

Abstract
Objective
Streptococcus agalactiae (GBS) is an important cause of chorioamnionitis. This study characterizes GBS colonization and stimulation of antimicrobial responses in human extraplacental membranes using an ex vivo transwell two-compartment system of full-thickness membranes and live GBS.
Study Design
Human extraplacental membranes were affixed to transwell frames (without synthetic membranes). Live GBS was added to the decidual side of membranes in transwell cultures, and cocultures were incubated for 4, 8 and 24 h. GBS recovery from homogenized membranes and culture medium was determined by enumerating colony forming units (CFU) on blood agar. Antimicrobial peptide expression was identified using immunohistochemistry and ELISA. GBS killing by HBDs was assessed in vitro by incubating GBS with different human beta defensins (HBDs) for 3 h, then enumerating CFU.
Results
GBS recovery from membranes markedly decreased over time (P < 0.05). The antimicrobial peptides HBD-1, HBD-2, HBD-3, and lactoferrin were expressed in both GBS-exposed and non-exposed tissues. Notably, a pattern of localized increased HBD-2 in the amnion of GBS-infected tissue was observed. Moreover, GBS-treated membranes released increased amounts of HBD-2 into the amniotic and decidual compartments of the transwell cultures after 24 h (P < 0.05). In bacterial cultures, HBD-2 decreased GBS viability in a concentration-dependent manner (P < 0.05).
Conclusion
Innate immune responses in ex vivo human extraplacental membranes suppress GBS growth. HBD-2 was implicated in this GBS suppression with evidence of signal transduction across the tissue. Antimicrobial peptides may be important for innate immune defense against intrauterine GBS infections during pregnancy.
Keywords: Human beta defensin, antimicrobial peptides, extraplacental membranes, placenta, group B Streptococcus, pregnancy
Introduction
Streptococcus agalactiae or Group B Streptococcus (GBS) is associated with adverse pregnancy and neonatal outcomes (1). Genital GBS colonization occurs in up to 30% of pregnant women (1) and is associated with neonatal sepsis and meningitis. Moreover, preterm infants with GBS disease have increased risk of mortality compared to GBS-infected infants delivered at term (2–3). In one study, GBS was the microorganism most commonly isolated from maternal and fetal tissues in women with midgestation spontaneous abortions (4). Likewise, histological chorioamnionitis and preterm birth at less than 32 weeks gestation were positively associated with GBS isolation from extraplacental membranes (5). In nonhuman primates, GBS induces preterm labor (6). Moreover, GBS adheres to and infects human extraplacental membranes and cells (6–8).
Despite evidence linking GBS infection with adverse birth outcomes, the processes of GBS colonization and infection of extraplacental membranes are poorly understood. Here we adapted an ex vivo transwell two-compartment system of full-thickness human extraplacental membranes for coculture with live GBS to characterize GBS colonization of membranes and test the hypothesis that antimicrobial peptides mediate tissue bactericidal activity.
Materials and Methods
Reagents and Materials
The GBS used in this study was strain A909, initially isolated from a septic newborn (9) and transformed with plasmid encoding genes for Green Fluorescent Protein and erythromycin resistance (construct RS020, a gift from Amanda Jones, University of Washington). GBS was grown at 37 °C in planktonic culture using Todd Hewitt Broth (THB, Becton-Dickinson, Franklin Lakes, NJ) or on sheep’s blood agar plates (Blood Agar Base #2, Remel, Lenexa, KS and BBL defibrinated sheep blood, Franklin Lakes, NJ) with 5 μg/mL erythromycin (Acros Organics, Geel, Belgium). Media, buffers, fetal bovine serum (FBS) and penicillin/streptomycin (pen/strep) were from GIBCO (Grand Island, NY).
Culture of Extraplacental Membranes
Human extraplacental gestational membranes were collected from healthy pregnancies undergoing scheduled cesarean delivery prior to onset of labor at the University of Michigan Birth Center. Only healthy, non-smoking, singleton mothers were included. Women were excluded if they had collagen vascular disease, evidence of bacterial vaginosis, cervical cerclage, third trimester bleeding, multifetal pregnancy, immunocompromised conditions, major medical conditions (e.g., chronic renal disease, sarcoidosis, hepatitis, HIV), or if pathological evaluation of the placenta or membranes was warranted. Except for pre-operatively administered antibiotics, women were excluded if prescription antibiotics were used during the two weeks preceding delivery. The University of Michigan Institutional Review Board approved this research (IRBMED#HUM0013915). No evidence of necrosis or infection was observed histologically.
Extraplacental membranes were cultured in a two-compartment transwell system as described previously published (10–11). Briefly, membranes were dissected from placenta immediately following delivery and transported to the lab in Dulbecco’s phosphate-buffered saline (DPBS). Membranes were rinsed with medium and blood clots were removed. The membranes were then mounted on sterile transwell frames that had no synthetic membrane (gift from Corning, NY) and held in place with sterile latex elastic bands (ORMCO, Orange, CA). The membranes were affixed with the choriodecidua facing the inner chamber of the transwell and the amnion facing the outer chamber. The transwell inserts with membranes were placed in wells of 12-well culture plates containing Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 1% FBS and pen/strep. Our laboratory previously demonstrated that 1% FBS was sufficient to maintain tissue health and no benefit was added with higher FBS concentration (11). To maintain equal medium levels between the inner and outer transwell chambers, and thereby avoid potential confounding of results due to hydrostatic pressure, 0.5 mL medium was added to the smaller inner chamber and 1.5 mL medium was added to the larger outer chamber. Cultures were incubated at 37 °C and 5% CO2. After 4 h, the medium was exchanged for DMEM/1% FBS without antibiotics. Sample size was based on power estimates using data generated in prior experiments (with cytokines) in our laboratory.
GBS Coculture with Extraplacental Membranes
GBS in early exponential growth phase was diluted with DMEM/1% FBS to 1×106 colony forming units/mL (CFU/mL). Inoculant concentrations were validated by overnight growth on 5% sheep blood agar with erythromycin. Following a 24-h acclimation, the medium of the transwell choriodecidual compartment was replaced with 0.5 mL GBS inoculant (1×106 CFU/mL) or fresh DMEM/1%FBS without GBS (controls). Amnion compartment medium was also replaced with DMEM/1%FBS. Cocultures were then incubated for 4, 8, or 24 h. Coculture experiments were conducted in triplicate using gestational membranes from five women.
At designated time points, medium from each transwell chamber was collected. An aliquot of 100 μL medium was diluted for CFU determination on 5% sheep blood agar with incubation for 12–24 h at 37 °C, and the remainder was stored at −80 °C. Transwell inserts with attached tissue were transferred to fresh wells with DPBS in both chambers. The plate was gently rocked for 5 min to rinse away non-adherent bacteria. Two 3-mm biopsy punches were taken from each transwell-mounted tissue piece, placed in 1 mL PBS, and homogenized on ice with two 40-sec pulses. Tissue homogenates were serially diluted 1:10 in PBS, plated on 5% sheep blood agar in triplicate, and grown 12–24 h at 37 °C. Colony counts of viable bacteria in the membrane (CFU/mL/cm2) were determined.
To control for altered GBS growth in tissue culture medium (used for coculture of tissue and GBS), GBS growth was observed using the above protocol in transwell culture wells with intact polycarbonate membranes but without extraplacental membranes. Medium from the upper and lower compartments was plated on 5% sheep blood agar in triplicate and incubated overnight at 37 °C.
Immunohistochemistry
While still attached to the transwell insert, extraplacental membranes were fixed with 10% phosphate-buffered formalin (Fisher, Waltham, MA) for 24–48 h at 4 °C. The fixed tissue was gently removed from the transwell insert and a strip of membrane was cut ≤ 4 mm in width. Processing and staining were performed by the University of Michigan’s Comprehensive Cancer Center Tissue Core. The tissue strips were embedded in paraffin “on edge”, sectioned, and mounted on slides. For antimicrobial peptide staining, heat-induced antigen retrieval was performed in citrate buffer (pH 6.0). Immunoperoxidase staining was completed on a DAKO AutoStainer at room temperature using the LSAB+ System-HRP kit from DAKO. Briefly, peroxidase block was followed by a 30-min incubation with primary antibody at the dilutions indicated for the following antimicrobial peptides: human beta defensins (HBD)-1 (1:25, rabbit polyclonal, Santa Cruz), HBD-2 (1:25, goat polyclonal, Santa Cruz), HBD-3 (1:100, rabbit polyclonal, Novus), HBD-5 (1:100, goat polyclonal, Santa Cruz), elafin (1:200, rabbit polyclonal, Santa Cruz), or lactoferrin (1:400, rabbit polyclonal, Abcam). Samples were then incubated sequentially with biotinylated LINK (30 min), streptavidin-HPR (30 min), and 3, 3′ diaminobenzidine (DAB+) chromogen solution (5 min), before being counterstained with hematoxylin.
Microscopy image capture and analysis were done using Nikon Elements Software. For amnion cell size analysis, images were captured from slides stained for HBD-2 after 24 h in culture. We analyzed 7 images in 6 tissues, but one tissue had the amnion tear off in a way that allowed us to get only one image from that tissue. Approximately 5–7 amnion cells were measured per image and cell size was averaged for each tissue (N=5).
HBD ELISAs
HBD-1 and HBD-2 release from the extraplacental membranes into medium was measured using a commercially available enzyme linked immunosorbant assay (ELISA) kit according to manufacturer’s instructions (Peprotech, Rocky Hill, NJ). The HBD-1 ELISA detection range was 4–1000 pg/mL and the HBD-2 ELISA detection range was 16–2000 pg/mL. Samples were not diluted. The HBD values are reported as pg protein/mL medium.
Antimicrobial Activity of HBDs to GBS
Antimicrobial activity assays were done according to previously published methods (12). Briefly, HBDs (Peprotech, Rocky Hill, NJ) were resuspended according to manufacturer’s directions in 10 mM acetic acid and diluted in 0.1% BSA in PBS. GBS were grown for 2.5 h to exponential log phase at 37 °C with shaking in THB, diluted 1:1000 (approximately 1 × 104 CFUs) in 10 mM sodium phosphate buffer (pH 7.4), and then treated with 0–20 μg/mL of HBD-1, HBD-2, or HBD-3 for 3 h at 37 °C in a 96 well plate. Samples were serially diluted and 10 μL of each diluent was plated on 5% sheep blood agar in duplicate. Following overnight incubation at 37 °C, CFUs were enumerated. Data are expressed as bacterial survival relative to vehicle control (0.1 mM acetic acid, 0.01% BSA).
Statistical Analysis
Data are expressed as mean ± SEM and were analyzed using GraphPad Prism 5 software (GraphPad Software, La Jolla, CA). For transwell coculture GBS CFU quantification and HBD antimicrobial activity, ANOVAs with Tukey’s post hoc test were performed. For HBD ELISA and cell size analysis, Student’s paired t-test was used for each time point. Data were considered significant if the p-value was < 0.05.
Results
GBS Growth in Gestational Membranes
To simulate an ascending intrauterine infection, GBS was applied to the choriodecdiual compartment of the transwell-mounted gestational membranes. No GBS colonies formed from membranes of unexposed control tissues after 4, 8 or 24 h of coculture (Figure 1). GBS-infected tissues initially yielded increased CFUs after 4 h of coculture, but CFUs subsequently decreased in GBS-exposed membranes at 8 and 24 h (Figure 1; P < 0.05). A similar time-dependent pattern was observed for CFU recovery from medium of the cultures (data not shown). No GBS was recovered from the amnion compartment medium (not shown), suggesting that GBS does not cross the gestational membranes. Furthermore, GBS growth was robust in standard transwells (with synthetic membrane) containing DMEM tissue culture medium but lacking gestational tissue (Supplemental Data Figure 1), indicating that the decreased CFU recovery from coculture with gestational tissues was dependent on the presence of the tissue and not an artifact of the DMEM medium or transwell system.
Figure 1.

Recovered GBS (expressed as CFU) from homogenized tissue punches of the gestational membranes following coculture with 1×106 CFU/mL GBS inoculants for 4, 8, and 24 h. Control tissues were not exposed to GBS. Columns are mean ± SEM (n = 4–5 women, 3 replicates per woman). Asterisks (*) represent significant differences when compared by Tukey’s post-hoc test following ANOVA (P < 0.05).
Immunohistochemical Detection of Antimicrobial Peptides in Extraplacental Membranes
Immunohistochemial staining assessed expression of antimicrobial peptides in GBS-exposed and unexposed extraplacental membranes. Although HBD-2 antibody stained throughout the membranes in both the nucleus and the cytoplasm, HBD-2 staining increased in GBS-treated membranes after 24 h, especially in the amnion epithelium (Figure 2). In addition, amnion cells in GBS-treated tissues were significantly larger (8.3 μm) in diameter compared to amnion cells in untreated tissue (6.7 μm) (P < 0.05) at 24 h. Regardless of whether membranes were exposed in vitro to GBS or not, the amnion epithelial cells, chorionic trophoblasts, and decidual cells stained positive for HBD-1, HBD-3, and lactoferrin at all time points (Supplemental Data Figure 2). Extraplacental membranes did not show staining for HBD-5 or elafin in control tissues or those exposed to GBS (Supplementary Data Figure 3).
Figure 2.

Immunohistochemical staining for HBD-2 in human gestational membranes cocultured with GBS added to the choriodecidual chamber of the Transwell cultures. The top row (A, B, and C) shows representative images for no treatment controls. The middle row (D, E, and F) shows representative images for GBS-treated tissues (decidual side only). The bottom row (G, H, and I) shows representative images of negative control sections incubated with secondary antibody only. Tissues within a column were cultured from the same subject. Intense staining is visible in amnion epithelium (AM) of GBS-stimulated membranes (arrows), despite lack of direct contact of the amnion with GBS.
HBD-2 Secretion from Extraplacental Membranes
Extraplacental membranes in transwell culture released increased amounts of HBD-2 into the medium of the choriodecidual (1.65-fold) and amniotic compartments (1.59-fold) after 24 h of exposure to GBS (Figure 3; P < 0.05 compared with controls). There were no statistically significant differences in HBD-2 at 4 h and 8 h. Likewise, a 3.6-fold difference of HBD-1 between GBS and control tissues in amniotic chamber culture medium at 24 h was not statistically significant (Supplementary Data Figure 3), though this negative result should be interpreted with caution due to the modest sample size.
Figure 3.
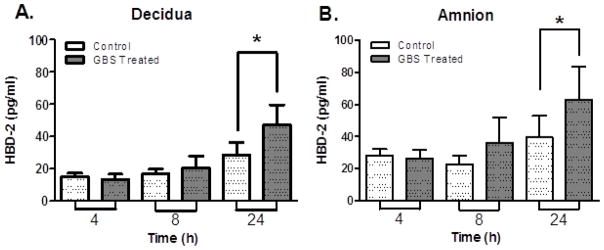
HBD-2 release into medium by extraplacental membranes in Transwell cultures with GBS (dark columns) and without GBS (light columns) exposure on the choriodecidual side. (A) Decidual compartment and (B) amniotic compartment. HBD-2 protein in the medium was measured by ELISA. Columns are mean ± SEM (n = 5 women, 3 replicates per woman). Asterisks (*) represent significant differences (P < 0.05) by paired student t-test.
HBD Bactericidal Activity against GBS
To investigate whether HBDs are capable of killing GBS directly, GBS was incubated with HBD-1, HBD-2, or HBD-3 in sodium phosphate buffer. HBD-2 killed GBS in a concentration-dependent manner, with nominal CFU recovered at 7.5 μg/ml and no GBS surviving exposure to the highest concentration of at 10 μg/mL of HBD-2 (Figure 4; P < 0.05). In contrast, HBD-1 and HBD-3 did not completely kill GBS at 10 μg/mL (data not shown).
Figure 4.
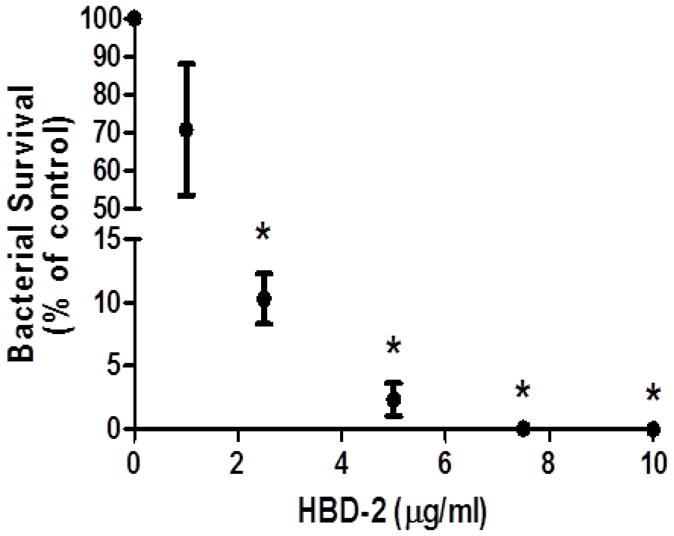
HBD-2 kills GBS. GBS were incubated in the presence of different concentrations of HBD-2 for 3 h. Data shown are mean ± SEM (n = 4 independent experiments), expressed as a percentage of bacterial survival relative to vehicle control. Asterisks (*) represent significant differences when compared by Tukey’s post-hoc test following ANOVA (P < 0.05).
Discussion
Although GBS remains the leading cause of infection-related neonatal morbidity and mortality, the mechanisms by which it interacts with and crosses extraplacental membranes as an ascending infection are poorly understood (1). Using a two-compartment tissue culture model, the present study provides evidence that GBS did not readily cross healthy term extraplacental membranes. Notably, antimicrobial innate immune responses, in particular HBD-2, were stimulated in amnion following GBS inoculation on the decidual side of the membranes, suggesting signal transduction across the tissue.
Although previous studies noted antimicrobial properties of extraplacental membranes to live bacteria (13–16), few studies quantified bactericidal activity of the tissue. The extraplacental membranes exhibited robust resistance to infection in vitro in our experiments. Viable GBS recovery from inoculum medium was reduced by at least half at 4, 8, and 24 h when cultured with extraplacental membranes, in agreement with Kjaergaard et al. who found reduced CFU counts of GBS recovered from inoculum medium over a 20-h incubation using a different two-compartment model of extraplacental membranes (17). In addition, we provide new information that GBS recovered from the tissue itself decreased over time, indicating that GBS adherence to and penetration into extraplacental membranes was inhibited.
HBDs are small cationic antimicrobial peptides thought to permeabilize microbial membranes (18). We show, for the first time to our knowledge, that extraplacental membranes express the antimicrobial peptide lactoferrin in the amniotic, chorionic trophoblast, and decidual cell layers, independent of exposure to GBS. Furthermore, similar to findings in extraplacental membranes of women delivering at term (18–19), we demonstrate that the antimicrobial peptides HBD-1, HBD-2, and HBD-3 were localized to the amnion, chorionic trophoblasts, and decidua of cultured extraplacental membrane explants, regardless of exposure to GBS. In particular, HBD-1 was highly expressed in all cell layers of extraplacental membranes irrespective of GBS treatment, consistent with other studies that suggest HBD-1 is constitutively expressed (20–21). Moreover, we found that secretion of HBD-1 into the medium did not change significantly with GBS stimulation. Our findings with HBD-1 are in agreement with reports by Garcia-Lopez et al. and Zaga-Clavellina et al. who had similar results with a different strain of GBS and E. coli. (18, 22–23). No evidence of necrosis or tissue degradation was observed in histological H&E slides, supporting evidence from the GBS killing and HBD-2 production data that the explants remained viable in culture throughout the experimental period.
GBS inoculation at the choriodecidual face of the membranes stimulated increased expression of HBD-2 in the amnion as visualized by immunostaining. The latter finding, in conjunction with reduced CFU recovery over time from the membranes, provides support that GBS stimulates signaling across the layers of the extraplacental membranes to increase HBD-2 production in amnion cells. Moreover, our observation that GBS stimulated an increase of amniotic cell size further supports the occurrence of cross-tissue signaling. Although novel with respect to HBD-2, previous reports suggest similar cross-tissue signaling in release of cytokines and prostaglandins by extraplacental membranes stimulated with LPS (24–26).
In addition to increased HBD-2 expression in amniotic cells, we observed increased HBD-2 in medium of the amniotic and choriodecidual compartments following choriodecidual stimulation with GBS, similar to findings reported by Zaga-Clavellina et al. (23). In contrast to the latter study, we used a different GBS strain and no antibiotics in our culture system, allowing us to quantify the bacterial death directly related to the extraplacental membranes innate immune response. In addition, this is the first report of the time-dependent increase of HBD-2 in both amniotic and choriodecidual compartments with GBS stimulation.
Recently, Garcia-Lopez and colleagues suggested that IL-1β may be a key mediator in the tissue-specific HBD-2 responses that they observed with Escherichia coli stimulation of extraplacental membranes in a similar transwell system (22). Just as GBS stimulation of the choriodecidua produced a tissue-specific increase of IL-1β secretion from the choriodecidua (10), E. coli stimulation of choriodecidua produced the same tissue-specific IL-1β secretion effect by the choriodecidua (27). A similar mechanism within the innate immune system of extraplacental membranes may be employed to protect against infection by Gram positive bacteria like GBS or Gram negative bacteria like E. coli. The cross-tissue signaling underlying the tissue-specific HBD responses observed in extraplacental membranes is not yet well characterized and warrants further study.
In studies comparing direct microbicidal activities of HBD-1, HBD-2 and HBD-3, we found that GBS strain A909 was most sensitive to killing by HBD-2 compared to HBD-1 and HBD-3, suggesting that increased HBD-2 secretion is linked to the GBS killing observed by us in extraplacental membranes. It is important to note that most in vitro bacterial viability studies (including ours) use synthetic or recombinant antimicrobial peptides at significantly higher levels than found in vivo (28), suggesting that antimicrobial peptides may be working synergistically in vivo (29).
Finally, formalin fixation of extraplacental membranes while mounted on the transwell frame, followed by “on edge” paraffin embedding of tissue strips, produced improved images for immunohistochemical assessment. The common technique for formalin fixation of extraplacental membranes is the “membrane roll,” which can produce histological sections with artifacts such as amnion shearing, in which amnion separates from choriodecidua. In the present study, amnion shearing was minimized and the sections better approximated a true cross section of the tissue.
In conclusion, this study utilized a two-compartment transwell extraplacental membrane tissue culture system to show that the antimicrobial response to GBS stimulation of extraplacental membrane explants was sufficiently robust to inhibit GBS colonization. We show for the first time signal transduction across the tissue increased HBD-2 production in the amnion epithelial cells in response to GBS. Moreover, we demonstrated that HBD-2 directly decreased GBS viability in a tissue-free system, suggesting a major role for HBD-2 in killing GBS in the extraplacental membranes.
Supplementary Material
Acknowledgments
We thank the University of Michigan pathology lab for help with embedding and staining our tissues. We also thank Mark Miller and Cassandra Korte for help with tissue culture experiments and helpful discussions.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Contributor Information
Erica Boldenow, Email: boldenow@umich.edu.
Sarah Jones, Email: jones.saraha@gmail.com.
Richard W. Lieberman, Email: jakrwl@med.umich.edu.
Mark C. Chames, Email: mchames@med.umich.edu.
David M. Aronoff, Email: daronoff@umich.edu.
Chuanwu Xi, Email: cxi@umich.edu.
Rita Loch-Caruso, Email: rlc@umich.edu.
References
- 1.Verani JR, McGee L, Schrag SJ. Prevention of perinatal group B streptococcal disease--revised guidelines from CDC, 2010. MMWR Recommendations and reports: Morbidity and mortality weekly report. Recommendations and reports/Centers for Disease Control. 2010 Nov 19;59(RR-10):1–36. [Practice Guideline] [PubMed] [Google Scholar]
- 2.Goldenberg RL, Hauth JC, Andrews WW. Intrauterine infection and preterm delivery. N Engl J Med. 2000 May 18;342(20):1500–7. doi: 10.1056/NEJM200005183422007. [DOI] [PubMed] [Google Scholar]
- 3.Jordan HT, Farley MM, Craig A, Mohle-Boetani J, Harrison LH, Petit S, et al. Revisiting the need for vaccine prevention of late-onset neonatal group B streptococcal disease: a multistate, population-based analysis. Pediatr Infect Dis J. 2008 Dec;27(12):1057–64. doi: 10.1097/INF.0b013e318180b3b9. [DOI] [PubMed] [Google Scholar]
- 4.McDonald HM, Chambers HM. Intrauterine infection and spontaneous midgestation abortion: is the spectrum of microorganisms similar to that in preterm labor? Infect Dis Obstet Gynecol. 2000;8(5–6):220–7. doi: 10.1155/S1064744900000314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hillier SL, Krohn MA, Kiviat NB, Watts DH, Eschenbach DA. Microbiologic causes and neonatal outcomes associated with chorioamnion infection. Am J Obstet Gynecol. 1991 Oct;165(4 Pt 1):955–61. doi: 10.1016/0002-9378(91)90447-y. [DOI] [PubMed] [Google Scholar]
- 6.Gravett MG, Haluska GJ, Cook MJ, Novy MJ. Fetal and maternal endocrine responses to experimental intrauterine infection in rhesus monkeys. Am J Obstet Gynecol. 1996 Jun;174(6):1725–31. doi: 10.1016/s0002-9378(96)70203-x. discussion 31–3. [DOI] [PubMed] [Google Scholar]
- 7.Galask RP, Varner MW, Petzold CR, Wilbur SL. Bacterial attachment to the chorioamniotic membranes. Am J Obstet Gynecol. 1984 Apr 1;148(7):915–28. doi: 10.1016/0002-9378(84)90534-9. [DOI] [PubMed] [Google Scholar]
- 8.Winram SB, Jonas M, Chi E, Rubens CE. Characterization of group B streptococcal invasion of human chorion and amnion epithelial cells In vitro. Infect Immun. 1998 Oct;66(10):4932–41. doi: 10.1128/iai.66.10.4932-4941.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lancefield RC, McCarty M, Everly WN. Multiple mouse-protective antibodies directed against group B streptococci. Special reference to antibodies effective against protein antigens. J Exp Med. 1975 Jul 1;142(1):165–79. doi: 10.1084/jem.142.1.165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Zaga V, Estrada-Gutierrez G, Beltran-Montoya J, Maida-Claros R, Lopez-Vancell R, Vadillo-Ortega F. Secretions of interleukin-1beta and tumor necrosis factor alpha by whole fetal membranes depend on initial interactions of amnion or choriodecidua with lipopolysaccharides or group B streptococci. Biol Reprod. 2004 Oct;71(4):1296–302. doi: 10.1095/biolreprod.104.028621. [DOI] [PubMed] [Google Scholar]
- 11.Thiex NW, Chames MC, Loch-Caruso RK. Tissue-specific cytokine release from human extra-placental membranes stimulated by lipopolysaccharide in a two-compartment tissue culture system. Reprod Biol Endocrinol. 2009;7:117. doi: 10.1186/1477-7827-7-117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Aronoff DM, Hao Y, Chung J, Coleman N, Lewis C, Peres CM, et al. Misoprostol impairs female reproductive tract innate immunity against Clostridium sordellii. J Immunol. 2008 Jun 15;180(12):8222–30. doi: 10.4049/jimmunol.181.4.2263. [Research Support, N.I.H., Extramural] [DOI] [PubMed] [Google Scholar]
- 13.Trelford JD, Trelford-Sauder M. The amnion in surgery, past and present. Am J Obstet Gynecol. 1979 Aug 1;134(7):833–45. doi: 10.1016/0002-9378(79)90957-8. [DOI] [PubMed] [Google Scholar]
- 14.Insausti CL, Alcaraz A, Garcia-Vizcaino EM, Mrowiec A, Lopez-Martinez MC, Blanquer M, et al. Amniotic membrane induces epithelialization in massive posttraumatic wounds. Wound Repair Regen. 2010 Jul-Aug;18(4):368–77. doi: 10.1111/j.1524-475X.2010.00604.x. [DOI] [PubMed] [Google Scholar]
- 15.Kjaergaard N, Hein M, Hyttel L, Helmig RB, Schonheyder HC, Uldbjerg N, et al. Antibacterial properties of human amnion and chorion in vitro. Eur J Obstet Gynecol Reprod Biol. 2001 Feb;94(2):224–9. doi: 10.1016/s0301-2115(00)00345-6. [DOI] [PubMed] [Google Scholar]
- 16.Gomes JA, Romano A, Santos MS, Dua HS. Amniotic membrane use in ophthalmology. Curr Opin Ophthalmol. 2005 Aug;16(4):233–40. doi: 10.1097/01.icu.0000172827.31985.3a. [DOI] [PubMed] [Google Scholar]
- 17.Kjaergaard N, Helmig RB, Schonheyder HC, Uldbjerg N, Hansen ES, Madsen H. Chorioamniotic membranes constitute a competent barrier to group b streptococcus in vitro. Eur J Obstet Gynecol Reprod Biol. 1999 Apr;83(2):165–9. doi: 10.1016/s0301-2115(99)00009-3. [DOI] [PubMed] [Google Scholar]
- 18.King AE, Paltoo A, Kelly RW, Sallenave JM, Bocking AD, Challis JR. Expression of natural antimicrobials by human placenta and fetal membranes. Placenta [Research Support, Non-US Gov’t] 2007 Feb-Mar;28(2–3):161–9. doi: 10.1016/j.placenta.2006.01.006. [DOI] [PubMed] [Google Scholar]
- 19.King AE, Kelly RW, Sallenave JM, Bocking AD, Challis JR. Innate immune defences in the human uterus during pregnancy. Placenta [Research Support, Non-US Gov’t Review] 2007 Nov-Dec;28(11–12):1099–106. doi: 10.1016/j.placenta.2007.06.002. [DOI] [PubMed] [Google Scholar]
- 20.Valore EV, Park CH, Quayle AJ, Wiles KR, McCray PB, Jr, Ganz T. Human beta-defensin-1: an antimicrobial peptide of urogenital tissues. J Clin Invest. 1998 Apr 15;101(8):1633–42. doi: 10.1172/JCI1861. [Research Support, U.S. Gov’t, P.H.S.] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Krisanaprakornkit S, Weinberg A, Perez CN, Dale BA. Expression of the peptide antibiotic human beta-defensin 1 in cultured gingival epithelial cells and gingival tissue. Infect Immun. 1998 Sep;66(9):4222–8. doi: 10.1128/iai.66.9.4222-4228.1998. [Research Support, Non-U.S. Gov’t Research Support, U.S. Gov’t, P.H.S.] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Garcia-Lopez G, Flores-Espinosa P, Zaga-Clavellina V. Tissue-specific human beta-defensins (HBD)1, HBD2, and HBD3 secretion from human extra-placental membranes stimulated with Escherichia coli. Reprod Biol Endocrinol. 2010;8:146. doi: 10.1186/1477-7827-8-146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zaga-Clavellina V, Garcia-Lopez G, Flores-Espinosa P. Evidence of in vitro differential secretion of human beta-defensins-1, -2, and -3 after selective exposure to Streptococcus agalactiae in human fetal membranes. J Matern Fetal Neonatal Med. 2011 Jun 1; doi: 10.3109/14767058.2011.578695. [DOI] [PubMed] [Google Scholar]
- 24.Thiex NW, Chames MC, Loch-Caruso RK. Tissue-specific induction of COX-2 and prostaglandins in lipopolysaccharide-stimulated extraplacental human gestational membranes in a 2-chamber transwell culture system. Reprod Sci. 2010 Dec;17(12):1120–9. doi: 10.1177/1933719110378344. [Research Support, N.I.H., Extramural Research Support, Non-U.S. Gov’t] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Thiex NW, Chames MC, Loch-Caruso RK. Tissue-specific cytokine release from human extra-placental membranes stimulated by lipopolysaccharide in a two-compartment tissue culture system. Reprod Biol Endocrinol. 2009;7:117. doi: 10.1186/1477-7827-7-117. [Research Support, N.I.H., Extramural Research Support, Non-U.S. Gov’t] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Zaga V, Estrada-Gutierrez G, Beltran-Montoya J, Maida-Claros R, Lopez-Vancell R, Vadillo-Ortega F. Secretions of interleukin-1beta and tumor necrosis factor alpha by whole fetal membranes depend on initial interactions of amnion or choriodecidua with lipopolysaccharides or group B streptococci. Biol Reprod. 2004 Oct;71(4):1296–302. doi: 10.1095/biolreprod.104.028621. [Comparative Study In Vitro Research Support, Non-U.S. Gov’t] [DOI] [PubMed] [Google Scholar]
- 27.Zaga-Clavellina V, Garcia-Lopez G, Flores-Herrera H, Espejel-Nunez A, Flores-Pliego A, Soriano-Becerril D, et al. In vitro secretion profiles of interleukin (IL)-1beta, IL-6, IL-8, IL-10, and TNF alpha after selective infection with Escherichia coli in human fetal membranes. Reprod Biol Endocrinol. 2007;5:46. doi: 10.1186/1477-7827-5-46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Starner TD, Agerberth B, Gudmundsson GH, McCray PB., Jr Expression and activity of beta-defensins and LL-37 in the developing human lung. J Immunol. 2005 Feb 1;174(3):1608–15. doi: 10.4049/jimmunol.174.3.1608. [Comparative Study Research Support, Non-U.S. Gov’t Research Support, U.S. Gov’t, P.H.S.] [DOI] [PubMed] [Google Scholar]
- 29.Chen X, Niyonsaba F, Ushio H, Okuda D, Nagaoka I, Ikeda S, et al. Synergistic effect of antibacterial agents human beta-defensins, cathelicidin LL-37 and lysozyme against Staphylococcus aureus and Escherichia coli. J Dermatol Sci. 2005 Nov;40(2):123–32. doi: 10.1016/j.jdermsci.2005.03.014. [In Vitro Research Support, Non-U.S. Gov’t] [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.