

Abstract
Calcium-signaling in cells requires a fine-tuned system of calcium-transport proteins involving ion channels, exchangers, and ion-pumps but also calcium-sensor proteins and their targets. Thus, control of physiological responses very often depends on incremental changes of the cytoplasmic calcium concentration, which are sensed by calcium-binding proteins and are further transmitted to specific target proteins. This Review will focus on calcium-signaling in vertebrate photoreceptor cells, where recent physiological and biochemical data indicate that a subset of neuronal calcium sensor proteins named guanylate cyclase-activating proteins (GCAPs) operate in a calcium-relay system, namely, to make gradual responses to small changes in calcium. We will further integrate this mechanism in an existing computational model of phototransduction showing that it is consistent and compatible with the dynamics that are characteristic for the precise operation of the phototransduction pathways.
Keywords: Calcium-signaling, cGMP, phototransduction
Critical biochemical reactions in rod and cone photoreceptor cells are under the control of negative feedback loops that determine the cell’s light sensitivity and its precise operation under different light regimes.1−4 One key factor that is involved in various feedback loops is Ca2+. Its concentration is balanced by two transport systems: In the dark state of the cell, the second messenger guanosine 3′,5′-cyclic monophosphate (cGMP) binds to a cyclic nucleotide-gated (CNG) channel in the plasma membrane, thereby opening the channel,5 which allows the influx of Na+ and Ca2+. The second transport system in the plasma membrane is a Na+/Ca2+, K+-exchanger that is extruding Ca2+. Illumination of a photoreceptor cell triggers the hydrolysis of cGMP, leading to the closure of CNG-channels, but the continuous operation of the exchanger is shuffling Ca2+ out of the cell resulting in a concomitant decrease of the cytoplasmic Ca2+-concentration.1,2 Ca2+-sensor proteins of the EF-hand superfamily of Ca2+-binding proteins are expressed in photoreceptor cells and detect these changes in Ca2+. Among them are guanylate cyclase-activating proteins (GCAPs) that constitute a subfamily of the neuronal Ca2+-sensor proteins.4,6−8 GCAPs control the activity of membrane bound guanylate cyclases in a Ca2+-dependent manner and are therefore essential key factors in shaping the rod and cone photoresponse under different light regimes.9,10 Two to eight different GCAP isoforms exist in species from fish to human. The apparently redundant expression of different GCAP forms in one cell type (rod or cone cell) was puzzling at the time, when these forms were first described. For example, mammalian GCAP1 and GCAP2 were found in rod and cone cells exhibiting almost identical biochemical properties.11−14 However, a few years later, electrophysiological recordings on transgenic mice lacking GCAP1, GCAP2, or both indicated that these neuronal calcium sensors have different roles in shaping the light response of a rod cell.15−17 Soon after this report, detailed biochemical studies showed that the Ca2+-sensitive activation profiles of bovine GCAP1 and 2 differ, which provided a molecular basis for interpreting the physiological data.18 The operation of different GCAP forms in rod or cone cells can be described in a Ca2+-relay model,19 for which we discuss in the following sections the experimental evidence and its relation to cone–rod diseases caused by GCAP1 mutations.
Contribution of GCAP1 and GCAP2 to the Flash Sensitivity of Photoreceptor Cells
Rods from transgenic mice lacking GCAP1 and 2 (GCAPs–/–) show a single flash response with a larger amplitude, a delayed time-to-peak, and a larger integration time when compared with wild-type mice (Figure 1).15 These characteristic features demonstrate the importance of guanylate cyclase regulation by GCAPs for the normal shape of the rod photoresponse. Expression of GCAP2 on a GCAP–/– genetic background resulted in variable flash responses, but in general GCAP2 expression could not rescue the initial rapid recovery phase typically observed in wildtype rods (see black trace in Figure 1).15 Consistent with these observations are single photon responses that were recorded from mice rods lacking GCAP2 (GCAP2–/–), which have a slower recovery to the baseline. Interestingly, the initial rapid recovery phase seems unchanged, whereas the later part of the recovery phase is delayed (see Figure 7 in ref (20)). A recent complementary study21 on mice lacking GCAP1 (GCAP1–/–) yields a qualitatively similar shape of the flash response as shown in Figure 1, that is, larger response amplitudes and delayed time-to-peak (the original recordings shown in Figure 6A from ref (21) are reproduced here in Figure 2A for comparison). However, Makino et al. report also in their study21 that the single photon responses from WT and GCAP1–/– rods show a remarkable heterogeneity (their Figure 6C–E is reproduced here in Figure 2B–D) in amplitudes and kinetics and some responses do not exhibit a full recovery to the baseline within 2 s (Figure 5 in ref (21)). It remains therefore an open question for further investigations, whether compensatory mechanisms change the transcription and translation pattern of GCAPs.21
Figure 1.
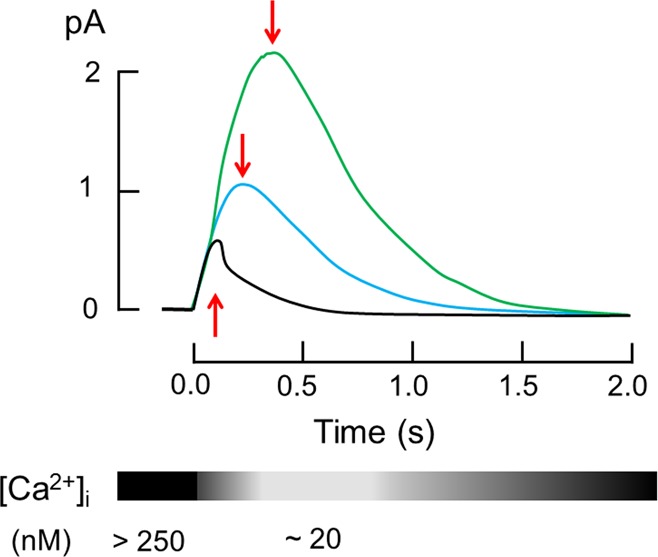
Idealized single photon responses of mice rods. Responses of rods from wild-type mice (black trace), from GCAPs–/– mice (green trace), and from mice expressing GCAP2 on a GCAPs–/– genetic background (blue trace) were superimposed. Red arrows indicate the time at which maximum amplitude is reached (time-to-peak). Traces represent typical idealized responses and were drawn according to published results in ref (15). The lower part of the figure shows the change of intracellular Ca2+-concentration during a light response.
Figure 2.
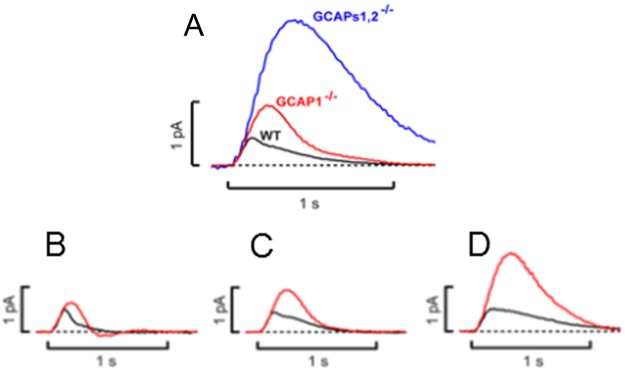
Heterogeneity of light responses from WT and transgenic mice. Original recordings were adapted from Figure 6 of ref (21) (Makino et al., PLoS One 7, e47637) in accordance with the Creative Commons Attribution License. (A) Comparison of averaged dim flash recordings from WT, GCAP1–/–, and GCAPs1,2–/– as indicated. (B–D) Variability of integration times of single photon responses as observed by Makino et al.21
Nevertheless, in comparison to WT rods, the time-to-peak is shifted by ∼100 ms in GCAP1–/– rods21 and only about 12 ms in GCAP2–/– rods20 indicating that GCAP1 and GCAP2 respond on a different time scale or on different free cytoplasmic Ca2+-concentration during the photoresponse.
The cytoplasmic Ca2+-concentration in mice is expected to drop from its initial dark value around 250 nM to 20 nM within a few hundred milliseconds.22 Different steady-state background light intensities probably adjust steady-state Ca2+-concentrations at intermediate levels. Although this has not been experimentally verified for mice rod or cone cells, one can anticipate it from data obtained on amphibian rods, where different intracellular Ca2+ concentrations were measured at different illumination conditions.23 While the precise value of the changing range of Ca2+ also depends on the species (dark values in the literature span a range from 250 to 800 nM, see, for instance, refs (22−29)), dynamic changes in Ca2+ are believed to strongly correlate with the illuminations state of the photoreceptor cell.
Collectively, these results point to a differential operation mode of GCAP1 and GCAP2 in rods and probably cones as well. In the next paragraph, we will review the biochemical studies that provide strong support of such a step-by-step activation modus of rod guanylate cyclases.
Activation Profiles of GCAP1 and GCAP2 Differ in Key Properties
Although GCAP1 and GCAP2 fold into similar three-dimensional structures that are also present as general folding motives in other neuronal calcium sensor proteins,30,31 they nevertheless display remarkable differences in their regulatory properties. This was explicitly stated for the first time in two biochemical in vitro studies by Hwang and Koch32 and Hwang et al.,18 in which the authors used bovine recombinant GCAP1 and GCAP2 reconstituted with either native guanylate cyclase 1 (ROS-GC1 or GC-E) in bovine rod outer segment membranes or tested GCAP function with ROS-GC1 expressed in HEK293 cells. The main findings of the studies were:
-
(a)
GCAP1 is activating ROS-GC1 at higher free Ca2+-concentration than GCAP2 (half-maximal activation expressed as IC50-value was at 700–1000 nM and at 100–200 nM, respectively).
-
(b)
GCAPs operate in a way that they increase the catalytic efficiency (kcat/Km) of the target ROS-GC1.
-
(c)
Myristoylation has a stronger impact on the activation profiles of GCAP1 than on the one of GCAP2.
While these studies were focused on ROS-GC1, subsequent work showed that GCAP1 and GCAP2 also activate bovine ROS-GC2 (alternatively named GC-F) with different Ca2+-sensitivities for each GCAP form (IC50 was 306 nM and 46 nM Ca2+ for GCAP1 and GCAP2, respectively).33 ROS-GC2 is a second sensory membrane guanylate cyclase in ROS that is expressed in a lower molar ratio to ROS-GC1. The exact ratio, however, seems to differ among species. For example, the ratios of ROS-GC1 to ROS-GC2 are 25:1 and 4:1 in bovine33 and mice34 outer segments, respectively. Biochemical studies on mice further supported the emerging picture that both guanylate cyclases are regulated in a differential manner by both GCAPs.34 A summarizing sketch of these results is shown in Figure 3: the Ca2+-dependent activation profiles of photoreceptor guanylate cyclases in the presence of either GCAP1 or GCAP2 represent a Ca2+-relay mode of GCAP action.
Figure 3.
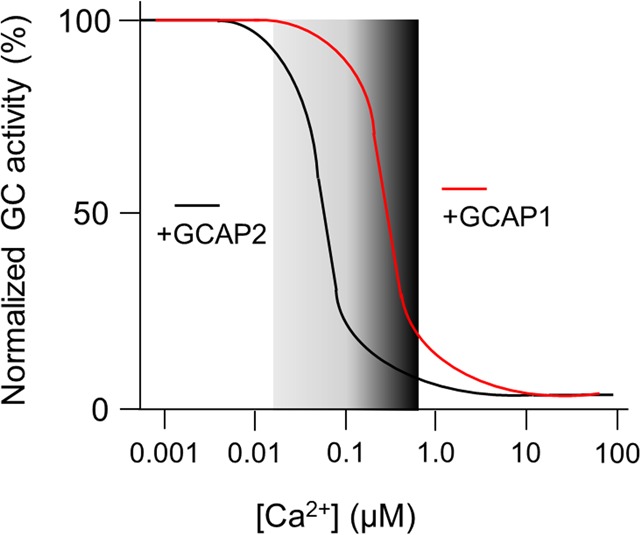
Ca2+-dependent activation profile of the photoreceptor guanylate cyclase is regulated by GCAP1 and GCAP2. The GC activity is normalized to 100%. The curves are typical for mammalian GCAPs and summarize the principal finding of many references (see main text). The gray bar symbolizes the physiological range of cytoplasmic Ca2+ that changes from a high (dark gray) to a low value (light gray).
To achieve a better comparison of electrophysiological data obtained from transgenic mice (see above) with data from biochemical measurements, Peshenko et al. performed a study using retinal homogenates from transgenic mice lacking the two ROS-GCs and GCAPs in different combinations.34 Further, Ca2+-dependent guanylate cyclase activities in retina homogenates of GCAP1–/– or GCAP2–/– displayed a characteristic shift of the Ca2+-dependent activation curves. While these studies confirm a differential regulatory mode of GCAP1 and GCAP2 action, the investigation by Peshenko et al. also revealed differences between mouse and bovine rod cells. For example, the ratio of the two GCs differed and their catalytic constants were in general higher in mice (see above).
In agreement with a differential action of GCAPs is the finding that GCAP1 and GCAP2 interact with different regions on ROS-GC1.35−39 Site-directed mutagenesis, the use of synthetic peptides representing the interaction domains, and cross-linking studies in combination with mass spectrometry revealed for GCAP1 a total of four interaction regions, where two are located in the juxtamembrane domain and two were found in the kinase homology domain.35,36 While these target regions are upstream from the cyclase catalytic domain, the interaction site of GCAP2 is located downstream on the C-terminal extension of the catalytic domain.37−39
In summary, the biochemical data obtained from mice, bovine, and purified recombinant samples is consistent with a model in which GCAP1 is turned on at higher free calcium concentration than GCAP2. Taking this observation into a physiological context means that both GCAPs are operating in a different time frame of the photoresponse, but also under different illumination conditions characterized by a changing pattern or oscillation of the intracellular Ca2+-concentration.
What Mechanisms Control the Tuning of Ca2+-Sensitivity?
Peshenko and Dizhoor demonstrated that GCAPs are Ca2+/Mg2+ sensors and that Mg2+ is a critical factor affecting the Ca2+-sensitivity.40 Decreasing the Mg2+-concentration in the incubation buffer can shift the Ca2+-sensitivity of ROS-GC regulation to lower Ca2+-concentration while keeping a difference between the IC50 values of GCAP1 and GCAP2. Moreover, recent results indicate that both the apparent affinity for Ca2+ of GCAP1 and GCAP2 and the dynamics of the conformational changes upon metal binding are significantly influenced by the presence of 1 mM free Mg2+.41 Another important factor involved in sensitivity regulation is the covalent fatty acylation, mainly a myristoylation (as mentioned already above) in GCAP1, which has almost no effect in GCAP2. Presence of the myristoyl group in GCAP1 is essential to shift the Ca2+-sensitivity into the physiological range and controls also the affinity of the GCAP1-ROS-GC1 interaction.18,42,43 Without an attached myristoyl group Ca2+-saturated GCAP1 remains in an active conformation and can significantly activate ROS-GC1 to about 20% of its maximal activity.32,43 The myristoyl group is located in a hydrophobic pocket31 and, in contrast to recoverin, it is not extruded by a switch mechanism upon changes in Ca2+-concentration. Interestingly, substitution of critical hydrophobic amino acids (Leu and Val) in the pocket by more bulky Phe residues restores almost the maximal activation of ROS-GC1 by GCAP1 and increases its affinity for ROS-GC1, but filling (or compensating) the hydrophobic pocket with bulky hydrophobic substituents does not change much the Ca2+sensitivity of ROS-GC1 regulation.43 A third parameter controlling the Ca2+-sensitivity could be the intermolecular tuning of the Ca2+-sensor with the corresponding target as this has been experimentally observed for calmodulin,44 but this is currently a controversial issue in the case of GCAPs. The observation that the control of GC by Ca2+ via the GCAPs is cooperative,45 while the binding of Ca2+ to individual GCAPs has been shown to occur sequentially and without cooperativity,41,46 suggests that the interaction with the target may influence the Ca2+-affinity of the sensor. This issue however awaits further clarification in future studies.
What about Cones?
Electroretinogram (ERG) recordings are sum potentials of retinal cells responding to light stimulation. ERG recordings on GCAPs–/– mice indicated that deletion of GCAPs delayed the recovery of cone light responses, which were partially restored in mice expressing transgenic GCAP1 on a GCAP null background.16,17 In a more recent study, single-cell recordings from cones of GCAPs–/– mice gave a detailed picture of the physiological function of GCAPs in cone photoreceptors.47 Characteristic features of light responses were a 2-fold longer time-to-peak and a 3-fold longer integration time, indicating that GCAPs are required for an efficient shutoff of the response on a short time scale, which is typical for cone response kinetics. However, as the authors of the study noted, the impact of GCAPs on modulation of the light response was smaller in comparison to rods despite the fact that cones adapt to a larger range of light intensities and have a wider span of dynamic changes in intracellular Ca2+. But mice are nocturnal animals and preferentially process olfactory information for orientation. Cones of mice might therefore express proteins with reduced capabilities. Thus, it will be worth testing whether animals with a cone system similar to humans have a more elaborated operation of GCAPs. Zebrafish, for example, possess four types of cone cells expressing a total set of nine cone opsins, thereby making the zebrafish retina responsive to visible and UV light.48 Due to the advantages of facilitated genetic manipulation and rapid larval development, work on zebrafish vision has increased tremendously in the past decade. Although we lack so far information from single-cell recordings of zebrafish cones with deletion of GCAPs, biochemical studies indicate a complex regulatory system for cGMP synthesis in zebrafish photoreceptor cells. Rod and cone cells express a total of three sensory guanylate cyclases and six different GCAP isoforms.49−51 Transcription of one guanylate cyclase (zGC3) and four GCAPs (zGCAP3, zGCAP4, zGCAP5, and zGCAP7) has been exclusively found in cones,49−51 and expression on the protein level has been demonstrated for zGCAP3.52 Two isoforms, zGCAP1 and zGCAP2, are expressed in rods and UV cones. During development, the cellular transcription pattern in the retina seems to change and may adapt to specific challenges of the aqueous habitat.50,51 Using purified samples of recombinant zGCAPs in biochemical reconstitution experiments showed a pattern of different activation profiles,53,54 which was similar to the ones observed with mammalian GCAPs (Figure 3). One can group the six zebrafish GCAP isoforms in two categories of different Ca2+- sensitivities, zGCAP4 and 7 switching between different GC activation profiles at higher Ca2+ compared to zGCAP1, 2, and 3. In addition, zGCAPs exhibit different properties with respect to Ca2+-binding and Ca2+-induced conformational changes pointing also to a step-by-step action of GCAPs in zebrafish cones.53
The Ca2+-Relay Mechanism in a Physiological Context
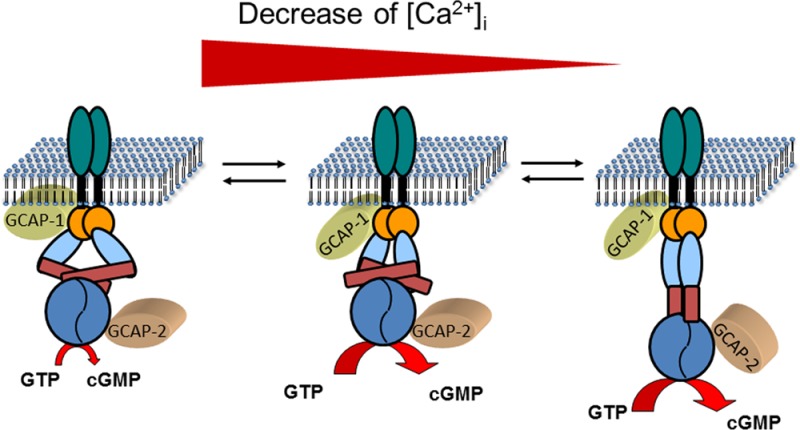
The cytoplasmic Ca2+-concentration decreases during the single flash response of a rod cell from the dark state value to a level well below 100 nM. Within the first 50 ms, the cytoplasmic Ca2+ has not reached its final value. This kind of intermediate state triggers activation of mammalian GCAP1 (or zGCAP4 and 7 in the zebrafish retina; see Figures 3 and 4 for comparison), which loses its bound Ca2+ and is transformed to its Mg2+-bound activator state (Figure 5). Several lines of experimental evidence show that GCAP1 undergoes a conformational change during this transformation, which is further transmitted to the catalytic center of the target GC (e.g., refs (11, 13, 31, 32, and 36)). Thus, GCAP1 steps into action and causes a first pulse of rapid cGMP synthesis, which contributes to effectively shut off the photoresponse. A characteristic fast response kinetics (Figure 1) is due to this step of cGMP synthesis, which is not seen in transgenic mice lacking GCAP1 or both GCAPs (Figures 1 and 2).15,21 A further decrease of cytoplasmic Ca2+ on a time scale of 0.5–1 s triggers the dissociation of Ca2+ from GCAP2 (or zGCAP1, 2, or 3 in the zebrafish retina), which now undergoes a conformational change leading to the activator state (Figure 5). The maximal activities of both GCAP-ROS-GC complexes are partially additive,20 but it remains unclear whether both GCAPs act synergistically on either ROS-GC1 or ROS-GC2 or whether they act on different GCs, which may even include a switch from a GCAP1 to a GCAP2 modus. In order to investigate the mechanistic implications of the Ca2+-relay model of guanylate cyclase regulation by GCAPs within its physiological context, a recently developed comprehensive kinetic model of phototransduction can be used. The model is optimized for amphibian rods and describes the synergic action of the biochemical events occurring in a rod photoreceptor cell upon light stimulation ranging over 5 orders of magnitude. The output consists of the simulated time course of the photocurrents, which can be compared with experimental electrophysiological recordings.55 The modular nature of the model allows the insertion of new mechanisms and the quantitative effects on the overall network can thus be directly assessed.56,57 In such a framework, the addition of the Ca2+-relay model can be realized by assuming that the rate of cGMP synthesis by ROS-GC1 is regulated by both GCAP1 and GCAP2 as follows:
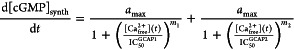 |
where amax = 60 μM/s45 is the maximal activation of GC1, assumed to be the same for both GCAP1 and GCAP2, [Ca2+free] is the free intracellular calcium depending on the illumination level, and IC50 (139 nM and 59 nM34) and m (1.7 and 2.4 (32)) indicate, respectively, the free Ca2+-concentration for half-maximal ROS-GC1 activation contributed by the specific GCAP and the Hill coefficient. We neglected any specific contribution of cGMP synthesis by ROS-GC2, because we lack at the moment essential data on putative switch mechanisms between a ROS-GC1 and ROS-GC2 operation modules (see above). However, the implementation of the Ca2+-relay mechanism based on ROS-GC1 regulation is fully compatible with the typical kinetics of photoresponses, as shown in Figure 6. The simulation of the photocurrent generated by six flashes of increasing intensity leading from 1.5 up to 11.5 × 104 photoisomerizations, that is, from very dim light to saturating conditions, is in line with the typical electrophysiological recordings. The significantly slower time-scale compared to mouse photoresponse is due to the fact that the model was developed for amphibian photoreceptors.
Figure 4.
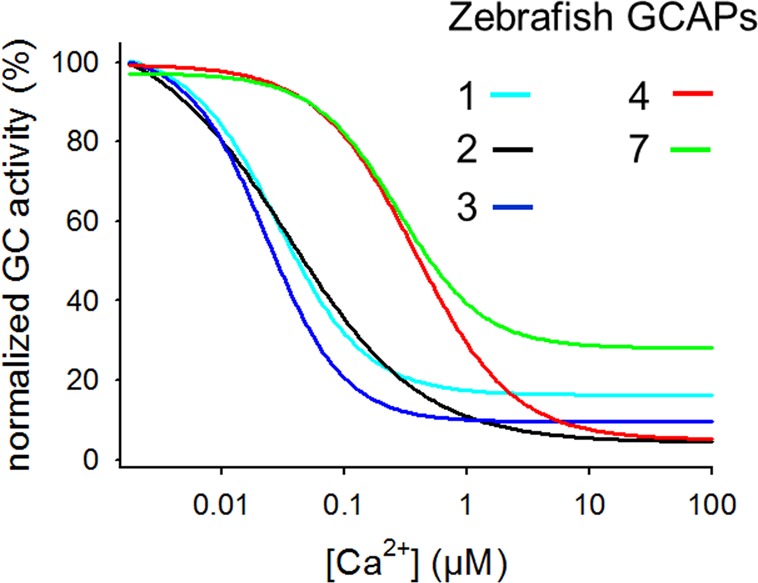
Activity profiles of cone specific GCAPs from zebrafish. The GC activity is normalized as in Figure 3. The curves were obtained by measuring the GC activity in the presence of zGCAPs as indicated. The figure is based on the original data published in ref (53), where a bovine ROS membrane preparation containing ROS-GCs was used for assaying zebrafish GCAPs.
Figure 5.

Ca2+-relay model of sequential GCAP action. Photoreceptor guanylate cyclases are dimers with one transmembrane domain in each monomer. In addition, each monomer consists of one extracelllar domain (in rods, this domain is in the lumen of the disks), one juxtamembrane domain (orange), and one kinase homology domain (light blue). Further, the dimerization domain (dark red rectangle) is important for the formation of an active enzyme controlling the correct positioning of the two catalytic domains (blue). In the dark state of the cell, GCAPs are fully or partially saturated with Ca2+ which keeps the guanylate cyclase activity at a very low level that is sufficient to keep a fraction of the CNG-channels in the plasma membrane open. GCAPs form with the target guanylate cyclase a complex in the presence and absence of Ca2+, which enables a rapid response to changing Ca2+-concentration after illumination. When the intracellular Ca2+-concentration falls to an intermediate level, Ca2+ dissociates form GCAP1. This process triggers a conformational change in GCAP1, leading to the activation of guanylate cyclase. When Ca2+ reaches its final lower intracellular level, GCAP2 turns into an activator. The different conformations of the guanylate cyclase are hypothetical and need to be verified in future experiments. It is further suggested that GCAPs stabilize the transition states of the cGMP catalytic step, but experimental proof is also lacking so far. The stepwise and reversible action of GCAPs would allow the cell to react on small incremental changes in Ca2+ with a fine-tuned response system.
Figure 6.

Simulated photocurrents of an amphibian rod stimulated by flashes of light of increasing intensity. The duration of each flash is 24 ms, and each stimulus leads to 1.54, 46, 280, 985, 30 400, and 115 000 photoisomerizations of rhodopsin. The mathematical model used for the simulation is the one reported in ref (57) in which the Ca2+-relay mechanism has been implemented as described here.
The computational implementation of the Ca2+-relay model allows assessing each term defining the rate of cGMP synthesis, thus deriving the specific contribution of GCAP1 and GCAP2 regulation of ROS-GC1 over time. Figure 7 shows that, at dim light intensities, when the Ca2+-concentration decreases mildly compared to the dark state, the most important contribution to the cGMP synthesis (black line) derives almost entirely from the GCAP1-mediated stimulation of ROS-GC1 (blue line), while the contribution of GCAP2 (red line) is modest. GCAP1 is thus the first calcium sensor stepping into action for a fast activation of ROS-GC1 (and maybe ROS-GC2 as well). However, already at intermediate illumination levels, the contribution of GCAP2 becomes more prominent, and at saturating conditions (Figure 7E and F) it quantitatively equals that of GCAP1, although the activation profile is slower and, due to the shorter time the cell spends at very low calcium, less broad over the whole photoresponse time range. Overall, the simulations show full consistence with the concept of Ca2+-relay mechanism shaping the photoresponse of vertebrates.
Figure 7.

Simulated time course of the synthesis of cGMP by GC1 under the Ca2+-mediated regulation by GCAP1 and GCAP2 according to the Ca2+-relay mechanism. Panels A–F refer to the same flashes of increasing intensity reported in Figure 6 (i.e., panel A refers to 1.54 photoisomerization, panel B to 46 photoisomerizations, and so forth). Blue lines report on the contribution of GCAP1 to the rate of synthesis, while red lines to that of GCAP2. The two contributions add up to form the overall rate (black line).
Disturbance of the Ca2+-Relay Modus in Retinal Diseases
Cone and cone–rod dystrophies are hereditary diseases that correlate with mutations in retinal proteins. One group of patients carries mutations in the GUCA1A gene coding for human GCAP1 (for a summary, see refs (58 and 59). So far, only one very rare mutation was identified in GCAP2.60 Nine mutations found in GCAP1 have been investigated in more detail including in particular the Ca2+-sensitive regulatory properties. Eight mutations are found in or near the EF-hands 3 and 4, indicating an impairment of Ca2+-binding and/or Ca2+-dependent properties. When recombinant forms of these GCAP1 mutants were investigated in reconstitution experiments with membranes containing photoreceptor guanylate cyclase, the Ca2+-dependent activation profiles of ROS-GC1 were shifted to higher free Ca2+-concentration.61−68 Thus, under normal physiological conditions these GCAP forms are permanently active leading to continuous high synthesis rates of cGMP. Some GCAP1 mutants would even require Ca2+-concentrations well above 100 μM to be turned off. These findings demonstrate that an imbalance of the finely tuned Ca2+-cGMP homeostasis can lead to severely impaired cellular functions. Since GCAP1 and GCAP2 respond sequentially to Ca2+ in a narrow range, the shifted response curve of GCAP1 is setting the Ca2+-relay operation modus out of control.
Other Ca2+-dependent Feedback Loops
Two other Ca2+-sensor proteins, calmodulin and recoverin, are also mainly involved in regulation of phototransduction processes. Ca2+-calmodulin binds directly to the CNG-channel and thereby decreases the affinity of the channel for its ligand cGMP69−71 and Ca2+-loaded recoverin inhibits rhodopsin kinase in the dark state of the cell.72,73 When Ca2+-levels drop after illumination, calmodulin dissociates from binding sites in the CNG channel, making the channel susceptible for lower cGMP concentration, which facilitates the reopening of the channel at lower cGMP levels that are present in dark adapted cells.
Experiments on photoreceptor cells of recoverin knockout mice revealed that recoverin is an essential component of the light-dependent modulation of the lifetime of light-activated rhodopsin.74 A more recent biochemical study also showed that the Ca2+-sensitive inhibition of rhodopsin kinase by recoverin can be increased by the synergetic effect of calmodulin placing the inhibitory action in a range of 0.05–1 μM free Ca2+-concentration.75 This mechanism of shifting the Ca2+-sensitive regulation of the target enzyme rhodopsin kinase is reminiscent of the Ca2+-relay model described above for the GCAPs and its physiological relevance is an open question for future studies.
Conclusion and Prospects
GCAPs in the vertebrate retina are calcium-sensor proteins that operate in a relay mode and make thereby the cell responsive to incremental changes in intracellular calcium. Biochemical and physiological data as well as results from a systems biology approach support this Ca2+-relay model, but also impose further questions. For example, flash responses delivered in the presence of background light showed that the flash sensitivity of rods and cones of GCAP–/–mice displayed a deviation from normal Weber-Fechner relation, indicating a partial loss of adaptation mechanisms.15,47 These results lead to unresolved questions such as how does a step-by-step action of GCAPs operate under constant illumination or under different background light intensities? Furthermore, is there a shift in the Ca2+-sensitivities of GCAPs under prolonged illumination? Which other mechanisms beside the Ca2+-dependent regulation of ROS-GCs control light adaptation of photoreceptor cells? Further, is the larger diversity of GCAPs in teleost fish the molecular basis for the dynamic range extension in cone adaptation of the fish retina? The heterogeneity and variability of flash responses obtained with transgenic mice21 impose also further questions about compensatory mechanisms (see above) and certainly reflect our incomplete understanding of GCAP function.
GCAP1 and GCAP2 are also localized in the synaptic layer of photoreceptor cells,13,14 and it has recently been reported that GCAP2 interacts with the protein ribeye that is part of the synaptic ribbon structure.76,77 Further, overexpression of GCAP2 leads to morphological changes of the ribbon synapse.78 Does GCAP2 operate as a Ca2+-sensor for more than one target in synaptic terminals including ribeye and ROS-GC1? The latter has also been localized in the outer plexiform layer of vertebrate retinae and could be under control of changing Ca2+-levels. Finally, a Ca2+-relay mode of operation could involve other Ca2+-sensors as well regulating, for example, synaptic vesicle release or light-dependent transport and translocation processes.
Glossary
Abbreviations
- GCAP
guanylate cyclase-activating protein
- ROS
rod outer segment
- GC
guanylate cyclase
- CNG
cyclic nucleotide-gated
Author Contributions
K.W.K. conceived the study, D.D.O. performed the computational simulation, both authors wrote and approved the paper.
The authors declare no competing financial interest.
References
- Pugh E. N. Jr., and Lamb T. D. (2000) Phototransduction in vertebrate rods and cones: molecular mechanisms of amplification, recovery and light adaptation. In Handbook of Biological Physics (Stavenga D. G., DeGrip W. J., and Pugh E. N. Jr., Eds.), Vol. 4, pp 183–255, Elsevier Science BV: Amsterdam, The Netherlands. [Google Scholar]
- Luo D. G.; Xue T.; Yau K.-W. (2008) How vision begins: an odyssey. Proc. Natl. Acad. Sci. U.S.A. 105, 9855–9862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stephen R.; Filipek S.; Palczewski K.; Sousa M. C. (2008) Ca2+-dependent regulation of phototransduction. Photochem. Photobiol. 84, 903–910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koch K.-W.; Duda T.; Sharma R. K. (2010) Ca2+-modulated vision-linked ROS-GC guanylate cyclase transduction machinery. Mol. Cell. Biochem. 334, 105–115. [DOI] [PubMed] [Google Scholar]
- Kaupp U. B.; Seifert R. (2002) Cyclic nucleotide-gated ion channels. Physiol. Rev. 82, 769–824. [DOI] [PubMed] [Google Scholar]
- Palczewski K.; Polans A. S.; Baehr W.; Ames J. B. (2000) Ca2+-binding protiens in the retina: structure, function, and the etiology of human visual diseases. BioEssays 22, 337–350. [DOI] [PubMed] [Google Scholar]
- Dizhoor A. M.; Olshevskaya E. V.; Peshenko I. V. (2010) Mg2+/Ca2+ cation binding cycle of guanylyl cyclase activating proteins (GCAPs): role in regulation of photoreceptor guanylyl cyclase. Mol. Cell. Biochem. 334, 117–124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burgoyne R. D. (2007) Neuronal calcium sensor proteins: generating diversity in neuronal Ca2+ signaling. Nat. Rev. Neurosci. 8, 182–193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burns M. E.; Mendez A.; Chen J.; Baylor D. A. (2002) Dynamics of cyclic GMP synthesis in retinal rods. Neuron 36, 81–91. [DOI] [PubMed] [Google Scholar]
- Gross O. P.; Pugh E. N. Jr; Burns M. (2012) Calcium feedback to cGMP synthesis strongly attenuates single-photon responses driven by long rhodopsin lifetimes. Neuron 76, 370–382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palczewski K.; Subbaraya I.; Gorczyca W. A.; Helekar B. S.; Ruiz C.; Ohguro H.; Huang J.; Zhao X.; Crabb J. W.; Johnson R. S.; Walsh K. A.; Gray-Keller M. P.; Detwiler P. B.; Baher W. (1994) Molecular cloning and characterization of retinal photoreceptor guanylyl cyclase-activating protein. Neuron 13, 395–404. [DOI] [PubMed] [Google Scholar]
- Gorczyca W. A.; Polans A. S.; Surgucheva I. G.; Subbaraya I.; Baehr W.; Palczewski K. (1995) Guanylyl cyclase activating protein. A calcium-sensitive regulator of phototransduction. J. Biol. Chem. 270, 22029–22036. [DOI] [PubMed] [Google Scholar]
- Dizhoor A. M.; Olshevskaya E. V.; Henzel W. J.; Wong S. C.; Stults J. T.; ankoudinova I.; Hurley J. B. (1995) Cloning, sequencing, and expression of a 24-kDa Ca(2+)-binding protein activating photoreceptor guanylyl cyclase. J. Biol. Chem. 270, 25200–25206. [DOI] [PubMed] [Google Scholar]
- Frins S.; Bonigk W.; Muller F.; Kellner R.; Koch K.-W. (1996) Functional characterization of a guanylyl cyclase-activating protein from vertebrate rods. Cloning, heterologous expression, and localization. J. Biol. Chem. 271, 8022–8027. [DOI] [PubMed] [Google Scholar]
- Mendez A.; Burns M. E.; Sokal I.; Dizhoor A. M.; Baehr W.; Palczewski K.; Baylor D.; Chen J. (2001) Role of guanylate cyclase-activating proteins (GCAPs) in setting the flash sensitivity of rod photoreceptors. Proc. Natl. Acad. Sci. U.S.A. 98, 9948–9953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howes K. A.; Pennesi M. E.; Sokal I.; Church-Kopish J.; Schmidt B.; Margolis D.; Frederick J. M.; Rieke F.; Palczewski K.; Wu S. M.; Detwiler P. B.; Baehr W. (2002) GCAP1 rescues rod photoreceptor response in GCAP1/GCAP2 knockout mice. EMBO J. 21, 1545–1554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pennesi M. E.; Howes K. A.; Baehr W.; Wu S. M. (2003) Guanylate cyclase-activating protein (GCAP) 1 rescues cone recovery kinetics in GCAP1/GCAP2 knockout mice. Proc. Natl. Acad. Sci. U.S.A 100, 6783–6788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hwang J. Y.; Lange C.; Helten A.; Höppner-Heitmann D.; Duda T.; Sharma R. K.; Koch K.-W. (2003) Regulatory modes of rod outer segment membrane guanylate cyclase differ in catalytic efficiency and Ca2+-sensitivity. Eur. J. Biochem. 270, 3814–3821. [DOI] [PubMed] [Google Scholar]
- Koch K.-W. (2006) GCAPs, the classical neuronal calcium sensors in the retina – a Ca2+-relay model of guanylate cyclase activation. Calcium Binding Proteins 1, 3–6. [Google Scholar]
- Makino C. L.; Peshenko I. V.; Wen X. H.; Olshevskaya E. V.; Barrett R.; Dizhoor A. M. (2008) A role for GCAP2 in regulating photoresponse. J. Biol. Chem. 283, 29135–29143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Makino C. L.; Wen X. H.; Olshevskaya E. V.; Peshenko I. V.; Savchenko A. B.; Dizhoor A. M. (2012) Enzymatic relay mechanism stimulates cyclic GMP synthesis in rod photoresponse: biochemical and physiological study in guanyly cyclase activating protein 1 knockout mice. PLoS One 7, e47637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woodruff M. L.; Sampath A. P.; Matthews H. R.; Krasnoperova N. V.; Lem J.; Fain G. L. (2002) Measurement of cytoplasmic calcium concentration in the rods of wild-type and transducin knock-out mice. J. Physiol. 543, 843–854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gray-Keller M. P.; Detwiler P. B. (1996) Ca2+ dependence of dark- and light-adapted flash responses in rod photoreceptors. Neuron 17, 323–331. [DOI] [PubMed] [Google Scholar]
- Korenbrot J. I.; Miller D. L. (1989) Cytoplasmic free calcium concentraton in dark-adapted retinal rod outer segments. Vision Res. 29, 939–948. [DOI] [PubMed] [Google Scholar]
- McCarthy S. T.; Younger J. P.; Owen W. G. (1996) Dynamic, spatially nonuniform calcium regulation in frog rods exposed to light. J. Neurophysiol. 76, 1991–2004. [DOI] [PubMed] [Google Scholar]
- Sampath A. P.; Matthews H. R.; Cornwall M. C.; Bandarchi J.; Fain G. L. (1999) Light-dependnet changes in outer segment free Ca2+ concentration in salamander cone photoreceptors. J. Gen. Physiol. 113, 267–277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakatani K.; Chen C.; Yau K.-W; Koutalos Y. (2002) Calcium and phototransduction. Adv. Exp. Med. Biol. 514, 1–20. [DOI] [PubMed] [Google Scholar]
- Matthews H. R.; Fain G. L. (2002) Time course and magnitude of the calcium release induced by bright light in salamander rods. J. Physiol. 542.3, 829–841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matthews H. R.; Fain G. L. (2003) The effect of light on outer segment calcium in salamander rods. J. Physiol. 552.3, 763–776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ames J. B.; Dizhoor A. M.; Ikura M.; Palczewski K.; Stryer L. (1999) Three-dimensional structure of guanylyl cyclase activating protein-2, a calcium-sensitive modulator of photoreceptor guanylyl cyclases. J. Biol. Chem. 274, 19329–19337. [DOI] [PubMed] [Google Scholar]
- Stephen R.; Bereta G.; Golczak M.; Palczewski K.; Sousa M. C. (2007) Stabilizing function for myristoyl group revealed by the crystal structure of a neuronal calcium sensor, guanylate cyclase-activating protein 1. Structure 15, 1392–1402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hwang J.-Y.; Koch K.-W. (2002) Calcium- and myristoyl-dependent properties of guanylate cyclase-activating protein-1 and protein-2. Biochemistry 41, 13021–13028. [DOI] [PubMed] [Google Scholar]
- Helten A.; Säftel W.; Koch K.-W. (2007) Expression level and activity profile of membrane bound guanylate cyclase type 2 in rod outer segments. J. Neurochem. 103, 1439–1446. [DOI] [PubMed] [Google Scholar]
- Peshenko I. V.; Olshevskaya E. V.; Savchenko A. B.; Karan S.; Palczewski K.; Baehr W.; Dizhoor W. (2011) Enzymatic properties and regulation of the native isozymes of retinal membrane guanylyl cyclase (RetGC) from mouse photoreceptors. Biochemistry 50, 5590–5600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lange C.; Duda T.; Beyermann M.; Sharma R. K.; Koch K.-W. (1999) Regions in vertebrate photoreceptor guanylyl cyclase ROS-GC1 involved in Ca2+-dependent regulation by guanylyl cyclase-activating protein GCAP-1. FEBS Lett. 460, 27–31. [DOI] [PubMed] [Google Scholar]
- Krylov D. M.; Hurley J. B. (2001) Identification of proximate regions in a complex of retinal guanylyl cyclase 1 and guanylyl cyclase-activating protein-1 by a novel mass spectrometry-based method. J. Biol. Chem. 276, 30648–30654. [DOI] [PubMed] [Google Scholar]
- Sokal I.; Haeseleer F.; Arendt A.; Adman E. T.; Hargrave P. A.; Palczewski K. (1999) Identification of a guanylyl cyclase-activating protein-binding site within the catalytic domain of retinal guanylyl cyclase 1. Biochemistry 38, 1387–1393. [DOI] [PubMed] [Google Scholar]
- Duda T.; Fik-Rymarkiewicz E.; Venkataraman V.; Krishnan R.; Koch K.-W.; Sharma R. K. (2005) The calcium-sensor guanylate cyclase activating protein type 2 specific site in rod outer segment membrane guanylate cyclase type 1. Biochemistry 44, 7336–7345. [DOI] [PubMed] [Google Scholar]
- Pettelkau J.; Schröder T.; Ihling C. H.; Olausson B. E. S.; Kölbel K.; Lange C.; Sinz A. (2012) Structural insights into retinal guanylycyclase-GCAP2 interaction determined by cross-linking and mass spectrometry. Biochemistry 51, 4932–4949. [DOI] [PubMed] [Google Scholar]
- Peshenko I. V.; Dizhoor A. M. (2004) Guanylyl cyclase-activating proteins (GCAPs) are Ca2+/Mg2+ sensors: implications for photoreceptor guanylyl cyclase (RetGC) regulation in mammalian photoreceptors. J. Biol. Chem. 279, 16903–16906. [DOI] [PubMed] [Google Scholar]
- Dell’Orco D.; Sulmann S.; Linse S.; Koch K.-W. (2012) Dynamics of conformational Ca2+-switches in signaling networks detected by a planar plasmonic device. Anal. Chem. 84, 2982–2989. [DOI] [PubMed] [Google Scholar]
- Peshenko I. V.; Olshevskaya E. V.; Dizhoor A. M. (2012) Interaction of GCAP1 with retinal guanylyl cyclase and calcium: sensitivity to fatty acylation. Front. Mol. Neurosci. 5, 19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peshenko I. V.; Olshevskaya E. V.; Lim S.; Ames J. B.; Dizhorr A. M. (2012) Calcium-myristoyl tug is a new mechanisms for intramolecular tuning of calcium sensitivity and tarbet enzyme interaction for guanylyl cyclase-activating protein 1. J. Biol. Chem. 287, 13972–13984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peersen O. B.; Madsen T. S.; Falke J. J. (1997) Intermolecular tuning of calmodulin by target peptides and proteins: differential effects on Ca2+ binding and implications for kinase activation. Protein Sci. 6, 794–807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koch K.-W.; Stryer L. (1988) Highly cooperative feedback control of retinal rod guanylate cyclase by calcium ions. Nature 334, 64–66. [DOI] [PubMed] [Google Scholar]
- Dell’Orco D.; Behnen P.; Linse S.; Koch K.-W. (2010) Calcium binding, structural stability and guanylate cyclase activation in GCAP1 variants associated with human cone dystrophy. Cell. Mol. Life. Sci. 67, 973–984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sakurai K.; Chen J.; Kefalov V. J. (2011) Role of guanylyl cyclase modulation in mouse cone phototransduction. J. Neurosci. 31, 7991–8000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chinen A.; Hamaoka T.; Yamada Y.; Kawamura S. (2003) Gene duplication and spectral diversification of cone visual pigments of zebrafish. Genetics 163, 663–675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Imanishi Y.; Yang L.; Sokal I.; Filipek S.; Palczewski K.; Baehr W. (2004) Diversity of guanylate cyclase-activating proteins (GCAPS) in teleost fish: characterization of three novel GCAPs (GCAP4, GCAP5, GCAP7) from zebrafish (Danio rerio) and prediction of eight GCAPs (GCAP1–8) in pufferfish (Fugu rubripes). J. Mol. Evol. 59, 2204–2217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rätscho N.; Scholten A.; Koch K.-W. (2009) Expression profiles of three novel sensory guanylate cyclases and guanylate cyclase-activating proteins in the zebrafish retina. Biochim. Biopyhs. Acta 1793, 1110–1114. [DOI] [PubMed] [Google Scholar]
- Rätscho N.; Scholten A.; Koch K.-W. (2010) Diversity of guanylate cyclases in teleost fishes. Mol. Cell. Biochem. 334, 207–214. [DOI] [PubMed] [Google Scholar]
- Fries R.; Scholten A.; Säftel W.; Koch K.-W. (2012) Operation profile of zebrafish guanylate cyclase-activating protein 3. J. Neurochem. 121, 54–65. [DOI] [PubMed] [Google Scholar]
- Scholten A.; Koch K.-W. (2011) Differential calcium signaling by cone specific guanylate cyclase-activating proteins from the zebrafish retina. PLoS One 6, e23117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Behnen P.; Scholten A.; Rätscho N.; Koch K.-W. (2009) The cone-specific calcium sensor guanylate cyclase activating protein 4 from the zebrafish retina. J. Biol. Inorg. Chem. 14, 89–99. [DOI] [PubMed] [Google Scholar]
- Dell’Orco D.; Schmidt H.; Mariani S.; Fanelli F. (2009) Network-level analysis of light adaptation in rod cells under normal and altered conditions. Mol. Biosyst. 5, 1232–1246. [DOI] [PubMed] [Google Scholar]
- Dell’Orco D.; Koch K.-W. (2010) Systems biochemistry approaches to vertebrate phototransduction: towards a molecular understanding of disease. Biochem. Soc. Trans. 38, 1275–1280. [DOI] [PubMed] [Google Scholar]
- Dell’Orco D.; Koch K.-W. (2011) A dynamic scaffolding mechanism for rhodopsin and transducin interaction in vertebrate vision. Biochem. J. 440, 263–271. [DOI] [PubMed] [Google Scholar]
- Hunt D. M.; Buch P.; Michaelides M. (2010) Guanylate cyclases and associated activator proteins in retinal diseases. Mol. Cell. Biochem. 334, 157–168. [DOI] [PubMed] [Google Scholar]
- Behnen P.; Dell’Orco D.; Koch K.-W. (2010) Involvement of the calcium sensor GCAP1 in hereditary cone dystrophies. Biol. Chem. 391, 631–637. [DOI] [PubMed] [Google Scholar]
- Sato M.; Nakazawa M.; Usui T.; Tanimoto N.; Abe H.; Ohguro H. (2005) Mutations in the gene coding for guanylate cyclase-activating protein 2 (GUCA1B gene) in patients with autosomal dominat retinal dystrophies. Graefés Arch. Clin. Exp. Ophthalmol. 243, 235–242. [DOI] [PubMed] [Google Scholar]
- Kitiratschky V. B.; Behnen P.; Kellner U.; Heckenlively J. R.; Zrenner E.; Jägle H.; Kohl S.; Wissinger B.; Koch K.-W. (2009) Mutations in the GUCA1A gene involved in hereditary cone dystrophies impair calcium-mediated regulation of guanylate cyclase. Hum. Mutat. 30, E782–E796. [DOI] [PubMed] [Google Scholar]
- Sokal I.; Li N.; Surgucheva I.; Warren M. J.; Payne A. M.; Bhattacharya S. S.; Baehr W.; Palczewski K. (1998) GCAP1 (Y99C) mutant is constitutively active in autosomal dominant cone dystrophy. Mol. Cell 2, 129–133. [DOI] [PubMed] [Google Scholar]
- Olshevskaya E. V.; Calvert P. D.; Woodruff M. L.; Peshenko I. V.; Savchenko A. B.; Makino C. L.; Ho Y. S.; Fain G. L.; Dizhoor A. M. (2004) The Y99C mutation in guanylyl cyclase-activating protein 1 increases intracellular Ca2+ and causes photoreceptor degeneration in transgenic mice. J. Neurosci. 24, 6078–6085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang L.; Wheaton D.; Bereta G.; Zhang K.; Palczewski K.; Birch D. G.; Baehr W. (2008) A novel GCAP1(N104K) mutation in EF-hand 3 (EF3) linked to autosomal dominant cone dystrophy. Vision Res. 48, 2425–2432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishiguchi K. M.; Sokal I.; Yang L.; Roychowdhury N.; Palczewski K.; Berson E. L.; Dryja T. P.; Baehr W. (2004) A novel mutation (I143NT) in guanylate cyclase-activating protein 1 (GCAP1) associated with autosomal dominant cone degeneration. Invest. Ophthalmol. Vis. Sci. 45, 3863–3870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sokal I.; Dupps W. J.; Grassi M. A.; Brown J. Jr.; Affatigato L. M.; Roychowdhury N.; Yang L.; Filipek S.; Palczewski K.; Stone E. M.; Baehr W. (2005) A novel GCAP1 missense mutation (L151F) in a large family with autosomal dominant cone-rod dystrophy (adCORD). Invest. Ophthalmol. Vis. Sci. 46, 1124–1132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang L.; Katz B. J.; Yang Z.; Zhao Y.; Faulkner N.; Hu J.; Baird J.; Baehr W.; Creel D. J.; Zhang K. (2005) Autosomal dominant cone dystrophy caused by a novel mutation in the GCAP1 gene (GUCA1A). Mol. Vis. 11, 143–151. [PubMed] [Google Scholar]
- Wilkie S. E.; Li Y.; Deery E. C.; Newbold R. J.; Garibaldir D.; Bateman J. B.; Zhang H.; Lin W.; Zack D. J.; Bhattacharya S. S.; Warren M. J.; Hunt D. M.; Zhang K. (2001) Identification and functional consequences of a new mutation (E155G) in the gene for GCAP1 that causes autosomal dominant cone dystrophy. Am. J. Hum. Genet. 69, 471–480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsu Y.-T.; Molday R. S. (1993) Modulation of the cGMP-gated channel of rod photoreceptor cells by calmodulin. Nature 361, 76–79. [DOI] [PubMed] [Google Scholar]
- Weitz D.; Zoche M.; Müller F.; Beyermann M.; Körschen H. G.; Kaupp U. B.; Koch K.-W. (1998) Calmodulin controls the rod photoreceptor CNG channel through an unconventional binding site in the N-terminus of the beta-subunit. EMBO J. 17, 2273–2284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grunwald M. E.; Yu W.-P.; Yu H.-H.; Yau K.-W. (1998) Identification of a domain on the β-subunit of the rod cGMP-gated cation channel that mediates inhibition by calcium-calmodulin. J. Biol. Chem. 273, 9148–9157. [DOI] [PubMed] [Google Scholar]
- Kawamura S.; Hisatomi O.; Kayada S.; Tokunaga F.; Kuo C.-H. (1993) Recoverin has S-modulin activity in frog rods. J. Biol. Chem. 268, 14579–14582. [PubMed] [Google Scholar]
- Gorodovikova E. N.; Senin I. I.; Philippov P. P. (1994) Calcium-sensitive control of rhodopsin phosphorylation in the reconstituted system consisting of photoreceptor membranes, rhodopsin kinase and recoverin. FEBS Lett. 353, 171–172. [DOI] [PubMed] [Google Scholar]
- Chen C.-K.; Woodruff M. L.; Chen F. S.; Chen D.; Fain G. L. (2010) Background light produces a recoverin-dependent modulation of activated-rhodopsin lifetime in mouse rods. J. Neurosci. 30, 1213–1220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grigoriev I. I.; Senin I. I.; Tikhomirova N. K.; Komolov K. E.; Permyakov S. E.; Zernii E. Y.; Koch K.-W.; Philippov P. P. (2012) Synergetic effect of recoverin and calmodulin on regulation of rhodopsin kinase. Front. Mol. Neurosci. 5, 1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Venkatesan J. K.; Natarajan S.; Schwarz K.; Mayer S. I.; Alpadi K.; Magupalli V. G.; Sung C.-H.; Schmitz F. (2010) Nicotinamide adenine dinucleotide-dependent binding of the neuronal Ca2+ sensor protein GCAP2 to photoreceptor synaptic ribbons. J. Neurosci. 30, 6559–6576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmitz F.; Natarajan S.; Venkatesan J. K.; Wahl S.; Schwarz K.; Grabner C. P. (2012) EF hand-mediated Ca2+- and cGMP signaling in photoreceptor synaptic terminals. Front. Mol. Neurosci. 5, 26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- López-del Hoyo N.; Fazioli L.; López-Begines S.; Fernández-Sánchez L.; Cuenca N.; Llorens J.; de la Villa P.d; Mendez A. (2012) Overexpression of guanylate cyclase activating protein 2 in rod photoreceptors in vivo leads to morphological changes at the synaptic ribbon. PLoS One 7, e42994. [DOI] [PMC free article] [PubMed] [Google Scholar]