

Abstract
Introduction
To date, isolated cell based blood-brain barrier (BBB) models have been widely used for brain drug delivery and targeting, due to their relatively proper bioelectrical and permeability properties. However, primary cultures of brain capillary endothelial cells (BCECs) isolated from different species vary in terms of bioelectrical and permeability properties.
Methods
To pursue this, in the current investigation, primary porcine and bovine BCECs (PBCECs and BBCECs, respectively) were isolated and used as an in vitro BBB model. The bioelectrical and permeability properties were assessed in BCECs co-cultured with C6 cells with/without hydrocortisone (550 nM). The bioelectrical properties were further validated by means of the permeability coefficients of transcellular and paracellular markers.
Results
The primary PBCECs displayed significantly higher trans-endothelial electrical resistance (~900 Ω.cm2) than BBCECs (~700 Ω.cm2) - both cocultured with C6 cells in presence of hydrocortisone. Permeability coefficients of propranolol/diazepam and mannitol/sucrose in PBCECs were ~21 and ~2 (×10-6 cm.sec-1), where these values for BBCECs were ~25 and ~5 (×10-6 cm.sec-1).
Conclusion
Upon our bioelectrical and permeability findings, both models display discriminative barrier functionality but porcine BCECs seem to provide a better platform than bovine BCECs for drug screening and brain targeting.
Keywords: Blood-brain barrier, Bioelectrical properties, Cell culture model, Brain targeting, Cell and tissue engineering
Introduction
Brain capillary endothelial cells provide restrictive barrier functionality to control CNS inward and outward transport of endogenous and exogenous molecules. On account of the barrier functions, enzymatic activity and functional presence of transporters and receptors, and studies on the permeation of pharmaceuticals across BBB represent a major challenge in neuropharmaceutical researches. In fact, specific/selective transportation of compounds/drugs through the biological membranes and barriers (e.g., BBB, blood ocular barriers) clearly emphasize importance of cellular transport machineries of such barriers (Barar et al. 2008; Barar et al. 2010; de Boer and Gaillard 2007; Omidi and Gumbleton 2005).
A number of cell-based in vitro BBB models have been so far exploited for brain drug screening, delivery, and targeting. Of these cell-based models, however, no immortalized cell line has been shown to represent a comprehensive model in terms of barrier restrictiveness and transport functions as seen in vivo (Gumbleton and Audus 2001). Ideally, a cell-based BBB model should represent discriminative barrier functionality as well as transport machineries. Since no immortalized cell line possesses high enough bioelectrical resistance and discriminative permeability coefficient ratio (i.e., for transcellular and paracellular markers), primary BCECs isolated from various species in particular bovine (Audus and Borchardt 1987) and porcine (Franke et al. 2000) have been exploited as in vitro BBB models. For example, Hurst and Fritz (1996) presented a co-culture BBB model of the immortalized human umbilical vein endothelial cells ECV304 with the rat C6 glioma cells with high transendotheila electrical resistance (TEER) about 400-600 Ω.cm 2 (Hurst and Fritz 1996). However, using DNA figure printing, ECV304 cells were later reassigned as the T24 bladder epithelial carcinoma cell (Kiessling et al. 1999; Suda et al. 2001).
We have also reported b.End3 cells as an appropriate in vitro BBB model for carrier-mediated transport studies, but not for drug screening due to its lower barrier discrimination (Omidi et al. 2003). Hence, to achieve a highly discriminative cell-based BBB model, we have later isolated primary BCECs from porcine brain and co-cultured with astrocytes (Smith et al. 2007; Omidi et al. 2003). This model showed TEER value almost as high as 1000 Ω.cm 2 . Similarly, Audus and Borchardt (1987) reported bovine brain microvessel endothelial cell monolayers as an in vitro model of BBB and later on Helms et al. (2010) reported an in vitro model of BBB established from bovine BCECs co-cultured with rat astrocytes showing TEER values from 250-750 (Ω.cm 2 ), which were significantly increased using cAMP elevators, dexamethasone and pH control by addition of HEPES, MOPS, or TES (Helms et al. 2010). It seems there is an inconsistency regarding the bioelectrical properties and permeability coefficients of two main isolated primary cultures of BCECs. Thus in the current study, we compare primary cultures of BCECs isolated from bovine and porcine.
Materials and methods
Materials
The following items were obtained from Sigma-Aldrich Chemical Co. (Poole, UK): glutaraldehyde, hydrocortisone and alkaline phosphatase (ALP). The M199 medium, DMEM/F12 medium, foetal bovine serum (FBS), heat-inactivated-FBS, penicillin G and streptomycin were obtained from InVitrogen (Paisley, UK). Percoll and the radioisotopes DL-[4- 3 H] propranolol hydrochloride and [U- 14 14C] sucrose were obtained from Amersham Life Science (Little Chalfont, UK). Tissue culture treated multi-well plates and Transwell-clearTM polyester membrane (pore size 0.4mm) inserts (diameter 6.5 and 24 mm) were obtained from Corning Costar (High Wycombe, UK). Neutrally-buffered 2% osmium tetroxide in veronal acetate buffer and Araldite (CY212) resin were obtained from TAAB (Aldermaston, UK). OptiPhase HiSafe3™ liquid scintillation fluid was obtained from Fisher Scientific Chemicals (Loughborough, UK). Dispase II, dispase/collagenase (from Vibrio alginolyticus/Bacillus polymyxa), and rat-tail collagen type I were obtained from Roche (East Sussex, UK). The rat glioma cell line, C6, was obtained from ECACC (Porton, UK).
Isolation of brain capillary endothelial cells
Isolations of BCECs from bovine and porcine were performed according to the previously reported methodologies (Audus and Borchardt 1987; Franke et al. 2000; Omidi et al. 2003; Smith et al. 2007) with slight modifications. Briefly, the bovine/porcine heads were obtained from slaughterhouse and transported to the lab within 0.5 h. Brains were carefully taken out and washed by ice-cold sterile phosphate buffered saline (PBS). Meninges and large surface vessels were removed from the cerebral cortex of the brains under sterile condition. The cortical grey matter of porcine brains minced to small pieces (<1 mm 3 03) and suspended in preparation media (i.e., M199 supplemented with 100 units/ml of penicillin G and 100 µg/ml of streptomycin) using 50 ml per brain. The preparation media was supplemented with dispase II at a final concentration of 0.5%. The mixture was incubated at 37oC for 2 h using Orbital Shaker SO5, Bibby Stenilin Ltd., (Staffordshire, UK). Then, 150 ml of 30% percoll solution (r=1.032 g/ml) was added per 100 ml of the digested suspension and the mixture centrifuged at 6000×g for 10 min at 4oC using a swing-out centrifuge, Avanti TM J-25 centrifuge, Beckman Coulter Inc., (California, USA). The pellet containing microvessels were resuspended in the culture media (i.e., preparation media supplemented with 10% heat-inactivated FBS) to a total volume of 10 ml per brain. Then, 1 mg/ml collagenase-dispase II (0.1% w/v) was added to the latter mixture and incubated at 37oC for 1 h. The suspension was kept at 4oC for 15-20 min to stop enzymatic digestion and passed through the 180 µm nylon mesh to obtain the detached cells. The resulted cells from 1 brain were suspended in 5 ml of culture media and centrifuged at 1300×g for 10 min at 4°C on a discontinuous percoll-gradient (heavy percoll, 1.07 g/cm, and light percoll, 1.03 g/cm 3 ). The endothelial cells, mainly present in cell clusters, were removed with a pipette from the middle band at a density between 1.052 and 1.05 g/cm 3 . The obtained cells were either used as fresh or frozen for later use.
Coating procedure with rat tail collagen type I
Rat tail collagen type I (RTCI) was prepared with sterile acetic acid 0.2% v/v. For coating, 25 and 75 cm 2 T-flasks were respectively introduced with 250 and 1000 (μl) of RTCI (0.2% v/v), i.e. at a final concentration of 3 mg of RTCI per cm 2 . The coated flasks were allowed to dry over night (16-18 hr) under laminar flow. For coating of the small (0.3318 cm 2 ) and large (4.7 cm 2 ) transwell membrane inserts (either polyester or polycarbonate inserts), they were covered with 50 and 500 (µl per cm 2 ) of RTCI (0.2% v/v), respectively at a final concentration of 5 mg per cm 2 02. The inserts were first kept at 37oC for 3 hr, and then let to dry under laminar flow at room temperature over night. To remove the acetic acid residual, the RTCI coated inserts were subjected to further washing (×3) using PBS before plating the cells.
Cell culture
For cultivation of BCECs, the isolated cells (at a seeding density of 5.0 ´ 10 4 cells per cm 2 ) were plated onto a T-flask or inserts precoated with RTCI. Once the freshly isolated PBCE cells were harvested, they were cultured as the primary alone or as cocultured with C6 glioma cells in presence or absence of hydrocortisone (550 nM). For cultivation as astrocytes coculture (ACC), BCECs were cultured onto the RTCI coated T-flasks or inserts while C6 cells were cultured on the basal of the plates. The cells were allowed to grow for 24-48 hr, then washed with PBS (37ºC) to remove cell debris and replenished with fresh culture media.
To generate tighter monolayers with high TEER, the culture medium was ‘switched’ to assay medium (i.e., DMEM/Ham's F12, 100 units/ml penicillin G, 100 mg/ml streptomycin supplemented with 550 nM hydrocortisone) without serum at 72 hr post-seeding according to previously reported method by Franke et al. (2000). At 24 hr after this ‘switch’ procedure, transport screening were undertaken.
Rat glioma C6 cells were cultured onto the 6- or 24-well plates at a seeding density of 5.0´10 4 cells/cm 2 using M199 medium supplemented with 2mM glutamine, heat inactivated FBS 10%, antibiotics penicillin G (100 units/ml) and streptomycin sulfate (100 µg/ml). For BCECs cocultures with astrocytic, C6 cells were cultivated 3 days prior to seeding BCECs to maximize the biosynthetic secretion of astrocyte derived factors (Rubin et al. 1991).
All cultured cells were maintained in a humidified atmosphere (5% CO2 / 95% air) and medium was replenished every 24 hr.
Light and electron microscopy
The morphology of monolayers of BCECs cultivated onto the cell culture flasks and Transwellä membranes (0.4 µm pore size) in the absence or presence of C6 coculture were examined by the light microscopy (LM) as well as the transmission electron microscopy (TEM). For TEM, cells were fixed and embedded in accordance with the methodology reported previously (Newman et al. 1999).
Trans-endothelial electrical resistance assessments
For bioelectrical properties, monolayers of porcine and bovine BCECs were assessed for formation of restrictive paracellular barrier. Trans-endothelial electrical resistance (TEER) was measured from day 3 to day 8 using EvomTM epithelial volt-ohm-meter, World Precision Instrument (Sarasota, USA).
Permeability coefficients of transcellular and paracellular markers
Monolayers of porcine and bovine BCECs cultivated on RTCI coated inserts were used for permeability experiments at day 5 or 6 post seeding, i.e., 24-48 hr after "switch" that cells showed the maximum TEER values. The transport of paracellular markers (0.5 µCi/well of [ 14 C] sucrose or [ 14 C] mannitol) and transcellular markers (0.5 µCi/well of [ 3 H] propranolol or [ 3 H] diazepam) were examined to assess the tight junction integrity and restrictiveness of monolayers. Sampling was performed at designated times (15, 30, 45, 60, 75 and 90 min) by removing 100 µl medium from either the apical or basal compartments (for experiment in either the A to B or B to A direction) and replenishing with the same amount of fresh medium. Analysis of radiolabeled probes was carried out using a liquid scintillation counter, Wallac 1409, PerkinElmer Life Sciences (Boston, USA). The apparent permeability coefficients were calculated according to our previously reported study (Omidi et al. 2003), using equation 1 (eq. 1):
J=-P×A×(C2-C1) (eq. 1)
Where, J is flux, P is the permeability of the BCECs for markers, A is the surface area over which diffusion is taking place, and C2-C1 is the difference in concentration of the marker across the membrane for the direction of flow.
Statistical analyses
All experiments were replicated and data represented as mean value of 4-6 replications ± standard deviation (S.D.). Statistical analyses were performed using either ANOVA followed with a multiple comparison test or the two-tailed Student T-test. A P- value less than 0.05 was assumed to present significance.
Results
Morphological examinations
For morphological characterization of BCECs, we looked at LM and TEM micrographs of the cells cultured onto the cell culture flasks and clear Transwell™ polycarbonate inserts. Fig. 1 shows LM images of BCECs at day 6 (panel A), which clearly demonstrates attenuated and elongated spindle form architecture. Both models displayed generation of tight junctions. Typical tight junction is displayed as higher protein density regions (shown with arrows) between two porcine BCECs coculture with astrocytes (Fig. 1B).
Fig. 1.
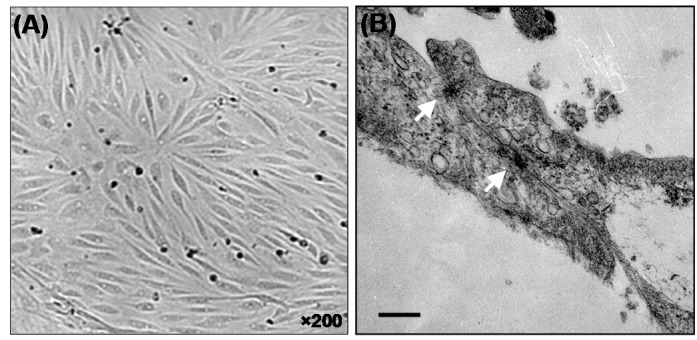
Light microscopy (A) and transmission electron micrographs (B) of the porcine brain capillary endothelial (PBCE) cells. Squamous morphology of the confluent cells (A) and two elongated primary PBCE cells cultured onto the Transwell™ polycarbonate membranes display typical morphology of the endothelial cells. The white arrows (B) represent the tight junctional elements between two flattened primary PBCE cells. Bar equals to 200 nm.
Bioelectrical properties
Figs. 2, 3 and 4 represent TEER values of both porcine and bovine BCECs at 100% confluency treated with HC alone, ACC and HC plus ACC, respectively. Cultured BCECs alone did not display a significant (p>0.05) induction in TEER values (not higher than 200 Ω.cm 2 ). The BCE cells (both porcine and bovine) treated with HC (550 nM) resulted in significant (p<0.05) increased in TEER values up to 501±62 Ω.cm 2 and 487±40 Ω.cm 2 , respectively. Similarly, PBCECs and BBCECs cultivated as ACC showed TEER values up to 546±59 Ω.cm 2 and 507±48 Ω.cm 2 , respectively. The highest TEER values were seen for PBCECs and BBCECs cultivated as ACC in presence of HC (550 nM), i.e. 893±89 Ω.cm 2 and 689±53 Ω.cm 2 , respectively.
Fig. 2.

Trans endothelial electrical resistance (TEER) properties of porcine and bovine brain capillary endothelial cells (BCECs) treated with 550 nM hydrocortisone (HC).
Fig. 3.

Trans endothelial electrical resistance (TEER) properties of porcine and bovine brain capillary endothelial cells (BCECs) as astrocyte coculture (ACC).
Fig. 4.

Trans endothelial electrical resistance (TEER) properties of porcine and bovine brain capillary endothelial cells (BCECs) as astrocyte coculture (ACC) and treated with 550 nM hydrocortisone (HC).
Permeability analyses
The permeability coefficients of transcellular and paracellular markers were measured in both porcine and bovine BCECs monolayers as shown in Table 1.
Table 1. Permeability coefficients of transcellular markers and paracellular markers in porcine and bovine brain capillary endothelial cells.
| Cell based model | Permeability coefficient (×10- 6 cm.sec- 1 ) | |||
| Porcine | Bovine | |||
| Propranolol | Mannitol | Propranolol | Mannitol | |
| BCECs + HC (550 nM) | 23.38 | 7.68 | 24.15 | 9.03 |
| BCECs + ACC | 24.23 | 5.93 | 27.41 | 8.26 |
| BCECs + ACC and HC (550 nM) | 21.47 | 2.31 | 25.32 | 5.18 |
Consistent with the bioelectrical properties, the permeability coefficients ratios of transcellular marker (propranolol) over paracellular marker (mannitol) in PBCECs and BBCECs both cultivated as ACC in presence of HC (550 nM) yielded approximately 9.3 and 4.9, respectively. The apical to basal (A to B) and basal to apical (B to A) permeability coefficients of transcellular marker (diazepam) and paracellular marker (sucrose) for porcine and bovine BCECs cultivated as ACC in the presence of hydrocortisone (550nM) are represented in Figs. 5 and 6, respectively.
Fig. 5.
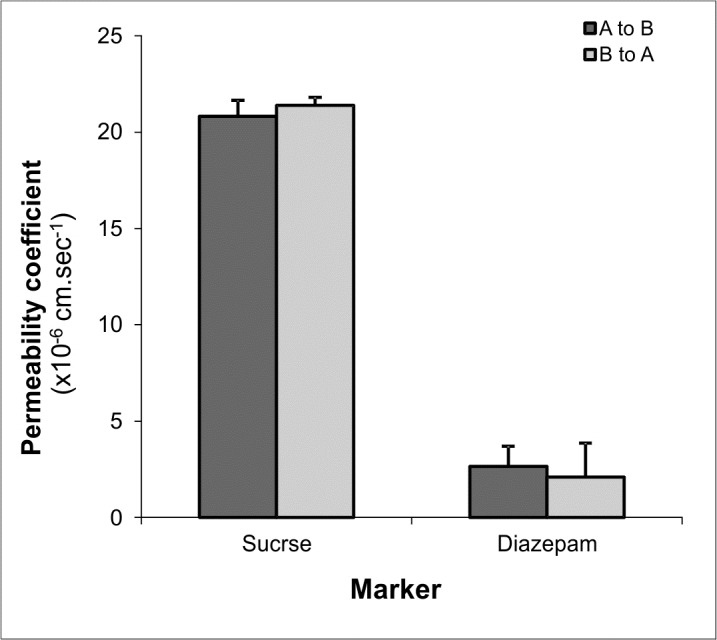
Permeability coefficients of transcellular and paracellular markers in porcine brain capillary endothelial (PBCE) cells in apical to basal (A to B) and basal to apical (B to A) directions. Data represents mean ± S.D. of four replications.
Fig. 6 .
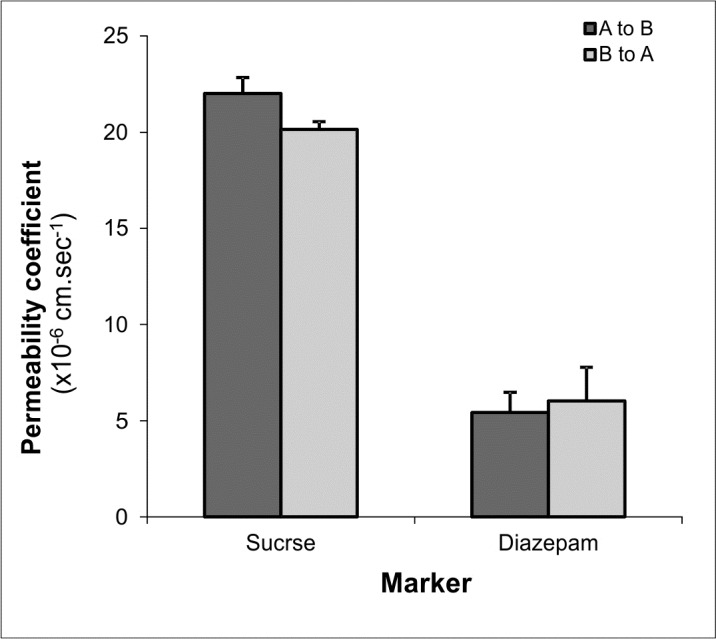
Permeability coefficients of transcellular and paracellular markers in bovine brain capillary endothelial (PBCE) cells in apical to basal (A to B) and basal to apical (B to A) directions. Data represents mean ± S.D. of 4 replications.
Discussion
Brain drug targeting appears to remain as one of the greatest challenging field of researches due to selective functionality of the blood-brain barrier. For drug screening across BBB, it is often essential to use a reliable in vitro BBB model with high enough barrier restrictiveness. Accordingly such model should possess high electrical resistance showing significantly high discrimination between transcellular and paracellular markers. Since no immortalized cell line grants such barrier characteristics (Barar et al. 2010; Gumbleton and Audus 2001), primary cultures of different species (Siakotos 1974) with ability to generate a restrictive barrier (Tsuji et al. 1992) have been used even though isolation of the primary cultures is often associated with some technical complexity. To obtain a high throughput BBB in vitro model, isolated primary BCECs have already been used (Audus and Borchardt 1987; Franke et al. 2000; Hoheisel et al. 1998; Tilling et al. 1998; Jeliazkova-Mecheva and Bobilya 2003; Lacombe et al. 2011; Nakagawa et al. 2009); however, it seems there is a controversy regarding the barrier restrictiveness of primary cultures isolated from different species. To pursue this issue, we have looked at the bioelectrical and permeability properties of porcine and bovine BCECs. It has been shown that astrocytes to be the major modulator of the BBB and regulate the permeability, enzymatic and transport functionalities of the BBB (Allt and Lawrenson 2000; Kuo and Lu 2011). Further, glucocorticoids such as hydrocortisone can modulate the barrier function of BCECs via induction of key molecules of tight junction, i.e. claudin-5 (Kashiwamura et al. 2011). Thus, we used these two modulation systems to improve the barrier functions of the PBCECs and BBCECs.
We witnessed that both BCECs displayed a spindle shape morphology 24 hr post-seeding and formed a homogenous monolayer at day 6 (Fig. 1A). The TEM micrographs displayed high dense interaction between two elongated PBCECs indicating formation of the tight junctions between two BCECs (Fig. 1B) cultivated as ACC in the presence of hydrocortisone (550 nM). It should be prompted that this finding is in agreement with previous results for BBB permeability properties in porcine and murine (Weidenfeller et al. 2005; Kashiwamura et al. 2011; Franke et al. 2000; Hoheisel et al. 1998).
The bioelectrical examinations resulted in significant responsiveness of the primary BCECs cells to ACC and hydrocortisone. Previously, we reported that the primary porcine BCECs cocultured with astrocytes and treated with HC can yield TEER values up to 900 Ω.cm 2 (Smith et al. 2007), which has been reproduced in this study too. The bovine BCECs cocultured with astrocytes and treated with HC also yielded high TEER value (up to ~700 Ω.cm 2 ), but these cells were not as responsive as the porcine BCECs. These differences in TEER values were also reflected in permeability coefficients of transcellular and paracellular markers (Table 1). Permeability coefficients of propranolol/diazepam and mannitol/sucrose in PBCECs were ~21 and ~2 (×10- 6 cm.sec- 1 ), where these values for BBCECs were ~25 and ~5 (×10- 6 cm.sec- 1 ). Overall, the permeability coefficients ratios of transcellular marker over paracellular marker in PBCECs and BBCECs (both cultivated as ACC in presence of 550 nM hydrocortisone) yielded approximately 9 and 5, respectively. Such difference between these two BCECs should be further investigated using molecular biology approach to find out why BCECs isolated from porcine display significantly higher responsiveness to those isolated from bovine. Intriguingly, similar barrier restrictiveness has been previously reported for PBCECs showing sucrose permeability values of 1×10- 6 cm.sec- 1 (Franke et al. 1999). It appears that the astrocytes impose greater impacts in terms of BCECs modulation. For example, Hamm et al. (2004) reported that the permeability of small tracers across BCE cell monolayers was increased upon removal of astrocytes from the coculture. Such changes in permeability were not in association with any detectable alteration in the molecular composition of the tight junction (Hamm et al. 2004). It should be also stated that some researchers have previously reported modulatory effects of some other compounds such as cAMP elevators (Rubin et al. 1991; Ishizaki et al. 2003; Wolburg et al. 1994).
Taken all these together, based on our findings, it can be proposed that PBCECs cocultured with astrocytes in the presence of hydrocortisone display better barrier restrictiveness and thus this model may be considered as an appropriate high-throughput in vitro model for drug screening experiments.
Ethical Issues
None to be declared.
Conflict of interests
No conflict of interest to be declared.
Acknowledgments
The technical supports and advice kindly provided by Dr Mark Gumbleton (Welsh School of Pharmacy, Cardiff University, Cardiff, UK) is highly appreciated. This work financially supported by Tabriz University of Medical Sciences.
References
- Barar J, Javadzadeh AR and Omidi Y . 2008 Ocular novel drug delivery: impacts of membranes and barriers. Expert Opin Drug Deliv, 5(5), 567-581 [DOI] [PubMed] [Google Scholar]
- Barar J, Gumbleton M, Asadi M and Omidi Y . 2010 Barrier functionality and transport machineries of human ECV304 cells. Med Sci Monit, 16(1), BR52-BR60 [PubMed] [Google Scholar]
- de Boer and Gaillard PJ . 2007 Drug Targeting to the Brain. Annu Rev Pharmacol Toxicol, 47, 323-355 [DOI] [PubMed] [Google Scholar]
- Omidi Y and Gumbleton M. 2005. Biological membranes and barriers, In: Biomaterials for delivery and targeting of proteins nucleic acids, Mahato RI, Ed. CRC Press, New York, pp 232-274.
- Gumbleton M and Audus KL . 2001 Progress and limitations in the use of in vitro cell cultures to serve as a permeability screen for the blood-brain barrier. J Pharm Sci, 90(11), 1681-1698 [DOI] [PubMed] [Google Scholar]
- Audus KL and Borchardt RT . 1987 Bovine brain microvessel endothelial cell monolayers as a model system for the blood-brain barrier. Ann N Y Acad Sci, 507(1), 9-18 [DOI] [PubMed] [Google Scholar]
- Franke H, Galla H and Beuckmann CT . 2000 Primary cultures of brain microvessel endothelial cells: a calid and flexible model to study drug transport through the blood-brain barrier in vitro. Brain Res Brain Res Protoc, 5(3), 248-256 [DOI] [PubMed] [Google Scholar]
- Hurst RD and Fritz IB . 1996 Properties of an immortalised vascular endothelial/glioma cell co- culture model of the blood-brain barrier. J Cell Physiol, 167(1), 81-88 [DOI] [PubMed] [Google Scholar]
- Kiessling F, Kartenbeck J and Haller C . 1999 Cell-Cell Contacts in the Human Cell Line ECV304 Exhibit Both Endothelial and Epithelial Characteristics. Cell Tissue Res, 297(1), 131-140 [DOI] [PubMed] [Google Scholar]
- Suda K, Rothen-Rutishauser B, Gunthert M and Wunderli-Allenspach H . 2001 Phenotypic characterization of human umbilical vein endothelial (ECV304) and urinary carcinoma (T24) cells: endothelial versus epithelial features. In Vitro Cell Dev Biol Anim, 37(8), 505-514 [DOI] [PubMed] [Google Scholar]
- Omidi Y, Campbell L, Barar J, Connell D, Akhtar S and Gumbleton M . 2003 Evaluation of the immortalised mouse brain capillary endothelial cell line, B .End3, as an in vitro blood-brain barrier model for drug uptake and transport studies. Brain Res, 990(1-2), 95-112 [DOI] [PubMed] [Google Scholar]
- Smith M, Omidi Y and Gumbleton M . 2007 Primary porcine brain microvascular endothelial cells: biochemical and functional characterisation as a model for drug transport and targeting. J Drug Target, 15(4), 253-268 [DOI] [PubMed] [Google Scholar]
- Helms HC, Waagepetersen HS, Nielsen CU and Brodin B . 2010 Paracellular tightness and claudin-5 expression is increased in the BCEC/Astrocyte blood-brain barrier model by increasing media buffer capacity during growth. AAPS J, 12(4), 759-770 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith M, Omidi Y and Gumbleton M . 2007 Primary porcine brain microvascular endothelial cells: biochemical and functional characterisation as a model for drug transport and targeting. J Drug Target, 15(4), 253-268 [DOI] [PubMed] [Google Scholar]
- Rubin LL, Hall DE, Porter S, Barbu K, Cannon C, Horner HC, et al. 1991 A cell culture model of the blood-brain barrier. J Cell Biol, 115(6), 1725-1735 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Newman GR, Campbell L, von Ruhland C, Jasani B and Gumbleton M . 1999 Caveolin and its cellular and subcellular immunolocalisation in lung alveolar epithelium: implications for alveolar epithelial type I cell function. Cell Tissue Res, 295(1), 111-120 [DOI] [PubMed] [Google Scholar]
- Siakotos AN . 1974 The isolation of endothelial cells from normal human and bovine brain. Methods Enzymol, 32(Part B), 717-722 [DOI] [PubMed] [Google Scholar]
- Tsuji A, Terasaki T, Takabatake Y, Tenda Y, Tamai I, Yamashima T, et al. 1992 P-glycoprotein as the drug efflux pump in primary cultured bovine brain capillary endothelial cells. Life Sci, 51(18), 1427-1437 [DOI] [PubMed] [Google Scholar]
- Hoheisel D, Nitz T, Franke H, Wegener J, Hakvoort A, Tilling T, et al. 1998 Hydrocortisone reinforces the blood-brain barrier properties in a serum free cell culture system [corrected and republished in Biochem Biophys Res Commun 1998 Jun 18;247(2):312-5]. Biochem Biophys Res Commun, 244(1), 312-316 [DOI] [PubMed] [Google Scholar]
- Tilling T, Korte D, Hoheisel D and Galla HJ . 1998 Basement membrane proteins influence brain capillary endothelial barrier function in vitro. J Neurochem, 71(3), 1151-1157 [DOI] [PubMed] [Google Scholar]
- Jeliazkova-Mecheva VV and Bobilya DJ . 2003 A Porcine Astrocyte/Endothelial Cell Co-Culture Model of the Blood-Brain Barrier. Brain Res Brain Res Protoc, 12(2), 91-8 [DOI] [PubMed] [Google Scholar]
- Lacombe O, Videau O, Chevillon D, Guyot AC, Contreras C, Blondel S, et al. 2011 In vitro primary human and animal cell-based blood-brain barrier models as a screening tool in drug discovery. Mol Pharm, 8(3), 651-663 [DOI] [PubMed] [Google Scholar]
- Nakagawa S, Deli MA, Kawaguchi H, Shimizudani T, Shimono T, Kittel A, et al. 2009 A new blood-brain barrier model using primary rat brain endothelial cells, pericytes and astrocytes. Neurochem Int, 54(3-4), 253-263 [DOI] [PubMed] [Google Scholar]
- Allt G and Lawrenson JG . 2000 The blood-nerve barrier: enzymes, transporters and receptors--a comparison with the blood-brain barrier. Brain Res Bull, 52(1), 1-12 [DOI] [PubMed] [Google Scholar]
- Kuo YC and Lu CH . 2011 Effect of human astrocytes on the characteristics of human brain-microvascular endothelial cells in the blood-brain barrier. Colloids Surf B Biointerfaces, 86(1), 225-231 [DOI] [PubMed] [Google Scholar]
- Kashiwamura Y, Sano Y, Abe M, Shimizu F, Haruki H, Maeda T, et al. 2011 Hydrocortisone enhances the function of the blood-nerve barrier through the up-regulation of claudin-5. Neurochem Res, 36(5), 849-855 [DOI] [PubMed] [Google Scholar]
- Weidenfeller C, Schrot S, Zozulya A and Galla HJ . 2005 Murine brain capillary endothelial cells exhibit improved barrier properties under the influence of hydrocortisone. Brain Res, 1053(1-2), 162-174 [DOI] [PubMed] [Google Scholar]
- Franke H, Galla H and Beuckmann CT . 2000 Primary cultures of brain microvessel endothelial cells: a valid and flexible model to study drug transport through the blood-brain barrier in vitro. Brain Res Brain Res Protoc, 5(3), 248-256 [DOI] [PubMed] [Google Scholar]
- Franke H, Galla HJ and Beuckmann CT . 1999 An improved low-permeability in vitro-model of the blood-brain barrier: transport studies on retinoids, sucrose, haloperidol, caffeine and mannitol. Brain Res, 818(1), 65-71 [DOI] [PubMed] [Google Scholar]
- Hamm S, Dehouck B, Kraus J, Wolburg-Buchholz K, Wolburg H, Risau W, et al. 2004 Astrocyte mediated modulation of blood-brain barrier permeability does not correlate with a loss of tight junction proteins from the cellular contacts. Cell Tissue Res, 315(2), 157-166 [DOI] [PubMed] [Google Scholar]
- Ishizaki T, Chiba H, Kojima T, Fujibe M, Soma T, Miyajima H, et al. 2003 Cyclic AMP induces phosphorylation of claudin-5 immunoprecipitates and expression of claudin-5 gene in blood-brain-barrier endothelial cells via protein kinase a-dependent and -independent pathways. Exp Cell Res, 290(2), 275-288 [DOI] [PubMed] [Google Scholar]
- Wolburg H, Neuhaus J, Kniesel U, Krauss B, Schmid EM, Ocalan M, et al. 1994 Modulation of tight junction structure in blood-brain barrier endothelial cells . effects of tissue culture, second messengers and cocultured astrocytes. J Cell Sci, 107, 1347-1357 [DOI] [PubMed] [Google Scholar]