

Abstract Abstract
The southeast Asian scarab beetle genus Peltonotus Burmeister (Scarabaeidae, Dynastinae, Cyclocephalini) is reviewed. New country records for Peltonotus morio Burmeister (Myanmar and Vietnam), Peltonotus nasutus Arrow (southern China and Cambodia), and Peltonotus favonius Jameson and Wada (Myanmar) are reported, including a new record in the Palearctic/Sino-Japanese biogeographic region. The first female specimen of Peltonotus favonius is described. Biological associations with aroid inflorescences are reviewed, and human consumption of Peltonotus beetles is reported. A key to all species, paralectotype designations for Peltonotus nasutus, diagnoses, and distributions using dynamic mapping tools are included.
Keywords: Edible insects, Palearctic region, Sino-Japanese region, Araceae, dynamic mapping
Introduction
The scarab beetle genus Peltonotus Burmeister (Scarabaeidae, Dynastinae) includes 25 species that are distributed in forest habitats in Southeast Asia and that are associated with aroid inflorescences (Araceae) (Jameson and Wada 2004). Adult beetles use inflorescences as sites for mating and feeding, and they serve as pollinators (Moore and Jameson in press, Maia et al. 2012). Species in the genus are intimately tied to host aroids and their forest habitats, and we predict that many species await discovery. Members of the genus form a natural group based on a unique, articulated maxillary tooth. The first monograph for the genus (Jameson and Wada 2004) included 19 species; since this time, six additional species have been described (Jameson and Wada 2009, Jameson and Jakl 2010), a 30% increase in species diversity.
Identification of species in the genus Peltonotus is hampered by sexual dimorphism that makes association of conspecific sexes difficult, absence of male or female specimens for some species, rarity of some species (perhaps due to brief activity patterns and host plant phenology), and color variability within species. For this reason, we amalgamate existing keys into one identification guide for males and females and provide diagnoses.
Species of Peltonotus are associated with aroid inflorescences (Araceae) (Jameson and Wada 2004). However, in comparison to the abundant research on aroid and scarab beetle interactions in the New World tropics (e.g., Gibernau et al. 2010; Maia et al. 2012; Young 1988), little research is being conducted on Peltonotus and aroids in the Old World. Peltonotus malayensis Arrow is associated with inflorescences of the climbing aroid, Epipremnum falcifolium Engl. (Araceae) (Jameson and Wada 2004). Male and female beetles (as well as many small beetles and arthropods) have been reported around the base of the spathe where adult Peltonotus malayensis were observed mating and feeding. Inflorescences of the cultivated aroid, Amorphophallus paeoniifolius (Dennst.) Nicolson (Araceae), attract aggregations of Peltonotus nasutus Arrow (Grimm 2009). This plant (also called the elephant foot yam or corpse plant) grows on the forest floor in dappled shade or in the open sun in secondary forest or highly disturbed areas. The large flower (up to 40 cm) smells like a rotting dead animal and deceptively attracts insects that may serve as pollinators (Schiestl and Dötterl 2012) including the carrion scarabs Phaeochrous dissimilis Arrow, Phaeochrous emarginatus Laporte, and Phaeochrous intermedius Pic (all Scarabaeoidea, Hybosoridae), and the aroid scarab Peltonotus nasutus (Grimm 2009). Additional research on aroids and Peltonotus species is needed in order to clarify plant-insect interactions including evolution, ecology, and pollination.
In addition to being associated with aroid inflorescences, adults are attracted to lights at night, and some have been collected in malaise traps. Adults may have short seasonal activity patterns. Some adults have been recorded for only two nights during season-long, intensive collecting efforts. Larvae are not known for any species in the genus.
Survey efforts and collecting in Southeast Asia have provided new distributional data for species in the genus, thus yielding a clearer understanding of distribution patterns. Herein, we report new distributional data for three species of Peltonotus. Because identification of species requires use of three publications (Jameson and Wada 2004, Jameson and Wada 2009, Jameson and Jakl 2010), we provide a comprehensive key to all species in the genus, short diagnoses, new paralectotype designations for Peltonotus nasutus, and maps with associated files for dynamic mapping capabilities.
Material and methods
Characters and specimens were examined using a dissecting microscope (6–48× magnification) and fiber-optic illumination. Digital images of specimens and structures were captured using the Leica Application Suite V3.8. Images were edited in Adobe Photoshop CS2 (background removed, contrast manipulated). In the absence of images for some specimens, illustrations are used. Specimen localities that were not recorded in latitude and longitude on original labels were translated using GoogleEarth (www.google.com/earth/index.html) or by using the Global Gazetteer Version 2.2 (www.fallingrain.com/world/). It should be noted that older localities have a wide margin of error, and their lack of precision is not conducive to ecological or niche modeling. Maps were generated by entering these data into Microsoft Excel 2008 and uploaded to EarthPoint (www.earthpoint.us/Excel-ToKml.aspx) and GoogleEarth (Appendix 1). These mapping tools allow for interactive mapping and addition of data by subsequent users. Locality information in species treatments is recorded with the country in bold letters, followed by the state/province/district, and the specific locality in parentheses.
This work unifies some character state definitions (e.g., form of labrum, male protibial teeth, female elytral epipleural pillow) previously used for identification of Peltonotus species. Species are characterized by combinations of characters including the form of the labrum (weakly sinuate, bi-emarginate/broadly emarginated, or deeply bilobed) (Figs 20–24), mentum apex and second labial palpomere (compared with palpomere 1) (Figs 25–35), mala of maxilla with or without thickened and strongly flattened setae (“lamellate setal brush”) (Figs 36–44), stipes of maxilla with or without curly setae (Figs 36–44), male protibia tri- or bidentate (Figs 45–49), form of male protarsomeres (Figs 50–54), form of the male parameres (Figs 55–72), and form of the female epipleuron in ventral view in relation to the position of the metacoxa (Figs 73–91). Expansions of the female elytral epipleuron may have an inflated area (or pillow) in dorsal view (Moore 2012). Setae are important for species diagnosis and are defined as minute if they are less than 0.2 mm, short if between 0.2–0.5 mm, moderately long if between 0.5–1.0 mm, and long if between 1.0–2.0 mm (as measured with an ocular micrometer). Punctures may lack setae, possess one seta (unisetigerous), or possess multiple setae (multisetigerous). Male parameres are highly asymmetrical, and we elected to illustrate the lateral view that best assists in identification.
Figures 20–24.

Head (dorsal view) showing characters of the labrum, mandible, and clypeus. 20 Peltonotus karubei (apex of labrum deeply bi-lobed; apex of mandible rounded laterally) 21 Peltonotus malayensis (apex of labrum bi-emarginate; apex of mandible rounded laterally) 22 Peltonotus vittatus (apex of labrum bi-emarginate; apex of mandible rounded laterally) 23 Peltonotus nasutus (apex of labrum weakly sinuate; apex of mandible quadrate laterally with broadly truncate apex; apex of clypeus with weak tubercle in male) 24 Peltonotus morio (apex of labrum weakly sinuate; apex of mandible quadrate laterally with broadly truncate apex; apex of clypeus without tubercle in male).
Figures 25–35.

Mentum, ventral view, showing form of apical half of mentum and form of labial palpomere 2 (in comparison to palpomere 1). 25 Peltonotus deltamentum 26 Peltonotus gracilipodus 27 Peltonotus kyojinus 28 Peltonotus malayensis 29 Peltonotus podocrassus 30 Peltonotus nasutus 31 Peltonotus nethis 32 Peltonotus pruinosus 33 Peltonotus similis 34 Peltonotus sisyrus 35 Peltonotus talangensis.
Figures 36–44.

Maxilla, ventral view, showing mala with or without lamellate setal brush (setae thick and strongly flattened), and showing stipes with or without setae curly at apices. 36 Peltonotus animus 37 Peltonotus cybele 38 Peltonotus deltamentum 39 Peltonotus favonius 40 Peltonotus karubei 41 Peltonotus malayensis 42 Peltonotus mushiyaus 43 Peltonotus talangensis 44 Peltonotus tigerus. Arrows indicate lamellate setal brush.
Figures 45–54.

Male prolegs, dorsal view (45–49), male protarsomeres, dorsal view (50–53), and male protarsomere 5, ventral view (54), of Peltonotus. 45 Peltonotus animus (male protibia tridentate with basal tooth obsolete) 46 Peltonotus cybele (male protibia tridentate with basal tooth well developed) 47 Peltonotus malayensis (male protibia bidentate) 48 Peltonotus talangensis (male protibia tridentate with basal tooth well developed) 49 Peltonotus rubripennis (male protibia tridentate with basal tooth weakly developed) 50 Peltonotus adelphosimilis (arrow showing protarsomere 5 of male with internoapical protuberance) 51 Peltonotus favonius (male protibia bidentate) 52 Peltonotus nasutus (male protibial claw greatly enlarged) 53 Peltonotus similis (arrow showing protarsomere 5 of male with internomedial protuberance) 54 Peltonotus deltamentum (male proclaw strongly arcuate in ventral view).
Figures 55–60.

Male parameres (with or without phallobase), dorsal and lateral views, in Peltonotus. Male parameres are highly asymmetrical, and we illustrate the lateral view that best assists in identification. 55 Peltonotus adelphosimilis 56 Peltonotus animus 57 Peltonotus brunnipennis 58 Peltonotus cybele 59 Peltonotus deltamentum 60 Peltonotus favonius.
Figures 67–72.

Male parameres (with or without phallobase), dorsal and lateral views, in Peltonotus. Male parameres are highly asymmetrical, and we illustrate the lateral view that best assists in identification. 67 Peltonotus rubripennis 68 Peltonotus silvanus 69 Peltonotus similis 70 Peltonotus sisyrus 71 Peltonotus talangensis 72 Peltonotus vittatus.
Figures 73–87.

Female elytral epipleuron (gray, ventral view) and position relative to metacoxa in Peltonotus. 73 Peltonotus brunnipennis 74 Peltonotus cybele 75 Peltonotus favonius 76 Peltonotus fujiokai 77 Peltonotus gracilipodus, Peltonotus podocrassus and Peltonotus silvanus 78 Peltonotus kyojinus 79 Peltonotus malayensis 80 Peltonotus morio 81 Peltonotus mushiyaus 82 Peltonotus nasutus 83 Peltonotus nethis 84 Peltonotus pruinosus 85 Peltonotus rubripennis 86 Peltonotus similis 87 Peltonotus sisyrus
Figures 88–91.
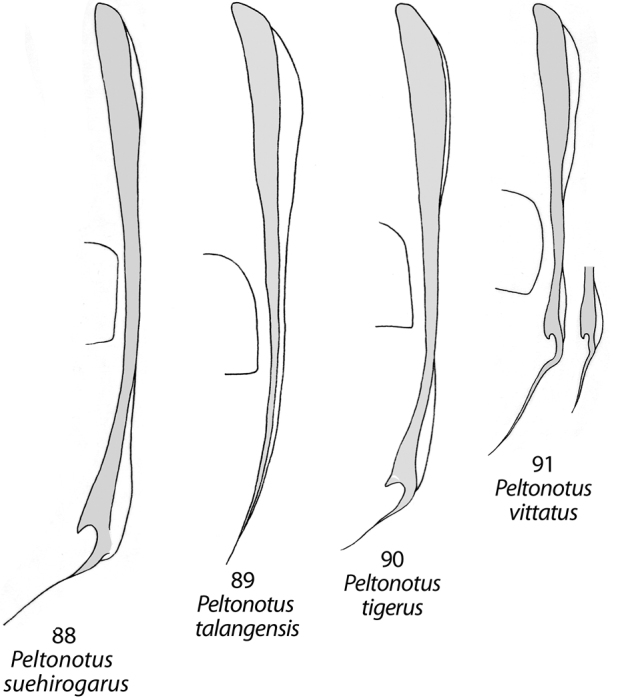
Female elytral epipleuron (gray, ventral view) and position relative to metacoxa in Peltonotus. 88 Peltonotus suehirogarus 89 Peltonotus talangensis 90 Peltonotus tigerus 91 Peltonotus vittatus.
We follow the phylogenetic species concept (Wheeler and Platnick 2000) that states that “A species is the smallest aggregation of (sexual) populations or (asexual) lineages diagnosable by a unique combination of character states.” Specimens examined for this research are deposited in the following institutions and private collections: the IRSNB, the Alain Drumont Collection, Brussels, Belgium; the FUJI; the MNHN; AREC; the MLJC; the Shinji Nagia Collection (Nagano, Japan); and BMNH.
New distributional records, human consumption, and paralectotype designations for Peltonotus nasutus
Peltonotus nasutus (Figs 14–15) is the most distinctive species within the genus Peltonotus due to its large body size (~20 mm), tubercle at the apex of the clypeus in the male (Fig. 23), and greatly enlarged protibial claw in the male (Fig. 52).
Figures 13–19.

Peltonotus species dorsal habitus. 13 Peltonotus mushiyaus, female 14–15 Peltonotus nasutus, male and female (respectively) 16 Peltonotus talangensis, male 17 Peltonotus tigerus, female 18–19 Peltonotus vittatus, male and female (respectively).
Large aggregations of adults (over 100) have been found in association with the large, fetid-smelling aroids in the genus Amorphophallus (Grimm 2009; label data at BMNH). In Thailand, the stench of flowering Amorphophallus paeoniifolius attracts a profusion of Peltonotus nasutus individuals that serve to pollinate the inflorescence. Seventy eight specimens were recorded in one flower, and these were collected, fried with fish sauce and salt, and then consumed by the Karen-speaking tribe in the Tak province in northern Thailand (Danell 2010). Thai people consume more insects per capita than other people and cultures (Chen et al. 1998), and this beetle species is a new record for human consumption.
The species is distributed in Myanmar, Thailand, Laos, and Vietnam (Jameson and Wada 2004; Li et al. 2012) (Fig. 92). Adults inhabit deciduous dipterocarp forests between 100–800 m elevation and have been collected at mercury vapor light traps. Examination of additional specimens provided new country records for Peltonotus nasutus in Cambodia and China. This species was not previously recorded as occurring in the Palaearctic region (as defined by Löbl and Smetana 2003). These records demonstrate that the species occurs in the Guangxi and Guizhou provinces of southern portion of China in what is considered the Palaearctic biogeographic region (Löbl and Smetana 2003) or the Sino-Japanese biogeographic region (Holt et al. 2013). New Country Record: CHINA (6 males, 2 females deposited in Drumont Collection; AREC): Guangxi Zhuang Autonomous Region (Guangxi), Guizhou (Weining, Mt. Ping-Qing-Liang-Zi), Yunnan (Jinggu, Mt. Longtanshan; Menglian, Mt. Daheishan). Specimens were collected from May to July: May (1), June (3), July (4). New Country Record: CAMBODIA (9 males, 12 females deposited at IRSNB): Pursat (Phnom Samkos Wildlife Sanctuary), Ratanakiri (Phumi Kalai Thum), Pailin (Pailin). Specimens were collected from April to June and November: April (3), May (2), June (3), November (13). The new country record in Weining, China extends the known range of the species over 600 km north.
Figure 92.

Distribution of Peltonotus morio (green icon) and Peltonotus nasutus (blue icon) in southeast Asia. Icons with stars indicate new country records for each species. Map was generated using data in Table 1.
Table 1.
Peltonotus Locality Table. Locality information for Peltonotus morio and Peltonotus nasutus. The Appendix file can be used for dynamic mapping using EarthPoint and GoogleEarth.
| Latitude, Longitude | Species name | Collection or Reference | Locality Information |
|---|---|---|---|
| 16°40'27"N, 98°17'59"E | Peltonotus morio | FUJI | S. Burma, Mt. Dawna, V.1992, 1 male, ele. 763m, NEW COUNTRY RECORD |
| 12°05'N, 99°00'E | Peltonotus morio | FUJI | S. Burma, Tenasserim, V.1992, 1 female, NEW COUNTRY RECORD |
| 26°52'41"N, 88°17'25"E | Peltonotus morio | BMNH | India, Kurseong Div., Lat Panchar, 4000 ft., VI.1934, 6 specimens, Col. Champion |
| 27°39'N, 84°19'E | Peltonotus morio | Dhoj et al. 2009 | Nepal, Chitwan Central region, Gunjanagar, 230 m |
| 27°39'N, 84°21'E | Peltonotus morio | Dhoj et al. 2009 | Nepal, Rampur, 230 m, amid maize-maize-vegetables in sandy soil from farming sites. |
| 22°29'N, 103°57'E | Peltonotus morio | IRSBN | Vietnam, Lao Cai Prov., VI.10.1917, 1 male NEW COUNTRY RECORD |
| 18°49'16"N, 98°55'11"E | Peltonotus morio | Jameson and Wada 2004 | Thailand, Doi Suthep |
| 27°18'42"N, 88°35'57"E | Peltonotus morio | Jameson and Wada 2004 | Sikkim, India |
| 24°39'32"N, 93°54'22"E | Peltonotus morio | Jameson and Wada 2004 | India, Manipur |
| 25°22'05"N, 91°45'13"E | Peltonotus morio | Jameson and Wada 2004 | India, Meghalaya, Khasi Hills |
| 27°09'33"N, 88°36'56"E | Peltonotus morio | Jameson and Wada 2004 | India, Pedong |
| 27°02'09"N, 88°14'08"E | Peltonotus morio | Jameson and Wada 2004 | India, Darjeeling |
| 28°16'N, 84°05'E | Peltonotus morio | Jameson and Wada 2004 | Nepal, Chhachok |
| 14°48'00"N, 106°49'59"E | Peltonotus nasutus | FUJI | S. Laos, Attapu, V.13.2007, 1 male, 1 female, ele 450m |
| 14°88'N, 105°87'E | Peltonotus nasutus | FUJI | S. Laos, Champasak Province, 2 females, |
| 16°42'18"N, 98°20'44"E | Peltonotus nasutus | FUJI | S. Burma, Mt. Dawna, V.1992, 1 female |
| 18°38'31"N, 94°42'56"E | Peltonotus nasutus | FUJI | Myanmar, Arakan Province, Nianjyo, 1070m, 1 male, 1 female |
| 15°N, 98°32'E | Peltonotus nasutus | BMNH | W. Thailand, Kanchanaburi Prov., Thung Yai Wildlife Sanctuary, mixed riverside forest, M. Brendell, V.8.1988, 10 specimens, within spathe of Amorphophallus inflorescence |
| 19°25'N, 103°30'E | Peltonotus nasutus | BMNH | Laos, Xiankhouang Prov. V.18.1919, 1 male |
| 26°51'22"N, 104°13'59"E | Peltonotus nasutus | Drumont | Chine, Guizhou, Mt. Ping-Qing-Liang-Zi, Weining county, 1-10/VII-2009, 1 male, 3 female NEW COUNTRY RECORD |
| 23°28'5"N, 100°41'E | Peltonotus nasutus | Drumont | Chine, Yunnan, Mt. Longtanshan, Jinggu county, VI.11-20, 3 male, Col. Li Jingke NEW COUNTRY RECORD |
| 22°35'N, 99°33'E | Peltonotus nasutus | Drumont | Chine, Yunnan, Mt. Daheishan, Menglian county, V.20-31-2009, Col. Li Jingke, 1 female NEW COUNTRY RECORD |
| 22°47'56"N, 108°19'44"E | Peltonotus nasutus | AREC | China, Guangxi Zhuang Automonus Region NEW COUNTRY RECORD |
| 17°28'59"N, 101°4'0"E | Peltonotus nasutus | IRSBN | Thailand, Changwat Loei, Na Haeo Bio. Sta., V-15-19-2003, light trap, Col. Constant, Smets, and Grootaert, 1 male, 2 female |
| 17°28'59"N, 101°4'0"E | Peltonotus nasutus | IRSBN | Thailand, Changwat Loei, Na Haeo Bio. Sta., V.17.2003, edge pond, Col. Constant and Smets, 2 female |
| 17°28'59"N, 101°4'0"E | Peltonotus nasutus | IRSBN | Thailand, Changwat Loei, Na Haeo Bio. Sta., V.5-12-2001, light trap, Col. Constant and Grootaert, 2 female |
| 19°27'N, 98°20'E | Peltonotus nasutus | IRSBN | N. Thailand, Mae Hong Son Prov., 600 m, 28-V to 2-VI-1999, Col. D. Hauck, 2 male, 2 female |
| 14°16'07"N, 98°59'12"E | Peltonotus nasutus | IRSBN | Thailand, Kanchanaburi Prov., Sai Yok NP, VI.4–5.2003, Constant and Smets, 1 male, 1 female |
| 13°49'59"N, 106°57'0"E | Peltonotus nasutus | IRSBN | Cambodia, Ratanakiri Prov., Phumi Kalai Thum., VI.1-19.2007, Col. Li Jingke, 1 male, 2 female NEW COUNTRY RECORD |
| 12°18'09"N, 102°59'20"E | Peltonotus nasutus | IRSBN | Cambodia, Pursat Prov., Phnum Samkos Wildlife Sanctuary, XI.15, 2005, light trapping, col. Smets and Van, 5 male, 4 female NEW COUNTRY RECORD |
| 12°18'09"N, 102°59'20"E | Peltonotus nasutus | IRSBN | Cambodia, Pursat Prov., Phnum Samkos Wildlife Sanctuary, IV.13-14, 2005, light trapping, primary forest edge, col. Smets and Van, 1 female, 1 male, |
| 12°18'09"N, 102°59'20"E | Peltonotus nasutus | IRSBN | Cambodia, Pursat Prov., Phnum Samkos Wildlife Sanctuary, IV.16, 2005, light trapping, col. Smets and Van, 3 female, 1 male NEW COUNTRY RECORD |
| 12°18'09"N, 102°59'20"E | Peltonotus nasutus | IRSBN | Cambodia, Pursat Prov., Phnum Samkos Wildlife Sanctuary, IV.15, 2005, light trapping, col. Smets and Van, 1 female NEW COUNTRY RECORD |
| 12°51'2"N, 102°36'34"E | Peltonotus nasutus | Drumont | Cambodia, Pailin Prov., 270m, V.6-16.2008, col. Murzin, 2 female NEW COUNTRY RECORD |
| 21°17'13"N, 101°10'02"E | Peltonotus nasutus | IRSBN | NW Laos, Louang Namtha Prov., Muang Sing, Houaylong-Kao, VI.2-19.2010, 6 male, 16 female |
| 17°58'0"N, 102°35'59"E | Peltonotus nasutus | IRSBN | Laos, Vientiane Prov., IV.4-1915, 1 female |
| 17°58'0"N, 102°35'59"E | Peltonotus nasutus | IRSBN | Laos, Vientiane Prov., V.18-1915, 1 male |
| 20°09'0"N, 101°19'53"E | Peltonotus nasutus | Li et al. 2012 | Laos, Bokeo Prov., Pha Ngam |
| 16°46'30"N, 102°37'10"E | Peltonotus nasutus | Jameson and Wada 2004 | Thailand, Khorat |
| 14°35'21"N, 98°44'29"E | Peltonotus nasutus | Jameson and Wada 2004 | Thailand, Pu Nam Long Hot Spring |
| 14°47'53"N, 98°44'29"E | Peltonotus nasutus | Jameson and Wada 2004 | Thailand, Khao Leam Dam |
| 19°05'47"N, 100°57'09"E | Peltonotus nasutus | Jameson and Wada 2004 | Thailand, Nan Province |
| 19°21'46"N, 98°59'01"E | Peltonotus nasutus | Jameson and Wada 2004 | Thailand, Ban Chiang Dao |
| 14°43'02"N, 102°01'23"E | Peltonotus nasutus | Jameson and Wada 2004 | Thailand, Khorat Prov., Pak Thong Chai |
| 17°57'46"N, 102°36'54"E | Peltonotus nasutus | Jameson and Wada 2004 | Laos, Vientane |
| 19°36'41"N, 103°43'44"E | Peltonotus nasutus | Jameson and Wada 2004 | Laos, Xiangkhouang |
| 22°20'59"N, 96°55'00"E | Peltonotus nasutus | Jameson and Wada 2004 | Myanmar, Gokhteik |
| 21°14'14"N, 106°22'34"E | Peltonotus nasutus | Jameson and Wada 2004 | Vietnam, Tonkin (north Vietnam) |
| 10°44'57"N, 106°40'43"E | Peltonotus nasutus | Jameson and Wada 2004 | Vietnam, Cochinchina (southern Vietnam) |
During the course of our research, we discovered two unrecorded paralectotype specimens. The male lectotype (at BMNH) and eight paralectotypes (6 at BMNH, 2 at MNHN) were previously designated (Jameson and Wada 2004). Two additional paralectotypes (1 male, 1 female) were found at IRSNB. The paralectotype male at IRSNB is labeled: a) “Cochinchina” (handwritten), b) “Collection E. Candèze” (type set with scribed, black box), c) “Type” (type set, red ink, with scribed, black box), d) “Peltonotus nasutus, Arrow co-type” (handwritten), e) “Peltonotus nasutus Type Arrow det Arrow 1908” (handwritten and type set), f) our paralectotype label. The paralectotype female at IRSNB is labeled: a) “Cochinch” (handwritten), b) “Collection E. Candèze” (type set with scribed, black box), c) “Type” (type set, red ink, with scribed, black box), d) “Peltonotus nasutus Type Arrow det Arrow 1908” (handwritten and type set), e) our paralectotype label.
New distributional records and description of first female specimen for Peltonotus favonius
Peltonotus favonius Jameson and Wada (Fig. 4) was previously known based only on one male specimen from Vietnam (Jameson and Wada 2009). This species is most similar to Peltonotus pruinosus, a species for which only the female holotype is known. The discovery of additional male specimens and the first female specimens facilitates identification of the species, expands the characteristics of the species, and broadens our understanding of the distribution of the species. New Country Record(2 male and 2 female specimens deposited in MLJC): MYANMAR, Mt. Nweezin, ~750m, 10 km NNE of Puta-o, North Kachin, June 16–21, 1998. The new record extends the known range of the species over 2000 km from Vietnam to Myanmar. Specimens were provided by Shinji Nagai. Male specimens from Myanmar (n=2) possess black and reddish-brown elytra (the holotype specimen from Vietnam possessed black elytra). Female specimens (n=2) differ from the male specimens in the following respects: Color: Head, pronotum, scutellum, propygidium, pygidium, and venter shining black; elytra black or dark reddish-brown with iridescent bloom. Elytron: Epipleuron in ventral view (Fig. 75) broadly expanded from base to apex of metacoxa, weakly convex, not incised at apex, with sparse, setose punctures, setae reddish, moderately long; in dorsal view expansion not developed (lacking dorsal pillow), instead with concave groove adjacent to epipleuron. Propygidium: Surface moderately densely punctate; punctures simple and ocellate, mixed, not setigerous. Pygidium: Surface moderately densely punctate; punctures simple and ocellate, not setigerous. Legs: Protibia tridentate. Proclaws of female 3/4 length of protarsomere 5, claw angled ventrally.
Figures 1–12.

Peltonotus species dorsal habitus. 1 Peltonotus animus, male 2–3 Peltonotus cybele, male and female (respectively) 4 Peltonotus favonius, male 5–7 Peltonotus fujiokai, males (showing variation) 8 Peltonotus fujiokai, female 9 Peltonotus karubei, male 10–11 Peltonotus malayensis, male and female (respectively) 12 Peltonotus morio, male.
New distributional records for Peltonotus morio
Peltonotus morio Burmeister (Fig. 12) is the type species for the genus Peltonotus and is one of the most wide-spread species in the genus (Fig. 92). It is distinguished from its close congener, Peltonotus nasutus Arrow (Figs 14–15), by its incomplete pronotal basal bead (complete in Peltonotus nasutus), form of the male parameres (Figs 65–66), lack of a small tubercle at the apex of the clypeus in the male (Fig. 24) (present in Peltonotus nasutus [Fig. 23]), and form of the epipleuron in females (Figs 80 versus 82).
Figures 61–66.

Male parameres (with or without phallobase), dorsal and lateral views, in Peltonotus. Male parameres are highly asymmetrical, and we illustrate the lateral view that best assists in identification. 61 Peltonotus fujiokai 62 Peltonotus gracilipodus and Peltonotus podocrassus 63 Peltonotus karubei 64 Peltonotus malayensis 65 Peltonotus morio 66 Peltonotus nasutus.
The species is found in northeastern India, Nepal, Bhutan, and Thailand (Jameson and Wada 2004). It can be collected at lights (Dhoj et al. 2009). Within the Palearctic region (Löbl and Smetana 2003) or Sino-Japanese region (Holt et al. 2013), it is the only recorded species of Cyclocephalini (Dynastinae), and it was recorded from Bhutan, Nepal, and Sikkim (Krell 2006). Examination of additional specimens provided new country records for Peltonotus morio in Myanmar and Vietnam. New Country Record: MYANMAR (2 specimens deposited in FUJI): Tanintharyi (near Tenasserim), May-1992, 1 male; Mt. Dawna, May-1992, 763 m elevation, 1 female. New Country Record: VIETNAM (1 specimen deposited in IRSNB): Lào Cai Province, June 10, 1917, 1 male. Despite the antiquity of the specimen (nearly 100 years old), the new record in Vietnam extends the known range of the species over 600 km from northern Thailand to northern Vietnam. Based on these distributional data, Peltonotus morio and Peltonotus nasutus may be narrowly sympatric in southern Myanmar and Thailand.
Key to Male Peltonotus Species
Males: Protibial claws with one claw enlarged and expanded; elytral epipleuron not developed in ventral view. Males of Peltonotus kyojinus, Peltonotus nethis, Peltonotus pruinosus, Peltonotus suehirogarus, Peltonotus mushiyaus, and Peltonotus tigerus are not known.
| 1 | Apical half of mentum acute, triangular (e.g., Figs 25, 34–35) | 2 |
| – | Apical half of mentum rounded (Figs 26–29, 31–33) or quadrate (Fig. 30) | 4 |
| 2 | Punctures of frons and clypeus unisetigerous; parameres as in Fig. 71 | Peltonotus talangensis Jameson & Jakl |
| – | Punctures of frons and clypeus multisetigerous (at least laterally); parameres not as in Fig. 71 | 3 |
| 3 | Smaller protarsal claw deeply arcuate (Fig. 54); parameres as in Fig. 59 | Peltonotus deltamentum Jameson & Wada |
| – | Smaller protarsal claw simply arched; parameres as in Fig. 70 | Peltonotus sisyrus Jameson & Wada |
| 4 | Apex of labrum weakly sinuate (Figs 23–24) | 5 |
| – | Apex of labrum bi-emarginate (Figs 21–22) to deeply bilobed (Fig. 20) | 6 |
| 5 | Protibia tridentate with well-developed basal tooth (e.g., Fig. 46); apex of clypeus at middle with tubercle (Fig. 23); parameres as in Fig. 66 | Peltonotus nasutus Arrow |
| – | Protibia tridentate with weakly developed basal tooth (e.g., Fig. 49); apex of clypeus lacking tubercle (Fig. 24); parameres as in Fig. 65 | Peltonotus morio Burmeister |
| 6 | Labrum with apex deeply bilobed (e.g.,Fig. 20) | 7 |
| – | Labrum with apex bi-emarginate (Figs 21–22) | 10 |
| 7 | Mala of maxilla with setae thick and strongly flattened (with well developed lamellate setal brush); Borneo, Malaysia, and Sumatra; parameres not as in Fig. 63 | 8 |
| – | Mala of maxilla with setae not thick and strongly flattened (lacking well developed lamellate setal brush) (Fig. 40); South Vietnam; parameres as in Fig. 63 | Peltonotus karubei Muramoto |
| 8 | Punctures of frons lacking setae; parameres as in Fig. 57 | Peltonotus brunnipennis Benderitter |
| – | Punctures of frons with dense, velutinous and/or moderately long setae; parameres not as in Fig. 57 | 9 |
| 9 | Protarsus with larger claw gracile, subequal at middle and base; maxillary stipes with setae curly at apex (e.g.,Fig. 41); Sarawak | Peltonotus gracilipodus Jameson & Wada |
| – | Protarsus with larger claw robust, much wider at middle than at base; maxillary stipes with setae straight, not curly at apex; Malaysia (Cameron Highlands) | Peltonotus podocrassus Jameson & Wada |
| 10 | Labial palpomere 2 greatly enlarged and dorsoventrally flattened, 2–3 times wider than apical palpomere 1 (Fig. 28) | 11 |
| – | Labial palpomere 2 not greatly enlarged and flattened, less than 1.5 times wider than apical palpomere 1 (Fig. 33) | 13 |
| 11 | Maxillary stipes with setae curly at apex (e.g., Fig. 36); parameres not as in Fig. 68 | 12 |
| – | Maxillary stipes with setae straight, not curly at apex; parameres as in Fig. 68 | Peltonotus silvanus Jameson & Wada |
| 12 | Elytral color reddish, lighter in color than pronotum and scutellum; punctures of pygidium multisetigerous, setae minute and moderate in length; parameres as in Fig. 64 | Peltonotus malayensis Arrow |
| – | Elytral color castaneous, similar in color to pronotum and scutellum (Fig. 1); punctures of pygidium unisetigerous, setae moderate in length; parameres as in Fig. 56 | Peltonotus animus Jameson & Wada |
| 13 | Protibia tridentate, basal tooth well developed or weakly developed (Figs 46, 48, 49) | 14 |
| – | Protibia bidentate (Fig. 47, 51) | 15 |
| 14 | Protibia with basal tooth well developed (Figs 46, 48), external margin without velutinous setae from middle to near base; parameres as in Fig. 61 | Peltonotus fujiokai Jameson & Wada |
| – | Protibia externally with basal tooth weakly developed (Fig. 49), external margin with velutinous setae from middle to near base; parameres as in Fig. 67 | Peltonotus rubripennis Miyake & Yamaya |
| 15 | Elytra reddish with castaneous vittae (Figs 18–19); parameres as in Fig. 72 | Peltonotus vittatus Arrow |
| – | Elytra lacking vittae, entirely reddish, castaneous, or black; parameres not as in Fig. 72 | 16 |
| 16 | Pronotal basal bead lacking, terminating at basolateral angle; length less than 15.0 mm; parameres as in Fig. 60 | Peltonotus favonius Jameson & Wada |
| – | Pronotal basal bead present, extending beyond basolateral angle (obscured anterior to scutellum); length greater than 17.0 mm; parameres not as in Fig. 60 | 17 |
| 17 | Protarsomere 5 with well-developed internoapical protrusion (Fig. 50), lacking weak medial protrusion; region surrounding Mt. Bawang, Kalimantan | Peltonotus adelphosimilis Jameson & Wada |
| – | Protarsomere 5 lacking internoapical protrusion; weak protrusion at middle (Fig. 53); Sabah | Peltonotus similis Arrow |
Key to Female Peltonotus Species
Females: Protibial claws similar in size and shape; elytral epipleuron developed or simple in ventral view. Females of Peltonotus deltamentum, Peltonotus karubei, and Peltonotus animus are not known.
| 1 | Apical half of mentum acute, triangular (Figs 25, 34–35) | 2 |
| – | Apical half of mentum rounded (Figs 26–29, 31–33) or quadrate (Fig. 30) | 3 |
| 2 | Punctures of frons and clypeus multisetigerous | Peltonotus sisyrus Jameson & Wada |
| – | Punctures of frons and clypeus unisetigerous | Peltonotus talangensis Jameson & Jakl |
| 3 | Apex of labrum weakly sinuate (Figs 23–24) | 4 |
| – | Apex of labrum bi-emarginate (Figs 21–22) to deeply bilobed (Fig. 20) | 5 |
| 4 | Apex of clypeus with weak, medial tubercle; lateral pillow of elytron (dorsal view) elongate-oval, extending more than half length of epipleuron; epipleuron as in Fig. 82 | Peltonotus nasutus Arrow |
| – | Apex of clypeus lacking weak tubercle; lateral pillow of elytron (dorsal view) narrower at apex and broader at base, extending less than half length of epipleuron; epipleuron as in Fig. 80 | Peltonotus morio Burmeister |
| 5 | Elytra with castaneous vittae or maculae (e.g., Figs 13, 18–19) | 6 |
| – | Elytra lacking vittae, entirely castaneous, reddish, or black | 7 |
| 6 | Elytral epipleuron in ventral view simple, lacking apical incision (Fig. 81) | Peltonotus mushiyaus Jameson & Wada |
| – | Elytral epipleuron in ventral view incised at apex (Fig. 91) | Peltonotus vittatus Arrow |
| 7 | Labrum with apex deeply bilobed (e.g., Fig. 20) | 8 |
| – | Labrum with apex bi-emarginate (e.g.,Figs 21–22) | 13 |
| 8 | Elytral epipleuron in ventral view simple, not emarginated (Fig. 83) | Peltonotus nethis Jameson & Wada |
| – | Elytral epipleuron in ventral view emarginated (e.g., Fig. 73) | 9 |
| 9 | Maxillary stipes with setae curly at apex (e.g., Fig. 41) | 10 |
| – | Maxillary stipes with setae straight, not curly at apex | 11 |
| 10 | Epipleural emargination with well-developed tooth in ventral view (Fig. 73) | Peltonotus brunnipennis Benderitter |
| – | Epipleural emargination with moderately developed tooth in ventral view (Fig. 77) | Peltonotus gracilipodus Jameson & Wada |
| 11 | Elytra entirely reddish (Fig. 17) | Peltonotus tigerus Jameson & Wada |
| – | Elytra entirely black | 12 |
| 12 | Lateral pillow of elytron (dorsal view) well-developed, extending medially at least ¼ elytral width, visible in ventral view (Fig. 88) | Peltonotus suehirogarus Jameson & Wada |
| – | Lateral pillow of elytron (dorsal view) moderately developed, extending medially about 1/8 elytral width, not visible in ventral view (Fig. 77) | Peltonotus podocrassus Jameson & Wada |
| 13 | Elytral epipleuron in ventral view broad, nearly parallel from base to near metacoxa, lacking emargination (Fig. 75, 84) | 14 |
| – | Elytral epipleuron in ventral view narrowing from base to near metacoxa (not parallel-sided), with or without emargination (e.g., Figs 76, 78–79) | 15 |
| 14 | Elytral epipleuron in ventral view with sparse, reddish, moderately long setae | Peltonotus favonius Jameson & Wada |
| – | Elytral epipleuron in ventral view without setae | Peltonotus pruinosus Arrow |
| 15 | Labial palpomere 2 greatly enlarged and dorsoventrally flattened, 2–3 times wider than palpomere 1 (e.g., Fig. 28) | 16 |
| – | Labial palpomere 2 not greatly enlarged and flattened, at most 1.5 times wider than palpomere 1 (e.g., Fig. 33) | 17 |
| 16 | Maxillary stipes with setae curly at apex (Fig. 41); lateral pillow of elytron (dorsal view) well-developed, visible in ventral view (Fig. 79) | Peltonotus malayensis Arrow |
| – | Maxillary stipes with setae straight, not curly at apex; lateral pillow of elytron (dorsal view) moderately developed, not visible in ventral view (Fig. 77) | Peltonotus silvanus Jameson & Wada |
| 17 | Body length more than 20 mm; epipleuron in ventral view simple, not emarginate (Fig. 78) | Peltonotus kyojinus Jameson & Wada |
| – | Body length less than 20 mm; epipleuron in ventral view simple or emarginate (Figs 74, 76, 85–86) | 18 |
| 18 | Elytral epipleuron emarginate in ventral view (Fig. 86) | 19 |
| – | Elytral epipleuron simple in ventral view (Figs 76, 85) | 21 |
| 19 | Elytral epipleuron in ventral view with round emargination (Figs 74, 86); not occurring in Mt. Bawang, Kalimantan region of Borneo | 20 |
| – | Elytral epipleuron in ventral view with elongate-oval emargination; Mt. Bawang, Kalimantan region of Borneo | Peltonotus adelphosimilis Jameson & Wada |
| 20 | Punctures of frons and clypeus mulitsetigerous, setae minute and/or short; elytral epipleuron as in Fig. 86; Borneo | Peltonotus similis Arrow |
| – | Punctures of frons and clypeus unisetigerous, setae minute; elytral epipleuron as in Fig. 74; Sumatra | Peltonotus cybele Jameson & Wada |
| 21 | Elytral epipleuron in ventral view terminating near metacoxa (Fig. 85) | Peltonotus rubripennis Miyake & Yamaya |
| – | Elytral epipleuron in ventral view extending posterior of metacoxa, terminating near sternite 3 (Fig. 76) Peltonotus fujiokai Jameson & Wada |
Peltonotus species diagnoses
Peltonotus adelphosimilis
Jameson & Wada, 2004
http://species-id.net/wiki/Peltonotus_adelphosimilis
Diagnosis (male and female).
Length 20.3–18.9 mm, color overall black or castaneous, elytra black or castaneous with or without iridescent bloom, head with some multisetigerous punctures, labrum bi-emarginate, mentum rounded in apical half, labial palpomere 2 not enlarged or obviously dorsoventrally flattened, mala lacking lamellate setal brush, maxillary stipes without setae curled at apices, male protibia bidentate, protarsomere 5 of male with internoapical protuberance (Fig. 50), form of parameres (Fig. 55), female epipleuron incised and with rounded emargination (similar to Peltonotus similis, Fig. 86).
Distribution.
Indonesia, Borneo Island (Kalimantan).
Peltonotus animus
Jameson & Wada, 2009
http://species-id.net/wiki/Peltonotus_animus
Diagnosis (male only).
Length ~16.5 mm, color overall castaneous, elytra castaneous with weak iridescent bloom (Fig. 1), frons with some multisetigerous punctures, labrum bi-emarginate, mentum rounded in apical half, labial palpomere 2 enlarged and dorsoventrally flattened, mala with dense lamellate setal brush (Fig. 36), maxillary stipes with some setae curled at apices (Fig. 36), male protibia tridentate with basal tooth obsolete (Fig. 45), and male parameres (Fig. 56).
Distribution.
Indonesia, Sumatra Island.
Peltonotus brunnipennis
Benderitter, 1934
http://species-id.net/wiki/Peltonotus_brunnipennis
Diagnosis (male and female).
Length 14.5–16.9 mm, color overall castaneous, elytra reddish-orange or black with iridescent bloom, head punctate and lacking setae, labrum deeply bi-lobed, mentum rounded in apical half, labial palpomere 2 enlarged and obviously dorsoventrally flattened, mala with lamellate setal brush, maxillary stipes with some setae curled at apices, male protibia tridentate, form of parameres (Fig. 57), female epipleuron incised and with oval emargination (Fig. 73).
Distribution.
Malaysia, Borneo Island (Sabah and Sarawak).
Peltonotus cybele
Jameson & Wada, 2009
http://species-id.net/wiki/Peltonotus_cybele
Diagnosis (male and female).
Length 14.5–16.5 mm, color overall castaneous, elytra castaneous suffused with dark red or reddish-brown and iridescent bloom (Figs 2–3), head with some unisetigerous punctures, labrum bi-emarginate, mentum rounded in apical half, labial palpomere 2 not enlarged or obviously dorsoventrally flattened, mala lacking lamellate setal brush (Fig. 37), maxillary stipes without setae curled at apices (Fig. 37), male protibia tridentate (Fig. 46), form of parameres (Fig. 58), female epipleuron incised and with rounded emargination (Fig. 74).
Distribution.
Indonesia, Sumatra Island.
Peltonotus deltamentum
Jameson & Wada, 2004
http://species-id.net/wiki/Peltonotus_deltamentum
Diagnosis (male only).
Length ~16.6 mm, color overall castaneous, elytra castaneous with weak iridescent bloom, head with some multisetigerous punctures, labrum bi-emarginate, mentum triangular in apical half (Fig. 25), labial palpomere 2 enlarged and dorsoventrally flattened, mala with dense lamellate setal brush (Fig. 38), maxillary stipes with setae curled at apices (Fig. 38), male protibia tridentate with basal tooth weakly developed, male proclaw strongly arcuate in ventral view (Fig. 54), form of parameres (Fig. 59).
Distribution.
Indonesia, Borneo Island (Kalimantan).
Peltonotus favonius
Jameson & Wada, 2009
http://species-id.net/wiki/Peltonotus_favonius
Diagnosis (male and female).
Length ~14.6 mm, color overall black, elytra black or dark reddish brown with iridescent bloom (Fig. 4), head with simple punctures (lacking setae), labrum bi-emarginate, mentum rounded in apical half, labial palpomere 2 not enlarged or obviously dorsoventrally flattened, mala lacking lamellate setal brush (Fig. 39), maxillary stipes without setae curled at apices (Fig. 39), male protibia bidentate (Fig. 51), form of parameres (Fig. 60), female epipleuron broadly expanded, weakly convex, extending from base to metacoxa, lacking incised apex (Fig. 75).
Distribution.
Vietnam and Myanmar.
Remarks.
This species is most similar to Peltonotus pruinosus, a species for which only the female holotype is known. Previously, this species was only known from the male holotype specimen from Vietnam.
Peltonotus fujiokai
Jameson & Wada, 2004
http://species-id.net/wiki/Peltonotus_fujiokai
Diagnosis (male and female).
Length 14.1–14.6 mm, color overall castaneous, elytra reddish-brown with castaneous vittae, reddish-brown, or black with iridescent bloom (Figs 5–8), head with some unisetigerous punctures, labrum bi-emarginate, mentum rounded in apical half, labial palpomere 2 not enlarged and not dorsoventrally flattened, mala without dense lamellate setal brush, maxillary stipes without setae curled at apices, male protibia tridentate, form of parameres (Fig. 61), female epipleuron simple, not incised and lacking emargination (Fig. 76).
Distribution.
Indonesia, Borneo Island (Kalimantan); Malaysia, Borneo Island (Sabah).
Peltonotus gracilipodus
Jameson & Wada, 2004
http://species-id.net/wiki/Peltonotus_gracilipodus
Diagnosis (male and female).
Length 14.4–16.8 mm, color overall castaneous, elytra castaneous with weak iridescent bloom, head with some multisetigerous punctures, labrum deeply bi-lobed, mentum rounded in apical half (Fig. 26), labial palpomere 2 enlarged and obviously dorsoventrally flattened (Fig. 26), mala with lamellate setal brush, maxillary stipes with some setae curled at apices, male protibia bidentate, form of parameres (Fig. 62), female epipleuron incised and with oblong-oval emargination (Fig. 77).
Distribution.
Indonesia, Sumatra Island.
Remarks.
Peltonotus gracilipodus and Peltonotus podocrassus (distributed in peninsular Malaysia) have quite similar male parameres and females have quite similar epipleura, perhaps indicating recent isolation of ancestral populations.
Peltonotus karubei
Muramoto, 2000
http://species-id.net/wiki/Peltonotus_karubei
Diagnosis (male only).
Length 13.4–14.5 mm, overall color black or castaneous, elytra reddish orange or black with iridescent bloom (Fig. 9), head with some multisetigerous punctures, labrum deeply bilobed (Fig. 20), labial palpomere 2 enlarged and obviously dorsoventrally flattened (Fig. 40), mala with weak lamellate setal brush (Fig. 40), maxillary stipes without setae curled at apices, male protibia bidentate, form of male parameres (Fig. 63).
Distribution.
Vietnam (southern).
Peltonotus kyojinus
Jameson & Wada, 2004
http://species-id.net/wiki/Peltonotus_kyojinus
Diagnosis (female only).
Length 21.3 mm, color overall castaneous, elytral disc brown with iridescent bloom, head with some multisetigerous punctures, labrum bi-emarginate, mentum rounded in apical half (Fig. 27), labial palpomere 2 not enlarged and not obviously dorsoventrally flattened, mala without lamellate setal brush, maxillary stipes without setae curled at apices, female epipleuron simple, not incised and lacking emargination (Fig. 78).
Distribution.
Indonesia, Borneo Island (Kalimantan).
Remarks.
Peltonotus kyojinus is the largest species of Peltonotus.
Peltonotus malayensis
Arrow, 1910
http://species-id.net/wiki/Peltonotus_malayensis
Figs 10–11 , 21 , 28 , 41 , 47 , 64 , 79
Diagnosis (male and female).
Length 14.4–17.2 mm, color overall castaneous or black, elytra reddish-brown or black with weak iridescent bloom (Figs 10–11), head with some multisetigerous punctures, labrum bi-emarginate (Fig. 21), mentum rounded in apical half (Fig. 28), labial palpomere 2 enlarged and obviously dorsoventrally flattened (Fig. 41), mala with weak lamellate setal brush, maxillary stipes setae curled at apices (Fig. 41), male protibia bidentate (Fig. 47), form of male parameres (Fig. 64), female epipleuron incised and with rounded emargination (Fig. 79).
Distribution.
Brunei; Indonesia, Borneo Island (Kalimantan); Malaysia, Borneo Island (Sarawak).
Peltonotus morio
Burmeister, 1847
http://species-id.net/wiki/Peltonotus_morio
Diagnosis (male and female).
Length 14.0–18.0 mm, color overall black or castaneous, elytra black or castaneous and shining (Fig. 12), head with unisetigerous punctures, labrum weakly sinuate (Fig. 24), mentum quadrate in apical half, labial palpomere 2 not enlarged and not dorsoventrally flattened, mala lacking lamellate setal brush, maxillary stipes without setae curled at apices, male protibia tridentate with basal tooth weakly developed, form of male parameres (Fig. 65), female epipleuron weakly, quadrately incised (Fig. 80) and with moderately developed dorsal pillow.
Distribution
(Fig. 92). Bhutan, India (northeastern), Myanmar, Nepal, Thailand, Vietnam.
Peltonotus mushiyaus
Jameson & Wada, 2009
http://species-id.net/wiki/Peltonotus_mushiyaus
Diagnosis (female only).
Length ~11.8 mm, overall color castaneous, elytral disc orangish-tan with castaneous maculae and iridescent bloom (Fig. 13), head with some unisetigerous punctures, labrum bi-emarginate, mentum rounded in apical half, labial palpomere 2 not enlarged or obviously dorsoventrally flattened, mala lacking lamellate setal brush (Fig. 42), maxillary stipes without setae curled at apices (Fig. 42), female epipleuron simple, not expanded (Fig. 81).
Distribution.
Malaysia, Borneo Island (Sabah).
Remarks.
Peltonotus mushiyaus is the smallest species in the genus. We hypothesize that males of this species will possess orangish-tan elytra with castaneous maculae, similar to males of Peltonotus vittatus.
Peltonotus nasutus
Arrow, 1910
http://species-id.net/wiki/Peltonotus_nasutus
Figs 14–15 , 23 , 30 , 52 , 66 , 82 , 92
Diagnosis (male and female).
Length 19.6–20.6 mm, color overall black or castaneous, elytra black or castaneous and shining (Fig. 14–15), head with unisetigerous punctures and apex of clypeus with weak tubercle medially (Fig. 23), labrum weakly sinuate (Fig. 23), mentum quadrate in apical half (Fig. 30), labial palpomere 2 not enlarged and not dorsoventrally flattened, mala lacking lamellate setal brush, maxillary stipes without setae curled at apices, male protibia tridentate with well developed basal tooth, male protibial claw greatly enlarged (Fig. 52), form of male parameres (Fig. 66), female epipleuron weakly, quadrately incised (Fig. 82) and with well developed dorsal pillow.
Distribution
(Fig. 92). Cambodia, China (southern), Laos, Myanmar, Thailand, Vietnam.
Remarks.
Peltonotus nasutus is the most common species in the genus and the only species with an apicomedial tubercle on the clypeus (male only).
Peltonotus nethis
Jameson & Wada, 2004
http://species-id.net/wiki/Peltonotus_nethis
Diagnosis (female only).
Length ~13.7 mm, color overall black, elytra black with iridescent bloom, head with unisetigerous punctures or lacking setae, labrum bi-emarginate, mentum rounded in apical half (Fig. 31), labial palpomere 2 greatly enlarged and dorsoventrally flattened, mala with lamellate setal brush, maxillary stipes without setae curled at apices, female epipleuron simple, not incised (Fig. 83).
Distribution.
Malaysia, Borneo Island (Sabah).
Peltonotus podocrassus
Jameson & Wada, 2004
http://species-id.net/wiki/Peltonotus_podocrassus
Diagnosis (male and female).
Length 17.6–18.7 mm, color overall castaneous, elytra castaneous with weak iridescent bloom, head with some multisetigerous punctures, labrum deeply bi-lobed, mentum rounded in apical half (Fig. 29), labial palpomere 2 enlarged and obviously dorsoventrally flattened (Fig. 29), mala with lamellate setal brush, maxillary stipes lacking setae curled at apices, male protibia bidentate, form of parameres (Fig. 62), female epipleuron incised and with oblong-oval emargination (Fig. 77).
Distribution.
Malaysia (Peninsular Malaysia).
Remarks.
Peltonotus podocrassus and Peltonotus gracilipodus (distributed in Sumatra) are similar with respect to the male parameres and female epipleura. This may be indicative of recent divergence from a common ancestor.
Peltonotus pruinosus
Arrow, 1910
http://species-id.net/wiki/Peltonotus_pruinosus
Diagnosis (female only).
Length ~15.7 mm, color overall black, elytra black with iridescent bloom, head punctate and lacking setae, labrum bi-emarginate, mentum rounded in apical half and moderately bi-lobed at middle (Fig. 32), labial palpomere 2 not enlarged and not obviously dorsoventrally flattened (Fig. 32), mala without lamellate setal brush, maxillary stipes without setae curled at apices, female epipleuron broadly expanded and lacking emargination at apex (Fig. 84).
Distribution.
India.
Peltonotus rubripennis
Miyake & Yamaya, 1994
http://species-id.net/wiki/Peltonotus_rubripennis
Diagnosis (male and female).
Length 12.0–12.5 mm, color overall castaneous, elytral disc brown with iridescent bloom, head with unisetigerous punctures, labrum bi-emarginate, mentum rounded in apical half, labial palpomere 2 slightly enlarged and not obviously dorsoventrally flattened, mala lacking lamellate setal brush, maxillary stipes lacking setae curled at apices, male protibia tridentate with basal tooth weakly developed (Fig. 49), form of parameres (Fig. 67), female epipleuron simple and lacking emargination at apex (Fig. 85).
Distribution.
Malaysia, Borneo Island (Sabah and Sarawak).
Peltonotus silvanus
Jameson & Wada, 2004
http://species-id.net/wiki/Peltonotus_silvanus
Diagnosis (male and female).
Length 16.3–17.8 mm, color overall castaneous, elytra castaneous, dark-brown, or black with weak iridescent bloom, head with some multisetigerous punctures, labrum bi-emarginate, mentum rounded in apical half, labial palpomere 2 enlarged and obviously dorsoventrally flattened, mala with lamellate setal brush, maxillary stipes lacking setae curled at apices, male protarsomeres 2–4 with apices expanded, male protibia bidentate, form of parameres (Fig. 68), female epipleuron incised and with oblong-oval emargination (Fig. 77).
Distribution.
Indonesia, Borneo Island (Kalimantan); Malaysia, Borneo Island (Sarawak).
Peltonotus similis
Arrow, 1931
http://species-id.net/wiki/Peltonotus_similis
Diagnosis (male and female).
Length 18.0–20.9 mm, color overall dark brown or black, elytra dark brown or black with or without iridescent bloom, head with some multisetigerous punctures, labrum bi-emarginate, mentum rounded in apical half (Fig. 33), labial palpomere 2 slightly enlarged and not obviously dorsoventrally flattened (Fig. 33), mala without lamellate setal brush, maxillary stipes without setae curled at apices, protarsomere 5 of male with internomedial protuberance (Fig. 53), male protibia bidentate, form of parameres (Fig. 69), female epipleuron incised and with rounded emargination (Fig. 86).
Distribution.
Malaysia, Borneo Island (Sabah).
Peltonotus sisyrus
Jameson & Wada, 2004
http://species-id.net/wiki/Peltonotus_sisyrus
Diagnosis (male and female).
Length 16.1–16.4 mm, overall castaneous, elytra castaneous with weak iridescent bloom, head with some punctures multisetigerous, labrum bi-emarginate, mentum triangular in apical half (Fig. 34), labial palpomere 2 enlarged and obviously dorsoventrally flattened (Fig. 34), mala with lamellate setal brush, maxillary stipes without setae curled at apices, male protibia bidentate, form of parameres (Fig. 70), female epipleuron incised and with broad, elongate emargination (Fig. 87).
Distribution.
Indonesia, Sumatra Island.
Peltonotus suehirogarus
Jameson & Wada, 2004
http://species-id.net/wiki/Peltonotus_suehirogarus
Diagnosis (female only).
Length 16.9–18.0 mm, color overall black, elytra black with iridescent bloom, head with some multisetigerous punctures, labrum bi-emarginate, mentum rounded in apical half, labial palpomere 2 enlarged and obviously dorsoventrally flattened, mala with lamellate setal brush, maxillary stipes with some setae weakly curled at apices, female epipleuron incised and with oblong-oval emargination (Fig. 88).
Distribution.
Indonesia, Borneo Island (Kalimantan); Malaysia, Borneo Island (Sarawak).
Peltonotus talangensis
Jameson & Jakl, 2010
http://species-id.net/wiki/Peltonotus_talangensis
Figs 16 , 35 , 43 , 48 , 71 , 89
Diagnosis (male and female).
Length 14.1–15.2 mm, color overall castaneous, elytra castaneous or with weak reddish tones and lacking iridescent bloom (Fig. 16), head with some punctures unisetigerous, labrum bi-emarginate, mentum triangular in apical half (Fig. 35), labial palpomere 2 enlarged and obviously dorsoventrally flattened (Fig. 35), mala with lamellate setal brush (Fig. 43), maxillary stipes without setae curled at apices (Fig. 43), male protibia tridentate (Fig. 48), form of parameres (Fig. 71), female epipleuron simple, not incised (Fig. 89).
Distribution.
Indonesia, Sumatra Island.
Peltonotus tigerus
Jameson & Wada, 2009
http://species-id.net/wiki/Peltonotus_tigerus
Diagnosis (female only).
Length ~13.7 mm, overall color black or castaneous, elytra reddish-brown with weak iridescent bloom (Fig. 17), head with some punctures multisetigerous, labrum bi-emarginate, labial palpomere 2 enlarged and dorsoventrally flattened, mala with well developed lamellate setal brush (Fig. 44), maxillary stipes without setae curled at apices (Fig. 44), female epipleuron incised with a round or oval emargination (Fig. 90).
Distribution.
Thailand.
Remarks.
We hypothesize that males of this species will possess reddish-brown elytra, similar to the coloration of the female.
Peltonotus vittatus
Arrow, 1910
http://species-id.net/wiki/Peltonotus_vittatus
Diagnosis (male and female).
Length 12.3–14.4 mm, color overall black or castaneous with pronotum reddish or black and with dark discal maculae, elytra reddish and with dark discal maculae and iridescent bloom (Figs 18–19), head with some multisetigerous punctures, labrum bi-emarginate (Fig. 22), mentum rounded in apical half, labial palpomere 2 not enlarged and not obviously dorsoventrally flattened, mala without lamellate setal brush, maxillary stipes without setae curled at apices, male protibia bidentate (or tridentate with basal tooth weakly developed), form of parameres (Fig. 72), female epipleuron narrowly incised (Fig. 91) with well developed dorsal pillow.
Distribution.
Malaysia, Borneo Island (Sabah and Sarawak).
Supplementary Material
Acknowledgements
We kindly thank the curators and collections managers who assisted in providing specimens for this research (see Materials and Methods): Max Barclay and Malcolm Kerley (BMNH), Patrick Grootaert (IRSNB), Matt J. Paulsen and Brett Ratcliffe (USNM currently housed as UNSM), and Andreas Reichenbach (AREC). Shinji Nagai (Japan) kindly provided specimens of Peltonotus favonius. Matt R. Moore and David Wickell (both Wichita State University) are thanked for imaging of some specimens and characters for this work, and Aura Paucar (University of Nebraska) is thanked for illustrations for some specimens and characters.
Appendix
Supplemental file for dynamic mapping. (doi: 10.3897/zookeys.320.5352.app) Microsoft Excel document (xls).
Explanation note: Distribution maps were generated by entering latitude and longitude data into Microsoft Excel 2008 and uploaded to EarthPoint (http://www.earthpoint.us/ExcelToKml.aspx) and GoogleEarth (http://www.google.com/earth/index.html). This supplementary file allows addition of data and interactive mapping or niche modeling. Please note, however, that older localities have a wide margin of error and their lack of precision is not conducive to ecological or niche modeling.
References
- Arrow GJ. (1910) On the lamellicorn beetles of the Peltonotus with descriptions of four new species. Annals and Magazine of the Natural History (Series 8) 5: 153–157. [Google Scholar]
- Chen PP, Wongsiri S, Jamyanya T, Rinderer TE, Vongsamanode S, Matsuka M, Sylvester HA, Oldroyd BP. (1998) Honey bees and other edible insects used as human food in Thailand. American Entomologist Spring: 24–29.
- Danell E. (2010) Dokmai dogma: Amorphophallus and its edible beetles. http://dokmaidogma.wordpress.com/2010/05/27/amorphophallus-and-its-edible-beetles/ [accessed March 19 2013]
- Dhoj YGC, Keller S, Nage P, Kafle L. (2009) Abundance and diversity of scarabaeid beetles (Coleoptera: Scarabaeidae) in different farming areas in Nepal. Formosan Entomology 29: 103–112. [Google Scholar]
- Gibernau M, Chartier M, Barabé D. (2010) Recent advances towards an evolutionary comprehension of Araceae pollination. In: Seberg O, Petersen G, Barfod AS, Davis JI, Editors. Diversity, phylogeny, and evolution in the Monocotyledons. >Aarhus University Press, Denmark, 101–114. [Google Scholar]
- Grimm R. (2009) Peltonotus nasutus Arrow, 1910 und Phaeochrous-Arten als Bestäuber von Amorphophallus paeoniifolius (Araceae) in Thailand (Coleoptera: Scarabaeidae). Entomologische Zeitschrift 119: 167-168. [Google Scholar]
- Holt BG, Lessard J-P, Borregaard MK, Fritz SA, Araújo MB, Dimitrov D, Fabre P-H, Graham CH, Graves GR, Jønsson KA, Nogués-Bravo D, Wang Z, Whittaker RJ, Fjeldså J, Rahbek (2013) An Update of Wallace’s Zoogeographic Regions of the World. Science 339: 74-78. doi: 10.1126/science.1228282 [DOI] [PubMed] [Google Scholar]
- Krell F-T. (2006) Dynastinae MacLeay, 1819. In: Löbl I. & Smetana A. 2006. Catalogue of Palaearctic Coleoptera, volume 3. Scarabaeoidea - Scirtoidea - Dascilloidea - Buprestoidea - Byrrhoidea. Eds Löbl I. & Smetana A. Apollo Books, Stenstrup, Denmark, 277–283. [Google Scholar]
- Jameson ML, Wada K. (2004) Revision of the genus Peltonotus Burmeister (Coleoptera: Scarabaeidae: Dynastinae) from Southeastern Asia. Zootaxa 502: 1-66. [Google Scholar]
- Jameson ML, Wada K. (2009) Five new species of Peltonotus Burmeister (Scarabaeidae: Dynastinae: Cyclocephalini) from Southeast Asia. Insecta Mundi 102: 1-16. [Google Scholar]
- Jameson ML, Jakl S. (2010) Synopsis of the aroid scarabs in the genus Peltonotus Burmeister (Scarabaeidae, Dynastinae, Cyclocephalini) from Sumatra and description of a new species. ZooKeys 34: 141-152. doi: 10.3897/zookeys.34.302 [DOI] [Google Scholar]
- Li J, Keith D, Gao M, Lin L. (2012) Coléoptères Scarabaeoidea de Pha Ngam (province de Bokeo, Laos). L’Entomologiste 68 (5): 273-276. [Google Scholar]
- Löbl I, Smetana A. (2003) Catalogue of Palaearctic Coleoptera, volume 1. Archostemata-Myxophaga-Adephaga. Löbl & Smetana, eds., Apollo Books, Stenstrup, Denmark, 819 pp. [Google Scholar]
- Maia ACD, Gibernau M, Carvalho AT, Gonçalves EG, Schlindwein C. (2012) The cowl does not make the monk: scarab beetle pollination of the Neotropical aroid Taccarum ulei (Araceae, Spathicarpeae). Biological Journal of the Linnean Society (early view online version), 13 pp. doi: 10.1111/j.1095-8312.2012.01985.x [DOI]
- Miyake Y, Yamaya S. (1994) Some new scarabaeid species from southern Asia preserved in the Nagaoka Municipal Science Museum (II). Bulletin of the Nagaoka Municiple Science Museum 29: 37-43. [Google Scholar]
- Moore MR. (2012) A new female elytral character for the tribe Cyclocephalini (Coleoptera: Scarabaeidae: Dynastinae) and an observation of its possible function. The Coleopterists Bulletin 63 (3): 200-202. doi: 10.1649/072.066.0303 [DOI] [Google Scholar]
- Moore MR, Jameson ML. (in press) Floral associations of cyclocephaline scarab beetles. Journal of Insect Science. [DOI] [PMC free article] [PubMed]
- Schiestl FP, Dötterl S. (2012) The evolution of floral scent and olfactory preferences in pollinators: coevolution or pre-existing bias? Evolution 66(7): 2042–2055. doi: 10.1111/j.1558-5646.2012.01593.x [DOI] [PubMed] [Google Scholar]
- Wheeler QD, Platnick NI. (2000) The phylogenetic species concept, 55–69. In: Wheeler QD, Meier R. (Eds) Species Concepts and Phylogenetic Theory: A Debate. Columbia University Press, New York, NY. [Google Scholar]
- Young HJ. (1988) Differential importance of beetle species pollinating Dieffenbachia longispatha (Araceae). Ecology 69: 832-844. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplemental file for dynamic mapping. (doi: 10.3897/zookeys.320.5352.app) Microsoft Excel document (xls).