

Abstract
Equine sarcoids are skin tumours of fibroblastic origin affecting equids worldwide. Bovine papillomavirus type-1 (BPV-1) and, less commonly, type-2 are recognized as etiological factors of sarcoids. The transforming activity of BPV is related to the functions of its major oncoprotein E5 which binds to the platelet-derived growth factor β receptor (PDGFβR) causing its phosphorylation and activation. In this study, we demonstrate, by coimmunoprecipitation and immunoblotting, that in equine sarcoid derived cell lines PDGFβR is phosphorylated and binds downstream molecules related to Ras-mitogen-activated protein kinase-ERK pathway thus resulting in Ras activation. Imatinib mesylate is a tyrosine kinase receptors inhibitor which selectively inhibits the activation of PDGFβR in the treatment of several human and animal cancers. Here we show that imatinib inhibits receptor phosphorylation, and cell viability assays demonstrate that this drug decreases sarcoid fibroblasts viability in a dose-dependent manner. This study contributes to a better understanding of the molecular mechanisms involved in the pathology of sarcoids and paves the way to a new therapeutic approach for the treatment of this common equine skin neoplasm.
1. Introduction
Sarcoids are skin tumours of fibroblastic origin affecting equids and are considered to be the most common equine cutaneous neoplasm worldwide. These tumours are locally invasive, nonmetastatic, and very rarely regress. Sarcoids may exist as single or multiple lesions, most frequently arising from sites of previous injuries such as the skin of the head, ventral abdomen, and the paragenital region, with six clinical types recognized: occult, verrucous, nodular, fibroblastic, mixed, and malignant [1]. They are all histologically characterized by proliferation of spindle-shaped dermal fibroblasts forming whorls and by epidermal hyperplasia, hyperkeratosis, and rete peg formation [2, 3]. Although the pathology of this equine neoplasm is not completely understood, a role for bovine papillomavirus type-1 (BPV-1) and, less commonly, BPV-2 infection had been recognized in the etiology of sarcoids [4, 5]. BPV-1 and -2 are oncogenic double-stranded DNA viruses belonging to the genus of Delta papillomaviruses, which are able to infect both epithelial cells and fibroblasts in their natural host [6]. The oncogenic potential of BPV-1/-2 in bovids is known to be related to the expression of viral oncoproteins E5, E6, and E7 [7]; BPV genes have been found to be expressed in sarcoids, suggesting that the viral proteins also play a role in the development of this equine skin neoplasm [8–12].
E5 is the major oncoprotein encoded by BPVs [13]; it is mostly localized in the endomembrane compartments of the Golgi apparatus (GA), endoplasmic reticulum, and plasma membrane of epithelial cells [14]; cytoplasmic and juxtanuclear expression of E5 has also been reported in sarcoid fibroblasts [11].
PDGFβR and other tyrosine kinase receptors are involved in human and animal papillomavirus-induced carcinogenesis since their physiological activity is impaired by E5 oncoproteins of PVs [13].
The major transforming activity of BPV-1 E5 is due to its ability to specifically bind and activate PDGFβR in a ligand independent manner, and, importantly, it does not activate other related receptors [15–18]. E5 binds to PDGFβR as a dimer thereby inducing receptor oligomerization, autophosphorylation, and thus activation [19–21]. PDGFβR is constitutively activated in transformed mouse cells expressing BPV-1 E5, and binding of BPV-1 E5 to PDGFβR induces its activation in mortal human fibroblasts [18, 22]. Importantly, this interaction also takes place in vivo, confirming the role of the E5-PDGFβR complex in carcinogenesis [15, 23]; additionally, PDGFβR has been found to be activated in BPV-E5 positive urinary bladder tumours and equine sarcoids [24, 25].
Activated tyrosine kinase receptors can stimulate intracellular-signalling pathways which regulate cell proliferation, such as the Ras-mitogen-activated protein kinase-ERK (Ras-MAPK-ERK) pathway [26, 27]; once phosphorylated, they recruit growth factor receptor bound protein 2 (GRB2) which is constitutively associated with Sos1 protein, a guanine nucleotide exchange factor of the small GTP-ase, Ras. When recruited to plasma membrane together with GRB2, Sos1 catalyses the switch of Ras from inactive GDP bound form (Ras-GDP) to a transductionally active GTP bound form (Ras-GTP). Ras-GTP binds to and stimulates downstream effectors (among these, Raf1), resulting in phosphorylation and activation of MEK and its downstream kinase ERK, which can regulate cell growth or differentiation [28]. In response to PDGFβR stimulation, GRB2 facilitates activation of the Ras-MAPK-ERK pathway [27] playing a role in brain cancer development [29]. BPV E5 induces recruitment of GRB2 to activated PDGFβR (pPDGFβR), contributing to cell transformation in vitro [22]; furthermore, pPDGFβR recruits GRB2-Sos1 which promote Ras activation in bovine urinary bladder cancer, suggesting a role of this pathway in BPV-induced carcinogenesis [25].
Imatinib mesylate is a tyrosine kinase receptor inhibitor formerly known as STI-571 or Gleevec; this small molecule, derived from 2-phenylaminopyrimidine, binds to the ATP-binding site, thus inhibiting the tyrosine kinase activity of the PDGFR (both α and β) and the downstream signals, including the Ras-MAPK-ERK cell proliferation pathway [30–32]. Imatinib has been largely studied in human solid tumours, where it has been shown to induce cell growth arrest [33–37]. Furthermore, imatinib selectively inhibits the Bcr-Abl tyrosine kinase in chronic myeloid leukemia and c-kit in several human cancers, resulting in either apoptosis or inhibition of proliferation [30, 31, 38–42]. Gleevec is used in veterinary medicine for the treatment of canine mast cells tumours (MCTs) [43–45] and has been proposed as therapy for several feline neoplasms, including vaccine associated sarcomas (VAS), in which the PDGFβR is selectively inhibited by the treatment [46, 47].
The aim of this study was to investigate the activation of PDGFβR, the Ras-MAPK-ERK pathway and to further evaluate the effects of imatinib mesylate on PDGFβR phosphorylation and cell viability in an equine sarcoid disease model.
2. Methods
2.1. Cells and Cell Culture
Equine sarcoid-derived cell lines EqS01 and EqS04b, expressing endogenous BPV-1 E5, EqS02a transfected with BPV-1 E5 (EqS02aE5) and EqS02a cells which do not express E5 and are not fully transformed, have all been described previously [48, 49]. Cells were cultured in Dulbecco's modified eagle medium (DMEM) supplemented with 10% fetal bovine serum (FBS) (Gibco) or in serum-deprived DMEM depending on the experiment, in a 37°C humidified atmosphere of 5% CO2 in air.
2.2. Coimmunoprecipitation, Immunoprecipitation, and Immunoblotting
To prepare protein extracts for immunoprecipitation, 80% confluent monolayers in 150 mm dishes were washed three times in phosphate-buffered saline (PBS; pH 7.4, 0.1 M) and then lysed in ice-cold JS buffer (50 mM HEPES, PH 7.5, 150 mM NaCl, 1% glycerol, 1% Triton-X100, 150 μM MgCl2, and 5 mM ethylene glycol tetra acetic acid) added with 20 mM sodium pyrophosphate, 0,1 mg mL−1 aprotinin, 2 mM phenylmethylsulfonyl fluoride, 10 mM disodium orthovanadate, and 50 mM sodium fluoride. Protein concentrations were determined by use of a protein assay kit (Bio-Rad Laboratories). Equal amounts of protein lysates (500 μg) were immunoprecipitated with 2 μg mL−1 rabbit anti-pPDGFβR antibody (Santa Cruz Biotechnology) overnight at 4°C. A-G/plus sepharose beads (Santa Cruz Biotechnology) (20 μL) were added to the samples, and the mixture was rotated for 1 h at 4°C. After 3 washings in lysis buffer, the immunoprecipitates were resuspended in Laemmli sample buffer [50] (sodium dodecyl sulphate (SDS), Tris-HCl pH 6.8, glycerol, bromophenol blue, and 2β-mercaptoethanol) and analyzed by SDS polyacrylamide gel electrophoresis (PAGE) and immunoblotting. Nitrocellulose membranes were blocked with 5% bovine serum albumin (BSA)-in-Tris-buffered saline (TBS: 12.5 mM TrisHCl pH 7.4; 125 mM NaCl) at room temperature (RT) and incubated O/N at 4°C with rabbit anti-pPDGFβR, mouse anti-GRB2 (Upstate Biotechnology) and rabbit anti-Sos1 (Santa Cruz Biotechnology) antibodies diluted 1 : 500, 1 : 1000, and 1 : 200, respectively. After 3 washing steps in TBS-Tween 0.1%, appropriate peroxidase-conjugated secondary antibodies (Amersham, Gel Health Care) were applied 1 h at RT at 1 : 1000 dilution. Membranes were washed again, and bound antibodies were visualized by enhanced chemiluminescence (ECL) (Western Blot Luminol Reagent, Santa Cruz Biotechnology). Protein levels were quantitatively estimated by densitometry using ChemiDoc gel scanner (Bio-Rad Laboratories) equipped with a densitometric workstation (Image Lab software, Bio Rad Laboratories).
Similarly, protein extracts were immunoprecipitated for Sos1 (2 μg mL−1) and analyzed by SDS-PAGE and immunoblotting with anti-GRB2 and anti-Sos1 antibodies. The reciprocal coimmunoprecipitation assay could not be performed since the anti-GRB2 antibody does not work for immunoprecipitation.
For evaluation of phosphorylation status of PDGFβR in starved serum-deprived cells, the same amount of protein lysates were immunoprecipitated with a rabbit anti-PDGFβR antibody (2 μg mL−1) (Santa Cruz Biotechnology) and probed for PDGFβR presence, using the aforementioned antibody; the membranes were stripped and reprobed with a mouse antiphosphotyrosine (pTyr) antibody at 1 : 1000 dilution (Upstate Biotechnology). Bound antibodies were visualized as mention above.
For the evaluation of activation status of PDGFβR after treatment with imatinib mesylate, cell lysates were subjected to immunoprecipitation with rabbit anti-PDGFβR and immunoblotting. The membranes were incubated with mouse anti-pTyr antibody, washed, and probed with appropriate secondary antibody. After stripping, rabbit anti-PDGFβR antibody was applied and revealed as mentioned above.
Equal volumes (30 μL) of cell lysates were analyzed by immunoblotting for actin levels detection (see the following for details) before coimmunoprecipitation and immunoprecipitation assays to ensure equal amounts of protein loading and allow normalization.
2.3. Ras-Pull-Down Assay
To assess Ras activation status, a Ras-pull-down assay (Millipore) was performed according to manufacturer protocol with slight modifications. Briefly, cell lysates were incubated with GST-Raf1-RBD agarose beads for 15 minutes at 4°C with gentle agitation. Beads containing activated Ras were collected by centrifugation, washed with JS lysis buffer, and then prepared and analyzed for immunoblotting with a mouse monoclonal anti-Ras antibody provided by the kit. Raf1-RBD-Ras levels were normalized to total Ras levels and expressed as densitometric ratio. Total Ras expression levels were normalised to actin levels.
2.4. Immunoblotting on Whole Cell Lysates
For protein expression analysis on whole cell lysates, equal amounts of proteins were boiled in 2x Laemmli sample buffer [50], electrophoresed, and subjected to immunoblotting; after blocking in TBS-BSA 5%, rabbit anti-PDGFβR diluted 1 : 500, rabbit anti-pMEK antibody diluted 1 : 1000, and rabbit anti-pERK (pERK) antibody at 1 : 2000 dilution (Cell Signaling Technology) were applied O/N at 4°C. The membranes were washed and incubated with peroxidase-conjugated anti-rabbit IgG diluted 1 : 1000, 1 h at RT. Following further washing, bound antibodies were visualized as above. The membranes were stripped and reprobed with mouse anti-actin antibody (Calbiochem) at 1 : 5000 dilution to ensure equal amount of proteins for each sample. The protein concentrations were normalised to the actin levels.
2.5. RT-PCR and Sequencing of Exons 11–20 of PDGFβR Gene
Total RNA from 80% confluent cell monolayers in 60 mm dishes was extracted using RNeasy mini Kit (Qiagen). Following DNase I treatment, first-strand cDNA from PDGFβR gene was synthesized using SuperScript III First-Strand Synthesis System for RT-PCR (Invitrogen) according to manufacturer's protocol. cDNA was amplified using different primers sets, which amplified different regions of all sequences from exon 11 to exon 20. The primers sets, the annealing position, the size of amplified fragments, and the annealing temperatures are summarized in Table 1. PCR conditions were as follows: denaturation at 94°C for 20 s, followed by 28 cycles at 94°C for 20 s, 52°C for 30 s, and 68°C for 20 s, with a final extension at 68°C for 10 min. PCR products were separated by electrophoresis in 1% agarose gels with Tris borate ethylene diamine tetra acetic acid (EDTA) buffer (TBE; 89 mMTris base, 89 mM Boric acid, and 2 mM EDTA), stained with ethidium bromide, and visualised under ultraviolet light. Amplicons were purified using the Charge Switch PCR Clean-Up Kit (Invitrogen) following manufacturer's instructions. Cycle sequencing reactions were performed using Applied Biosystems BigDye Terminator Ready Reaction v3.1 Kit, followed by purification of sequence reactions and electrophoresing using ABI Prism 3130XL Genetic Analyser (Applied Biosystems). The sequences were aligned to Equus caballus wild type PDGFβR using Basic Local Alignment Search Tool (NCBI/BLAST).
Table 1.
Primers sets used to amplify cDNA from exons 11–20 of PDGFβR in EqS cells.
| Primers sets | Annealing position | Annealing T° | Fragments size |
|---|---|---|---|
| F1: ATCCTCATCATGCTCTGGCAG | F1: 2119 | 52°C | 628 bp |
| R1: TCTCCTTTCATGTCCAGCATG | R1: 2747 | ||
| F1: ATCCTCATCATGCTCTGGCAG | F1: 2119 | 52°C | 1226 bp |
| R3: TCGAGAAGCAGCACCAGCTG | R3: 3344 | ||
| F3: TGGCTACATGGACATGAGCAAG | F3: 2682 | 52°C | 663 bp |
| R3: TCGAGAAGCAGCACCAGCTG | R3: 3344 | ||
| F3: TGGCTACATGGACATGAGCAAG | F3: 2682 | 52°C | 593 bp |
| R2: ATCTCGTAGATCTCGTCGGAG | R2: 3275 |
T°: temperature; F: forward; R: reverse; bp: base pairs.
2.6. Double Immunofluorescence and Confocal Microscopy
EqS cell lines were grown for 2 days on coverslips, washed with PBS, fixed in 4% paraformaldehyde for 20 min, and permeabilized with 0,1% triton X-100 in PBS 5 min. The slides were blocked with 2% BSA for 30 min. Sheep anti-E5 (a kind gift of Professor Maria Saveria Campo, University of Glasgow, Scotland [51, 52]) and rabbit anti-PDGFβR primary antibodies were applied O/N at 4°C in a humidified chamber at 1 : 50 dilution in PBS. The slides were washed three times with PBS, then incubated with Alexa Fluor 488 donkey anti-sheep and Alexa Fluor 546 goat anti-rabbit 30 min at RT (Molecular Probes) in a humidified chamber at 1 : 100 dilution. Finally, after washing with PBS, the slides were mounted in aqueous medium PBS : Glycerol 1 : 1 (Sigma). For scanning and photography, a confocal laser-scanning microscope LSM-510 (Zeiss) was used. Alexa Fluor 488/546 was irradiated at 488 nm and 543 nm and detected with a 505–530 nm and 506–615 nm bandpass filters, respectively.
2.7. Imatinib Mesylate, Inhibition of PDGFβR Activation, and Cell Viability Assay
Imatinib mesylate was provided by LC laboratories (Woburn, USA). For our studies, 50 mM stock solutions were prepared in DMSO and stored at −80°C. Dilutions of imatinib were made from the stock solutions in serum-free medium.
For the evaluation of activation status of PDGFβR after drug treatment, 80% confluent cells monolayers in 60 mm dishes were incubated in serum-free medium for 24 h and treated for 15 min with imatinib at the following concentrations: 0,01 μM, 0,1 μM, 1 μM, 5 μM, with or without 30 ng mL−1 of human recombinant PDGF-BB (PDGFβR natural ligand) (Sigma). Cells were then lysed and subjected to immunoprecipitation of PDGFβR and immunoblotting for pTyr and PDGFβR as described previously.
For cell viability assay, cells were plated in 96-well microtiter plates at 10000 cells/well and incubated in DMEM 10% FBS in standard conditions. After 24 h, the plates were washed, and serum-free medium containing various concentrations of imatinib with or without 30 ng mL−1 of human recombinant PDGF-BB was added. Each condition was replicated in five wells. Relative viable cell numbers were measured after 72 h using the Cell Titer 96 AQ ueous one solution assay (Promega), a colorimetric system based on the tetrazolium salt MTS, according to manufacturer's protocol. Absorbance at 490 nm was measured using a Sirio-S Reader (Seac and Radim Diagnostics) and Sirio-S v7.0 software.
3. Results
3.1. PDGFβR Activation and Its Binding to Downstream Molecules in Equine Sarcoid Fibroblasts
In order to investigate the activation of PDGFβR, we first assessed PDGFβR expression levels, which were very similar among all EqS cell lines (Figure 1), and then the interaction of activated receptor with its molecular substrates, GRB2 and Sos1. pPDGFβR immunoprecipitation followed by immunoblotting for pPDGFβR, GRB2, and Sos1-yielded the following results: pPDGFβR is immunoprecipitated in higher amounts in EqS02aE5 and EqS04b, both expressing higher levels of BPV-1 E5, and, to a lesser extent, in EqS01a, which expresses lower levels of the oncogene, when compared to EqS02a; the latter cell line harbours very few copies of viral genome, and E5 expression is not detectable [48, 49]; the levels of GRB2 and Sos1 co-immunoprecipitated with pPDGFβR were increased in EqS01a, EqS02aE5, and EqS04b correlating with pPDGFβR levels, when compared to EqS02a (Figure 2).
Figure 1.
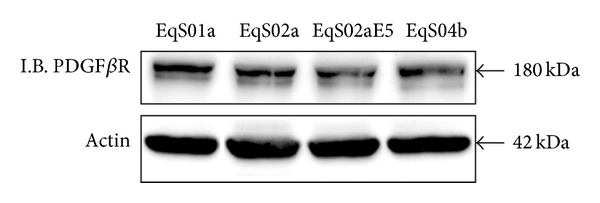
Immunoblotting (I.B.) analysis of PDFGβR expression in sarcoid-derived cell lines (EqS). PDFGβR was expressed at similar levels in all the analyzed cell lines. Actin protein levels confirmed equal amount of protein loading in each lane.
Figure 2.
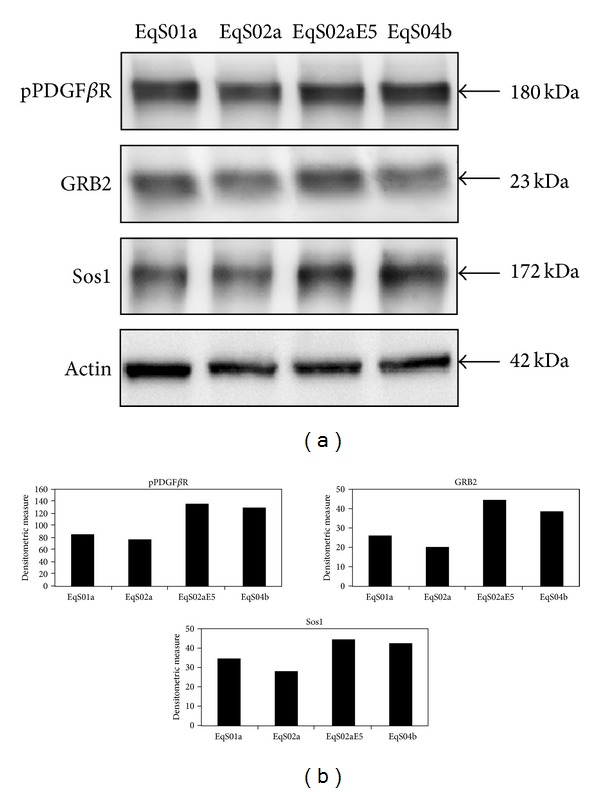
pPDFGβR-GRB2-Sos1 coimmunoprecipitation in EqS cell lines. (a) The presence of GRB2 and Sos1 was detected in pPDFGβR immunoprecipitates in higher amount in cell lines EqS01a, EqS02aE5, and EqS04b expressing BPV-1 E5 when compared to EqS02a which do not express the oncoprotein. Actin protein levels were detected on whole cell lysates before immunoprecipitation to ensure equal protein loading and allow normalization. (b) Quantitative densitometric analysis of the bands was performed with Image Lab software (ChemiDoc, Bio-Rad Laboratories).
Since Sos1 could be recruited to activated PDGFβR via GRB2, we next analyzed the physical interaction between Sos1 and GRB2 by coimmunoprecipitation. GRB2 bound Sos1 in all the analyzed cell lines; however, the complex was found in larger amounts in EqS02aE5 and EqS04b (both highly expressing E5) (Figure 3).
Figure 3.
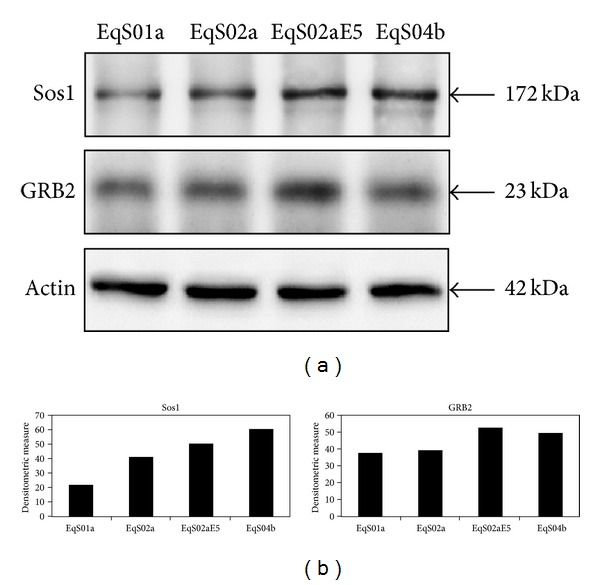
Sos1-GRB2 coimmunoprecipitation. (a) Binding of GRB2 to Sos1 was recorded in all EqS cell lines; the complex was coimmunoprecipitated in higher amount in EqS02aE5 and EqS04b, which express the highest levels of BPV-1 E5 among the analyzed cell lines. Actin protein levels were detected on whole cell lysates before immunoprecipitation to ensure equal protein loading and allow normalization. (b) Quantitative densitometric analysis of the bands was performed with Image Lab software (ChemiDoc, Bio-Rad Laboratories).
3.2. Molecular Analysis of Ras-MAPK-ERK Pathway
To further analyze the downstream signalling molecules of pPDGFβR complexed with GRB2-Sos1, we investigated the activation of Ras using a pull-down assay. Firstly, immunoblotting on whole cell lysates collected before performing the pull-down assay showed similar Ras-expression levels in EqS02a, EqS02aE5 and EqS04b. Ras was found to be overexpressed and, consistently, activated at higher levels in EqS01a when compared to other cell lines. Raf1-RBD-Ras was also detected at higher levels in EqS02aE5 and EqS04b when compared to EqS02a (Figures 4(a) and 4(b)).
Figure 4.
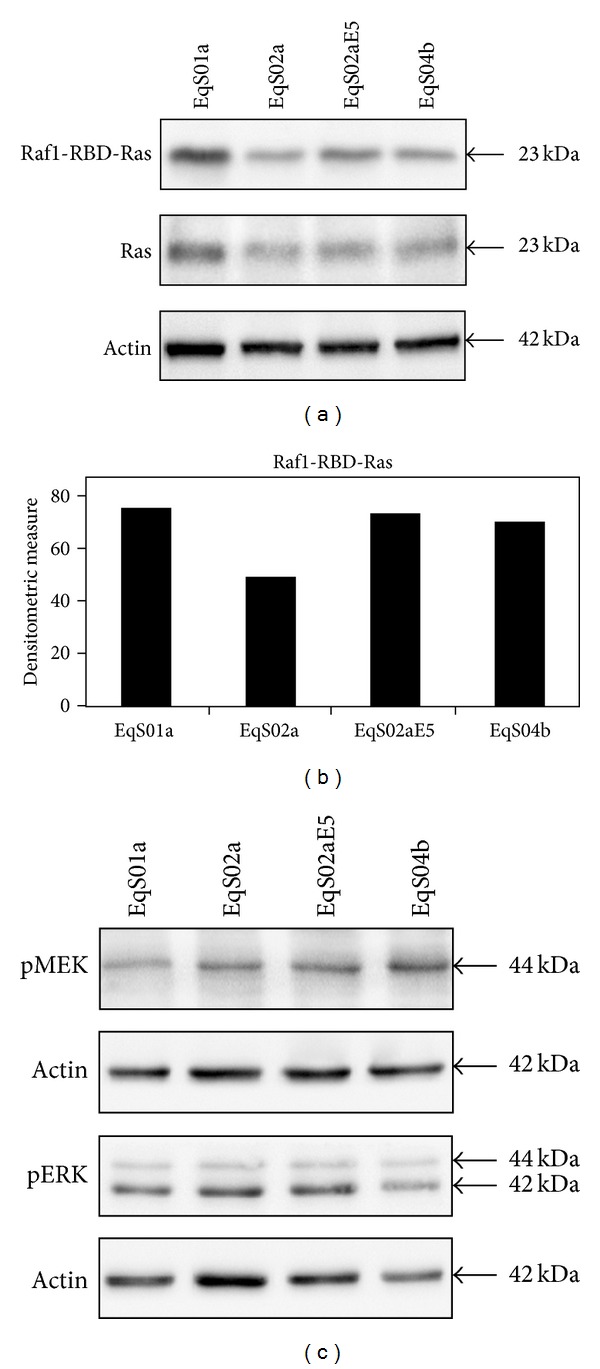
Analysis of Ras-mitogen-activated protein kinase-ERK pathway in sarcoid-derived cell lines (EqS). (a) Immunoblotting analysis of Ras in a pull-down assay of activated Ras (Raf1-RBD-Ras). Raf1-RBD-Ras was increased in cell lines EqS01a (to higher extent), EqS02aE5, and EqS04b when compared to EqS02a (not expressing E5). Total Ras levels were detected on whole cell lysates before pull-down assay to allow normalization of Raf1-RBD-Ras. Ras was overexpressed in EqS01a. Ras expression levels were normalized to actin levels. (b) Quantitative densitometric analysis of the bands was performed with Image Lab software (ChemiDoc, Bio-Rad Laboratories). (c) Immunoblotting analysis of pMEK and pERK expression. No differences in phosphorylation status of both MEK and ERK kinases were recorded. Actin protein levels were detected to ensure equal amounts of proteins for each lane.
The phosphorylation status of ERK (pERK) and its upstream kinase MEK (pMEK) were also determined by immunoblotting using phosphospecific antibodies; however, no differences in expression levels were observed in EqS cell lines (Figure 4(c)).
3.3. Analysis of PDGFβR Activation in Serum-Starved EqS Cells
EqS cell lines were grown in serum-deprived medium to exclude the possibility that receptor activation could be due to growth factors from FBS. Immunoprecipitation assay, performed by using an anti-PDGFβR antibody followed by immunoblotting with anti-PDGFβR and anti-pTyr antibodies, revealed that PDGFβR is phosphorylated only in EqS02aE5 and EqS04b, both expressing high levels of E5 (Figure 5).
Figure 5.
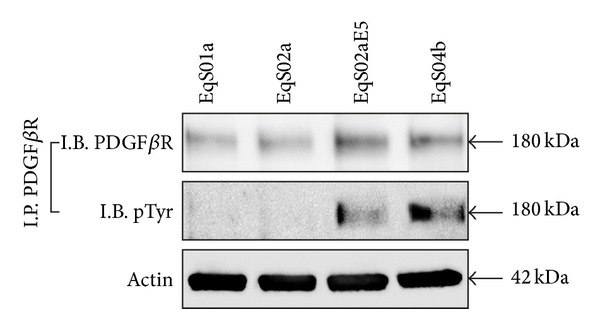
Activation of PDFGβR in serum-starved sarcoid-derived cell lines (EqS). The PDFGβR immunoprecipitated in serum-deprived cells is phosphorylated only in EqS02aE5 and EqS04b, which express the higher levels of BPV-1 E5 among the analyzed cell lines (I.P.: immunoprecipitation; I.B.: immunoblotting).
3.4. Sequence Analysis of Exons 11–20 of PDGFβR Gene
To further evaluate whether the sustained activation of PDGFβR in EqS cell lines might be caused by activating mutations in the transmembrane and/or cytosolic domain, cDNAs of exons 11–20 of PDGFβR were amplified by RT-PCR using different primers sets followed by sequencing. Sequence analysis and alignment with wild type sequence revealed that no mutations occurred in these exons in EqS cell lines (data not shown).
3.5. Colocalization of PDGFβR with BPV-1 E5
As gene mutations that may induce activation of PDGFβR have been excluded, we next sought to address whether the receptor c-localizes with BPV-1 E5. EqS cell lines were analyzed by double labelling immunofluorescence using rabbit anti-PDGFβR and sheep anti-E5 primary antibodies (green fluorescence for E5 and red fluorescence for PDGFβR). Interestingly, both proteins appeared to be mostly expressed in a juxtanuclear position in EqS02aE5, where they markedly colocalize as judged by the yellow fluorescence of merged images (Figure 6(a)). PDGFβR was found to be expressed also in the cytoplasm of EqS01a, EqS04b (data not shown), and EqS02a cells (Figure 6(b)); the latter cell line showed, as expected, no signal for E5 (Figure 6(b)). BPV-1 E5 staining was not recorded in EqS01a and EqS04b, which is probably due to undetectable expression levels of endogenous protein by immunofluorescence (expression of E5 in these cells has been shown previously by RT-PCR [48]).
Figure 6.
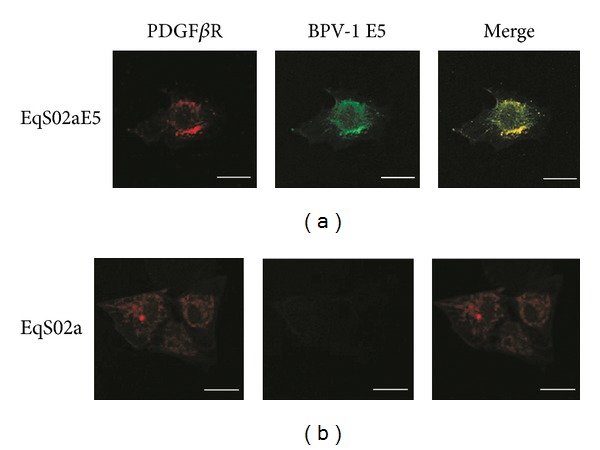
BPV-1E5-PDFGβR colocalization. (a) BPV-1 E5 (green fluorescence) and PDFGβR (red fluorescence) markedly colocalized (merged panel, yellow fluorescence) in a juxtanuclear Golgi-like position in sarcoid-derived EqS02a cells transfected with BPV-1 E5 (EqS02aE5). A representative image is shown. (b) EqS02a cells expressed PDFGβR in the cytoplasm and showed no green signal for E5. Bar = 20 μm.
3.6. Effects of Imatinib Mesylate on PDGFβR Phosphorylation and Cell Viability
Serum-starved cells were exposed to various concentration of imatinib with or without PDGF-BB in order to determine the effects of the drug on PDGFβR activation and cell viability in EqS cell lines. Protein extracts were subjected to immunoprecipitation of PDGFβR followed by immunoblotting with anti-pTyr and anti-PDGFβR antibodies. Figure 7 clearly shows that PDGF-BB was able to induce PDGFβR transphosphorylation, which indeed was not observed in the absence of ligand; furthermore imatinib is shown to inhibit PDGFβR autophosphorylation induced by PDGF-BB in a dose-dependent manner, with near complete inhibition at a concentration of 5 μM in all the cell lines. Additionally, PDGFβR expression levels decreased parallel to the increase of its phosphorylation degree, whereas they increased concomitantly with inhibition of phosphorylation by imatinib with a dose-dependent manner.
Figure 7.
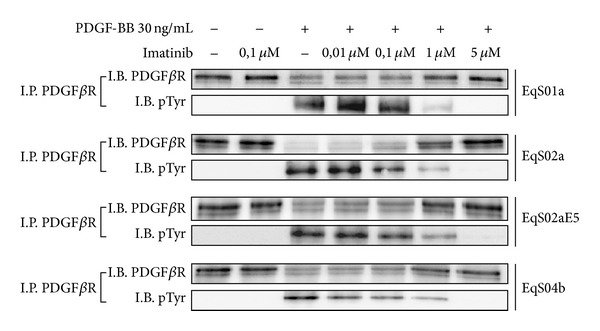
Dose-dependent inhibition of PDGF-BB-induced PDFGβR activation by imatinib mesylate. In sarcoid-derived cell lines (EqS) treated with increasing doses of imatinib with or without PDGF-BB, the drug inhibited PDFGβR autophosphorylation. PDGFβR downregulation was another measure of its activation induced by PDFG-BB, and, conversely, increasing of PDGFβR expression in cells treated with higher doses of imatinib confirmed the inhibition of receptor activation (I.P.: immunoprecipitation; I.B.: immunoblotting).
Similarly, cells were exposed to 10% FBS or various concentrations of imatinib with or without PDGF-BB and subjected to MTS-based cell viability assay. As shown in Figure 8, all the cell lines proliferated in the presence of both FBS and PDGF-BB, and imatinib at a concentration of 0.1 μM is already sufficient to severely reduce cell viability in EqS01a, EqS02aE5, and EqS04b; treatment with increasing doses of imatinib caused a further gradual decrease in cell viability levels. All these biological effects appear to be milder in EqS02a, in which no difference in cell viability can be observed at the highest doses of imatinib.
Figure 8.
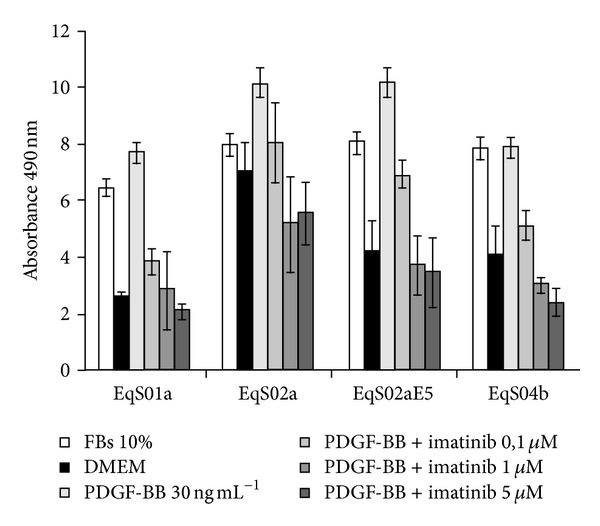
Effects of imatinib mesylate on PDGF-BB-stimulated sarcoids-derived cells (EqS) viability. MTS-based assay revealed that PDGF-BB stimulates cell growth as well as FBS and that imatinib decreases cell viability in all the analyzed cell lines in a dose-dependent manner, but with lower efficiency in EqS02a (not expressing E5). The data represent the mean of repeated independent experiments.
4. Discussion
E5 is the major oncoprotein encoded by BPV-1 and plays a key role in the tumorigenic process [13]; the main transforming activity of BPV E5 is due to its specifical binding to PDGFβR, causing receptor autophosphorylation and activation, which results in mitogenic signalling and neoplastic transformation [18, 20].
Among the downstream pathways of pPDGFβR, the Ras-MAPK-ERK signalling is known to regulate cell proliferation [26, 27]. In this study we investigated the activation of PDGFβR and Ras-MAPK-ERK pathway in equine sarcoid-derived cell lines EqS01a, EqS02a, EqS02aE5, and EqS04b [48, 49]. We found that PDGFβR was phosphorylated and bound its downstream partners GRB2-Sos1 in higher amounts in EqS02aE5 and EqS04b (both containing high levels of oncoproteins transcripts) and, to a lesser extent, in EqS01a when compared to EqS02a (which expresses low and not detectable levels of oncoproteins, resp.). Consistently, active Ras was expressed at higher levels in EqS cell lines expressing E5, suggesting that BPV-1 E5 may contribute to activation of this pathway and lead to sarcoid fibroblasts full transformation. Accordingly, PDGFβR was found to be constitutively activated in transformed mouse cells expressing BPV-1 E5, and pPDGFβR binds GRB2 in mortal human fibroblasts transfected with the oncoprotein [18, 22]; moreover, PDGFβR has been found to be activated in vivo in BPV-E5 positive equine sarcoids, and pPDGFβR recruits GRB2-Sos1 which enhance Ras activation in bovine urinary bladder cancer [24, 25]; our finding of total Ras overexpression in EqS01a cell line is in agreement with previous studies which reported an association between PV infection in bovids and activation of Ras gene [53]: these data may suggest a role of this pathway in BPV-induced carcinogenesis. In addition, Ras had been found to be activated in cultured fibroblasts also by Rhesus-PV E5, indicating that E5 genes may play a major role in the regulation of this transduction pathway [54].
Surprisingly, no differences were found in phosphorylation status of the downstream kinases of activated Ras, namely, pMEK and pERK. Previous studies showed that E5 does not promote any changes in ERK activity in cultured fibroblasts [22, 55] as well as in BPV-induced tumours in vivo: it is therefore possible that pPDGFβR activates GRB2, Sos1, and Ras which may deviate on phosphatidylinositol-3-kinase/AKT pathway (PI3 K/AKT). As matter of the fact, PI3 K/AKT pathway, rather than MEK-ERK signalling, had been found to be activated in BPV-induced tumours, thus contributing to neoplastic transformation [25, 56].
To further investigate the possible factors contributing to PDGFβR activation, we first assessed PDGFβR phosphorylation status on serum starved cells: the receptor was found to be phosphorylated only in EqS02aE5 and EqS04b, which express the highest levels of BPV-1 E5 among all cell lines, suggesting that its activation may be due to the interaction with the E5 oncoprotein rather than to the presence of growth factors added to the medium. Accordingly, Petti et al. demonstrated that BPV-1 E5 activates PDGFβR in a ligand-independent manner [18]. Taken together, all these findings may indicate that the extent of PDGFβR activation may correlate with E5 expression levels and the number of viral copies; direct association of viral load with impaired expression of tyrosine kinase receptors has been reported for human cervical tumours harboring human papillomavirus, suggesting that the impairing of these cellular functions may depend on the viral load in PVs induced cancer [57].
PDGFR and other related tyrosine kinase receptors may be activated by mutations or genetic rearrangements causing tumours in both human and animal species, particularly in the transmembrane and cytosolic domains [58–64]. Sequence analysis of PDGFβR in our cell lines revealed that no mutations occurred along these domains, suggesting that the receptor is not activated by itself in our experimental model.
Furthermore, double labeling immunofluorescence showed that PDGFβR perfectly colocalized with BPV-1 E5 in EqS02aE5 where it was mostly expressed in a juxtanuclear position, consistently with the location of E5 in the Golgi apparatus (GA) [14, 65]. The intracellular colocalization of E5 and PDGFβR indicates that this protein interaction may take place in the GA and may be another proof of evidence of the possible receptor activation upon E5 interaction; juxtanuclear colocalization of E5 with PDGFβR was also reported in bovine urinary bladder cancer by this research group, suggesting that this finding could be common in BPV-induced tumours [15]. Many studies have reported the physical interaction between these two proteins and subsequent activation of PDGFβR both in vitro and in vivo [23, 66], thus we speculate that BPV-1 E5 may bind to PDGFβR also in our sarcoid-derived cell lines expressing E5, inducing its phosphorylation and activation of downstream pathways leading to transformation.
Although many therapeutical strategies have been proposed for treatment of sarcoid, no 100% effective therapy is available so far [1]. Many tyrosine kinase receptors inhibitors are used in veterinary medicine [67]; among these, imatinib mesylate (also known as STI-571 or Gleevec) has been shown to selectively inhibit PDGFβR activation in both human and animal tumours [33–36, 47]. In this study we showed that sarcoid fibroblasts proliferation was mostly dependent on PDGFβR stimulation; furthermore, we demonstrated that imatinib inhibited PDGFβR phosphorylation in a dose-dependent manner. In this case no phosphorylation of PDGFβR could be observed in serum-free conditions, whilst the receptor was phosphorylated after 24 h of starvation, suggesting that longer times are needed to reach detectable levels of its activation. The finding of total PDGFβR downregulation concomitant to the augmentation of its phosphorylation degree induced by PDGF-BB, was not surprising: for the PDGFβR and other tyrosine kinase receptors, ligand binding induces receptor phosphorylation and thus triggers its clustering in coated pits, followed by endocytosis and lysosomal degradation of receptor-ligand complexes [68, 69]. Thus the augmentation of PDGFβR expression levels at increasing doses of drug was another measure of the efficiency of treatment. Here we also demonstrated that imatinib was able to decrease sarcoid fibroblasts cell viability in a dose-dependent manner. However, lower effects were observed on cell viability in EqS02a by imatinib treatment at all the experimental doses; lacking E5 expression, this cell line may have lower levels of activated PDGFβR, thus the drug may have less access to the ATP-binding site and, as a consequence, milder biological effects when compared to other cell lines. These findings strengthen our hypothesis of a PDGFβR activation by E5 and suggest that imatinib may target more efficiently on fully transformed sarcoid fibroblasts.
5. Conclusions
Finally, our study demonstrates that PDGFβR is activated thus binding downstream molecular partners in sarcoid-derived cell lines expressing BPV-1 E5; this activation results in Ras activation but not major phosphorylation of MEK and ERK kinases, suggesting that this signalling cascade may possibly cross-talk with other transduction pathways which had been found to be activated in BPV-induced tumours. Further investigations are needed to clarify the specific roles of activated PDGFβR and downstream pathways in the pathology of equine sarcoid. Furthermore, the data obtained by treatment of sarcoid-derived fibroblasts with imatinib, suggest that this drug could be proposed for a clinical trial leading to therapy of sarcoid in vivo. We have developed a translational approach, and a clinical trial is ongoing to test on sarcoid-affected donkeys the therapeutic effects of a dermatological cream based on Imatinib mesylate (CESA protocol number 2012/0052665).
Authors' Contribution
Gennaro Altamura and Annunziata Corteggio have contributed equally to this work.
Acknowledgments
The authors wish to express their gratitude to Ph.D. student F. Bocaneti for her technical help. The authors are grateful to Professor Maria Saveria Campo (Emeritus Professor, Glasgow University) for her kind gift of sheep anti-E5 antibody and for her critical reading of the paper. This work was supported by grant from MIUR: PRIN 2009: L ‘EMATURIA ENZOOTICA CRONICA NEL BOVINO. CUP E6J11000300001.
References
- 1.Borzacchiello G, Corteggio A. Equine sarcoid: state of the art. Ippologia. 2009;20(4):7–14. [Google Scholar]
- 2.Martens A, De Moor A, Demeulemeester J, Ducatelle R. Histopathological characteristics of five clinical types of equine sarcoid. Research in Veterinary Science. 2000;69(3):295–300. doi: 10.1053/rvsc.2000.0432. [DOI] [PubMed] [Google Scholar]
- 3.Scott DW, Miller WH. Equine dermatology. Journal of Equine Veterinary Science. 2003;23(2):65–67. [Google Scholar]
- 4.Lancaster WD, Theilen GH, Olson C. Hybridization of bovine papilloma virus type 1 and type 2 DNA to DNA from virus-induced hamster tumors and naturally occurring equine tumors. Intervirology. 1979;11(4):227–233. doi: 10.1159/000149038. [DOI] [PubMed] [Google Scholar]
- 5.Gorman NT. Equine sarcoid—time for optimism. Equine Veterinary Journal. 1985;17(6):412–414. doi: 10.1111/j.2042-3306.1985.tb02543.x. [DOI] [PubMed] [Google Scholar]
- 6.Nasir L, Campo MS. Bovine papillomaviruses: their role in the aetiology of cutaneous tumours of bovids and equids. Veterinary Dermatology. 2008;19(5):243–254. doi: 10.1111/j.1365-3164.2008.00683.x. [DOI] [PubMed] [Google Scholar]
- 7.Borzacchiello G, Roperto F. Bovine papillomaviruses, papillomas and cancer in cattle. Veterinary Research. 2008;39(5, article 45) doi: 10.1051/vetres:2008022. [DOI] [PubMed] [Google Scholar]
- 8.Nasir L, Reid SWJ. Bovine papillomaviral gene expression in equine sarcoid tumours. Virus Research. 1999;61(2):171–175. doi: 10.1016/s0168-1702(99)00022-2. [DOI] [PubMed] [Google Scholar]
- 9.Carr EA, Théon AP, Madewell BR, Hitchcock ME, Schlegel R, Schiller JT. Expression of a transforming gene (E5) of bovine papillomavirus in sarcoids obtained from horses. American Journal of Veterinary Research. 2001;62(8):1212–1217. doi: 10.2460/ajvr.2001.62.1212. [DOI] [PubMed] [Google Scholar]
- 10.Chambers G, Ellsmore VA, O’Brien PM, et al. Sequence variants of bovine papillomavirus E5 detected in equine sarcoids. Virus Research. 2003;96(1-2):141–145. doi: 10.1016/s0168-1702(03)00175-8. [DOI] [PubMed] [Google Scholar]
- 11.Borzacchiello G, Russo V, Della Salda L, Roperto S, Roperto F. Expression of platelet-derived growth factor-beta receptor and bovine papillomavirus E5 and E7 oncoproteins in equine sarcoid. Journal of Comparative Pathology. 2008;139(4):231–237. doi: 10.1016/j.jcpa.2008.07.006. [DOI] [PubMed] [Google Scholar]
- 12.Corteggio A, Altamura G, Roperto F, Borzacchiello G. Bovine papillomavirus E5 and E7 oncoproteins in naturally occurring tumors: are two better than one? Infectious Agents and Cancer. 2013;8, article 1 doi: 10.1186/1750-9378-8-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Venuti A, Paolini F, Nasir L, et al. Papillomavirus E5: the smallest oncoprotein with many functions. Molecular Cancer. 2011;10, article 140 doi: 10.1186/1476-4598-10-140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Pennie WD, Grindlay GJ, Cairney M, Campo MS. Analysis of the transforming functions of bovine papillomavirus type 4. Virology. 1993;193(2):614–620. doi: 10.1006/viro.1993.1169. [DOI] [PubMed] [Google Scholar]
- 15.Borzacchiello G, Russo V, Gentile F, et al. Bovine papillomavirus E5 oncoprotein binds to the activated form of the platelet-derived growth factor β receptor in naturally occurring bovine urinary bladder tumours. Oncogene. 2006;25(8):1251–1260. doi: 10.1038/sj.onc.1209152. [DOI] [PubMed] [Google Scholar]
- 16.DiMaio D, Mattoon D. Mechanisms of cell transformation by papillomavirus E5 proteins. Oncogene. 2001;20(54):7866–7873. doi: 10.1038/sj.onc.1204915. [DOI] [PubMed] [Google Scholar]
- 17.Goldstein DJ, Li W, Wang L-M, et al. The bovine papillomavirus type 1 E5 transforming protein specifically binds and activates the β-type receptor for the platelet-derived growth factor but not other related tyrosine kinase-containing receptors to induce cellular transformation. Journal of Virology. 1994;68(7):4432–4441. doi: 10.1128/jvi.68.7.4432-4441.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Petti L, Nilson LA, DiMaio D. Activation of the platelet-derived growth factor receptor by the bovine papillomavirus E5 transforming protein. The EMBO Journal. 1991;10(4):845–855. doi: 10.1002/j.1460-2075.1991.tb08017.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Nappi VM, Petti LM. Multiple transmembrane amino acid requirements suggest a highly specific interaction between the bovine papillomavirus E5 oncoprotein and the platelet-derived growth factor beta receptor. Journal of Virology. 2002;76(16):7976–7986. doi: 10.1128/JVI.76.16.7976-7986.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Lai C-C, Henningson C, Dimaio D. Bovine papillomavirus E5 protein induces oligomerization and trans-phosphorylation of the platelet-derived growth factor β receptor. Proceedings of the National Academy of Sciences of the United States of America. 1998;95(26):15241–15246. doi: 10.1073/pnas.95.26.15241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Klein O, Kegler-Ebo D, Su J, Smith S, DiMaio D. The bovine papillomavirus E5 protein requires a juxtamembrane negative charge for activation of the platelet-derived growth factor β receptor and transformation of C127 cells. Journal of Virology. 1999;73(4):3264–3272. doi: 10.1128/jvi.73.4.3264-3272.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Petti LM, Ricciardi EC, Page HJ, Porter KA. Transforming signals resulting from sustained activation of the PDGFβ receptor in mortal human fibroblasts. Journal of Cell Science. 2008;121(8):1172–1182. doi: 10.1242/jcs.018713. [DOI] [PubMed] [Google Scholar]
- 23.Roperto S, Borzacchiello G, Esposito I, et al. Productive infection of bovine papillomavirus type 2 in the placenta of pregnant cows affected with urinary bladder tumors. PLoS ONE. 2012;7(3) doi: 10.1371/journal.pone.0033569.e33569 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Borzacchiello G, Mogavero S, De Vita G, Roperto S, Della Salda L, Roperto F. Activated platelet-derived growth factor β receptor expression, PI3K-AKT pathway molecular analysis, and transforming signals in equine sarcoids. Veterinary Pathology. 2009;46(4):589–597. doi: 10.1354/vp.08-VP-0191-B-FL. [DOI] [PubMed] [Google Scholar]
- 25.Corteggio A, Di Geronimo O, Roperto S, Roperto F, Borzacchiello G. Activated platelet-derived growth factor β receptor and Ras-mitogen-activated protein kinase pathway in natural bovine urinary bladder carcinomas. Veterinary Journal. 2012;191(3):393–395. doi: 10.1016/j.tvjl.2011.03.019. [DOI] [PubMed] [Google Scholar]
- 26.Carnero A. The PKB/AKT pathway in cancer. Current Pharmaceutical Design. 2010;16(1):34–44. doi: 10.2174/138161210789941865. [DOI] [PubMed] [Google Scholar]
- 27.Moeller SJ, Head ED, Sheaff RJ. p27Kip1 inhibition of GRB2-SOS formation can regulate Ras activation. Molecular and Cellular Biology. 2003;23(11):3735–3752. doi: 10.1128/MCB.23.11.3735-3752.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Dance M, Montagner A, Salles J-P, Yart A, Raynal P. The molecular functions of Shp2 in the Ras/Mitogen-activated protein kinase (ERK1/2) pathway. Cellular Signalling. 2008;20(3):453–459. doi: 10.1016/j.cellsig.2007.10.002. [DOI] [PubMed] [Google Scholar]
- 29.Lokker NA, Sullivan CM, Hollenbach SJ, Israel MA, Giese NA. Platelet-derived growth factor (PDGF) autocrine signaling regulates survival and mitogenic pathways in glioblastoma cells: evidence that the novel PDGF-C and PDGF-D ligands may play a role in the development of brain tumors. Cancer Research. 2002;62(13):3729–3735. [PubMed] [Google Scholar]
- 30.Druker BJ, Tamura S, Buchdunger E, et al. Effects of a selective inhibitor of the Ab1 tyrosine kinase on the growth of Bcr-Ab1 positive cells. Nature Medicine. 1996;2(5):561–566. doi: 10.1038/nm0596-561. [DOI] [PubMed] [Google Scholar]
- 31.Buchdunger E, Zimmermann J, Mett H, et al. Inhibition of the Abl protein-tyrosine kinase in vitro and in vivo by a 2- phenylaminopyrimidine derivative. Cancer Research. 1996;56(1):100–104. [PubMed] [Google Scholar]
- 32.Buchdunger E, Cioffi CL, Law N, et al. Abl protein-tyrosine kinase inhibitor STI571 inhibits in vitro signal transduction mediated by c-Kit and platelet-derived growth factor receptors. The Journal of Pharmacology and Experimental Therapeutics. 2000;295(1):139–145. [PubMed] [Google Scholar]
- 33.Kilic T, Alberta JA, Zdunek PR, et al. Intracranial inhibition of platelet-derived growth factor-mediated glioblastoma cell growth by an orally active kinase inhibitor of the 2-phenylaminopyrimidine class. Cancer Research. 2000;60(18):5143–5150. [PubMed] [Google Scholar]
- 34.Rubin BP, Schuetze SM, Eary JF, et al. Molecular targeting of platelet-derived growth factor B by imatinib mesylate in a patient with metastatic dermatofibrosarcoma protuberans. Journal of Clinical Oncology. 2002;20(17):3586–3591. doi: 10.1200/JCO.2002.01.027. [DOI] [PubMed] [Google Scholar]
- 35.Sjöblom T, Shimizu A, O’Brien KP, et al. Growth inhibition of dermatofibrosarcoma protuberans tumors by the platelet-derived growth factor receptor antagonist STI571 through induction of apoptosis. Cancer Research. 2001;61(15):5778–5783. [PubMed] [Google Scholar]
- 36.McGary EC, Weber K, Mills L, et al. Inhibition of platelet-derived growth factor-mediated proliferation of osteosarcoma cells by the novel tyrosine kinase inhibitor STI571. Clinical Cancer Research. 2002;8(11):3584–3591. [PubMed] [Google Scholar]
- 37.Gilbert RE, Kelly DJ, McKay T, et al. PDGF signal transduction inhibition ameliorates experimental mesangial proliferative glomerulonephritis. Kidney International. 2001;59(4):1324–1332. doi: 10.1046/j.1523-1755.2001.0590041324.x. [DOI] [PubMed] [Google Scholar]
- 38.Druker BJ, Lydon NB. Lessons learned from the development of an Abl tyrosine kinase inhibitor for chronic myelogenous leukemia. The Journal of Clinical Investigation. 2000;105(1):3–7. doi: 10.1172/JCI9083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Mauro MJ, O’Dwyer M, Heinrich MC, Druker BJ. STI571: a paradigm of new agents for cancer therapeutics. Journal of Clinical Oncology. 2002;20(1):325–334. doi: 10.1200/JCO.2002.20.1.325. [DOI] [PubMed] [Google Scholar]
- 40.Krystal GW, Honsawek S, Litz J, Buchdunger E. The selective tyrosine kinase inhibitor STI571 inhibits small cell lung cancer growth. Clinical Cancer Research. 2000;6(8):3319–3326. [PubMed] [Google Scholar]
- 41.van Oosterom AT, Judson I, Verweij J, et al. Safety and efficacy of imatinib (STI571) in metastatic gastrointestinal stromal tumours: a phase I study. The Lancet. 2001;358(9291):1421–1423. doi: 10.1016/s0140-6736(01)06535-7. [DOI] [PubMed] [Google Scholar]
- 42.van Oosterom AT, Judson IR, Verweij J, et al. Update of phase I study of imatinib (STI571) in advanced soft tissue sarcomas and gastrointestinal stromal tumors: a report of the EORTC Soft Tissue and Bone Sarcoma Group. European Journal of Cancer. 2002;38(supplement 5):S83–S87. doi: 10.1016/s0959-8049(02)80608-6. [DOI] [PubMed] [Google Scholar]
- 43.Isotani M, Ishida N, Tominaga M, et al. Effect of tyrosine kinase inhibition by imatinib mesylate on mast cell tumors in dogs. Journal of Veterinary Internal Medicine. 2008;22(4):985–988. doi: 10.1111/j.1939-1676.2008.00132.x. [DOI] [PubMed] [Google Scholar]
- 44.Marconato L, Bettini G, Giacoboni C, et al. Clinicopathological features and outcome for dogs with mast cell tumors and bone marrow involvement. Journal of Veterinary Internal Medicine. 2008;22(4):1001–1007. doi: 10.1111/j.1939-1676.2008.0128.x. [DOI] [PubMed] [Google Scholar]
- 45.Yamada O, Kobayashi M, Sugisaki O, et al. Imatinib elicited a favorable response in a dog with a mast cell tumor carrying a c-kit c.1523A>T mutation via suppression of constitutive KIT activation. Veterinary Immunology and Immunopathology. 2011;142(1-2):101–106. doi: 10.1016/j.vetimm.2011.04.002. [DOI] [PubMed] [Google Scholar]
- 46.Lachowicz JL, Post GS, Brodsky E. A phase I clinical trial evaluating imatinib mesylate (Gleevec) in tumor-bearing cats. Journal of Veterinary Internal Medicine. 2005;19(6):860–864. doi: 10.1892/0891-6640(2005)19[860:apicte]2.0.co;2. [DOI] [PubMed] [Google Scholar]
- 47.Katayama R, Huelsmeyer MK, Marr AK, Kurzman ID, Thamm DH, Vail DM. Imatinib mesylate inhibits platelet-derived growth factor activity and increases chemosensitivity in feline vaccine-associated sarcoma. Cancer Chemotherapy and Pharmacology. 2004;54(1):25–33. doi: 10.1007/s00280-004-0780-7. [DOI] [PubMed] [Google Scholar]
- 48.Yuan ZQ, Gault EA, Gobeil P, Nixon C, Campo MS, Nasir L. Establishment and characterization of equine fibroblast cell lines transformed in vivo and in vitro by BPV-1: Model systems for equine sarcoids. Virology. 2008;373(2):352–361. doi: 10.1016/j.virol.2007.11.037. [DOI] [PubMed] [Google Scholar]
- 49.Yuan Z, Gault EA, Saveria Campo M, Nasir L. Different contribution of bovine papillomavirus type 1 oncoproteins to the transformation of equine fibroblasts. The Journal of General Virology. 2011;92(4):773–783. doi: 10.1099/vir.0.028191-0. [DOI] [PubMed] [Google Scholar]
- 50.Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;227(5259):680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- 51.Borzacchiello G, Russo V, Spoleto C, et al. Bovine papillomavirus type-2 DNA and expression of E5 and E7 oncoproteins in vascular tumours of the urinary bladder in cattle. Cancer Letters. 2007;250(1):82–91. doi: 10.1016/j.canlet.2006.09.022. [DOI] [PubMed] [Google Scholar]
- 52.Silva MA, Altamura G, Corteggio A, et al. Expression of connexin 26 and bovine papillomavirus E5 in cutaneous fibropapillomas of cattle. The Veterinary Journal. 2013;195(3):337–343. doi: 10.1016/j.tvjl.2012.07.009. [DOI] [PubMed] [Google Scholar]
- 53.Campo MS, McCaffery RE, Doherty I, Kennedy IM, Jarrett WFH. The Harvey ras 1 gene is activated in papillomavirus-associated carcinomas of the upper alimentary canal in cattle. Oncogene. 1990;5(3):303–308. [PubMed] [Google Scholar]
- 54.Ghai J, Ostrow RS, Tolar J, et al. The E5 gene product of rhesus papillomavirus is an activator of endogenous Ras and phosphatidylinositol-3′-kinase in NIH 3T3 cells. Proceedings of the National Academy of Sciences of the United States of America. 1996;93(23):12879–12884. doi: 10.1073/pnas.93.23.12879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Zago M, Campo MS, O’Brien V. Cyclin A expression and growth in suspension can be uncoupled from p27 deregulation and extracellular signal-regulated kinase activity in cells transformed by bovine papillomavirus type 4 E5. The Journal of General Virology. 2004;85(12):3585–3595. doi: 10.1099/vir.0.80313-0. [DOI] [PubMed] [Google Scholar]
- 56.Corteggio A, Di Geronimo O, Roperto S, Roperto F, Borzacchiello G. Bovine papillomavirus E7 oncoprotein binds to p600 in naturally occurring equine sarcoids. The Journal of General Virology. 2011;92(2):378–382. doi: 10.1099/vir.0.025866-0. [DOI] [PubMed] [Google Scholar]
- 57.Song SH, Lee JK, Hur JY, Kim I, Saw HS, Park YK. The expression of epidermal growth factor receptor, vascular endothelial growth factor, matrix metalloproteinase-2, and cyclooxygenase-2 in relation to human papilloma viral load and persistence of human papillomavirus after conization with negative margins. International Journal of Gynecological Cancer. 2006;16(6):2009–2017. doi: 10.1111/j.1525-1438.2006.00727.x. [DOI] [PubMed] [Google Scholar]
- 58.Golub TR, Barker GF, Lovett M, Gary Gilliland D. Fusion of PDGF receptor β to a novel ets-like gene, tel, in chronic myelomonocytic leukemia with t(5;12) chromosomal translocation. Cell. 1994;77(2):307–316. doi: 10.1016/0092-8674(94)90322-0. [DOI] [PubMed] [Google Scholar]
- 59.Heinrich MC, Corless CL, Duensing A, et al. PDGFRA activating mutations in gastrointestinal stromal tumors. Science. 2003;299(5607):708–710. doi: 10.1126/science.1079666. [DOI] [PubMed] [Google Scholar]
- 60.Hirota S, Ohashi A, Nishida T, et al. Gain-of-function mutations of platelet-derived growth factor receptor α gene in gastrointestinal stromal tumors. Gastroenterology. 2003;125(3):660–667. doi: 10.1016/s0016-5085(03)01046-1. [DOI] [PubMed] [Google Scholar]
- 61.Hirota S. Gain-of-function mutation of c-kit gene and molecular target therapy in GISTs. Japanese Journal of Gastroenterology. 2003;100(1):13–20. [PubMed] [Google Scholar]
- 62.Sirvent N, Maire G, Pedeutour F. Genetics of dermatofibrosarcoma protuberans family of tumors: from ring chromosomes to tyrosine kinase inhibitor treatment. Genes Chromosomes and Cancer. 2003;37(1):1–19. doi: 10.1002/gcc.10202. [DOI] [PubMed] [Google Scholar]
- 63.Giantin M, Vascellari M, Morello EM, et al. c-KIT messenger RNA and protein expression and mutations in canine cutaneous mast cell tumors: correlations with post-surgical prognosis. Journal of Veterinary Diagnostic Investigation. 2012;24(1):116–126. doi: 10.1177/1040638711425945. [DOI] [PubMed] [Google Scholar]
- 64.Matsumura I, Mizuki M, Kanakura Y. Roles for deregulated receptor tyrosine kinases and their downstream signaling molecules in hematologic malignancies. Cancer Science. 2008;29(3):479–485. doi: 10.1111/j.1349-7006.2007.00717.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Burkhardt A, Willingham M, Gay C, Jeang K-T, Schlegel R. The E5 oncoprotein of bovine papillomavirus is oriented asymmetrically in Golgi and plasma membranes. Virology. 1989;170(1):334–339. doi: 10.1016/0042-6822(89)90391-7. [DOI] [PubMed] [Google Scholar]
- 66.Talbert-Slagle K, DiMaio D. The bovine papillomavirus E5 protein and the PDGF β receptor: it takes two to tango. Virology. 2009;384(2):345–351. doi: 10.1016/j.virol.2008.09.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.London CA. Tyrosine kinase inhibitors in veterinary medicine. Topics in Companion Animal Medicine. 2009;24(3):106–112. doi: 10.1053/j.tcam.2009.02.002. [DOI] [PubMed] [Google Scholar]
- 68.Nilsson J, Thyberg J, Heldin CH, Westermark B, Wasteson A. Surface binding and internalization of platelet-derived growth factor in human fibroblasts. Proceedings of the National Academy of Sciences of the United States of America. 1983;80(181):5592–5596. doi: 10.1073/pnas.80.18.5592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Goldstein JL, Brown MS, Anderson RG, Russell DW, Schneider WJ. Receptor-mediated endocytosis: concepts emerging from the LDL receptor system. Annual Review of Cell Biology. 1985;1:1–39. doi: 10.1146/annurev.cb.01.110185.000245. [DOI] [PubMed] [Google Scholar]