy [57].
Role of meninges in CNS homeostasis
The protective function of meninges was considered the major functional significance of this structure. Indeed, meninges were considered to physically protect the brain from traumas and to be devoid of any functional connection with the brain parenchyma. A continuous mat of extracellular matrix molecules, including laminin, fibronectin, collagens IV, XV and XVIII and heparan sulfate proteoglycans [58], is localized at the surface of the brain as well as around the blood vessels inside the brain. This material was considered to form a sharp interface separating (both anatomically and functionally) the brain parenchyma (neurons and glia) from the extraparenchymal tissues (meninges and vessels).
More in-depth studies of the meninges ultrastructure have contributed to change this limited view of meninges function [21]. Meninges contain a laminin-enriched ECM organized in fractones relevant for sequestering and concentrating growth factors [59] that have the potential to modulate stem cell homeostasis and cortical function. Furthermore, meninges are an important source of several trophic factors [47-49], including FGF-2 [60], insulin-like growth factor-II [61,62], CXCL12 [45,63-65], and retinoic acid [49-51]. Of note is also that numerous growth factors and cytokines, including those promoting stem cell proliferation and differentiation (i.e. FGF2, EGF), are heparin-binding molecules [66,67] that can bind to the heparan sulfate chains of heparan sulfate proteoglycans that are abundant in meninges. Interestingly, cells of the meninges have been shown to be highly responsive to principal mitogens such as EGF, FGF-2, and BDNF [68,69]. Heparan sulfate proteoglycans enrichments have been found in regions associated to neural stem cell proliferation [9,59]. These observations suggest the existence of a functional system involved in the regulation of growth factors in neurogenic regions and in meninges. It has been proposed by Mercier et al. that heparan sulfate proteoglycans regions course as a single anatomical system that comprises the olfactory bulb, the rostral migratory stream, the subventricular zone, the sub-callosum and subcapsule zones and the meninges of the ventral hippocampal neurogenic zone [9].
A further indication that meninges are functionally linked to the neural tissue is the presence of gap junction proteins. Cx43, Cx30 and Cx26 have been found along a network of cells in the meninges and in their projections into the brain, including meningeal sheaths of blood vessels and stroma of the choroid plexus [60,70-72]. The distribution of these proteins suggests the existence of anatomical and functional interactions between meningeal cells, meningeal-perivascular cells, ependymocytes and astrocytes [60,71,73] capable to provide a rapid mean to spread signals. Altogether the data indicate that meninges are an important structure that modulates brain function during embryogenesis and adult life.
Meninges: a novel stem cell niche
Functional complexity linked to cellular complexity
Different cell populations have been described in meninges such as fibroblasts, perivascular and meningeal macrophages, mast cells, pericytes, smooth muscle cells and endothelial cells. Recently, interstitial cells characterized by a small cell body and extremely long, moniliform, cell processes - telopodes named tenocytes [73] have been described. Adding complexity to the potential role of meninges in CNS function was the identification of a stem cell population sharing many features with the bona fide neural stem cells [28,74-76].
Here, we provide a description of the current knowledge on the in vitro cultured meningeal cells, reviewing the established studies on meningeal fibroblasts, the more recent findings on pericytes and boundary cap cells and the latest observations suggesting the presence of a stem cell population in meninges.
Meningeal fibroblasts
The most commonly used markers to identify meningeal fibroblasts are fibronectin, vimentin, chondroitin sulfates (recognized also by CS-56 antibody) and retinaldehyde dehydrogenase type 2 [77]. Meningeal fibroblasts have been shown to play a primary role in the acute and subacute phases of injury-induced parenchymal reaction in CNS, as they promptly infiltrate the lesion site [78,79]. Here, the reactive astrogliosis and the meningeal cells form a glial-fibroblast interface that produces new basal lamina that, in combination with the glial endfeet, reforms the glia limitans [80,81]. This process is believed to be essential for restoring the blood-brain barrier and reestablishing CNS homeostasis [80]. In a long series of experiments stretching back for many years, the lesion scar has been identified as one critical element that impedes axonal regeneration in the adult mammalian CNS [82,83]. After lesion, CNS axons start to sprout over short distances, but almost all of them stop abruptly at the lesion scar border and fail to traverse it [84-86]. In vitro cultures of meningeal fibroblasts have been classically used as an in vitro model of injury-induced scar, allowing investigation of the mechanisms involved in the formation of the barrier to axon regeneration [80,87,88]. Nevertheless, some reports described in vitro [89] and in vivo [90,91] axonal growth-promoting properties of cultured meningeal fibroblasts. Moreover, cells from meninges were able to induce neuronal differentiation of embryonic stem cells [92]. As observed in vivo at CNS lesion sites [93-96], in vitro cultured meningeal fibroblasts are induced by TGF-β1 to produce high levels of ECM components, like Col IV, and various CSPGs, such as biglycan, versican, decorin, neurocan and phosphocan, tenascin-C, semaphorin3A and EphB, while astrocytes are induced to increase the expression of neurocan, phosphocan and ephrin-B2 [89,97,98].
In vitro modelling of the scar confirms that attachment of neurons and extension of neurites are suppressed when they are cultured on a layer of fibroblasts, and they are dramatically diminished if co-cultured on fibroblasts and astrocytes [79,99]. These results suggest that inhibitory effects are mainly triggered i) by an indirect negative effect of fibroblasts on the growth-promoting abilities of astrocytes and ii) by a direct inhibition of fibroblast on neurite growth mediated by expression of inhibitory molecules such as NG2 and Sema3A [100].
Pericytes
CNS microvascular pericytes reside in the perivascular space, an extrofection of the leptomeninges, and play an important role in vascular homeostasis by producing different ECM components [101,102]. Pericytes respond to insults to the CNS by secreting different regulatory molecules and migrating into the perivascular space [103] and have been reported to differentiate in vitro into macrophage-like cells and osteoblasts. In 2006, Dore-Duffy et al. showed that pericytes isolated from the microvessels of the brain cortex exhibit multipotential stem cell activity [104]. These stem cells express NG2 and nestin and can be cultured in vitro as neurospheres. Cultured cells showed cell renewal properties and could be induced to differentiate into pericytes, neurons, astrocytes and oligodendrocytes. Other numerous studies reported pericytes differentiation into mesenchymal lineage cells, such as adipocytes, smooth-muscle cells and endothelial cells [105].
Boundary cap cells
During development, boundary cap (BC) cells regulate the entry of the forming afferent nerve in the spinal cord and cannot be found after postnatal day 6. BC cells are neural crest-derived cells that are found in clusters in the dorsal entry zone and the ventral exit zone of nerve roots in the spinal cord, at the border between the CNS and the PNS in direct contact with the pial lamina [106-109]. BC cells are easily identified in meninges by the exclusive expression of monoamine oxidase type B and, between embryonic days 10.5-15.5, of the zinc finger transcription factor Krox20/Egr2. BC cells have been extracted from meninges of mouse embryo at day 12 [110,111]. In vivo, BC cells generate Schwann cells in the nerve root region and differentiate into nociceptive neurons and glial satellite cells after migration in the dorsal root ganglia [112]; cultured BC cells express both Schwann cell (i.e. Sox9, nestin, Sox2 and Musashi) and neural crest related markers (Sox10, p75, polysialylated neural cells adhesion molecule), suggesting that they are in an intermediate stage of differentiation [113]. In vitro cultured BC cells were capable to differentiate in glia, sensory neurons and smoothmuscle-like cells [114]. Moreover, more recent studies showed that cultured BC cells could differentiate in vitro into mature Schwann cells capable of remyelinating DRG neurons [115-117]. In vivo differentiation potential of cultured BC cells has also been analysed [112,116,117]. Interestingly, boundary cap cells have been found to differentiate differently depending on the site of transplantation. They differentiate into neurons and astrocytes in the CNS, into oligodendrocytes in the demyelinated spinal cord and in Schwann cells, glial satellite cells and nociceptive neurons in the DRG.
Meningeal stem/progenitor cells
Supporting the concept of meninges as putative neural stem cell niche, are the peculiar morphofunctional properties of meninges themselves. Indeed, meninges may have the potential to modulate stem cell homeostasis since they contain laminin-enriched ECM organized in fractones [21], N-sulfated heparan sulphate molecules, functional gap junctions and secrete several trophic factors. More recently, Popescu et al. presented evidence for the presence of telocytes in meninges and choroid plexus. Telocytes are interstitial cells that appear to be in close contact with stem cells and to be able to regulate the stem cell niche by generation of intercellular signaling [73]. We have analyzed the leptomeningeal compartment of the rat brain and have identified a nestin-positive cell population in brain meninges of embryonic and adult rodents [28,74]. Nestin is an intermediate filament of neuroepithelial derivation [118] that has been detected in stem/progenitor cells of neural and non-neural tissues. The cells extracted from the meningeal biopsies could be grown as neurospheres, as in the case of cells extracted from classic neurogenic regions [28]. Meningeal cultured cells could be differentiated in vitro and in vivo into either neurons (identified by neuronal phenotypic and electrophysiological properties) or into mature MBP-expressing oligodendrocytes (Figure 2A). Nestin-positive cells have been identified also in human encephalic [119] and spinal cord meninges [74], indicating the interspecies relevance of our observation. The data are in agreement with previous indications that NSCs were present in the choroid plexus of the adult rat [120] and that cells from human meninges expressed some neural markers, such as neurofilament protein and neuron-specific enolase when cultured in vitro, while they express GFAP after transplantation in rat brains [121-123].
Figure 2.
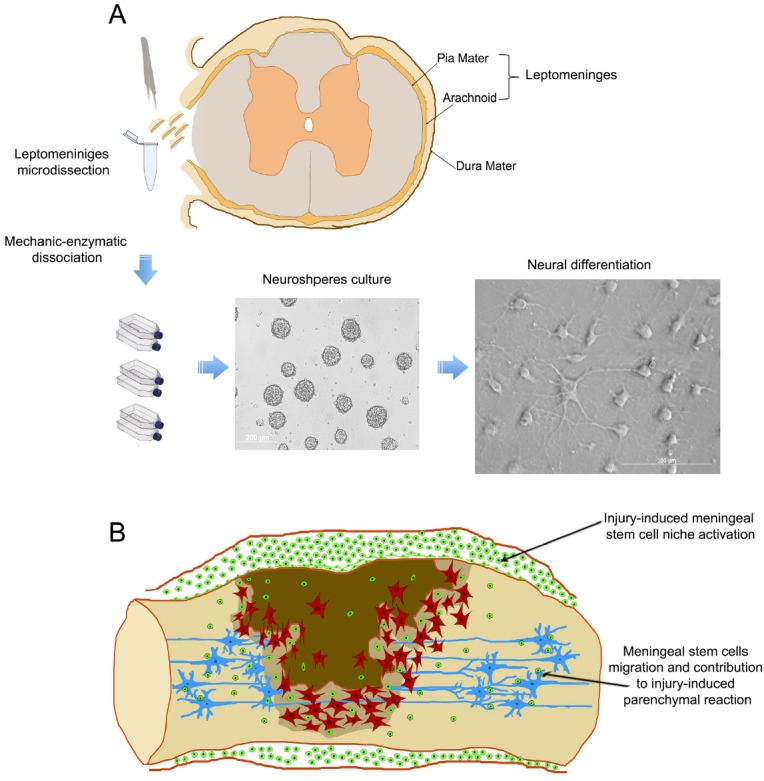
Meningeal stem cells. (A). Meningeal stem cells from adult spinal cord can be microdissected, expanded in vitro and induced to differentiate into neural cells. (B). schematic representation of the activation of the stem cell niche in meninges following spinal cord injury. Meningeal stem cells proliferate, increase in number and migrate inside the parenchyma contributing to the parenchymal reaction.
Meninges: an injury responsive stem cells niche
To assess a possible significance of meninges as a niche for stem cells with functional implication in CNS physiopathology, our group investigated the presence of relatively quiescent, mitotically-active transient-amplifying cells and neuroblasts in spinal cord meninges. We found that nestin-positive cells endowed with self-renewal and proliferative properties, and doublecortin (DCX)-positive cells are present in adult spinal cord meninges. A further paradigm to define a stem cell niche is activation by diseases [124-128]. Activation is defined by proliferation and increased number of all the cell subsets [129,130] participating in a stem cell niche [131] and migration of precursor cells in the parenchyma undergoing reaction. Detailed analysis of changes in gene expression in meningeal spinal cord cells induced by injury showed increase in several stemness-related genes, including Pou5f1/Oct4 and Nanog, and of neural precursor markers, such as Nestin, Dcx, Pax6 and Klhl1 [74]. Moreover, we observed a significant increase of both nestin- and DCX-positive cells in meninges, indicating a general amplification of the meningeal stem cell pools associated to the injury-induced activation [74]. In addition, nestin-negative/DCX-positive cells appeared, suggesting a progression toward the neural fate. We also used an in vivo labeling approach to show that meningeal cells migrated and accumulated in the fibrotic scar and were also present in the glial scar and in the perilesion parenchyma. Interestingly, migrating meningeal cells also penetrated the dorsal horn 30 days after the injury. These data suggest that at least part of the proliferating cells present in the lesioned parenchyma originate from meninges (Figure 2B). Moreover, some of the migrating meningeal cells expressed the same markers (nestin and DCX) that are transiently expressed by neural precursors within classic neurogenic niches of the embryo and the adult brain, providing new insights into the complexity of the parenchymal reaction to a traumatic injury [74]. Almost at the same time, Nakagomi et al. demonstrated that the leptomeninges exhibit neural stem/progenitor cell (NSPC) activity in response to ischemia in adult brains [75]. Pial ischemia-induced NSPCs (iNSPCs) expressed the NSPC marker nestin, formed neurosphere-like cell clusters with self-renewal ability, and differentiated into neurons, astrocytes, and oligodendrocytes [75], indicating that they have stem cell capacity similar to other NSPC types. Moreover, the same group shows that leptomeningeal cells in post-stroke brain express the immature neuronal marker doublecortin as well as nestin [76] and that these cells can migrate into the post-stroke cortex.
These new findings highlight a new role for leptomeninges in CNS repair in response to CNS injury and suggest the existence of a new stem cell niche in the meninges that participates in the reaction occurring in the parenchyma following injury [29]. Although the in vivo role and the fate of the meningeal stem/precursor cells remain to be fully elucidated, these observations may have relevant consequences for understanding the mechanisms of stem cell activation in CNS diseases and the nature and origin of neural cell precursors appearing in ectopic non neurogenic regions of the brain. The superficial location and widespread distribution of meninges make this site an attractive source of neural cell precursors to be used for regenerative medicine applied to pathologies of the spinal cord and/or the brain, especially considering their possible use in an autologous setting.
Conclusion
Data in literature indicate a complex role of the meninges in CNS homeostasis.
The idea that the continuous membranous mat of extracellular matrix localized at the surface of the brain and spinal cord as well as around the blood vessels in the CNS, forms a sharp interface separating (both anatomically and functionally) the brain parenchyma (neurons and glia) from the extraparenchymal tissues (meninges and vessels) is changing. Meninges are considered to form a functional network together with parenchymal tissue contributing to CNS physiology and injury-induced reactions.
Primitive meninges form as early as the neural tube develops. They are necessary for the development of the whole forebrain and for the generation of the primitive brain vasculature. In adult, meninges cover and penetrate the CNS deeply at every level of its organization, including formation of large projections between major brain structures. Moreover, all the major arteries supplying the brain pass through leptomeninges and form branches while penetrating the cortex. Leptomeninges form a complex microenvironment by producing trophic factors and extracellular matrix components that have key functions for the normal cortex development.
On top of these anatomical considerations, our discovery of a new stem cell population endowed with neural differentiation potential in the meninges sheds a new light on meninges function in physiology and in pathology. This stem cell population shows in vivo self renewal and proliferation properties that are activated by spinal cord injury and brain stroke. Meningeal activated stem/precursor cells are able to migrate to the parenchyma contributing to parenchymal reaction, providing new insights for understanding the complexity of the parenchymal reaction to injury. Altogether, the data suggest that meninges are a functional stem cell niche. The nature and the origin of these meningeal stem/precursor cells have to be further characterized as far as their potential contribution to the neural tissue both in physiological and pathological conditions. Given their distribution, meninges may be a strategic net helping intracerebral migration and distribution of activated precursor cells at specific sites. Meninges are also more accessible than other neural stem cell niches, an aspect that has interesting implication for sampling of NSCs for regenerative medicine.
In conclusion, we reviewed the several functions of meninges showing that meninges are not merely a protecting sac coverings the CNS but form a functional syncytium with the neural tissue. Further studies are needed to clarify the presence and the role of the stem cells in meninges during development and the possible function of meninges as anatomical net for stem cell migration to sites of integration into the normal and injured tissue [29]. This will open new prospectives for the pharmacological modulation of meningeal endogenous stem cells for the regenerative therapies of neurode-generative diseases.
Acknowledgement
This work was supported by Italian Ministry of University and Scientific Research (PRIN 2008-2009), italian spinal cord injured patients association, FAIP (Federazione delle Associazioni Italiane Para-tetraplegici) and GALM (Gruppo Animazione Lesionati Midollari) and International Foundation for Research in Paraplegie-IRP 2012.
References
- 1.Jacobson S, Marcus EM. Neuroanatomy for the Neuroscientist. Springer; 2008. pp. 325–331. [Google Scholar]
- 2.Wilkins RH. Neurosurgical Classics. Thieme; 1992. p. 1. [Google Scholar]
- 3.Beatriz M, Lopes S Meninges. Embryology. In: Joung H. Lee., editor. Meningiomas. Springer; 2009. pp. 25–29. [Google Scholar]
- 4.Etchevers HC, Couly G, Vincent C, Le Douarin NM. Anterior cephalic neural crest is required for forebrain viability. Development. 1999;126:3533–3543. doi: 10.1242/dev.126.16.3533. [DOI] [PubMed] [Google Scholar]
- 5.O’Rahilly R, Müller F. The meninges in human development. J Neuropathol Exp Neurol. 1986;45:588–608. [PubMed] [Google Scholar]
- 6.Vivatbutsiri P, Ichinose S, Hytönen M, Sainio K, Eto K, Iseki S. Impaired meningeal development in association with apical expansion of calvarial bone osteogenesis in the Foxc1 mutant. J Anat. 2008;212:603–611. doi: 10.1111/j.1469-7580.2008.00893.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Angelov DN, Vasilev VA. Morphogenesis of rat cranial meninges. A light- and electron-microscopic study. Cell Tissue Res. 1989;257:207–216. doi: 10.1007/BF00221652. [DOI] [PubMed] [Google Scholar]
- 8.Mercier F, Kitasako JT, Hatton GI. Fractones and other basal laminae in the hypothalamus. J Comp Neurol. 2003;455:324–340. doi: 10.1002/cne.10496. [DOI] [PubMed] [Google Scholar]
- 9.Mercier F, Arikawa-Hirasawa E. Heparan sulfate niche for cell proliferation in the adult brain. Neurosci Lett. 2012;510:67–72. doi: 10.1016/j.neulet.2011.12.046. [DOI] [PubMed] [Google Scholar]
- 10.Mercier F, Hatton GI. Meninges and perivasculature as mediators of CNS plasticity. Adv Mol Cell Biol. 2003;31:215–253. [Google Scholar]
- 11.Kuban KC, Gilles FH. Human telencephalic angiogenesis. Ann Neurol. 1985;17:539–548. doi: 10.1002/ana.410170603. [DOI] [PubMed] [Google Scholar]
- 12.Kurz H, Horn J, Christ B. Morphogenesis of embryonic CNS vessels. Cancer Treat Res. 2004;117:33–50. doi: 10.1007/978-1-4419-8871-3_2. [DOI] [PubMed] [Google Scholar]
- 13.Marin-Padilla M. Early vascularization of the embryonic cerebral cortex: Golgi and electron microscopic studies. J Comp Neurol. 1985;241:237–249. doi: 10.1002/cne.902410210. [DOI] [PubMed] [Google Scholar]
- 14.Nelson MD, Gonzalez-Gomez I, Gilles FH. The search for human telencephalic ventriculofugal arteries. AJNR Am J Neuroradiol. 1991;12:215–222. [PMC free article] [PubMed] [Google Scholar]
- 15.Marín-Padilla M, Knopman DS. Developmental aspects of the intracerebral microvasculature and perivascular spaces: insights into brain response to late-life diseases. J Neuropathol Exp Neurol. 2011;70:1060–1069. doi: 10.1097/NEN.0b013e31823ac627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Zhang ET, Inman CB, Weller RO. Interrelationships of the pia mater and the perivascular (Virchow-Robin) spaces in the human cerebrum. J Anat. 1990;170:111–123. [PMC free article] [PubMed] [Google Scholar]
- 17.Reina-De La Torre F, Rodriguez-Baeza A, Sahuquillo-Barris J. Morphological characteristics and distribution pattern of the arterial vessels in human cerebral cortex: a scanning electron microscope study. Anat Rec. 1998;251:87–96. doi: 10.1002/(SICI)1097-0185(199805)251:1<87::AID-AR14>3.0.CO;2-7. [DOI] [PubMed] [Google Scholar]
- 18.Rodriguez-Baeza A, Reina-De La Torre F, Ortega-Sanchez M, Sahuquillo-Barris J. Perivascular structures in corrosion casts of the human central nervous system: a confocal laser and scanning electron microscope study. Anat Rec. 1998;252:176–184. doi: 10.1002/(SICI)1097-0185(199810)252:2<176::AID-AR3>3.0.CO;2-1. [DOI] [PubMed] [Google Scholar]
- 19.Jones EG. On the mode of entry of blood vessels into the cerebral cortex. J Anat. 1970;106:507–520. [PMC free article] [PubMed] [Google Scholar]
- 20.Nonaka H, Akima M, Nagayama T, Hatori T, Zhang Z, Ihara F. Microvasculature of the human cerebral meninges. Neuropathology. 2003;23:129–135. doi: 10.1046/j.1440-1789.2003.00487.x. [DOI] [PubMed] [Google Scholar]
- 21.Mercier F, Kitasako JT, Hatton GI. Anatomy of the brain neurogenic zones revisited: fractones and the fibroblast/macrophage network. J Comp Neurol. 2002;451:170–188. doi: 10.1002/cne.10342. [DOI] [PubMed] [Google Scholar]
- 22.Pollock H, Hutchings M, Weller RO, Zhang ET. Perivascular spaces in the basal ganglia of the human brain: their relationship to lacunes. J Anat. 1997;191:337–346. doi: 10.1046/j.1469-7580.1997.19130337.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Krahn V. The pia mater at the site of the entry of blood vessels into the central nervous system. Anat Embryol (Berl) 1982;164:257–263. doi: 10.1007/BF00318509. [DOI] [PubMed] [Google Scholar]
- 24.Krisch B, Leonhardt H, Oksche A. Compartments and perivascular arrangement of the meninges covering the cerebral cortex of the rat. Cell Tissue Res. 1984;238:459–474. doi: 10.1007/BF00219861. [DOI] [PubMed] [Google Scholar]
- 25.Hutchings M, Weller RO. Anatomical relationships of the pia mater to cerebral blood vessels in man. J Neurosurg. 1986;65:316–325. doi: 10.3171/jns.1986.65.3.0316. [DOI] [PubMed] [Google Scholar]
- 26.Alcolado R, Weller RO, Parrish EP, Garrod D. The cranial arachnoid and pia mater in man: anatomical and ultrastructural observations. Neuropatol App Neurobiol. 1988;14:1–17. doi: 10.1111/j.1365-2990.1988.tb00862.x. [DOI] [PubMed] [Google Scholar]
- 27.Nicholas DS, Weller RO. The fine anatomy of the human spinal meninges. A light and scanning electron microscopy study. J Neurosurg. 1988;69:276–282. doi: 10.3171/jns.1988.69.2.0276. [DOI] [PubMed] [Google Scholar]
- 28.Bifari F, Decimo I, Chiamulera C, Bersan E, Malpeli G, Johansson J, Lisi V, Bonetti B, Fumagalli G, Pizzolo G, Krampera M. Novel stem/progenitor cells with neuronal differentiation potential reside in the leptomeningeal niche. J Cell Mol Med. 2009;13:3195–3208. doi: 10.1111/j.1582-4934.2009.00706.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Decimo I, Bifari F, Krampera M, Fumagalli G. Neural Stem Cell Niches in Health and Diseases. Curr Pharm Des. 2012;18:1755–1783. doi: 10.2174/138161212799859611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Alliot F, Rutin J, Leenen PJ, Pessac B. Pericytes and periendothelial cells of brain parenchyma vessels co-express aminopeptidase N, aminopeptidase A, and nestin. J Neurosci Res. 1999;58:367–378. [PubMed] [Google Scholar]
- 31.Suzuki S, Namiki J, Shibata S, Mastuzaki Y, Okano H. The neural stem/progenitor cell marker Nestin is expressed in proliferative endothelial cells, but not in mature vasculature. J Histochem Cytochem. 2010;58:721–730. doi: 10.1369/jhc.2010.955609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Peters A, Palay SL, Webster HD. The fine structure of the nervous system: neurons and their supporting cells. Oxford University Press; 1990. p. 428. [Google Scholar]
- 33.Mercier F, Hatton GI. Immunocytochemical basis for a meningeo-glial network. J Comp Neurol. 2000;420:445–465. doi: 10.1002/(sici)1096-9861(20000515)420:4<445::aid-cne4>3.0.co;2-3. [DOI] [PubMed] [Google Scholar]
- 34.Schwartz M, Shechter R. Protective autoimmunity functions by intracranial immunosurveillance to support the mind: The missing link between health and disease. Mol Psychiatry. 2010;15:342–354. doi: 10.1038/mp.2010.31. [DOI] [PubMed] [Google Scholar]
- 35.Halfter W, Dong S, Yip YP, Willem M, Mayer U. A critical function of the pial basement membrane in cortical histogenesis. J Neurosci. 2002;22:6029–6040. doi: 10.1523/JNEUROSCI.22-14-06029.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Hartmann D, Schulze M, Sievers J. Meningeal cells stimulate and direct the migration of cerebellar external granule cells in vitro. J Neurocytol. 1998;27:395–409. doi: 10.1023/a:1006998609999. [DOI] [PubMed] [Google Scholar]
- 37.Hartmann D, Ziegenhagen MW, Sievers J. Meningeal cells stimulate neuronal migration and the formation of radial glial fascicles from the cerebellar external granular layer. Neurosci Lett. 1998;244:129–132. doi: 10.1016/s0304-3940(98)00154-2. [DOI] [PubMed] [Google Scholar]
- 38.Sievers J, von Knebel Doeberitz C, Pehlemann FW, Berry M. Meningeal cells influence cerebellar development over a critical period. Anat Embryol (Berl) 1986;175:91–100. doi: 10.1007/BF00315459. [DOI] [PubMed] [Google Scholar]
- 39.Sievers J, Pehlemann FW, Gude S, Berry M. Meningeal cells organize the superficial glia limitans of the cerebellum and produce components of both the interstitial matrix and the basement membrane. J Neurocytol. 1994;23:135–149. doi: 10.1007/BF01183867. [DOI] [PubMed] [Google Scholar]
- 40.von Knebel Doeberitz C, Sievers J, Sadler M, Pehlemann FW, Berry M, Halliwell P. Destruction of meningeal cells over the newborn hamster cerebellum with 6-hydroxydopamine prevents foliation and lamination in the rostral cerebellum. Neuroscience. 1986;17:409–426. doi: 10.1016/0306-4522(86)90256-3. [DOI] [PubMed] [Google Scholar]
- 41.Hartmann D, Sievers J, Pehlemann FW, Berry M. Destruction of meningeal cells over the medial cerebral hemisphere of newborn hamsters prevents the formation of the infrapyramidal blade of the dentate gyrus. J Comp Neurol. 1992;320:33–61. doi: 10.1002/cne.903200103. [DOI] [PubMed] [Google Scholar]
- 42.Siegenthaler JA, Pleasure SJ. We have got you 'covered': how the meninges control brain development. Curr Opin Genet Dev. 2011;21:249–255. doi: 10.1016/j.gde.2010.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Mithal DS, Banisadr G, Miller RJ. CXCL12 Signaling in the Development of the Nervous System. J Neuroimmune Pharmacol. 2012 Jan 21; doi: 10.1007/s11481-011-9336-x. [Epub ahead of print] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Zou YR, Kottmann AH, Kuroda M, Taniuchi I, Littman DR. Function of the chemokine receptor CXCR4 in haematopoiesis and in cerebellar development. Nature. 1998;393:595–599. doi: 10.1038/31269. [DOI] [PubMed] [Google Scholar]
- 45.Borrell V, Marin O. Meninges control tangential migration of hem- derived Cajal-Retzius cells via CXCL12/CXCR4 signaling. Nat Neurosci. 2006;9:1284–1293. doi: 10.1038/nn1764. [DOI] [PubMed] [Google Scholar]
- 46.Paredes MF, Li G, Berger O, Baraban SC, Pleasure SJ. Stromal-derived factor-1 (CXCL12) regulates laminar position of Cajal-Retzius cells in normal and dysplastic brains. J Neurosci. 2006;26:9404–9012. doi: 10.1523/JNEUROSCI.2575-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.López-Bendito G, Sánchez-Alcañiz JA, Pla R, Borrell V, Picó E, Valdeomillos M, Marin O. Chemokine signaling controls intracortical migration and final distribution of GABAergic interneurons. J Neurosci. 2008;28:1613–1624. doi: 10.1523/JNEUROSCI.4651-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Zarbalis K, Choe Y, Siegenthaler JA, Orosco LA, Pleasure SJ. Meningeal defects alter the tangential migration of cortical interneurons in Foxc1hith/hith mice. Neural Dev. 2012;7:2. doi: 10.1186/1749-8104-7-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Siegenthaler JA, Ashique AM, Zarbalis K, Patterson KP, Hecht JH, Kane MA, Folias AE, Choe Y, May SR, Kume T, Napoli JL, Peterson AS, Pleasure SJ. Retinoic acid from the meninges regulates cortical neuron generation. Cell. 2009;139:597–609. doi: 10.1016/j.cell.2009.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Zhang J, Smith D, Yamamoto M, Ma L, McCaffery P. The meninges is a source of retinoic acid for the late-developing hindbrain. J Neurosci. 2003;23:7610–7620. doi: 10.1523/JNEUROSCI.23-20-07610.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Choe Y, Siegenthaler JA, Pleasure SJ. A cascade of morphogenic signaling initiated by the meninges controls corpus callosum formation. Neuron. 2012;73:698–712. doi: 10.1016/j.neuron.2011.11.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Montagnani S, Castaldo C, Di Meglio F, Sciorio S, Giordano-Lanza G. Extra cellular matrix features in human meninges. Ital J Anat Embryol. 2000;105:167–177. [PubMed] [Google Scholar]
- 53.Radakovits R, Barros CS, Belvindrah R, Patton B, Müller U. Regulation of radial glial survival by signals from the meninges. J Neurosci. 2009;29:7694–7705. doi: 10.1523/JNEUROSCI.5537-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Ichikawa-Tomikawaa N, Ogawa J, Douet V, Xu Z, Kamikubo Y, Sakurai T, Kohsaka S, Chiba H, Hattori N, Yamada Y, Arikawa-Hirasawa E. Laminin α1 is essential for mouse cerebellar development. Matrix Biol. 2012;31:17–28. doi: 10.1016/j.matbio.2011.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Nagy JI, Patel D, Ochalski PA, Stelmack GL. Connexin30 in rodent, cat and human brain: selective expression in gray matter astrocytes, co-localization with connexin43 at gap junctions and late developmental appearance. Neuroscience. 1999;88:447–468. doi: 10.1016/s0306-4522(98)00191-2. [DOI] [PubMed] [Google Scholar]
- 56.Wiencken-Barger AE, Djukic B, Casper KB, McCarthy KD. A role for Connexin43 during neurodevelopment. Glia. 2007;55:675–686. doi: 10.1002/glia.20484. [DOI] [PubMed] [Google Scholar]
- 57.Beggs HE, Schahin-Reed D, Zang K, Goebbels S, Nave KA, Gorski J, Jones KR, Sretavan D, Reichardt LF. FAK deficiency in cells contributing to the basal lamina results in cortical abnormalities resembling congenital muscular dystrophies. Neuron. 2003;40:501–514. doi: 10.1016/s0896-6273(03)00666-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Choi BH. Role of the basement membrane in neurogenesis and repair of injury in the central nervous system. Microsc Res Tech. 1994;28:193–203. doi: 10.1002/jemt.1070280304. [DOI] [PubMed] [Google Scholar]
- 59.Kerever A, Schnack J, Vellinga D, Ichikawa N, Moon C, Arikawa-Hirasawa E, Efird JT, Mercier F. Novel extracellular matrix structures in the neural stem cell niche capture the neurogenic factor FGF-2 from the extracellular milieu. Stem Cells. 2007;25:2146–2157. doi: 10.1634/stemcells.2007-0082. [DOI] [PubMed] [Google Scholar]
- 60.Mercier F, Hatton GI. Connexin 26 and basic fibroblast growth factor are expressed primarily in the subpial and subependymal layers in adult brain parenchyma: Roles in stem cell proliferation and morphological plasticity? J Comp Neurol. 2001;431:88–104. doi: 10.1002/1096-9861(20010226)431:1<88::aid-cne1057>3.0.co;2-d. [DOI] [PubMed] [Google Scholar]
- 61.Stylianopoulou F, Herbert J, Soares MB, Efstratiadis A. Expression of the insulin-like growth factor II gene in the choroid plexus and the leptomeninges of the adult rat central nervous system. Proc Natl Acad Sci USA. 1988;85:141–145. doi: 10.1073/pnas.85.1.141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Brar AK, Chernausek SD. Localization of insulin-like growth factor binding protein-4 expression in the developing and adult rat brain: Analysis by in situ hybridization. J Neurosci Res. 1993;35:103–114. doi: 10.1002/jnr.490350112. [DOI] [PubMed] [Google Scholar]
- 63.Stumm R, Kolodziej A, Schulz S, Kohtz JD, Höllt V. Patterns of SDF-1a and SDF-1b mRNAs, migration pathways, and phenotypes of CXCR4-expressing neurons in the developing rat telencephalon. J Comp Neurol. 2007;502:382–399. doi: 10.1002/cne.21336. [DOI] [PubMed] [Google Scholar]
- 64.Reiss K, Mentlein R, Sievers J, Hartmann D. Stromal cell-derived factor 1 is secreted by meningeal cells and acts as chemotactic factor on neuronal stem cells of the cerebellar external granular layer. Neuroscience. 2002;115:295–305. doi: 10.1016/s0306-4522(02)00307-x. [DOI] [PubMed] [Google Scholar]
- 65.Berger O, Li G, Han SM, Paredes M, Pleasure SJ. Expression of SDF-1 and CXCR4 during reorganization of the postnatal dentate gyrus. Dev Neurosci. 2007;29:48–58. doi: 10.1159/000096210. [DOI] [PubMed] [Google Scholar]
- 66.Aviezer D, Hecht D, Safran M, Elsinger M, David G, Yayon A. Perlecan, basal lamina proteoglycan, promotes basic fibroblast growth factor-receptor binding, mitogenesis and angiogenesis. Cell. 1994;79:1005–1013. doi: 10.1016/0092-8674(94)90031-0. [DOI] [PubMed] [Google Scholar]
- 67.Arikawa-Hirasawa E, Le AH, Nishino I, Nonaka I, Ho NC, Francomano CA, Govindraj P, Hassell JR, Devaney JM, Spranger J, Stevenson RE, Iannaccone S, Dalakas MC, Yamada Y. Structural and functional mutations of the perlecan gene cause Schwartz-jampel syndrome, with mitotic myopathy and chondrodysplasia. Am J Hum Genet. 2002;70:1368–1375. doi: 10.1086/340390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Day-Lollini PA, Stewart GR, Taylor MJ, Johnson RM, Chellman GJ. Hyperplastic changes within the leptomeninges of the rat and monkey in response to chronic intracerebroventricular infusion of nerve growth factor. Exp Neurol. 1997;145:24–37. doi: 10.1006/exnr.1997.6448. [DOI] [PubMed] [Google Scholar]
- 69.Parr AM, Tator CH. Intrathecal epidermal growth factor and fibroblast growth factor-2 exacerbate meningeal proliferative lesions associated with intrathecal catheters. Neurosurgery. 2007;60:926–933. doi: 10.1227/01.NEU.0000255441.59612.98. [DOI] [PubMed] [Google Scholar]
- 70.Nagy JI, Li X, Rempel J, Stelmack G, Patel D, Staines WA, Yasumura T, Rash JE. Connexin26 in adult rodent central nervous system: demonstration at astrocytic gap junctions and colocalization with connexin30 and connexin43. J Comp Neurol. 2001;441:302–323. doi: 10.1002/cne.1414. [DOI] [PubMed] [Google Scholar]
- 71.Lynn BD, Tress O, May D, Willecke K, Nagy JI. Ablation of connexin30 in transgenic mice alters expression patterns of connexin26 and connexin32 in glial cells and leptomeninges. Eur J Neurosci. 2011;34:1783–1793. doi: 10.1111/j.1460-9568.2011.07900.x. [DOI] [PubMed] [Google Scholar]
- 72.Arishima H, Sato K, Kubota T. Immunohistochemical and ultrastructural study of gap junction proteins connexin26 and 43 in human arachnoid villi and meningeal tumors. J Neuropathol Exp Neurol. 2002;61:1048–1055. doi: 10.1093/jnen/61.12.1048. [DOI] [PubMed] [Google Scholar]
- 73.Popescu BO, Gherghiceanu M, Kostin S, Ceafalan L, Popescu LM. Telocytes in meninges and choroid plexus. Neurosci Lett. 2012;516:265–269. doi: 10.1016/j.neulet.2012.04.006. [DOI] [PubMed] [Google Scholar]
- 74.Decimo I, Bifari F, Rodriguez FJ, Malpeli G, Dolci S, Lavarini V, Pretto S, Vasquez S, Sciancalepore M, Montalbano A, Berton V, Krampera M, Fumagalli G. Nestin- and DCX-positive cells reside in adult spinal cord meninges and participate to injury- induced parenchymal reaction. Stem Cells. 2011;29:2062–2076. doi: 10.1002/stem.766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Nakagomi T, Molnár Z, Nakano-Doi A, Taguchi A, Saino O, Kubo S, Clausen M, Yoshikawa H, Nakagomi N, Matsuyama T. Ischemia-induced neural stem/progenitor cells in the pia mater following cortical infarction. Stem Cells Dev. 2011;20:2037–2051. doi: 10.1089/scd.2011.0279. [DOI] [PubMed] [Google Scholar]
- 76.Nakagomi T, Molnár Z, Taguchi A, Nakano-Doi A, Lu S, Kasahara Y, Nakagomi N, Matsuyama T. Leptomeningeal-derived doublecortin-expressing cells in poststroke brain. Stem Cells Dev. 2012;21:2350–4. doi: 10.1089/scd.2011.0657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Avnur Z, Geiger B. Immunocytochemical localization of native chondroitin-sulfate in tissues and cultured cells using specific monoclonal antibody. Cell. 1984;38:811–822. doi: 10.1016/0092-8674(84)90276-9. [DOI] [PubMed] [Google Scholar]
- 78.Stichel CC, Müller HW. The CNS lesion scar: new vistas on an old regeneration barrier. Cell Tissue Res. 1998;294:1–9. doi: 10.1007/s004410051151. [DOI] [PubMed] [Google Scholar]
- 79.Shearer MC, Niclou SP, Brown D, Asher RA, Holtmaat AJ, Levine JM, Verhaagen J, Fawcett JW. The astrocyte/meningeal cell interface is a barrier to neurite outgrowth which can be overcome by manipulation of inhibitory molecules or axonal signalling pathways. Mol Cell Neurosci. 2003;24:913–925. doi: 10.1016/j.mcn.2003.09.004. [DOI] [PubMed] [Google Scholar]
- 80.Abnet K, Fawcett JW, Dunnett SB. Interactions between meningeal cells and astrocytes in vivo and in vitro. Brain Res Dev Brain Res. 1991;59:187–196. doi: 10.1016/0165-3806(91)90099-5. [DOI] [PubMed] [Google Scholar]
- 81.Bundesen LQ, Scheel TA, Bregman BS, Kromer LF. Ephrin-B2 and EphB2 regulation of astrocyte–meningeal fibroblast interactions in response to spinal cord lesions in adult rats. J Neurosci. 2003;23:7789–7800. doi: 10.1523/JNEUROSCI.23-21-07789.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Ramón y Cajal S. Degeneration and regeneration of the nervous system. Hafner; 1928. pp. 21–53. [Google Scholar]
- 83.Guth L, Reier PJ, Barrett CP, Donati EJ. Repair of the mammalian spinal cord. Trends Neurosci. 1983;6:20–24. [Google Scholar]
- 84.Stichel CC, Müller HW. Relationship between injury-induced astrogliosis, laminin expression and axonal sprouting in the adult rat brain. J Neurocytol. 1994;23:615–630. doi: 10.1007/BF01191556. [DOI] [PubMed] [Google Scholar]
- 85.Blaugrund E, Lavie V, Cohen J, Solomon A, Schreyer DJ, Schwartz M. Axonal regeneration is associated with glial migration: comparison between the injured optic nerves of fish and rats. J Comp Neurol. 1993;330:105–112. doi: 10.1002/cne.903300109. [DOI] [PubMed] [Google Scholar]
- 86.Schnell L, Schwab ME. Sprouting and regeneration of lesioned corticospinal tract fibres in the adult rat spinal cord. Eur J Neurosci. 1993;5:1156–1171. doi: 10.1111/j.1460-9568.1993.tb00970.x. [DOI] [PubMed] [Google Scholar]
- 87.Kimura-Kuroda J, Teng X, Komuta Y, Yoshioka N, Sango K, Kawamura K, Raisman G, Kawano H. An in vitro model of the inhibition of axon growth in the lesion scar formed after central nervous system injury. Mol Cell Neurosci. 2010;43:177–187. doi: 10.1016/j.mcn.2009.10.008. [DOI] [PubMed] [Google Scholar]
- 88.McKeon RJ, Schreiber RC, Rudge JS, Silver J. Reduction of neurite outgrowth in a model of glial scarring following CNS injury is correlated with the expression of inhibitory molecules on reactive astrocytes. J Neurosci. 1991;11:3398–3411. doi: 10.1523/JNEUROSCI.11-11-03398.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Ness R, David S. Leptomeningeal cells modulate the neurite growth promoting properties of astrocytes in vitro. Glia. 1997;19:47–57. doi: 10.1002/(sici)1098-1136(199701)19:1<47::aid-glia5>3.0.co;2-7. [DOI] [PubMed] [Google Scholar]
- 90.Franzen R, Martin D, Daloze A, Moonen G, Schoenen J. Grafts of meningeal fibroblasts in adult rat spinal cord lesion promote axonal regrowth. Neuroreport. 1999;10:1551–1556. doi: 10.1097/00001756-199905140-00029. [DOI] [PubMed] [Google Scholar]
- 91.Zukor KA, Kent DT, Odelberg SJ. Meningeal cells and glia establish a permissive environment for axon regeneration after spinal cord injury in newts. Neural Dev. 2011;6:1. doi: 10.1186/1749-8104-6-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Hayashi H, Morizane A, Koyanagi M, Ono Y, Sasai Y, Hashimoto N, Takahashi J. Meningeal cells induce dopaminergic neurons from embryonic stem cells. Eur J Neurosci. 2008;27:261–268. doi: 10.1111/j.1460-9568.2008.06027.x. [DOI] [PubMed] [Google Scholar]
- 93.Berry M, Maxwell WL, Logan A, Mathewson A, McConnell P. Deposition of scar tissue in the central nervous system. Acta Neurochir Suppl (Wien) 1983;32:31–53. doi: 10.1007/978-3-7091-4147-2_3. [DOI] [PubMed] [Google Scholar]
- 94.Tang X, Davies JE, Davies SJ. Changes in distribution, cell associations, and protein expression levels of NG2, neurocan, phosphacan, brevican, versican V2, and tenascin-C during acute to chronic maturation of spinal cord scar tissue. J Neurosci Res. 2003;71:427–444. doi: 10.1002/jnr.10523. [DOI] [PubMed] [Google Scholar]
- 95.Pasterkamp RJ, Giger RJ, Ruitenberg MJ, Holtmaat AJ, De Wit J. Expression of the gene encoding the chemorepellent semaphorin III is induced in the fibroblast component of neural scar tissue formed following injuries of adult but not neonatal CNS. Mol Cell Neurosci. 1999;13:143–116. doi: 10.1006/mcne.1999.0738. [DOI] [PubMed] [Google Scholar]
- 96.Zhang Y, Anderson PN, Campbell G, Mohajeri H, Schachner M, Lieberman AR. Tenascin-C expression by neurons and glial cells in the rat spinal cord: changes during postnatal development and after dorsal root or sciatic nerve injury. J Neurocytol. 1995;24:585–601. doi: 10.1007/BF01257374. [DOI] [PubMed] [Google Scholar]
- 97.Kähäri VM, Larjava H, Uitto J. Differential regulation of extracellular matrix proteoglycan (PG) gene expression. Transforming growth factor-beta 1 upregulates biglycan (PGI), and versican (large fibroblast PG) but down-regulates decorin (PGII) mRNA levels in human fibroblasts in culture. J Biol Chem. 1991;266:10608–10615. [PubMed] [Google Scholar]
- 98.Komuta Y, Teng X, Yanagisawa H, Sango K, Kawamura K, Kawano H. Expression of transforming growth factor-beta receptors in meningeal fibroblasts of the injured mouse brain. Cell Mol Neurobiol. 2010;30:101–11. doi: 10.1007/s10571-009-9435-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Fallon JR. Preferential outgrowth of central nervous system neurites on astrocytes and Schwann cells as compared with nonglial cells in vitro. J Cell Biol. 1985;100:198–207. doi: 10.1083/jcb.100.1.198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Niclou SP, Franssen EH, Ehlert EM, Taniguchi M, Verhaagen J. Meningeal cell-derived semaphorin 3A inhibits neurite outgrowth. Mol Cell Neurosci. 2003;24:902–912. doi: 10.1016/s1044-7431(03)00243-4. [DOI] [PubMed] [Google Scholar]
- 101.Balabanov R, Dore-Duffy P. Role of the CNS microvascular pericyte in the blood brain barrier. J Neurosci Res. 1998;53:637–664. doi: 10.1002/(SICI)1097-4547(19980915)53:6<637::AID-JNR1>3.0.CO;2-6. [DOI] [PubMed] [Google Scholar]
- 102.Thomas WE. Brain macrophages: on the role of pericytes and perivascular cells. Brain Res Brain Res Rev. 1999;31:42–57. doi: 10.1016/s0165-0173(99)00024-7. [DOI] [PubMed] [Google Scholar]
- 103.Dore-Duffy P, Owen C, Balabanov R, Murphy S, Beaumont T, Rafols JA. Pericyte migration from the vascular wall in response to traumatic brain injury. Microvasc Res. 2000;60:55–69. doi: 10.1006/mvre.2000.2244. [DOI] [PubMed] [Google Scholar]
- 104.Dore-Duffy P, Katychev A, Wang X, Van Buren EJ. CNS microvascular pericytes exhibit multipotential stem cell activity. J Cereb Blood Flow Metab. 2006;26:613–624. doi: 10.1038/sj.jcbfm.9600272. [DOI] [PubMed] [Google Scholar]
- 105.Crisan M, Yap S, Casteilla L, Chen CW, Corselli M, Park TS, Andriolo G, Sun B, Zheng B, Zhang L, Norotte C, Teng PN, Traas J, Schugar R, Deasy BM, Badylak S, Buhring HJ, Giacobino JP, Lazzari L, Huard J, Péault B. A perivascular origin for mesenchymal stem cells in multiple human organs. Cell Stem Cell. 2008;3:301–313. doi: 10.1016/j.stem.2008.07.003. [DOI] [PubMed] [Google Scholar]
- 106.Golding JP, Cohen J. Border controls at the mammalian spinal cord: Late surviving neural crest boundary cap cells at dorsal root entry sites may regulate sensory afferent ingrowth and entry zone morphogenesis. Mol Cell Neurosci. 1997;9:381–396. doi: 10.1006/mcne.1997.0647. [DOI] [PubMed] [Google Scholar]
- 107.Vitalis T, Alvarez C, Chen K, Shih JC, Gaspar P, Cases O. Developmental expression pattern of monoamine oxidases in sensory organs and neural crest derivatives. J Comp Neurol. 2003;464:392–403. doi: 10.1002/cne.10804. [DOI] [PubMed] [Google Scholar]
- 108.Coulpier F, Le Crom S, Maro GS, Manent J, Giovannini M, Maciorowski Z, Fischer A, Gessler M, Charnay P, Topilko P. Novel features of boundary cap cells revealed by the analysis of newly identified molecular markers. Glia. 2009;57:1450–1457. doi: 10.1002/glia.20862. [DOI] [PubMed] [Google Scholar]
- 109.Wilkinson DG, Bhatt S, Chavrier P, Bravo R, Charnay P. Segment-specific expression of a zinc-finger gene in the developing nervous system of the mouse. Nature. 1989;337:461–464. doi: 10.1038/337461a0. [DOI] [PubMed] [Google Scholar]
- 110.Topilko P, Schneider-Maunoury S, Levi G, Baron-Van Evercooren A, Chennoufi AB, Seitanidou T, Babinet C, Charnay P. Krox-20 controls myelination in the peripheral nervous system. Nature. 1994;371:796–799. doi: 10.1038/371796a0. [DOI] [PubMed] [Google Scholar]
- 111.Murphy P, Topilko P, Schneider-Maunoury S, Seitanidou T, Baron-Van Evercooren A, Charnay P. The regulation of Krox-20 expression reveals important steps in the control of peripheral glial development. Development. 1996;122:2847–2857. doi: 10.1242/dev.122.9.2847. [DOI] [PubMed] [Google Scholar]
- 112.Maro GS, Vermeren M, Voiculescu O, Melton L, Cohen J, Charnay P, Topilko P. Neural crest boundary cap cells constitute a source of neuronal and glial cells of the PNS. Nat Neurosci. 2004;7:930–938. doi: 10.1038/nn1299. [DOI] [PubMed] [Google Scholar]
- 113.Zujovic V, Thibaud J, Bachelin C, Vidal M, Deboux C, Coulpier F, Stadler N, Charnay P, Topilko P, Baron-Van Evercooren A. Boundary cap cells are peripheral nervous system stem cells that can be redirected into central nervous system lineages. Proc Natl Acad Sci USA. 2011;108:10714–10719. doi: 10.1073/pnas.1018687108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Herling-Leffler J, Marmigère F, Heglind M, Cederberg A, Koltzenburg M, Enerback S, Ernfors P. The boundary cap: A source of neural crest stem cells that generate multiple sensory neuron subtypes. Development. 2005;132:2623–2632. doi: 10.1242/dev.01852. [DOI] [PubMed] [Google Scholar]
- 115.Aquino JB, Hjerling-Leffler J, Koltzenburg M, Edlund T, Villar MJ, Ernfors P. In vitro and in vivo differentiation of boundary cap neural crest stem cells into mature Schwann cells. Exp Neurol. 2006;198:438–449. doi: 10.1016/j.expneurol.2005.12.015. [DOI] [PubMed] [Google Scholar]
- 116.Zujovic V, Thibaud J, Bachelin C, Vidal M, Deboux C, Coulpier F, Stadler N, Charnay P, Topilko P, Baron-Van Evercooren A. Boundary cap cells are peripheral nervous system stem cells that can be redirected into central nervous system lineages. Proc Natl Acad Sci USA. 2011;108:10714–10719. doi: 10.1073/pnas.1018687108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Zujovic V, Thibaud J, Bachelin C, Vidal M, Coulpier F, Charnay P, Topilko P, Baron-Van Evercooren A. Boundary cap cells are highly competitive for CNS remyelination: fast migration and efficient differentiation in PNS and CNS myelin-forming cells. Stem Cells. 2010;28:470–479. doi: 10.1002/stem.290. [DOI] [PubMed] [Google Scholar]
- 118.Lendahl U, Zimmerman LB, McKay RD. CNS stem cells express a new class of intermediate filament protein. Cell. 1990;60:585–595. doi: 10.1016/0092-8674(90)90662-x. [DOI] [PubMed] [Google Scholar]
- 119.Petricevic J, Forempoher G, Ostojic L, Mardesic-Brakus S, Andjelinovic S, Vukojevic K, Saraga-Babic M. Expression of nestin, mesothelin and epithelial membrane antigen (EMA) in developing and adult human meninges and meningiomas. Acta Histochem. 2011;113:703–711. doi: 10.1016/j.acthis.2010.09.005. [DOI] [PubMed] [Google Scholar]
- 120.Itokazu Y, Kitada M, Dezawa M, Mizoguchi A, Matsumoto N, Shimizu A, Ide C. Choroid plexus ependymal cells host neural progenitor cells in the rat. Glia. 2006;53:32–42. doi: 10.1002/glia.20255. [DOI] [PubMed] [Google Scholar]
- 121.DeGiorgio LA, Sheu KF, Blass JP. Culture from human leptomeninges of cells containing neurofilament protein and neuron-specific enolase. J Neurol Sci. 1994;124:141–148. doi: 10.1016/0022-510x(94)90319-0. [DOI] [PubMed] [Google Scholar]
- 122.DeGiorgio LA, Bernstein JJ, Blass JP. Implantation of cultured human leptomeningeal cells into rat brain. Int J Dev Neurosci. 1997;15:231–238. doi: 10.1016/s0736-5748(96)00076-7. [DOI] [PubMed] [Google Scholar]
- 123.Bernstein JJ, Karp SM, Goldberg WJ, DeGiorgio LA, Blass JP. Human leptomeningeal-derived cells express GFAP and HLADR when grafted into rat spinal cord. Int J Dev Neurosci. 1996;14:681–687. doi: 10.1016/0736-5748(95)00096-8. [DOI] [PubMed] [Google Scholar]
- 124.Moreno-Manzano V, Rodriguez-Jimenez FJ, Garcia-Rosello M, Lainez S, Erceq S, Calvo MT, Ronaghi M, Lloret M, Planells-Cases R, Sanchez-Puelles JM, Stojkovic M. Activated spinal cord ependymal stem cells rescue neurological function. Stem Cells. 2009;27:733–743. doi: 10.1002/stem.24. [DOI] [PubMed] [Google Scholar]
- 125.Arvidsson A, Collin T, Kirik D, Kokaia Z, Lindvall O. Neuronal replacement from en- dogenous precursors in the adult brain after stroke. Nat Med. 2002;8:963–970. doi: 10.1038/nm747. [DOI] [PubMed] [Google Scholar]
- 126.Nakatomi H, Kuriu T, Okabe S, Yamamoto S, Hatano S, Kawahara N, Tamura A, Kirino T, Nakafuku M. Regeneration of hippocampal pyramidal neurons after ischemic brain injury by recruitment of endogenous neural progenitors. Cell. 2002;110:429–441. doi: 10.1016/s0092-8674(02)00862-0. [DOI] [PubMed] [Google Scholar]
- 127.Gan L, Qiao S, Lan X, Chi L, Luo C, Lien L, Yan Liu Q, Liu R. Neurogenic responses to amyloid-beta plaques in the brain of Alzheimer’s disease-like transgenic (pPDGF-APPSw, Ind) mice. Neurobiol Dis. 2008;29:71–80. doi: 10.1016/j.nbd.2007.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Meletis K, Barnabe-Heider F, Carlen M, Evergren E, Tomilin N, Shupliakov O, Frisén J. Spinal cord injury reveals multilineage differentiation of ependymal cells. PLoS Biol. 2008;6:e182. doi: 10.1371/journal.pbio.0060182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Komitova M, Perfilieva E, Mattsson B, Eriksson PS, Johansson BB. Effects of cortical ische- mia and postischemic environmental enrichment on hippocampal cell genesis and differentiation in the adult rat. J Cereb Blood Flow Metab. 2002;22:852–860. doi: 10.1097/00004647-200207000-00010. [DOI] [PubMed] [Google Scholar]
- 130.Fallon J, Reid S, Kinyamu R, Opole I, Opole R, Baratta J, Korc M, Endo TL, Duong A, Nguyen G, Karkehabadhi M, Twardik D, Patel S, Loughlin S. In vivo induction of massive prolifer- ation, directed migration, and differentiation of neural cells in the adult mammalian brain. Proc Natl Acad Sci USA. 2000;97:14686–14691. doi: 10.1073/pnas.97.26.14686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Doetsch F, Garcia-Verdugo JM, Alvarez-Buylla A. Cellular composition and three-dimensional organization of the subventricular geminal zone in the adult mammalian brain. J Neurosci. 1997;17:5046–5061. doi: 10.1523/JNEUROSCI.17-13-05046.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]