

Summary
Cell migration is an important biological process which has been intensively studied in the past decades. Numerous techniques, mainly involving two-dimensional cell culture systems, have contributed to dissecting the essential mechanisms underlying this process. However, the development of three-dimensional cell culture and in vivo systems has shown some differences with what was previously believed to be well-established cell migration mechanisms, suggesting that two-dimensional cell motility would be a poor predictor of in vivo behaviour. Drosophila is a widely recognized model organism to study developmental and homeostatic processes and has been widely used to investigate cell migration. Here, we focus on the migration of small groups of pupal hemocytes that accumulate during larval stages in dorsal patches. We show that integrins, and other known nascent adhesion-related proteins such as Rhea and Fermitin 1, are crucial for this process and that their depletion does not affect polarization in response to environmental cues. We also present evidence for the importance of adhesion maturation-related proteins in hemocyte migration, namely Zyxin. Zyxin depletion in hemocytes leads to a significant increase of cell speed without affecting their response to a chemotactic cue. This is the first report of a systematic analysis using Drosophila melanogaster hemocytes to study adhesion-related proteins and their function in cell migration in vivo. Our data point to mechanisms of cell migration similar to those described in three-dimensional in vitro systems and other in vivo model organisms.
Keywords: Drosophila, Hemocyte, Integrin, Migration
Introduction
Cell migration is a key mechanism that occurs during developmental and homeostatic processes such as tissue repair, immune surveillance and morphogenesis. The basic mechanisms underlying cell migration have been intensively studied in the past decades using different two-dimensional (2D) in vitro methods. These have allowed researchers to establish a model for cell migration that shows that an intermediate strength of adhesion is required for maximal cell speed (DiMilla et al., 1991; Palecek et al., 1997). This suggested that weak cell-adhesive interactions with the substratum would not provide enough traction and at strong adhesion, the cell would be too attached to the substratum to move efficiently. Interestingly, recent findings using three-dimensional (3D) in vitro and recently developed in vivo techniques have highlighted new features and intriguing differences in cell behaviour in comparison to what was previously described using 2D in vitro systems, namely in terms of cell morphology and signalling pathways controlling the process of cell migration (Baker and Chen, 2012; Beerling et al., 2011; Toetsch et al., 2009). As such, more and more researchers are turning to in vivo model systems (believed to be better mimicked by 3D in vitro systems) to clarify important aspects regarding cell migration.
Drosophila melanogaster has been a model organism of choice to look at cell migration in vivo. Border cell migration or epithelial cell migration, are some of best studied examples of collective cell migration (Montell et al., 2012; Weijer, 2009). To study single cell migration, immune circulatory cells, also known as hemocytes, which are equivalent to mammalian leukocytes in terms of functions and features, have emerged as a very useful model (Tirouvanziam et al., 2004; Wood and Jacinto, 2007).
Integrins are well-known transmembrane adhesion receptors involved in cell–extracellular matrix (ECM) interactions and play critical roles in cell signalling. They are one of the core components of adhesion sites where the ECM connects to the cellular cytoskeleton (Huttenlocher and Horwitz, 2011; Zaidel-Bar et al., 2007). The first step in adhesion establishment is the formation of nascent adhesions (NA), also referred to as focal complexes, which can then either disassemble or mature into focal adhesions (FA). The latter can undergo an additional maturation step which will give rise to fibrillar adhesions. In a study by Zaidel-Bar et al., the dynamics of the formation and maturation of these adhesion structures has been reported and some central differences in their composition have been shown (Zaidel-Bar et al., 2003). Although adhesion structures have been intensively studied in both 2D and more recently 3D in vitro systems (Fraley et al., 2010; Tolde et al., 2012), very little is known about their existence, formation and roles in an in vivo context. In Drosophila, the integrin family consists of five αPS subunits (αPS1 to 5) and two β subunits, βν and βPS, the latter also known as myospheroid (Brown et al., 2000). The only previously reported links between hemocytes and integrins in Drosophila demonstrated that integrins act downstream of the GEF Dizzy and the Rap1 small GTPase to control both hemocyte shape changes and allow invasive migration during early embryonic development (Huelsmann et al., 2006; Siekhaus et al., 2010).
In this paper, we show that hemocytes that are located in dorsal patches, and are sessile during larval stages, depend on integrins and other known adhesion-related proteins to migrate after pupal formation. Whereas depletion of proteins described to be involved in NA formation leads to a decrease in hemocyte speed without affecting cell polarization, depletion of a recognized adhesion maturation-related protein increases cell speed. These findings suggest that the degree of maturation of the adhesions alters cell migration speed in a biphasic manner thus providing new molecular insights to further understand the biphasic model proposed by DiMilla et al., where an optimal adhesion strength maximises cell speed (DiMilla et al., 1991).
Results
Mutations in the Drosophila βPS integrin subunit, myospheroid, lead to hemocyte migration defects
During late 3rd instar larvae and early white pre-pupal stages of Drosophila melanogaster development, a dorsal population of hemocytes, hereupon referred to as dorsal patch-hemocytes, can be found attached to the dorsal epithelium aligned in periodic patches along the dorsal vessel (Lanot et al., 2001; Makhijani et al., 2011; Stofanko et al., 2008; Zettervall et al., 2004). During this period, hemocytes maintain a round morphology, a low migration speed and are unresponsive upon wounding of the epithelium (Babcock et al., 2008) in contrast to the hemocytes in embryonic (Stramer et al., 2005) or late pupal stages (Moreira et al., 2011). However, by 2 hours after pupa formation (APF), the dorsal sub-epidermal hemocytes have acquired a spread morphology with filopodium and lamellipodium-like structures and collectively initiate random single cell migration (supplementary material Movie 1).
Based on the MARCM (Mosaic Analysis with a Repressible Cell Marker) system described by Wu and Luo (Wu and Luo, 2007), we established a protocol (DEMON – Deleted in Hemocytes Only) that allows to test the effects of homozygous lethal mutations in hemocytes only, in an otherwise phenotypic wild-type heterozygote background (Fig. 1A,B). Using this method we were able to generate GFP-positive wild-type and myospheroid mutant (mys1) hemocytes in the dorsal patches area (Fig. 1C), with a maximum of three GFP-positive cells in the selected region.
Fig. 1. Myospheroid is required for proper hemocyte migration.
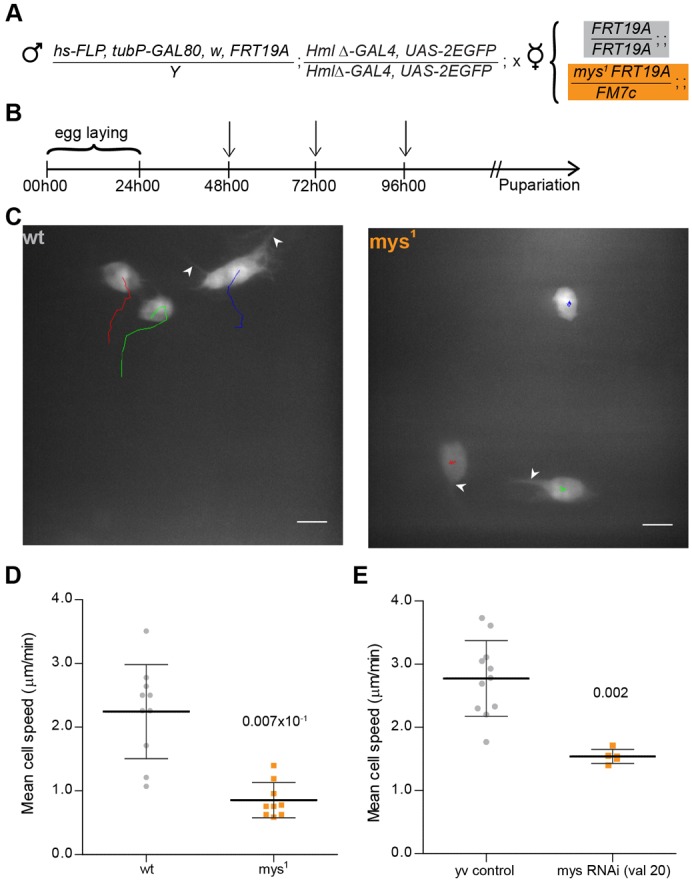
(A) Outline of the MARCM protocol in hemocytes. Cross between DEMON males and FRT19a control or mys1 mutant virgin female flies for MARCM analysis. (B) Crosses are placed at 25°C for 24 hours. The progeny is submitted to three 1 hour heat-shocks (indicated by arrows) at 37°C before selection of 3rd instar females containing GFP-expressing hemocytes. (C) Movement of wild-type and mys1 GFP-expressing hemocytes in 3 to 4 hour APF flies, tracked for 12 minutes (1 min time-lapse interval). Arrowheads indicate filopodium-like protrusions produced by both wild-type and mys1 mutant hemocytes. Scale bars: 10 µm. (D) Graph showing individual mean cell speed for wild-type and mys1 clones (2 to 4 hours APF). (E) Graph showing average mean cell speeds for yv control and mys (valium 20) RNAi-expressing flies (between 3 to 4 hours APF). Mann–Whitney tests for non-Gaussian populations were used. Black lines indicate the samples' means; Error bars = standard deviation; P-values shown above tested groups.
Between 2 and 4 hours APF, mys1 hemocytes present striking differences in terms of cell movement when compared to controls (Fig. 1C,D): during this period, both wild-type and mys1 hemocytes extend filopodium-like protrusions, typical of migrating cells. In terms of cell displacement however, it was evident that mys1 hemocytes show an aberrant migratory behaviour. Calculation of individual cell speeds shows a significant difference between controls and mys1 hemocytes with the former reaching an average speed of 2.24±0.74 µm.min−1 whereas the latter had a residual calculated mean cell speed of 0.86±0.28 µm.min−1. The movement in the mys1 hemocytes is likely to be a consequence of the constant wiggling and cell shape changes that lead to permanent adjustments in the midpoint of the cell (supplementary material Movie 2). This result led us to hypothesize that, like in in vitro systems, other known adhesion components besides integrins, could play a role in dorsal patch-hemocyte migration upon pupal formation.
To test this hypothesis we decided to use the RNAi based methodology combined with the UAS/GAL4 system (Duffy, 2002) to specifically knockdown the genes of interest in hemocytes. Initially, we established that knockdown of myospheroid led to a significant decrease in cell speed (mean cell speed 1.54±0.11 µm.min−1) when compared to wild-type (mean cell speed 2.77±0.60 µm.min−1) in a similar manner as observed in the mys1 hemocytes (Fig. 1E). This ensured that the RNAi based approach was suitable and sufficient for identifying anomalies in hemocyte migratory behaviour upon knockdown of genes of interest (supplementary material Movie 4). It is important to notice that in the case of RNAi knockdown some migration was still detected, compared to mys1 mutant hemocytes, probably because this method does not result in a complete mys loss of function.
Knockdown of integrins, integrin-activating and other adhesion-associated proteins affects hemocyte migration
To understand the role of other integrin-related proteins known to be involved in the formation of NA (Choi et al., 2008; Zaidel-Bar et al., 2003) we decided to follow a candidate gene approach based protocol for specifically knocking down the genes of interest. Between 3 and 4 hours APF, whereas control hemocytes are already widely scattered cells, myospheroid deficient hemocytes remain closely clustered together forming a densely packed group of single cells (Fig. 2A; supplementary material Movies 3, 4). Two independent mys RNAi fly lines were used for comparison: a significant decrease in cell migration speed was observed with both (1.54±0.11 µm.min−1 and 1.52±0.56 µm.min−1 for the valium 20 and the valium 10 lines, respectively) (Fig. 2B). It is also interesting to notice that the patches form independently of the integrin knockdown.
Fig. 2. NA and FA proteins are essential for optimal hemocyte migration.
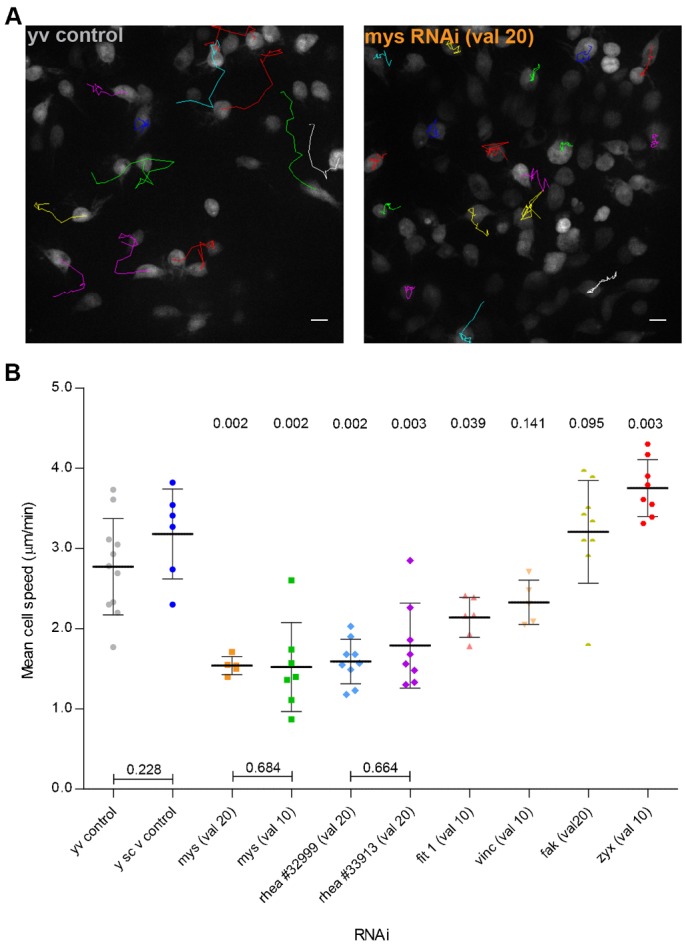
(A) Example tracks of yv controls and mys RNAi-expressing flies. Scale bars: 10 µm. (B) Graph showing average mean cell speeds for different genetic backgrounds in 3 to 4 hour APF flies. val 10 = TRIP valium 10; val 20 = TRIP valium 20. Mann–Whitney tests for non-Gaussian populations were used. Both rhea RNAi lines were compared to the y1sc1v1 control line. All other lines were compared to the y1v1 control line. Black lines indicate the samples' means; Error bars = standard deviation; P-values shown above tested groups.
Depletion of defined integrin-activating proteins such as Rhea (Talin homologue) or Fermitin 1 (Kindlin 1 homologue) (Anthis et al., 2009; Larjava et al., 2008) led to significant decreases in the mean cell speed (Fig. 2B; supplementary material Movies 5, 6). rhea knockdown reduces hemocyte cell speed to values comparable to the ones observed with the mys knockdown hemocytes (1.80±0.52 µm.min−1 and 1.59±0.21 µm.min−1 for the 33913 and the 32999 lines, respectively). fermitin 1 knockdown, although to a lesser extent than mys and rhea, also causes hemocyte migratory defects, with the latter reaching average speeds of 2.14±0.25 µm.min−1.
Depletion of either Vinculin or Focal Adhesion Kinase (FAK), both described to be involved in early adhesion formation (Deramaudt et al., 2011; Zaidel-Bar et al., 2003), has no significant effect on hemocyte migration in vivo (Fig. 2B; supplementary material Movies 6, 7). These results correlate with previous findings which suggested that either molecule is non-essential for Drosophila's viability (Alatortsev et al., 1997; Grabbe et al., 2004).
Following the stabilization of a NA, which in part involves the organized recruitment and assembly of the above discussed proteins, adhesion structures can develop into a FA, a process that relies on several factors, including tension which depends on myosin II activity (Parsons et al., 2010). This maturation process is accompanied by an increase in the number and diversity of proteins that assemble at adhesion sites, like zyxin. As such, we decided to investigate the effect of knocking down zyxin expression during hemocyte migration. Interestingly, zyxin knocked down hemocytes showed a significant increase in the mean cell speed (3.75±0.36 µm.min−1) when compared to controls (Fig. 2B; supplementary material Movie 7), a phenotype which has been previously described in in vitro cultured cells (Fraley et al., 2010; Hoffman et al., 2006).
Polarization in response to wounds is not affected by the knockdown of NA related proteins
Integrins also appear as key molecules in the establishment and maintenance of a polarity axis in directionally migrating cells; therefore we decided to analyse their role in hemocyte polarization and migration upon epithelial wounding.
Three to four hours APF, the hemocytes in the dorsal patches area show a random migratory pattern with a filopodial protrusion extending in the direction of the migration. Upon wounding, part of the nearby hemocyte population responds by extending the filopodial protrusion towards the wound site (Fig. 3A; supplementary material Movie 8). Quantification of the number of hemocytes at the wound site over time in a control situation suggests that within the first 30 minutes after wounding, a first wave of rapidly responding hemocytes reaches the wound site (Fig. 3B). Over the next few minutes, some cells still respond to the chemotactic cue although the rate at which they reach the area of interest is diminished compared to the initial response phase. This behaviour is enhanced in zyxin knockdown hemocytes thus suggesting that blocking adhesion maturation does not affect hemocyte chemotaxis. For myospheroid and rhea-depleted hemocytes the response to a wound was less perceptible since only a few cells ever reach the area of interest during the time of analysis. However, since blocking the initial formation of NA leads to striking migratory defects, we decided to further characterize the hemocytes' response to a chemotactic cue: for that, we analysed the mean angle of the hemocytes' polarization arm relative to the wound before and after wounding (Fig. 3C). In the control population, we calculated a significant change in the mean angle of the polarization arm relative to the wound site. Interestingly, the knockdown of myospheroid, rhea or fermitin 1 had no impact in the extension of the filopodial protrusion towards wounds: as for controls, in all situations, there was a shift to lower values of the mean angle of polarization, which is indicative of the arm pointing towards the wound site. This was observed independently of the capacity of hemocytes to migrate towards wounds (Fig. 3C; supplementary material Movies 9, 10, 11), thus suggesting that integrin-containing adhesions, although crucial for hemocyte migration, are not necessary for the establishment of polarity upon wounding.
Fig. 3. Polarization towards a chemotactic cue does not depend on NA proteins.

(A) yv control flies immediately before and 28 minutes after wounding, respectively. Zoom of a selected cell presented at the top right hand corners, showing changes in the polarization arm. Red arrows point in the direction of the polarization arm of cells at the pictured time point. Green arrows point towards wound center (depicted by a white cross). Scale bars: 10 µm. (B) Graph showing the mean hemocyte numbers at the wound area along time for different control and RNAi flies. Dotted line under x axis depicts the chosen time interval for the polarization analysis. (C) Graph showing the mean polarization angle for individual cells before and after wounding in 3 to 4 hour APF flies. val 10 = TRIP valium 10; val 20 = TRIP valium 20. Black lines indicate the samples' means; Error bars = standard deviation; P-values shown above tested groups.
In opposition to what happens with NA-related proteins, zyxin-depleted hemocytes showed no significant changes between before and after wounding in terms of the mean polarization angle (Fig. 3C; supplementary material Movie 12) which could suggest that zyxin knocked down hemocytes failed to reorient themselves towards the wound and/or were less sensitive to environmental cues. However, we have shown that the response of zyxin-depleted hemocytes occurs more rapidly than in a control situation thus probably making it a lot more difficult to detect shifts in the mean polarization angle as in all other analysed situations.
Discussion
In this paper, we have looked at cell migration in vivo using Drosophila dorsal patches-hemocytes as our model system. These patches of immune cells are found attached to the dorsal epithelium aligned along the dorsal vessel of the late 3rd instar and young white pre-pupas (Lanot et al., 2001; Stofanko et al., 2008). The origin of these tightly packed groups of cells is partially unknown, but it has been suggested that these serve as a major hematopoietic compartment in the larva that can be activated upon an immune challenge (Márkus et al., 2009). How these cells remain attached to the epithelium prior to the onset of metamorphosis is also not clear. Our results suggest that integrins (more specifically myospheroid, Drosophila's main beta integrin subunit) and integrin-containing adhesions play no role in the original attachment of these hemocytes to the dorsal patches areas, thus indicating that alternative adhesions systems may also be involved in hemocyte attachment to these locations.
Interestingly, in spite of the numerous similarities between Drosophila hemocytes and mammalian leukocytes, we seem to have found what appears to be a striking difference in terms of their migratory behaviour: whereas vertebrate immune cells can move in complex in vivo, ex vivo and 3D in vitro environments in a integrin-independent fashion, using the force of the actin-network expansion (Lämmermann et al., 2008), Drosophila dorsal patch-hemocytes are dependent on integrin-containing adhesions to migrate. Hemocytes seem to therefore fit a more conventional model which describes integrin-containing adhesions as essential for cell migration (Huttenlocher and Horwitz, 2011). Besides myospheroid, we have also shown that rhea (talin homologue) and fermitin 1 (kindlin 1 homologue) depleted hemocytes suffer from similar migratory defects, both showing significant decreases in the mean cell speed, a phenotype that has been previously observed in HT-1080 talin-depleted cells in a 3D in vitro assay (Fraley et al., 2010). Also, both Talin and Kindlin 1 have been shown to bind directly to the β integrin cytoplasmic tail causing integrin conformational changes and promoting integrin activation (Harburger et al., 2009; Ye et al., 2011). Our data suggest that a similar integrin activation mechanism could occur in migrating hemocytes in vivo and that depletion of either molecule could impair the formation of NAs.
Vinculin is another core component of NAs (Zaidel-Bar et al., 2003). Vinculin binds directly to talin and the actin cytoskeleton (Humphries et al., 2007) thus acting both as a bridge and a force-transmitter (Grashoff et al., 2010). Disruption of vinculin in 2D cultured cells was shown to play a critical role in cell migration, leading to a significant increase of cell speed (Coll et al., 1995). However, more recently it was also shown that in a 3D environment, vinculin depletion decreases cell motility, a phenotype that was associated to a decrease in adhesion strength and lowering of the traction forces (Mierke et al., 2010). Our results in vivo show that vinculin depleted hemocytes suffer no significant changes in terms of cell speed when compared to controls, which is in line with previous findings by Alatortsev et al., who have shown that vinculin is non-essential for Drosophila's survival and development (Alatortsev et al., 1997). As vinculin is one of the main force transmitters identified in integrin-containing adhesions, it has yet to be clarified whether other molecules could be playing this role in migrating hemocytes.
Focal Adhesion Kinase (FAK) is another molecule which plays a critical role in both NA formation and adhesion turnover, depending on the state of phosphorylation (Deramaudt et al., 2011). In mice, FAK depletion causes numerous defects that ultimately lead to embryonic death (Ilić et al., 2004). In Drosophila, fak expression was shown to be non-essential, and the absence of FAK has no effect on integrin-dependent mechanisms nor does it seem to influence border cell migration (Grabbe et al., 2004). Similarly, we observe that fak knockdown had no effect on hemocyte migration. fak over-expression however, has been shown to act negatively on integrin adhesion in Drosophila, and is potentially critical for adhesion remodelling. It would therefore be interesting to see if any migration defects would also occur upon over-expression of fak in hemocytes.
It is known that upon stabilization, NAs can overcome several maturation steps which imply the arrival of adhesion maturation-related proteins such as Zyxin to the adhesion sites. Zyxin is a stretch-sensitive mechanosensor (Yoshigi et al., 2005) important for the maintenance and repair of actin stress fibers (Smith et al., 2010) and it has been described as a molecular hallmark of the transition from an NA to an FA (Zaidel-Bar et al., 2003). We have shown that zyxin depletion enhances hemocyte migration speed, a phenotype which has already been shown by others and was correlated with an enhanced adhesion capacity that did not affect integrin expression levels (Fraley et al., 2010; Hoffman et al., 2006). Further analysis should elucidate whether adhesion strength and/or myospheroid expression are affected in zyxin-depleted hemocytes. Our results therefore suggest that zyxin knockdown may prevent the transition of NAs into FAs as in cell culture studies. Zyxin may be involved in the maintenance of hemocyte integrin activity to regulate migration. This is similar to the biphasic model (DiMilla et al., 1991), where an intermediate strength of cell–substratum interaction is essential for maximal cell migration. It is possible that the disruption of initial NA formation decreases cell adhesion and leads to a decrease in cell speed due to a lack of the necessary traction for migration. On the other hand, disrupting the maturation of FAs (stronger adhesive structures than NAs) decreases what should be a stronger cell–substratum interaction, and therefore causes an increase in cell migration. This suggests that cell adhesion maturation may be an important factor for controlling cell speed.
Immune cell migration, besides being a naturally occurring developmental process, can also be triggered by external environmental cues such as wounds or infections (Niethammer et al., 2009). Migration, a complex multi-step process, starts with the formation of an internal cellular asymmetry of molecules and structures that allows the establishment of a front–rear cell polarity that eventually translates itself in a directional movement towards the site of interest (Bornens, 2008; Li and Gundersen, 2008; Mellman and Nelson, 2008; Stramer et al., 2010). Amongst the many molecules involved in the establishment of this polarity axis are the integrins. Using a peviously described migration assay (Moreira et al., 2011), we were able to conclude that, in hemocytes, integrins and other NA-related proteins are not essential for the establishment of a polarization axis towards a wound. This is not the first report of immune cells responding to environmental cues in an integrin-independent fashion (Lämmermann et al., 2008), which implies that other receptors apart from integrins are functioning in detecting environmental changes and in establishing a front–rear polarity axis in dorsal patch-hemocytes.
The polarization results obtained upon zyxin knockdown suggested that zyxin-depleted hemocytes could be less sensitive to environmental cues, similar to what was described for zyxin-depleted fibroblasts in a haptotatic migration assay towards different integrin ligands (Hoffman et al., 2006). However, we have shown that zyxin-depleted hemocytes still reached the wound area at a higher rate than control cells within the first 30 minutes post wounding. Altogether, the data suggest that zyxin depletion solely affects cell migration speed without affecting a cell's response to a chemotactic cue.
Ultimately, our results clarify the importance of a tight regulation of adhesion formation and maturation for optimal cell migration in an in vivo context, which, similarly to the previously described biphasic model by DiMilla et al. based on cell adhesion (DiMilla et al., 1991), suggests that integrin-containing adhesions formation and maturation is the major force controlling hemocyte migration in vivo.
Materials and Methods
Fly stocks
w; hmlΔGAL4,UAS-GFP; (Sinenko and Mathey-Prevot, 2004) was used to visualize dorsal patches-hemocytes and to drive the expression of other UAS constructs in hemocytes specifically. The choice of a truncated version of the original hml driver by Goto et al. (Goto et al., 2001) relied on the hemocyte-specific pattern of expression that recovers all the wild-type hemocyte specific characteristics that had been lost in the original construct. To generate the DEMON stock the following stocks from the Bloomington Stock Center were used: hsFLP, tubP-GAL80, w*, FRT19A; Pin/CyO; (BL# 5133), FRT19A;; (BL# 1709), mys1FRT19A/FM7c;; (BL# 23862). For the migration and polarization analysis the following lines were used: y1v1 (BL# 1509) and y1sc1v1 (BL# 25710) were used as controls; myospheroid (valium 10) (BL# 27735), myospheroid (valium 20) (BL# 33642), rhea (valium 20) (BL# 33913), rhea (valium 20) (BL# 32999), zyxin (valium 10) (BL# 29591), vinculin (valium 10) (BL# 25965), fermitin 1 (valium 10) (BL# 25966), FAK (valium 20) (BL# 33617). Flies were raised on standard medium and maintained at 29°C until imaging, unless otherwise stated. Beginning of pupariation was identified according to what is described by Bainbridge and Bownes (Bainbridge and Bownes, 1981).
MARCM clones generation
Crosses were maintained at 25°C between heat-shocks. Heat-shocks were carried out in a 37°C waterbath, followed by 1 hour at 18°C to extend G2 phase and improve MARCM efficiency.
Live imaging
Live pupas were mounted as previously described (Moreira et al., 2011) except that the imaging was done directly through the transparent cuticle. For cell speed analysis, pupas were imaged in an Andor Revolution spinning disc confocal microscope (Andor Technology, UK) using a 40×/1.30 Plan Fluor PFS oil immersion objective (Nikon Instruments) in conjunction with a 488 nm OPSL CW laser and a 500–550 nm band-pass emission filter. For the polarization assay, pupas were imaged using a Zeiss LSM 5 Live line-scanning confocal microscope (Carl Zeiss, Jena) using a 40×/1.30 Plan-Neofluar oil immersion objective in conjunction with a 488 nm Sapphire laser and a LP 505 nm emission filter. Images and time lapses were analysed using Fiji software.
Polarization assay
Pupas were imaged during 20 min (2 min interval) before wounding. Wounding was performed with a pulsed UV laser (355 nm) using a UGA-40 spot illumination scanning system (Rapp OptoElectronic, Germany) fitted to the line-scanning confocal microscope. Wounds were induced on six rectangle-forming spots on the pupa's dorsal epithelium close to the middle dorsal patch. After wounding, pupas were imaged during 1 hour.
To calculate the mean of the polarization angle, we chose three time points before and after wounding, per cell (time points 0, 10 and 18 min before wounding and time points 10, 18 and 28 min after wounding, respectively). The polarization angle corresponds to the angle formed between the wound center, the cell's center and its polarization arm. During migration, hemocytes can sometimes leave behind a trailing edge or present multiple long filopodium-like extensions that can resemble polarity arms. In such cases the cell displacement between flanking time points was used to determine which of the extensions was the leading protrusion. Only hemocytes in which polarity arms were clearly identified were used in the analysis.
Image analysis and quantification
For cell speed quantification, the Manual Tracking and the Chemotaxis and Migration Tool 1.01 (Integrated BioDiagnostics) plugins were used. Maximum projections were used for the analysis. Hemocyte centers were marked manually and their coordinates used to calculate cell speed. A minimum of 15 cells per individual fly were used for these measurements. For polarization angle measurements, wound limits were manually marked and the geometrical center was selected as the wound center. The Angle tool from Fiji was used to measure the polarization angles according to what was previously described. A minimum of three individual flies for each genotype was used in the quantification. For counting the number of hemocytes at the wound area, only cells with protrusions touching the delimited wound area were considered.
Supplementary Material
Acknowledgments
We thank Ângela Dias for the technical help and the Jacinto group members for insightful discussions. The work was supported by a fellowship (SFRH/BD/62345/2009) to C.G.A.M. and a grant (PTDC/SAU-OBD/101259/2008) from Fundação para a Ciência e Tecnologia, and a European Research Council Starting Grant (2007-StG-208631).
Footnotes
Author contributions: C.G.A.M., A.J. and S.P. conceived and designed the experiments. C.G.A.M. performed the experiments and analyzed the data. C.G.A.M., A.J. and S.P. wrote the paper.
Competing interests: The authors have no competing interests to declare.
References
- Alatortsev V. E., Kramerova I. A., Frolov M. V., Lavrov S. A., Westphal E. D. (1997). Vinculin gene is non-essential in Drosophila melanogaster. FEBS Lett. 413, 197–201 10.1016/S0014-5793(97)00901-0 [DOI] [PubMed] [Google Scholar]
- Anthis N. J., Wegener K. L., Ye F., Kim C., Goult B. T., Lowe E. D., Vakonakis I., Bate N., Critchley D. R., Ginsberg M. H. et al. (2009). The structure of an integrin/talin complex reveals the basis of inside-out signal transduction. EMBO J. 28, 3623–3632 10.1038/emboj.2009.287 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Babcock D. T., Brock A. R., Fish G. S., Wang Y., Perrin L., Krasnow M. A., Galko M. J. (2008). Circulating blood cells function as a surveillance system for damaged tissue in Drosophila larvae. Proc. Natl. Acad. Sci. USA 105, 10017–10022 10.1073/pnas.0709951105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bainbridge S. P., Bownes M. (1981). Staging the metamorphosis of Drosophila melanogaster. J. Embryol. Exp. Morphol. 66, 57–80. [PubMed] [Google Scholar]
- Baker B. M., Chen C. S. (2012). Deconstructing the third dimension: how 3D culture microenvironments alter cellular cues. J. Cell Sci. 125, 3015–3024 10.1242/jcs.079509 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beerling E., Ritsma L., Vrisekoop N., Derksen P. W. B., van Rheenen J. (2011). Intravital microscopy: new insights into metastasis of tumors. J. Cell Sci. 124, 299–310 10.1242/jcs.072728 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bornens M. (2008). Organelle positioning and cell polarity. Nat. Rev. Mol. Cell Biol. 9, 874–886 10.1038/nrm2524 [DOI] [PubMed] [Google Scholar]
- Brown N. H., Gregory S. L., Martin-Bermudo M. D. (2000). Integrins as mediators of morphogenesis in Drosophila. Dev. Biol. 223, 1–16 10.1006/dbio.2000.9711 [DOI] [PubMed] [Google Scholar]
- Choi C. K., Vicente-Manzanares M., Zareno J., Whitmore L. A., Mogilner A., Horwitz A. R. (2008). Actin and alpha-actinin orchestrate the assembly and maturation of nascent adhesions in a myosin II motor-independent manner. Nat. Cell Biol. 10, 1039–1050 10.1038/ncb1763 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coll J. L., Ben-Ze'ev A., Ezzell R. M., Rodríguez Fernández J. L., Baribault H., Oshima R. G., Adamson E. D. (1995). Targeted disruption of vinculin genes in F9 and embryonic stem cells changes cell morphology, adhesion, and locomotion. Proc. Natl. Acad. Sci. USA 92, 9161–9165 10.1073/pnas.92.20.9161 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deramaudt T. B., Dujardin D., Hamadi A., Noulet F., Kolli K., De Mey J., Takeda K., Rondé P. (2011). FAK phosphorylation at Tyr-925 regulates cross-talk between focal adhesion turnover and cell protrusion. Mol. Biol. Cell 22, 964–975 10.1091/mbc.E10-08-0725 [DOI] [PMC free article] [PubMed] [Google Scholar]
- DiMilla P. A., Barbee K., Lauffenburger D. A. (1991). Mathematical model for the effects of adhesion and mechanics on cell migration speed. Biophys. J. 60, 15–37 10.1016/S0006-3495(91)82027-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duffy J. B. (2002). GAL4 system in Drosophila: a fly geneticist's Swiss army knife. Genesis 34, 1–15 10.1002/gene.10150 [DOI] [PubMed] [Google Scholar]
- Fraley S. I., Feng Y., Krishnamurthy R., Kim D.-H., Celedon A., Longmore G. D., Wirtz D. (2010). A distinctive role for focal adhesion proteins in three-dimensional cell motility. Nat. Cell Biol. 12, 598–604 10.1038/ncb2062 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goto A., Kumagai T., Kumagai C., Hirose J., Narita H., Mori H., Kadowaki T., Beck K., Kitagawa Y. (2001). A Drosophila haemocyte-specific protein, hemolectin, similar to human von Willebrand factor. Biochem. J. 359, 99–108 10.1042/0264-6021:3590099 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grabbe C., Zervas C. G., Hunter T., Brown N. H., Palmer R. H. (2004). Focal adhesion kinase is not required for integrin function or viability in Drosophila. Development 131, 5795–5805 10.1242/dev.01462 [DOI] [PubMed] [Google Scholar]
- Grashoff C., Hoffman B. D., Brenner M. D., Zhou R., Parsons M., Yang M. T., McLean M. A., Sligar S. G., Chen C. S., Ha T. et al. (2010). Measuring mechanical tension across vinculin reveals regulation of focal adhesion dynamics. Nature 466, 263–266 10.1038/nature09198 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harburger D. S., Bouaouina M., Calderwood D. A. (2009). Kindlin-1 and -2 directly bind the C-terminal region of beta integrin cytoplasmic tails and exert integrin-specific activation effects. J. Biol. Chem. 284, 11485–11497 10.1074/jbc.M809233200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoffman L. M., Jensen C. C., Kloeker S., Wang C.-L. A., Yoshigi M., Beckerle M. C. (2006). Genetic ablation of zyxin causes Mena/VASP mislocalization, increased motility, and deficits in actin remodeling. J. Cell Biol. 172, 771–782 10.1083/jcb.200512115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huelsmann S., Hepper C., Marchese D., Knöll C., Reuter R. (2006). The PDZ-GEF dizzy regulates cell shape of migrating macrophages via Rap1 and integrins in the Drosophila embryo. Development 133, 2915–2924 10.1242/dev.02449 [DOI] [PubMed] [Google Scholar]
- Humphries J. D., Wang P., Streuli C., Geiger B., Humphries M. J., Ballestrem C. (2007). Vinculin controls focal adhesion formation by direct interactions with talin and actin. J. Cell Biol. 179, 1043–1057 10.1083/jcb.200703036 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huttenlocher A., Horwitz A. R. (2011). Integrins in cell migration. Cold Spring Harb. Perspect. Biol. 3, a005074 10.1101/cshperspect.a005074 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ilić D., Kovacic B., Johkura K., Schlaepfer D. D., Tomasević N., Han Q., Kim J.-B., Howerton K., Baumbusch C., Ogiwara N. et al. (2004). FAK promotes organization of fibronectin matrix and fibrillar adhesions. J. Cell Sci. 117, 177–187 10.1242/jcs.00845 [DOI] [PubMed] [Google Scholar]
- Lämmermann T., Bader B. L., Monkley S. J., Worbs T., Wedlich-Söldner R., Hirsch K., Keller M., Förster R., Critchley D. R., Fässler R. et al. (2008). Rapid leukocyte migration by integrin-independent flowing and squeezing. Nature 453, 51–55 10.1038/nature06887 [DOI] [PubMed] [Google Scholar]
- Lanot R., Zachary D., Holder F., Meister M. (2001). Postembryonic hematopoiesis in Drosophila. Dev. Biol. 230, 243–257 10.1006/dbio.2000.0123 [DOI] [PubMed] [Google Scholar]
- Larjava H., Plow E. F., Wu C. (2008). Kindlins: essential regulators of integrin signalling and cell-matrix adhesion. EMBO Rep. 9, 1203–1208 10.1038/embor.2008.202 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li R., Gundersen G. G. (2008). Beyond polymer polarity: how the cytoskeleton builds a polarized cell. Nat. Rev. Mol. Cell Biol. 9, 860–873 10.1038/nrm2522 [DOI] [PubMed] [Google Scholar]
- Makhijani K., Alexander B., Tanaka T., Rulifson E., Brückner K. (2011). The peripheral nervous system supports blood cell homing and survival in the Drosophila larva. Development 138, 5379–5391 10.1242/dev.067322 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Márkus R., Laurinyecz B., Kurucz E., Honti V., Bajusz I., Sipos B., Somogyi K., Kronhamn J., Hultmark D., Andó I. (2009). Sessile hemocytes as a hematopoietic compartment in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 106, 4805–4809 10.1073/pnas.0801766106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mellman I., Nelson W. J. (2008). Coordinated protein sorting, targeting and distribution in polarized cells. Nat. Rev. Mol. Cell Biol. 9, 833–845 10.1038/nrm2525 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mierke C. T., Kollmannsberger P., Zitterbart D. P., Diez G., Koch T. M., Marg S., Ziegler W. H., Goldmann W. H., Fabry B. (2010). Vinculin facilitates cell invasion into three-dimensional collagen matrices. J. Biol. Chem. 285, 13121–13130 10.1074/jbc.M109.087171 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Montell D. J., Yoon W. H., Starz-Gaiano M. (2012). Group choreography: mechanisms orchestrating the collective movement of border cells. Nat. Rev. Mol. Cell Biol. 13, 631–645 10.1038/nrm3433 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moreira C. G. A., Regan J. C., Zaidman-Rémy A., Jacinto A., Prag S. (2011). Drosophila hemocyte migration: an in vivo assay for directional cell migration. Methods Mol. Biol. 769, 249–260 10.1007/978-1-61779-207-6_17 [DOI] [PubMed] [Google Scholar]
- Niethammer P., Grabher C., Look A. T., Mitchison T. J. (2009). A tissue-scale gradient of hydrogen peroxide mediates rapid wound detection in zebrafish. Nature 459, 996–999 10.1038/nature08119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palecek S. P., Loftus J. C., Ginsberg M. H., Lauffenburger D. A., Horwitz A. F. (1997). Integrin-ligand binding properties govern cell migration speed through cell-substratum adhesiveness. Nature 385, 537–540 10.1038/385537a0 [DOI] [PubMed] [Google Scholar]
- Parsons J. T., Horwitz A. R., Schwartz M. A. (2010). Cell adhesion: integrating cytoskeletal dynamics and cellular tension. Nat. Rev. Mol. Cell Biol. 11, 633–643 10.1038/nrm2957 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siekhaus D., Haesemeyer M., Moffitt O., Lehmann R. (2010). RhoL controls invasion and Rap1 localization during immune cell transmigration in Drosophila. Nat. Cell Biol. 12, 605–610 10.1038/ncb2063 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sinenko S. A., Mathey-Prevot B. (2004). Increased expression of Drosophila tetraspanin, Tsp68C, suppresses the abnormal proliferation of ytr-deficient and Ras/Raf-activated hemocytes. Oncogene 23, 9120–9128 10.1038/sj.onc.1208156 [DOI] [PubMed] [Google Scholar]
- Smith M. A., Blankman E., Gardel M. L., Luettjohann L., Waterman C. M., Beckerle M. C. (2010). A zyxin-mediated mechanism for actin stress fiber maintenance and repair. Dev. Cell 19, 365–376 10.1016/j.devcel.2010.08.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stofanko M., Kwon S. Y., Badenhorst P. (2008). A misexpression screen to identify regulators of Drosophila larval hemocyte development. Genetics 180, 253–267 10.1534/genetics.108.089094 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stramer B., Wood W., Galko M. J., Redd M. J., Jacinto A., Parkhurst S. M., Martin P. (2005). Live imaging of wound inflammation in Drosophila embryos reveals key roles for small GTPases during in vivo cell migration. J. Cell Biol. 168, 567–573 10.1083/jcb.200405120 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stramer B., Moreira S., Millard T., Evans I., Huang C.-Y., Sabet O., Milner M., Dunn G., Martin P., Wood W. (2010). Clasp-mediated microtubule bundling regulates persistent motility and contact repulsion in Drosophila macrophages in vivo. J. Cell Biol. 189, 681–689 10.1083/jcb.200912134 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tirouvanziam R., Davidson C. J., Lipsick J. S., Herzenberg L. A. (2004). Fluorescence-activated cell sorting (FACS) of Drosophila hemocytes reveals important functional similarities to mammalian leukocytes. Proc. Natl. Acad. Sci. USA 101, 2912–2917 10.1073/pnas.0308734101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Toetsch S., Olwell P., Prina-Mello A., Volkov Y. (2009). The evolution of chemotaxis assays from static models to physiologically relevant platforms. Integr. Biol. 1, 170–181 10.1039/b814567a [DOI] [PubMed] [Google Scholar]
- Tolde O., Rösel D., Janoštiak R., Veselý P., Brábek J. (2012). Dynamics and morphology of focal adhesions in complex 3D environment. Folia Biologica 58, 177–184. [DOI] [PubMed] [Google Scholar]
- Weijer C. J. (2009). Collective cell migration in development. J. Cell Sci. 122, 3215–3223 10.1242/jcs.036517 [DOI] [PubMed] [Google Scholar]
- Wood W., Jacinto A. (2007). Drosophila melanogaster embryonic haemocytes: masters of multitasking. Nat. Rev. Mol. Cell Biol. 8, 542–551 10.1038/nrm2202 [DOI] [PubMed] [Google Scholar]
- Wu J. S., Luo L. (2007). A protocol for mosaic analysis with a repressible cell marker (MARCM) in Drosophila. Nat. Protoc. 1, 2583–2589 10.1038/nprot.2006.320 [DOI] [PubMed] [Google Scholar]
- Ye F., Kim C., Ginsberg M. H. (2011). Molecular mechanism of inside-out integrin regulation. J. Thromb. Haemost. 9 Suppl. 1, 20–25 10.1111/j.1538-7836.2011.04355.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshigi M., Hoffman L. M., Jensen C. C., Yost H. J., Beckerle M. C. (2005). Mechanical force mobilizes zyxin from focal adhesions to actin filaments and regulates cytoskeletal reinforcement. J. Cell Biol. 171, 209–215 10.1083/jcb.200505018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaidel-Bar R., Ballestrem C., Kam Z., Geiger B. (2003). Early molecular events in the assembly of matrix adhesions at the leading edge of migrating cells. J. Cell Sci. 116, 4605–4613 10.1242/jcs.00792 [DOI] [PubMed] [Google Scholar]
- Zaidel-Bar R., Itzkovitz S., Ma'ayan A., Iyengar R., Geiger B. (2007). Functional atlas of the integrin adhesome. Nat. Cell Biol. 9, 858–867 10.1038/ncb0807-858 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zettervall C.-J., Anderl I., Williams M. J., Palmer R., Kurucz E., Ando I., Hultmark D. (2004). A directed screen for genes involved in Drosophila blood cell activation. Proc. Natl. Acad. Sci. USA 101, 14192–14197 10.1073/pnas.0403789101 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.