

Abstract
Pseudomonas aeruginosa PAO1 has two differentially expressed methionine sulfoxide reductase genes: msrA (PA5018) and msrB (PA2827). The msrA gene is expressed constitutively at a high level throughout all growth phases, whereas msrB expression is highly induced by oxidative stress, such as sodium hypochlorite (NaOCl) treatment. Inactivation of either msrA or msrB or both genes (msrA msrB mutant) rendered the mutants less resistant than the parental PAO1 strain to oxidants such as NaOCl and H2O2. Unexpectedly, msr mutants have disparate resistance patterns when exposed to paraquat, a superoxide generator. The msrA mutant had a higher paraquat resistance level than the msrB mutant, which had a lower paraquat resistance level than the PAO1 strain. The expression levels of msrA showed an inverse correlation with the paraquat resistance level, and this atypical paraquat resistance pattern was not observed with msrB. Virulence testing using a Drosophila melanogaster model revealed that the msrA, msrB, and, to a greater extent, msrA msrB double mutants had an attenuated virulence phenotype. The data indicate that msrA and msrB are essential genes for oxidative stress protection and bacterial virulence. The pattern of expression and mutant phenotypes of P. aeruginosa msrA and msrB differ from previously characterized msr genes from other bacteria. Thus, as highly conserved genes, the msrA and msrB have diverse expression patterns and physiological roles that depend on the environmental niche where the bacteria thrive.
INTRODUCTION
Pseudomonas aeruginosa is a Gram-negative metabolically versatile bacterium commonly found in the environment, including hospitals. This bacterium can grow under nutrient-limited conditions and in stressful environments (1). In health care settings, P. aeruginosa is an opportunistic pathogen responsible for serious bacterial infections in immunocompromised hospitalized patients with cancer, AIDS, cystic fibrosis, and burn injuries. During infection, P. aeruginosa is exposed to toxic levels of reactive oxygen species (ROS) generated by host innate immune cells. In the environment outside the host, P. aeruginosa is exposed to ROS produced by other microbes; moreover, ROS is generated as a by-product of its own aerobic metabolism (2). The ROS can oxidize cellular macromolecules, including proteins and nucleic acids, leading to cell damage and eventual bacterial cell death. To survive oxidative stress, P. aeruginosa has evolved a wide variety of mechanisms to protect itself and to repair the damage caused by ROS toxicity. Several antioxidant enzymes capable of degrading ROS, such as catalases, superoxide dismutases, peroxiredoxins, alkyl hydroperoxide reductases, organic hydroperoxide resistance protein, and thiol peroxidase, have been characterized (2–6). P. aeruginosa also produces a number of enzymes that are required for repairing oxidative DNA damage (7). However, the repair system for oxidized proteins in this microorganism is poorly understood.
Both free and bound methionine (Met) are highly sensitive to oxidation by ROS. In biological systems, methionine oxidation often results in the formation of methionine sulfoxide (MetSO) that could alter protein conformation and/or inactivate protein functions. Methionine sulfoxide can be directly repaired by methionine sulfoxide reductase (Msr), which catalyzes a reverse reaction to convert MetSO to Met. Generally, oxidation of Met produces two S- and R-MetSO diastereoisomers that are referred to as Met-S-SO and Met-R-SO. The reduction of Met-S-SO and Met-R-SO to Met is catalyzed by MsrA and MsrB, respectively. Both MsrA and MsrB can reduce either free or protein-bound MetSO residues (8, 9). Msr catalyzes reduction of MetSO to Met through oxidation of its reactive cysteine residue to form a sulfenic acid intermediate prior to forming an intramolecular disulfide bond between the catalytic Cys and the resolving Cys residues (8, 10). The oxidized enzymes are reactivated using the NADPH/thioredoxin reductase/thioredoxin system (10, 11). Both msrA and msrB are highly conserved and exist in a diverse range of organisms (12, 13). In pathogenic bacteria, a lack of functional msrA not only increases sensitivity to oxidative stress but is also associated with an attenuated virulence phenotype (14–19). Oxidation of bacterial Met to MetSO has been shown to contribute to pathogen killing in neutrophils by the myeloperoxidase (MPO) system, probably via generation of hypochlorous acid (HOCl), a potent oxidant inducing Met oxidation (20).
In this study, the roles of P. aeruginosa msrA (PA5018) and msrB (PA2827) in oxidative stress protection, gene expression profiles, and bacterial virulence were evaluated.
(Adisak Romsang conducted this research in partial fulfillment of the requirements for a Ph.D. from Mahidol University, Bangkok, Thailand.)
MATERIALS AND METHODS
Bacterial strains, plasmids, and growth conditions.
All bacterial strains and plasmids used in this study are listed in Table 1. Both Escherichia coli and Pseudomonas aeruginosa strains were aerobically cultivated in Luria-Bertani (LB) broth at 37°C with shaking at 180 rpm unless otherwise stated. The medium for E. coli growth was supplemented with 100 μg ml−1 ampicillin (Ap), 15 μg ml−1 gentamicin (Gm), 30 μg ml−1 kanamycin (Km), or 15 μg ml−1 tetracycline (Tc) as required, whereas the medium for P. aeruginosa was supplemented with 200 μg ml−1 carbenicillin (Cb), 25 μg ml−1 chloramphenicol (Cm), 30 μg ml−1 Gm, or 200 μg ml−1 Tc as required. To produce synchronous growth, an overnight culture was inoculated into fresh LB medium to give an optical density at 600 nm (OD600) of 0.1. Exponential-phase cells (OD600 of approximately 0.6 after 3 h of growth) were used in all experiments.
Table 1.
Bacterial strains and plasmids used in this study
| Strain or plasmid | Relevant characteristic(s)a | Source or reference |
|---|---|---|
| P. aeruginosa strains | ||
| PAO1 | Wild-type strain | ATCC 15692 |
| msrA strain | msrA mutant, derivative of PAO1 in which msrA was disrupted by pKNOCKmsrA, Gmr | This study |
| msrB strain | msrB mutant, derivative of PAO1 in which a part of msrB was deleted by Cre-loxP system | This study |
| msrA msrB strain | msrA msrB mutant, derivative of msrB in which msrA was disrupted by pKNOCKmsrA, Gmr | This study |
| E. coli strains | ||
| DH5α | λ− ϕ80dlacZΔM15 recA1 endA1 gyrA96 thi-1 hsdR17(rK− mK+) supE44 relA1 deoR Δ(lacZYA-argF)U169 | Stratagene Inc. (USA) |
| BW20767 | leu-63::IS10 recA1 creC510 hsdR17 endA1 zbf-5 uidA(ΔMluI)::pir+ thi RP4-2-tet::Mu-1kan::Tn7 | 60 |
| Plasmids | ||
| pKNOCKGm | Suicide vector, Gmr | 24 |
| pKNOCKmsrA | pKNOCKGm containing a 351-bp fragment of the msrA coding region, Gmr | This study |
| pBBR1MCS-4 | Medium-copy-number expression vector, Apr | 26 |
| pBBRmsrA | Full-length msrA coding region cloned into pBBR1MCS-4, Apr | This study |
| pBBRmsrB | Full-length msrB coding region cloned into pBBR1MCS-4, Apr | This study |
| pUFR047 | Low-copy-number expression vector, Apr | 27 |
| pUFRmsrA | Full-length msrA coding region cloned into pUFR047, Apr | This study |
| pUFRmsrB | Full-length msrB coding region cloned into pUFR047, Apr | This study |
| pCM351 | Vector containing the loxP-Gmr-loxP region, Gmr | 25 |
| pCM157 | Vector containing the Cre-encoding gene, Tcr | 25 |
Gmr, gentamicin resistance; Apr, ampicillin resistance; Tcr, tetracycline resistance.
Molecular techniques.
General molecular techniques were performed using standard protocols (21). Transformation of plasmids into P. aeruginosa strains was carried out using electroporation as previously described (22).
Northern blot analysis.
Total RNA isolation, agarose formaldehyde gel electrophoresis, blotting, and hybridization were performed as previously described (21). For analysis of msrA and msrB expression, 10 μg and 50 μg of purified total RNA were loaded into the gel. Radioactively labeled probes were prepared using [α-32P]dCTP and a DNA labeling bead (Amersham, GE Healthcare). A 351-bp fragment of the msrA coding region and a 193-bp fragment of the msrB coding region used as gene-specific probes were amplified from pBBRmsrA using primers BT3002 (5′-ACAGGTGCTGTTCGGCATGG-3′) and BT3003 (5′-AGTTCGGCCTGGAAGCTCTC-3′) and from pBBRmsrB using primers BT2203 (5′-ATCTATCATTGCACCTGCTG-3′) and BT2204 (5′-GGAAGACGTGCCCCAGGTG-3′).
Quantification of msrA and msrB expression by endpoint RT-PCR and real-time PCR.
Total RNA samples were prepared from PAO1 cultures induced with 1 mM H2O2, 1 mM paraquat (PQ), or 0.2% NaOCl for 15 min that were treated with DNase I (Fermentas, Lithuania) according to the manufacturer's protocol. First-strand cDNA synthesis was accomplished using RevertAid Moloney murine leukemia virus (M-MuLV) reverse transcriptase (Fermentas, Lithuania) with random hexamer primers and total RNA (1 μg). Endpoint reverse transcription-PCR (RT-PCR) was performed to preliminarily determine the expression levels of msrA and msrB using primers BT3002 (5′-ACAGGTGCTGTTCGGCATGG-3′) and BT3003 (5′-AGTTCGGCCTGGAAGCTCTC-3′) for msrA and primers BT2203 (5′-ATCTATCATTGCACCTGCTG-3′) and BT2204 (5′-GGAAGACGTGCCCCAGGTG3′) for msrB and 10 ng of cDNA as a template for 25 cycles, as follows: 95°C for 15 s, 55°C for 15 s, and 72°C for 15 s. The 16S rRNA gene, amplified by RT-PCR using primers BT2781 (5′-GCCCGCACAAGCGGTGGAG-3′) and BT2782 (5′-ACGTCATCCCCACCTTCCT-3′), was used as an internal control to normalize cDNA samples. The PCR products were visualized by 1.8% agarose gel electrophoresis. Real-time PCR experiments were performed as described previously (23). Briefly, 10 ng of cDNA was used as a template in a Kapa SYBR Fast quantitative PCR (kit containing Master Mix 2×; ABI Prism) reaction in an Applied Biosystems StepOnePlus real-time PCR system. All reactions were set up according to the manufacturer's instructions. In this test, the comparative threshold cycle (CT) method is used to determine the relative target quantity in the samples. With the comparative CT method, the StepOne software measures the amplification of the targets, msrA and msrB, and of the endogenous control, 16S rRNA, in the samples under induced conditions and in the reference sample (the uninduced condition). Measurements are normalized using the endogenous control. The cycling parameters used were as follows: enzyme activation at 95°C for 20 s, followed by 40 cycles at 95°C for 3 s and 60°C for 60 s. All real-time PCR experiments were performed in independent triplicate trials.
Construction of P. aeruginosa msrA mutant.
Insertional inactivation of msrA (PA5018) was performed using the pKNOCK suicide plasmid vector (24). The internal DNA fragment of the msrA coding region was PCR amplified with primers BT3002 (5′-ACAGGTGCTGTTCGGCATGG-3′) and BT3003 (5′-AGTTCGGCCTGGAAGCTCTC-3′), using genomic DNA isolated from wild-type PAO1 as the template. The 351-bp PCR product was cloned into pKNOCK-Gm cut with SmaI, generating pKNOCKGmmsrA. This recombinant plasmid was introduced into PAO1 by conjugation. The transconjugants were selected for by the gentamicin resistance (Gmr) phenotype. A single homologous recombination event between the msrA fragment on pKNOCKGmmsrA and its counterpart on the chromosome results in the insertion of the plasmid and inactivation of msrA. The msrA mutant was confirmed by PCR and Southern blot analyses.
Construction of P. aeruginosa msrB mutant.
The unmarked deletion of PAO1 msrB was constructed using gene replacement with the Cre-loxP system (25). A 1,453-bp fragment covering the entire msrB gene and the 5′ and 3′ flanking regions was amplified with the primers BT1187 (5′-CCTGGATCCGCTGGTAATACTGCAT-3′) and BT190 (5′-GGTGCTCATTCAAGCAGCG-3′) and cloned into pUC18 digested with EcoRI-BamHI (blunted with Klenow fragment), yielding pUCmsrB. A KpnI and SacII fragment containing a Gmr cassette flanked with loxP sites from pCM351 (25) was cloned into pUCmsrB cut with KpnI and SacII, yielding pUCΔmsrB::Gm. Digestion of pUCmsrB with KpnI and SacII resulted in a deletion of 305 bp of the msrB coding region. The pUCΔmsrB::Gm was transferred into PAO1, and the transformants were selected for a double crossover event by means of the Gmr and Cb-sensitive phenotypes. To obtain an unmarked msrB mutant, a Cre recombinase expression plasmid, pCM157 (25), was transferred into the transformant, and Cre-loxP recombination facilitated the excision of the Gmr cassette. The pCM157 was cured by growing cells under a nonselective condition for several generations. The deletion of msrB was confirmed by PCR and Southern blot analyses.
Construction of a double msrA msrB mutant.
The msrA msrB mutant was constructed by the transfer of pKNOCKGmmsrA into the ΔmsrB mutant and selected by the Gm resistance phenotype. The double msrA msrB mutant was confirmed by PCR and Southern blot analyses.
Cloning of full-length msr genes.
Full-length msrA and msrB genes were PCR amplified from PAO1 genomic DNA using the primers BT3004 (5′-CGCGAGTCATTCGCCTACC-3′) and BT3005 (5′-CTCGACTGGCTGGTGGCG-3′) for msrA and the primers BT2201 (5′-CGAGGTAGAGCCGATGAG-3′) and BT2202 (5′-CGGGTCGTCGCTCTCTAGCT-3′) for msrB. The PCR products were cloned into the medium-copy-number expression vector pBBR1MCS-4 (26) and the low-copy-number expression vector pUFR047 (27) at the SmaI site. The resulting plasmids were named pBBRmsrA, pBBRmsrB, pUFRmsrA, and pUFRmsrB.
Viability test with H2O2 and NaOCl exposures.
Exponential-phase cultures were normalized to an OD600 of 0.5 before being treated with 100 mM H2O2 or 0.3% NaOCl for 30 min at 37°C. After treatment, cells were washed twice with fresh LB broth. Cells that survived the treatment were scored using a viable cell count. The resistance levels against H2O2 or NaOCl were expressed as percent survival, defined as the percentage of CFU in treated cultures divided by the percentage of CFU in untreated cultures.
Determination of oxidant resistance levels.
A plate sensitivity assay was performed to determine the oxidant resistance level as previously described (28). Briefly, exponential-phase cells were 10-fold serially diluted in fresh LB medium, and 10 μl of each dilution was spotted onto LB plates containing 500 μM hydrogen peroxide (H2O2), 1,600 μM cumene hydroperoxide (CuOOH), 300 μM N-ethylmaleimide (NEM), or 175 μM paraquat (PQ). The plates were incubated at 37°C for 18 h before the numbers of CFU were scored. The resistance level against an oxidant was expressed as the percent survival, defined as the percentage of CFU on the plates containing oxidant divided by the percentage of CFU on plates without oxidant. For anaerobic conditions, the culture plates were incubated in a 3.5-liter anaerobic jar containing an anaerobic gas pack (AN35; AnaeroGen, Oxoid, United Kingdom) and an anaerobic indicator and placed in a 37°C incubator for 48 h before the CFU were scored.
SOD activity assay.
Xanthine-xanthine oxidase-coupled reduction of cytochrome c was used to measure total superoxide dismutase (SOD) activity (29). One unit of SOD activity was defined as the amount of enzyme required to inhibit the rate of reduction of cytochrome c by 50%.
Drosophila virulence tests.
The virulence of P. aeruginosa was investigated using the Drosophila melanogaster feeding assay performed as previously described (30, 31). Each of the exponential-phase cultures of each P. aeruginosa strain was adjusted to an OD600 of 0.5 before 800 μl of the homogenized bacterial cells was overlaid to completely cover the surface of the preservative-free corn flour Drosophila medium (500 ml of water, 62 g of corn flour, 8 g of yeast extract, 40 g of sugar, and 4 g of agar) at the bottom of a glass fly culture vial. Approximately 1-week-old adult flies were starved for 3 h prior to the feeding assay. Twenty flies each were added to every vial, and each strain of P. aeruginosa was tested at least for three replications. Then, all of the tested flies were incubated at 25°C for 15 h before the number of viable flies was observed. The experiments were performed in double-blind fashion and analyzed from nine experiments of three different batches of flies.
RESULTS AND DISCUSSION
Identification and transcription organization of msr genes in P. aeruginosa.
The P. aeruginosa PAO1 genome (32) contains PA5018, a 648-bp open reading frame (ORF) annotated as msrA encoding the peptide methionine sulfoxide reductase A. The deduced amino acid sequence of P. aeruginosa MsrA shares 58.5%, 56.7%, and 56.3% identity with MsrA from E. coli (33), bovine (Bos taurus) (34), and mouse (Mus musculus) (35), respectively (Fig. 1A). Lower percent identities (27.7% to 34.3%) between P. aeruginosa MsrA and MsrA from Erwinia pyrifoliae, Neisseria meningitidis, and Saccharomyces cerevisiae were observed (Fig. 1A). The MsrA signature motif GCFWG (8, 34) and a thioredoxin binding domain at the C terminus (36) are conserved in P. aeruginosa MsrA. The msrA gene is located 93 bp downstream of PA5017, a hypothetical protein. Analysis of transcriptional organization of these genes by Northern blotting and RT-PCR using primers located on the PA5017 and msrA genes indicated that they are separately transcribed (Fig. 2A and data not shown). A 399-bp PA2827 annotated as methionine sulfoxide reductase B (msrB) shares 70.0% identity with MsrB from E. coli (Fig. 1B). The highly conserved catalytic motif RXCXN of MsrB (36, 37) is also conserved in PAO1 MsrB. The msrB gene is located immediately upstream of a putative operon consisting of the PA2826 gene, a glutathione peroxidase homolog (gpx) and the PA2825 gene, a transcription regulator (ospR) (38). Northern blot analyses of msrA and msrB showed that these genes are transcribed mostly as a monocistronic transcript (Fig. 2A).
Fig 1.

Genetic organization and multiple amino acid sequence alignments of P. aeruginosa MsrA and MsrB. Genetic organization and alignments of MsrA (A) and MsrB (B) from P. aeruginosa with other characterized enzymes from various organisms were performed using the CLUSTALW program. The arrow direction indicates the transcription orientation. Dark gray boxes indicate the MsrA or MsrB signature motif. The asterisk, colon, and period symbols indicate identical residues, conserved substitutions, and semiconserved substitutions, respectively. Values at right are percent identities of the aligned proteins with the P. aeruginosa protein. gpx, glutathione peroxidase homolog (PA2826); and ospR, oxidative stress response and pigment production regulator.
Fig 2.

Expression analysis of msrA and msrB during normal growth and in response to oxidants. (A) Autoradiograms of Northern blots of uninduced and 0.2% NaOCl-induced samples probed with either radioactively labeled msrA or msrB. Equal amounts of purified total RNA from uninduced (UN) and 0.2% NaOCl induced (IN) samples were loaded into each lane (10 μg for msrA and 50 μg for msrB). The number below each band represents the fold change in band intensity relative to the level of the uninduced culture, determined using densitometric analysis. (B) Real-time RT-PCR analysis of msrA and msrB expression. Total RNA was isolated from uninduced exponential-phase cultures. Real-time RT-PCR and subsequent analysis were performed on equal amounts of cDNA (10 ng) prepared from total RNA samples as described in Materials and Methods. Data shown are fold change of expression levels using msrB as the reference sample. (C) Real-time RT-PCR analysis of the expression levels of P. aeruginosa msrA and msrB in cultures induced with either 1 mM H2O2, 1 mM PQ, or 0.2% NaOCl. Oxidant induction, RNA isolation, real-time RT-PCR, and subsequent analyses were performed as described in Materials and Methods. Data presented are means ± standard deviations of three independent experiments. The asterisk indicates statistically significant difference (P < 0.01) compared with the uninduced condition.
The expression patterns of msrA and msrB in response to oxidative stress.
There is a clear need to comprehensively analyze the patterns of gene expression of both msrA and msrB during standard growth phases and in response to stresses. Currently, analyses of bacterial msrA and msrB show highly diverse patterns of gene expression in terms of growth phase and stress response. Bacteria in different growth phases confront different levels of oxidative stress resulting from physiologically generated ROS from aerobic metabolism. The msr genes in some bacteria have been shown to have growth phase-dependent expression (15, 33). Endpoint RT-PCR was performed to determine the levels of msrA and msrB at early log, mid-log, early stationary, and late stationary phases of growth of PAO1 cultures. The levels of msrA and msrB transcripts were not significantly altered during the different growth phases tested (data not shown).
Adaptive gene expression is an important component of bacterial defense against stresses. The expression of many genes involved in oxidative stress protection and repair processes are frequently induced by exposure to oxidants. The expression patterns of msrA and msrB in the PAO1 strain cultivated under inducing concentrations of various oxidants were determined using real-time PCR. The results illustrated that exposure of PAO1 to H2O2 (1 mM), paraquat (1 mM), and sodium hypochlorite (NaOCl; 0.2%) did not induce expression of msrA (Fig. 2C). Similarly, no significant induction of msrB expression was observed in paraquat- and H2O2-treated samples. In contrast, NaOCl treatment of PAO1 highly induced msrB expression by 17.6-fold (Fig. 2C). PAO1 cultures were induced with H2O2 at concentrations ranging from 250 μM to 10 mM. H2O2 treatment has been shown to oxidize free and bound Met, leading to the formation of both S and R epimers of MetSO. In many bacteria H2O2 is reported to be a potent inducer of msr expression (39–41). At all H2O2 concentrations tested, no induction of either msrA or msrB expression was observed (data not shown). Hence, lack of H2O2 induction of msrA and msrB expression could be due partly to the highly efficient H2O2 removal systems of PAO1 coupled with the uninduced levels of msrA and msrB that are most likely sufficient to repair H2O2 oxidation of Met in both protein and free forms.
The pattern of oxidants capable of inducing msrB and the oxidant concentration required for induction do not match any known oxidative stress sensor/transcriptional regulators in P. aeruginosa. We tested the NaOCl induction of msrB in mutants of transcription regulators involved in oxidative stress response, such as oxyR, ohrR, ospR, and iscR. The results showed that the fold induction by NaOCl of msrB expression in these mutants was induced to levels similar to the level attained in PAO1 (data not shown).
Recently, the hypochlorite-specific transcriptional regulator YjiE has been reported (42). A mutant of a yjiE homolog (PA5428) was constructed, and NaOCl induction of msrB was tested (S. Mongkolsuk, K. Saninjuk, and W. Wirathorn, unpublished data). The PA5428 mutant gave a similar level of NaOCl-induced msrB expression as a similarly treated wild-type PAO1, indicating that PA5428 (yjiE homolog) played no role in regulation of hypochlorite-induced msrB expression (data not shown). Moreover, sigma factor-mediated regulation of msr has been reported; for example, msrA and msrAB are members of σF and σE regulons in Caulobacter crescentus and N. meningitidis, respectively (43, 44). Further experiments are required to investigate the contribution of sigma factors in msrA gene regulation in P. aeruginosa. Thus, at present, it is not known which transcriptional regulator controls msrB expression in P. aeruginosa. In a few bacteria where oxidant-induced expression of msr genes has been characterized, the transcriptional regulator responsible for this adaptation has not been identified (15, 41, 45).
MsrA and MsrB recognize the S and R forms of MetSO and catalyze their conversion to Met. However, MsrA has much higher efficiency in repairing oxidation of free Met than MsrB (46). Under normal physiological growth conditions, the basal level of msrA transcripts was 17.2-fold higher than the level of msrB transcripts (Fig. 2B). This indicates that MsrA is present at relatively high concentrations throughout various growth phases and during exposure to oxidants, ensuring that it is ready to repair both free and bound MetSO formation upon oxidant exposure. In contrast, msrB expression is maintained at a low level under normal physiological conditions. Once the bacteria are exposed to oxidants such as NaOCl from external sources, rapid induction (over 17.6-fold) of msrB expression occurs (Fig. 2C). NaOCl is a strong oxidant, and in aqueous solutions, NaOCl ionizes to generate Na+ and hypochlorite (OCl−), which rapidly forms hypochlorous acid (HOCl). Hence, NaOCl and derivatives could directly react with various macromolecules. Moreover, as a secondary effect, NaOCl treatment also generates arrays of ROS (47, 48).
Mutations in msrA and msrB altered resistance levels to NaOCl and H2O2.
HOCl is an important component of the host-generated immune response against bacterial infection. Thus, during their interactions with the host, bacteria are exposed to harmful levels of highly reactive HOCl. Recently, genes involved in protection from oxidative stress have been implicated in the role of protecting bacteria from NaOCl toxicity (39, 49, 50). In this study, experiments were performed to elucidate the role of P. aeruginosa msr genes in protecting the bacteria from NaOCl toxicity. Specifically, resistance levels to NaOCl of PAO1 and various msr mutants were assessed. The results showed that msrA, msrB, and msrA msrB mutants were 10-, 102-, and 103-fold more sensitive to 0.3% NaOCl treatment, respectively, than the PAO1 parental strain. The difference in the reduction of NaOCl resistance levels in the single msrA and msrB mutants and the additive effects of reduction in NaOCl resistance levels in the double mutant indicate that msrA and msrB have functionally different capacities in protection against NaOCl toxicity. Next, the ability of msrA or msrB in a medium-copy-number expression vector (pBBR1MCS-4) to complement the reduced NaOCl resistance phenotype in PAO1 and the mutant strains was determined. First, PAO1 strains harboring either pBBRmsrA or pBBRmsrB have similar resistance levels to NaOCl as PAO1 harboring a vector control. This result indicates that high expression levels of either msrA or msrB do not confer additional resistance to NaOCl. Second, pBBRmsrA and pBBRmsrB plasmids fully complemented the NaOCl-sensitive phenotypes of msrA and msrB mutants to resistance levels attained by the parental strain, supporting the idea that the observed phenotype of the mutants was due to a lack of either MsrA or MsrB. Because MsrA and MsrB have different substrate ranges and biochemical properties, the experiments were extended to test cross-complementation using either pBBRmsrA or pBBRmsrB in the single mutants. The results showed that pBBRmsrB partially complemented the NaOCl-sensitive phenotype of the msrA mutant to a level that was 5-fold lower than that of PAO1, while pBBRmsrA could also partially complement the phenotype of the msrB mutant to a level 10-fold lower than that of the wild-type strain (Fig. 3). In addition, the ectopic expression of either pBBRmsrA or pBBRmsrB also partially complemented the NaOCl resistance levels of the double msrA msrB mutant to a resistance level 10-fold lower than that of PAO1 (Fig. 3).
Fig 3.

Determination of H2O2 and NaOCl resistance levels. The resistance levels of P. aeruginosa strains against 100 mM H2O2 or 0.3% NaOCl were determined using a viability test performed as described in Materials and Methods. The percent survival was defined as the percentage of the CFU with treatment divided by the percentage of CFU without treatment. Data presented are means ± standard deviations of three independent experiments. The asterisk indicates a statistically significant difference (P < 0.01) compared with the PAO1 strain.
Exposure of bacteria to NaOCl results in the generation of many types of ROS (47, 48, 51), which could oxidize both free and bound Met residues in proteins, leading to a loss of biological functions. One could assume that, upon Met oxidation, S and R epimers of MetSO are formed at equal efficiencies, and the repair capacity would reside in the quantity and efficiency of MsrA and MsrB. The expression of either msrA or msrB from an expression vector did not confer additional resistance to NaOCl upon wild-type PAO1 strains, supporting the notion that NaOCl treatment produced other forms of cellular damage in addition to oxidation of Met. The mutational analysis revealed that both msrA and msrB have differential capacities to reduce free or peptidyl MetSO in the repair of oxidized methionine resulting from the NaOCl treatment. The ability of ectopic expression of msrA and msrB to partially cross-complement the NaOCl-sensitive phenotypes of the msrA, msrB, and double msrA msrB mutants supports the separate roles the two enzymes have in the overall protection of Met from oxidants. The partial cross-complementation phenotype could arise from the ability of each enzyme to functionally substitute for the lack of either MsrA or MsrB activities or the interconversion between S and R epimers of MetSO.
P. aeruginosa is likely to encounter toxic levels of H2O2 during interaction with hosts and in the environment. H2O2 can oxidize both free and bound Met (52). Genetic analyses in other bacteria have shown that inactivation of either msrA or msrB renders the mutants less resistant to H2O2 treatment (45, 53). The resistance levels to H2O2 in PAO1 and various msr mutants were determined. Unexpectedly, the single mutants msrA and msrB did not show significant alteration in H2O2 (100 mM) resistance levels under the conditions tested. Nonetheless, the H2O2 resistance level of the msrA msrB double mutant was 8-fold more sensitive to H2O2 treatment than PAO1. This small but reproducible reduction in the H2O2 resistance level of the double mutant could be fully complemented by either pBBRmsrA or pBBRmsrB. The lack of H2O2 phenotypes in single msr mutants and a small increase in H2O2 sensitivity in the msrA msrB mutant are both likely due to the presence of numerous peroxide-scavenging enzymes, such as catalases, peroxiredoxins, and alkyl hydroperoxide reductases, which efficiently removed H2O2 to mitigate the peroxide oxidation of Met. A minor susceptibility to H2O2 could be observed only when methionine sulfoxide reductases were totally deficient. The data support a minor role for msrA and msrB in protecting the cell from H2O2-induced death. This is consistent with the strategy used by many bacteria in that oxidant removal has a primary role over the repair of oxidatively damaged macromolecules. Once the oxidant removal capacity is exceeded and oxidation of macromolecules begins, repair of the oxidatively damaged molecules becomes a major component of the oxidant removal systems that protect the bacteria.
No significant alteration in resistance levels was observed in the msrA, msrB, and msrA msrB mutants when the strains were treated with 1.6 mM CuOOH and 300 μM NEM.
Inactivation of either msrA or msrB disparately alters the mutants' ability to cope with superoxide stress.
Superoxide anion-mediated oxidation of Met residues in proteins and free Met has been reported (53, 54). In other bacteria, inactivation of msr genes has been shown to increase sensitivity to the superoxide generator paraquat (53); therefore, the physiological roles of P. aeruginosa msrA and msrB in superoxide stress protection were evaluated. The resistance levels of P. aeruginosa strains against paraquat were determined. Unexpectedly, the msrA mutant was 70-fold more resistant to paraquat (175 μM) killing than the PAO1 parental strain (Fig. 4). In contrast, an msrB mutant was 102-fold more sensitive to paraquat than a parental strain (Fig. 4). The resistance level of the double msrA msrB mutant was equivalent to the sum of the resistance levels of the msrA and msrB mutants and comparable to the level of PAO1. These findings support a notion that the altered paraquat phenotypes of msrA and msrB mutants arise from disparate pathways. A recent observation indicated that paraquat toxicity to bacteria results either directly from the redox cycling property of the drug or from superoxide anions generated as by-products of the reaction (55). We extended the investigation to determine the probable mode of paraquat killing of various msr mutants by performing a paraquat plate sensitivity assay under aerobic and anaerobic conditions using LB medium supplemented with 1% nitrate. The PAO1, msrA, msrB, and double mutant strains had similar patterns of paraquat resistance under aerobic growth conditions, performed using either LB or LB medium plus nitrate. In contrast, under anaerobic growth conditions, the addition of 175 μM paraquat to LB medium plus nitrate did not significantly reduce the percent survival of either wild-type PAO1 or msr mutants (Fig. 5). Moreover, when a higher paraquat concentration (500 μM) was used in the anaerobic plate sensitivity assay, no reduction in the survival of PAO1 and mutant strains was detected (Fig. 5). Clearly, paraquat-induced killing of P. aeruginosa required aerobic conditions; hence, the toxicity likely arises from superoxide anions generated by the drug. Paraquat resistance levels have been known to correlate with the ability of bacterial cells to remove superoxide anions and superoxide dismutase (SOD) activity. Total SOD activity was measured in the msrA, msrB, and double mutants, all grown aerobically. The results showed nonsignificant differences in the levels of total SOD activity in the wild-type and mutant strains tested (data not shown). Therefore, alterations in levels of paraquat resistance of the msrA and msrB mutants are independent of total SOD enzyme activity.
Fig 4.

Determination of paraquat resistance levels. The resistance levels of P. aeruginosa strains against 0.175 mM paraquat were determined using a plate sensitivity assay as described in Materials and Methods and are expressed as percent survival. The percent survival was defined as the percentage of CFU on plates containing paraquat divided by the percentage of CFU on plates without paraquat. Data presented are means ± standard deviations of three independent experiments. The asterisk indicates a statistically significant difference (P < 0.01) compared with the PAO1 harboring pBBR1MCS-4 or the pUFR047 vector control.
Fig 5.

Determination of paraquat resistance levels under aerobic and anaerobic conditions. A plate sensitivity assay was performed using LB medium with and without supplementation of 1% KNO3 and incubated under aerobic and anaerobic conditions. The concentrations of paraquat were 0.175 mM for aerobic experiments and 0.5 mM for anaerobic experiments. The resistance levels of P. aeruginosa strains against paraquat were determined using a plate sensitivity assay and are expressed as percent survival as defined in the legend of Fig. 4. Data presented are means ± standard deviations of three independent experiments. The asterisk indicates a statistically significant difference (P < 0.01) compared with PAO1.
We extended the investigation by examining the ability of msrA and msrB to complement and cross-complement the paraquat resistance phenotypes of the msrA and msrB mutants. Paraquat resistance levels of PAO1, msrA, msrB, and the double msrA msrB mutant strains harboring either pBBRmsrA, pBBRmsrB, or a vector control were measured. Unexpectedly, high levels of expression of msrA from pBBRmsrA reduced the percentage of survival after paraquat treatment by 103-fold for the PAO1 and msrA strains and by more than 10-fold for the msrB and msrA msrB mutants relative to strains harboring the vector control (Fig. 4). We examined the effect of lowering msrA expression levels using pUFR047 (a low-copy-number expression vector; 2 to 4 copies) instead of pBBR1MCS (a medium-copy-number vector; 30 to 40 copies) (56). Decreasing the copy number of the expression vectors, and hence reducing msrA expression levels, in strains harboring pUFRmsrA resulted in a 102-fold increase in percent survival of PAO1 and the msrA mutant. However, in the msrB strain and the double msrA msrB mutant, there was a 50-fold increase in the percent survival with paraquat treatment compared to the strains harboring a medium-copy-number vector (Fig. 4). The results from expression analysis of pBBRmsrA and pUFRmsrA clearly reveal a correlation between the levels of msrA expression and the magnitude of reduction in the paraquat resistance levels in all strains tested, regardless of the host genetic background. The effects of msrB expression in PAO1 and the three msr mutant strains were also examined. The expression of msrB in the msrA mutant lowered its paraquat resistance level by 8-fold, whereas both pBBRmsrB and pUFRmsrB could fully complement the paraquat-sensitive phenotype of the msrB mutant to levels similar to the level of PAO1. There were no alterations in the paraquat resistance phenotype associated with msrB expression levels in PAO1 and all mutant strains harboring either pBBRmsrB or pUFRmsrB.
The phenotypes of paraquat resistance of the msrA mutant and strains with overexpression of msrA have not been observed in other bacteria and are highly unexpected. Theoretically, high expression of a gene encoding a repair enzyme for oxidized Met such as MsrA should enhance the resistance level against oxidant treatments. In contrast, the results here showed that the level of MsrA correlated with a reduction in the paraquat resistance level, as illustrated by two independent observations. First, inactivation of msrA resulted in an increase in the paraquat resistance level; second, a high expression level of msrA from the expression vectors decreased the paraquat resistance level. These observations suggest that MsrA possesses other enzymatic activity besides the methionine sulfoxide reductase function that repairs free and bound MetSO. A recent report that MsrA is also a stereospecific methionine oxidase, producing Met-S-SO, raises the possibility that P. aeruginosa MsrA also has additional oxidase-like activity (57). Similar to findings of a previous report (57), the depletion of thioredoxin by paraquat treatment could lead to enhanced methionine oxidase activity and the formation of MetSO in target proteins that render the bacteria more susceptible to paraquat killing.
msrA and msrB mutants are attenuated for virulence in the Drosophila host model.
The role of msr genes in virulence and pathogenicity varied widely in different bacteria. Nonetheless, msr genes have important roles in virulence and pathogenesis in many bacteria. Hence, the virulence of the PAO1, msrA, msrB, and double msrA msrB mutant strains was tested using Drosophila melanogaster as a host model. As shown in Fig. 6, feeding the flies with PAO1 cultures resulted in 25.0% ± 5.0% fly survival, compared with 95.0% ± 2.6% fly survival when LB medium was fed as a negative control. When either an msrA or msrB mutant was used to feed D. melanogaster, the fly survival increased to 64.5% ± 6.1% or 61.1% ± 8.0%, respectively. Moreover, the percent survival of the fly was further increased to 71.7% ± 4.1% when the double msrA msrB mutant was used in the feeding experiment. The significant increase in the percent survival of Drosophila flies ranging from 40 to 50% after feeding with the msrA, msrB, and the msrA msrB double mutant strains compared to levels with PAO1 indicated that these mutant strains have an attenuated virulence phenotype in the test model (P < 0.01 by Student t test). This phenotype in both the msrA and msrB mutants could be rescued in the complemented strains expressing functional a copy of each gene (Fig. 6). The attenuated virulence phenotype of the double msrA msrB mutant could only be partially complemented by expression of either msrA or msrB from expression vectors (Fig. 6). These data suggest that msrA and msrB are virulence determinants for P. aeruginosa in the D. melanogaster model, and they indicate the involvement of MsrA and MsrB in P. aeruginosa virulence.
Fig 6.
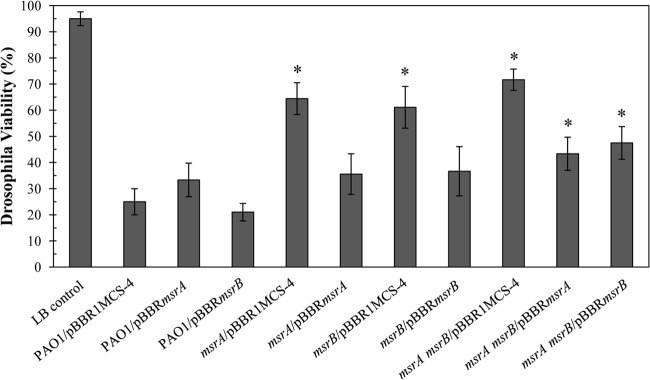
Virulence assay of P. aeruginosa strains. The virulence of P. aeruginosa strains was determined using the Drosophila melanogaster feeding method (30, 31). The flies that survived were counted after 15 h of incubation, and results are expressed as the percent viability. The percent viability was defined as the percentage of the surviving flies after 15 h of incubation with P. aeruginosa cell cultures. Data presented are means ± standard deviations of three independent experiments. The asterisk indicates a statistically significant difference (P < 0.01) compared with PAO1.
The roles of methionine sulfoxide reductases in virulence have been observed in other pathogenic bacteria; for example, Salmonella enterica serovar Typhimurium MsrA, but not MsrB, has an important role in oxidative stress protection and in virulence (58). MsrA has been described as a virulence determinant in various plant- and human-pathogenic bacteria such as Erwinia chrysanthemi (16), Staphylococcus aureus (17), Mycobacterium smegmatis (18), and Mycoplasma genitalium (19). Recently, msrB has been shown to have a role in virulence in Enterococcus faecalis (59).
Loss of either msrA or msrB in P. aeruginosa attenuated its ability to cause death of the flies. The greater attenuated virulence of the msrA msrB mutant confirmed that the two genes functioned in different pathways in protecting the bacteria from host defense responses (Fig. 6). We speculated that the phenotypic difference between the msrA and msrB mutants could be due to these genes likely having roles in different virulence pathways during the bacterial infection, which could arise from different expression patterns of the two msr genes and their different enzymatic activities.
ACKNOWLEDGMENTS
This work was supported by grants from the Chulabhorn Research Institute, Center of Excellence on Environmental Health and Toxicology, Ministry of Education, Thailand, and from Mahidol University. A.R. was supported by a Royal Golden Jubilee Scholarship (PHD/0294/2550) from the Thailand Research Fund.
We thank Tatsanee Chuchue for technical assistance.
Footnotes
Published ahead of print 17 May 2013
REFERENCES
- 1.Grundmann H, Kropec A, Hartung D, Berner R, Daschner F. 1993. Pseudomonas aeruginosa in a neonatal intensive care unit: reservoirs and ecology of the nosocomial pathogen. J. Infect. Dis. 168:943–947 [DOI] [PubMed] [Google Scholar]
- 2.Hassett DJ, Schweizer HP, Ohman DE. 1995. Pseudomonas aeruginosa sodA and sodB mutants defective in manganese- and iron-cofactored superoxide dismutase activity demonstrate the importance of the iron-cofactored form in aerobic metabolism. J. Bacteriol. 177:6330–6337 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Baysse C, De Vos D, Naudet Y, Vandermonde A, Ochsner U, Meyer JM, Budzikiewicz H, Schafer M, Fuchs R, Cornelis P. 2000. Vanadium interferes with siderophore-mediated iron uptake in Pseudomonas aeruginosa. Microbiology 146:2425–2434 [DOI] [PubMed] [Google Scholar]
- 4.An BC, Lee SS, Lee EM, Lee JT, Wi SG, Jung HS, Park W, Chung BY. 2010. A new antioxidant with dual functions as a peroxidase and chaperone in Pseudomonas aeruginosa. Mol. Cells 29:145–151 [DOI] [PubMed] [Google Scholar]
- 5.Ochsner UA, Hassett DJ, Vasil ML. 2001. Genetic and physiological characterization of ohr, encoding a protein involved in organic hydroperoxide resistance in Pseudomonas aeruginosa. J. Bacteriol. 183:773–778 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Somprasong N, Jittawuttipoka T, Duang-Nkern J, Romsang A, Chaiyen P, Schweizer HP, Vattanaviboon P, Mongkolsuk S. 2012. Pseudomonas aeruginosa thiol peroxidase protects against hydrogen peroxide toxicity and displays atypical patterns of gene regulation. J. Bacteriol. 194:3904–3912 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ochsner UA, Vasil ML, Alsabbagh E, Parvatiyar K, Hassett DJ. 2000. Role of the Pseudomonas aeruginosa oxyR-recG operon in oxidative stress defense and DNA repair: OxyR-dependent regulation of katB-ankB, ahpB, and ahpC-ahpF. J. Bacteriol. 182:4533–4544 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Moskovitz J, Poston JM, Berlett BS, Nosworthy NJ, Szczepanowski R, Stadtman ER. 2000. Identification and characterization of a putative active site for peptide methionine sulfoxide reductase (MsrA) and its substrate stereospecificity. J. Biol. Chem. 275:14167–14172 [DOI] [PubMed] [Google Scholar]
- 9.Grimaud R, Ezraty B, Mitchell JK, Lafitte D, Briand C, Derrick PJ, Barras F. 2001. Repair of oxidized proteins. Identification of a new methionine sulfoxide reductase. J. Biol. Chem. 276:48915–48920 [DOI] [PubMed] [Google Scholar]
- 10.Boschi-Muller S, Gand A, Branlant G. 2008. The methionine sulfoxide reductases: catalysis and substrate specificities. Arch. Biochem. Biophys. 474:266–273 [DOI] [PubMed] [Google Scholar]
- 11.Moskovitz J, Weissbach H, Brot N. 1996. Cloning the expression of a mammalian gene involved in the reduction of methionine sulfoxide residues in proteins. Proc. Natl. Acad. Sci. U. S. A. 93:2095–2099 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Rodrigo MJ, Moskovitz J, Salamini F, Bartels D. 2002. Reverse genetic approaches in plants and yeast suggest a role for novel, evolutionarily conserved, selenoprotein-related genes in oxidative stress defense. Mol. Genet. Genomics 267:613–621 [DOI] [PubMed] [Google Scholar]
- 13.Kryukov GV, Kumar RA, Koc A, Sun Z, Gladyshev VN. 2002. Selenoprotein R is a zinc-containing stereo-specific methionine sulfoxide reductase. Proc. Natl. Acad. Sci. U. S. A. 99:4245–4250 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wizemann TM, Moskovitz J, Pearce BJ, Cundell D, Arvidson CG, So M, Weissbach H, Brot N, Masure HR. 1996. Peptide methionine sulfoxide reductase contributes to the maintenance of adhesins in three major pathogens. Proc. Natl. Acad. Sci. U. S. A. 93:7985–7990 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ezraty B, Aussel L, Barras F. 2005. Methionine sulfoxide reductases in prokaryotes. Biochim. Biophys. Acta 1703:221–229 [DOI] [PubMed] [Google Scholar]
- 16.Hassouni ME, Chambost JP, Expert D, Van Gijsegem F, Barras F. 1999. The minimal gene set member msrA, encoding peptide methionine sulfoxide reductase, is a virulence determinant of the plant pathogen Erwinia chrysanthemi. Proc. Natl. Acad. Sci. U. S. A. 96:887–892 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Singh VK, Moskovitz J. 2003. Multiple methionine sulfoxide reductase genes in Staphylococcus aureus: expression of activity and roles in tolerance of oxidative stress. Microbiology 149:2739–2747 [DOI] [PubMed] [Google Scholar]
- 18.Douglas T, Daniel DS, Parida BK, Jagannath C, Dhandayuthapani S. 2004. Methionine sulfoxide reductase A (MsrA) deficiency affects the survival of Mycobacterium smegmatis within macrophages. J. Bacteriol. 186:3590–3598 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Dhandayuthapani S, Blaylock MW, Bebear CM, Rasmussen WG, Baseman JB. 2001. Peptide methionine sulfoxide reductase (MsrA) is a virulence determinant in Mycoplasma genitalium. J. Bacteriol. 183:5645–5650 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Rosen H, Klebanoff SJ, Wang Y, Brot N, Heinecke JW, Fu X. 2009. Methionine oxidation contributes to bacterial killing by the myeloperoxidase system of neutrophils. Proc. Natl. Acad. Sci. U. S. A. 106:18686–18691 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Sambrook J, Russell DW. 2001. Molecular cloning: a laboratory manual, 3rd ed Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY [Google Scholar]
- 22.Choi KH, Schweizer HP. 2006. Mini-Tn7 insertion in bacteria with single attTn7 sites: example Pseudomonas aeruginosa. Nat. Protoc. 1:153–161 [DOI] [PubMed] [Google Scholar]
- 23.Buranajitpakorn S, Piwkam A, Charoenlap N, Vattanaviboon P, Mongkolsuk S. 2011. Genes for hydrogen peroxide detoxification and adaptation contribute to protection against heat shock in Xanthomonas campestris pv. campestris. FEMS Microbiol. Lett. 317:60–66 [DOI] [PubMed] [Google Scholar]
- 24.Alexeyev MF. 1999. The pKNOCK series of broad-host-range mobilizable suicide vectors for gene knockout and targeted DNA insertion into the chromosome of gram-negative bacteria. Biotechniques 26:824–826, 828 [DOI] [PubMed] [Google Scholar]
- 25.Marx CJ, Lidstrom ME. 2002. Broad-host-range cre-lox system for antibiotic marker recycling in gram-negative bacteria. Biotechniques 33:1062–1067 [DOI] [PubMed] [Google Scholar]
- 26.Kovach ME, Elzer PH, Hill DS, Robertson GT, Farris MA, Roop RMII, Peterson KM. 1995. Four new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene 166:175–176 [DOI] [PubMed] [Google Scholar]
- 27.DeFeyter R, Kado CI, Gabriel DW. 1990. Small, stable shuttle vectors for use in Xanthomonas. Gene 88:65–72 [DOI] [PubMed] [Google Scholar]
- 28.Prapagdee B, Vattanaviboon P, Mongkolsuk S. 2004. The role of a bifunctional catalase-peroxidase KatA in protection of Agrobacterium tumefaciens from menadione toxicity. FEMS Microbiol. Lett. 232:217–223 [DOI] [PubMed] [Google Scholar]
- 29.McCord JM, Fridovich I. 1969. Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein). J. Biol. Chem. 244:6049–6055 [PubMed] [Google Scholar]
- 30.D'Argenio DA, Gallagher LA, Berg CA, Manoil C. 2001. Drosophila as a model host for Pseudomonas aeruginosa infection. J. Bacteriol. 183:1466–1471 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Apidianakis Y, Rahme LG. 2009. Drosophila melanogaster as a model host for studying Pseudomonas aeruginosa infection. Nat. Protoc. 4:1285–1294 [DOI] [PubMed] [Google Scholar]
- 32.Stover CK, Pham XQ, Erwin AL, Mizoguchi SD, Warrener P, Hickey MJ, Brinkman FS, Hufnagle WO, Kowalik DJ, Lagrou M, Garber RL, Goltry L, Tolentino E, Westbrock-Wadman S, Yuan Y, Brody LL, Coulter SN, Folger KR, Kas A, Larbig K, Lim R, Smith K, Spencer D, Wong GK, Wu Z, Paulsen IT, Reizer J, Saier MH, Hancock RE, Lory S, Olson MV. 2000. Complete genome sequence of Pseudomonas aeruginosa PAO1, an opportunistic pathogen. Nature 406:959–964 [DOI] [PubMed] [Google Scholar]
- 33.Moskovitz J, Rahman MA, Strassman J, Yancey SO, Kushner SR, Brot N, Weissbach H. 1995. Escherichia coli peptide methionine sulfoxide reductase gene: regulation of expression and role in protecting against oxidative damage. J. Bacteriol. 177:502–507 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lowther WT, Brot N, Weissbach H, Matthews BW. 2000. Structure and mechanism of peptide methionine sulfoxide reductase, an “anti-oxidation” enzyme. Biochemistry 39:13307–13312 [DOI] [PubMed] [Google Scholar]
- 35.Kim HY, Gladyshev VN. 2005. Role of structural and functional elements of mouse methionine-S-sulfoxide reductase in its subcellular distribution. Biochemistry 44:8059–8067 [DOI] [PubMed] [Google Scholar]
- 36.Ranaivoson FM, Neiers F, Kauffmann B, Boschi-Muller S, Branlant G, Favier F. 2009. Methionine sulfoxide reductase B displays a high level of flexibility. J. Mol. Biol. 394:83–93 [DOI] [PubMed] [Google Scholar]
- 37.Moskovitz J, Singh VK, Requena J, Wilkinson BJ, Jayaswal RK, Stadtman ER. 2002. Purification and characterization of methionine sulfoxide reductases from mouse and Staphylococcus aureus and their substrate stereospecificity. Biochem. Biophys. Res. Commun. 290:62–65 [DOI] [PubMed] [Google Scholar]
- 38.Lan L, Murray TS, Kazmierczak BI, He C. 2010. Pseudomonas aeruginosa OspR is an oxidative stress sensing regulator that affects pigment production, antibiotic resistance and dissemination during infection. Mol. Microbiol. 75:76–91 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Lee WL, Gold B, Darby C, Brot N, Jiang X, de Carvalho LP, Wellner D, St John G, Jacobs WR, Jr, Nathan C. 2009. Mycobacterium tuberculosis expresses methionine sulphoxide reductases A and B that protect from killing by nitrite and hypochlorite. Mol. Microbiol. 71:583–593 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Singh VK, Moskovitz J, Wilkinson BJ, Jayaswal RK. 2001. Molecular characterization of a chromosomal locus in Staphylococcus aureus that contributes to oxidative defence and is highly induced by the cell-wall-active antibiotic oxacillin. Microbiology 147:3037–3045 [DOI] [PubMed] [Google Scholar]
- 41.Vattanaviboon P, Seeanukun C, Whangsuk W, Utamapongchai S, Mongkolsuk S. 2005. Important role for methionine sulfoxide reductase in the oxidative stress response of Xanthomonas campestris pv. phaseoli. J. Bacteriol. 187:5831–5836 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Gebendorfer KM, Drazic A, Le Y, Gundlach J, Bepperling A, Kastenmuller A, Ganzinger KA, Braun N, Franzmann TM, Winter J. 2012. Identification of a hypochlorite-specific transcription factor from Escherichia coli. J. Biol. Chem. 287:6892–6903 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Alvarez-Martinez CE, Baldini RL, Gomes SL. 2006. A Caulobacter crescentus extracytoplasmic function sigma factor mediating the response to oxidative stress in stationary phase. J. Bacteriol. 188:1835–1846 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Huis in't Veld RA, Willemsen AM, van Kampen AH, Bradley EJ, Baas F, Pannekoek Y, van der Ende A. 2011. Deep sequencing whole transcriptome exploration of the σE regulon in Neisseria meningitidis. PLoS One 6:e29002. 10.1371/journal.pone.0029002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Alamuri P, Maier RJ. 2006. Methionine sulfoxide reductase in Helicobacter pylori: interaction with methionine-rich proteins and stress-induced expression. J. Bacteriol. 188:5839–5850 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Kwak GH, Hwang KY, Kim HY. 2012. Analyses of methionine sulfoxide reductase activities towards free and peptidyl methionine sulfoxides. Arch. Biochem. Biophys. 527:1–5 [DOI] [PubMed] [Google Scholar]
- 47.Assinger A, Koller F, Schmid W, Zellner M, Koller E, Volf I. 2010. Hypochlorite-oxidized LDL induces intraplatelet ROS formation and surface exposure of CD40L—a prominent role of CD36. Atherosclerosis 213:129–134 [DOI] [PubMed] [Google Scholar]
- 48.Dukan S, Touati D. 1996. Hypochlorous acid stress in Escherichia coli: resistance, DNA damage, and comparison with hydrogen peroxide stress. J. Bacteriol. 178:6145–6150 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Chesney JA, Eaton JW, Mahoney JR., Jr 1996. Bacterial glutathione: a sacrificial defense against chlorine compounds. J. Bacteriol. 178:2131–2135 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Small DA, Chang W, Toghrol F, Bentley WE. 2007. Toxicogenomic analysis of sodium hypochlorite antimicrobial mechanisms in Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 74:176–185 [DOI] [PubMed] [Google Scholar]
- 51.Rutala WA. 1996. APIC guideline for selection and use of disinfectants. 1994, 1995, and 1996 APIC Guidelines Committee. Association for Professionals in Infection Control and Epidemiology, Inc. Am. J. Infect. Control 24:313–342 [DOI] [PubMed] [Google Scholar]
- 52.Le HT, Chaffotte AF, Demey-Thomas E, Vinh J, Friguet B, Mary J. 2009. Impact of hydrogen peroxide on the activity, structure, and conformational stability of the oxidized protein repair enzyme methionine sulfoxide reductase A. J. Mol. Biol. 393:58–66 [DOI] [PubMed] [Google Scholar]
- 53.Atack JM, Kelly DJ. 2008. Contribution of the stereospecific methionine sulphoxide reductases MsrA and MsrB to oxidative and nitrosative stress resistance in the food-borne pathogen Campylobacter jejuni. Microbiology 154:2219–2230 [DOI] [PubMed] [Google Scholar]
- 54.Bobrowski K, Hug GL, Pogocki D, Marciniak B, Schoneich C. 2007. Stabilization of sulfide radical cations through complexation with the peptide bond: mechanisms relevant to oxidation of proteins containing multiple methionine residues. J. Phys. Chem. B. 111:9608–9620 [DOI] [PubMed] [Google Scholar]
- 55.Gu M, Imlay JA. 2011. The SoxRS response of Escherichia coli is directly activated by redox-cycling drugs rather than by superoxide. Mol. Microbiol. 79:1136–1150 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Tao L, Jackson RE, Cheng Q. 2005. Directed evolution of copy number of a broad host range plasmid for metabolic engineering. Metab. Eng. 7:10–17 [DOI] [PubMed] [Google Scholar]
- 57.Lim JC, You Z, Kim G, Levine RL. 2011. Methionine sulfoxide reductase A is a stereospecific methionine oxidase. Proc. Natl. Acad. Sci. U. S. A. 108:10472–10477 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Denkel LA, Horst SA, Rouf SF, Kitowski V, Bohm OM, Rhen M, Jager T, Bange FC. 2011. Methionine sulfoxide reductases are essential for virulence of Salmonella Typhimurium. PLoS One 6:e26974. 10.1371/journal.pone.0026974 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Zhao C, Hartke A, La Sorda M, Posteraro B, Laplace JM, Auffray Y, Sanguinetti M. 2010. Role of methionine sulfoxide reductases A and B of Enterococcus faecalis in oxidative stress and virulence. Infect. Immun. 78:3889–3897 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Metcalf WW, Jiang W, Daniels LL, Kim SK, Haldimann A, Wanner BL. 1996. Conditionally replicative and conjugative plasmids carrying lacZα for cloning, mutagenesis, and allele replacement in bacteria. Plasmid 35:1–13 [DOI] [PubMed] [Google Scholar]