

Abstract
The repair of chromosomal double strand breaks (DSBs) is crucial in the maintenance of genomic integrity. However, the repair of DSBs can also destabilize the genome by causing mutations and chromosomal rearrangements, the driving forces for carcinogenesis and hereditary diseases. Break induced replication (BIR) is one of the DSB repair pathways that is highly prone to genetic instability1–3. BIR proceeds by invasion of one broken end into a homologous DNA sequence followed by replication that can copy hundreds of kilobasepairs of DNA from a donor molecule all the way through its telomere4,5. The resulting repaired chromosome comes at a great cost to the cell, as BIR promotes mutagenesis, loss of heterozygosity, translocations, and copy number variations, all hallmarks of carcinogenesis4–9. BIR employs the majority of known replication proteins to copy large portions of DNA, similar to S-phase replication10,11. It has thus been suggested that BIR proceeds by semiconservative replication; however, the model of a bona-fide, stable replication fork contradicts the known instabilities associated with BIR such as a 1000-fold increase in mutation rate compared to normal replication9. Here we demonstrate that the mechanism of replication during BIR is significantly different from S-phase replication, as it proceeds via an unusual bubble-like replication fork that results in conservative inheritance of the new genetic material. We provide the evidence that this atypical mode of DNA replication, dependent on Pif1 helicase, is responsible for the dramatic increase in BIR-associated mutations. We propose that the BIR-mode of synthesis presents a powerful mechanism that can initiate bursts of genetic instability in eukaryotes including humans.
Theoretically, BIR might constitute a unidirectional, bona fide replication fork producing two semi-conservatively replicated molecules4,11 (Fig. 1a(i)). Alternatively, a D-loop formed by invasion of the broken chromosome may persist throughout BIR, migrating down the length of the chromosome, creating an unusual condition of conservative inheritance of newly synthesized DNA1,12,13 (Fig. 1a(ii–iv)).
Figure 1. The mode of DNA synthesis during BIR.

a, The models of BIR. (i), Replication fork proceeds semiconservatively. (ii–iv), Migrating bubble leads to conservative inheritance of new DNA. Synchronous (ii) and asynchronous (iii,iv) synthesis of leading and lagging DNA strands. b, (i), The BIR frameshift mutation assay. A DSB is induced at MATa of the recipient chromosome (Chr) III. lys2 reporter is inserted in the donor chromosome 16 or 36 kb telomere-proximal from MATα-inc. Lys+ mutations would be inherited equally by the donor (D) or recipient (R) if BIR is semiconservative (ii), but only by recipient if BIR is conservative (iii).
To distinguish between these models, we used a disomic yeast system (Fig. 1b(i)) containing a second, truncated copy of chromosome III (Chr III), cleaved by HO endonuclease under control of a galactose-inducible promoter2. The HO-induced DSB possesses only one efficiently repairable end that invades the second copy of Chr III, and initiates BIR that copies over 100 kb of the distal part of the chromosome. Using this system, we recently demonstrated that BIR stimulates mutagenesis along the path of DNA synthesis at a series of lys2 frameshift reporters9. Here, we examined these Lys+ mutations to determine whether errors during BIR were acquired semi-conservatively (inherited by either the donor or recipient molecule; Fig.1b(ii)) or conservatively (inherited only by the recipient molecule; Fig. 1b(iii)). Pulse-field gel electrophoresis (PFGE) was used to separate donor and recipient molecules from Lys+ BIR outcomes resulting from mutations in a lys2 reporter located 16 or 36 kb distal to the site of BIR initiation (Fig. 2a, b). Sequencing of the PCR products derived from the separated chromosomes revealed that the great majority of heterozygous frameshift mutations (58 of 58 and 68 of 77 from strains with reporters at 16 and 36 kb, respectively) were inherited by the recipient molecule, while the donor sequence remained unchanged (see also Supplementary Discussion). Overall, the mutation pattern supports a conservative replication mechanism for BIR. However, since this conclusion was based on analysis of selected mutation events, we developed a non-selective test to analyze BIR microscopically by DNA combing.
Figure 2. BIR-induced mutations.

a, The sequencing of the separated donor and recipient chromosomes of heterozygous Lys+ mutants. b, The effect of pif1Δ on BIR-induced frameshifts. Medians of mutation rates are shown. The arrows represent a reduction as compared to wt. c, The assay to study BIR-induced base substitutions in ura3–29 reporter. d, Depending on orientation, the selectable position of ura3–29 leading strand includes cytosine (C) or guanine (G). e, MMS amplifies BIR-induced base substitutions in orientation-dependent way. The arrows indicate an increase as compared to no-MMS control. See Extended Data Tables 1 and 2 for the details of statistical analysis and for the ranges of medians shown in e and b.
The experiments were conducted in nocodazole-arrested cells of disomic BIR strain bearing a cassette facilitating BrdU incorporation in yeast14 (Fig. 3a, b). BrdU was added 3.5 hours following DSB induction. After completion of BIR, PFGE-separated donor and recipient molecules (Fig. 3c; Extended Data Fig. 1), were analyzed by molecular combing and fluorescent in situ hybridization (FISH). We used an anti-BrdU antibody, the P1 probe specific to the tandem repeat of TEF1/BSD inserted 14 kb centromere-proximal to MAT in the donor chromosome, the P2 probe specific to the 20 kb region of Chr III where invasion occurs, and the P3 probe specific to the 15 kb region near the telomere (Fig. 3a) to characterize BIR. We observed BrdU tracts approximately 100 kb in length in 70 of the 98 repaired recipient molecules analyzed (Fig. 3d, e, Extended Data Fig. 2a). These tracts include the entire chromosome region marked by P2 and P3 and, therefore, represent BIR that copied the donor chromosome through to its telomere. Additionally, 14% of recipient molecules contained long (>30 kb) BrdU tracts that overlapped with P2 but not with P3 (Fig. 3d; Extended Data Fig. 3a). These molecules likely represent repair events where BIR was interrupted, resulting in half-crossover formation2,15 (see also Supplementary Discussion).
Figure 3. DNA synthesis during BIR is conservative.

a, Experimental system to assay BIR using dynamic molecular combing including the position of hybridization probes P1, P2 and P3. b, (i), BrdU incorporation in the recipient is expected from conservative BIR (red). (ii), Formation of half-crossovers in pif1Δ leads to short patches of BrdU in recipient. c, Donor and recipient chromosomes separated using PFGE. d, The summary of molecular combing analysis. e, The donors and recipients of wt (PIF1) and pif1Δ. Each molecule was hybridized with P1, P2, P3 probes (green tracts) and treated with anti-BrdU antibodies (red tracts).
Our analysis of donor molecules supports a conservative mode of DNA replication during BIR, as only 4 out of 103 donor molecules were illuminated by >30 kb BrdU tracts (Fig. 3d, e; Extended Data Fig. 2a). These data confirm a strong bias (P<0.0001) towards BrdU tracts present only in the recipient chromosome. The 4 cases of BrdU incorporation in the donor could result from rare semi-conservative synthesis or from BIR initiated >30 kb proximal to the DSB site, which would result in a donor-like size and hybridization pattern due to copying of regions unique to the donor molecule16. Based on these data, we estimate that, even if semiconservative synthesis occurs, it can account for no more than 8% of the BIR events we analyzed (see Supplementary Discussion and Extended Data Fig. 4 for the results of another series of experiments supporting this conclusion).
The unusual mode of replication prompted us to characterize the structure of BIR molecular intermediates at LYS2 inserted ~16 kb from the point of strand invasion. Genomic DNA extracted from nocodazole-arrested cells undergoing BIR was digested with PstI, and analyzed by 2-dimensional (2D) gel electrophoresis using a LYS2-specific probe (Fig. 4a(i)). We detected bubble-like structures between 3 and 7 hours after DSB induction (Fig. 4b–d), but not at 13 hours, consistent with the timing of BIR progression9 (Extended Data Fig. 5). All bubble-like intermediates were markedly different from the Y structures indicative of S-phase replication forks observed before addition of nocodazole and induction of the break (Fig. 4c, 0 Hr). Furthermore, no bubble-like structures were observed in control strains in which HO endonuclease cannot initiate a DSB (Fig. 4d, No-cut), thus linking these structures to BIR exclusively. The bubble-like structures observed in BIR were reminiscent of bubbles routinely detected at replication origins17, with one important difference: the BIR bubbles included a long, high-molecular-weight tail that extended well beyond the size expected for complete replication (arrows in Fig. 4c, d). We hypothesized that initiation of BIR lagging-strand synthesis is often delayed compared to leading strand, resulting in accumulation of single-stranded DNA (ssDNA) behind the BIR bubble, which makes the region around LYS2 refractory to PstI digestion. Indeed, pre-incubation of genomic DNA with oligonucleotides (PstO3 and PstO4; Fig. 4a(ii, iii)) complimentary to the Watson strand of two PstI sites flanking the LYS2 gene eliminated the tail and resulted in a second arc that likely corresponds to molecular intermediates with bubbles consisting of one double-stranded branch (leading-strand synthesis) and one single-stranded branch (lagging-strand synthesis) (Fig. 4a, b, d; Extended Data Fig. 6). Similar results were also obtained using BglII digestion (Extended Data Fig. 7). Importantly, while simultaneous addition of oligonucleotides BglO3 and BglO4, complimentary to the Watson strand of two BglII sites, eliminated the ssDNA tail, the addition of each of these oligos individually failed to eliminate the tail. This confirms that two types of DNA intermediates contribute to the observed ssDNA tail: those containing ssDNA centromere proximal to LYS2 and those with ssDNA distal to LYS2 (Fig. 4a; Extended Data Fig. 7a (ii, iii)). Addition of oligonucleotides complimentary to the Crick strand did not have any effect (data not shown). Bubble migration intermediates were also detected with an HPH-specific probe that hybridizes to the end of the donor chromosome (Fig. 4a, e). These data strongly support a migrating D-loop type of DNA replication18,19.
Figure 4. Molecular intermediates of BIR.

a, D-loop migration during coordinated (i) and uncoordinated (ii, iii) leading- and lagging-strand synthesis. b. Schematic of 2D gel with BIR bubbles forming an arc (1,2) with an extension (3) representing ssDNA tail. Annealing with PstO3 and PstO4 allows PstI digestion changing the mobility of the intermediate (red, 2’). c, 2D analysis of Y-arc during normal replication (0Hr) and bubble-like structures at time points following BIR induction hybridized to LYS2-specific probe. d, High molecular-weight tails (arrows) disappear following annealing with PstO3 and PstO4. The arc is absent in no-cut controls. E, BIR intermediates highlighted with HPH-specific probe.
We hypothesized that ssDNA accumulated behind the migrating BIR bubble is the cause of BIR-associated mutagenesis because of the propensity of ssDNA to accumulate unrepaired DNA lesions20. This was tested by employing methyl methanesulfonate (MMS), a DNA damaging agent that predominantly creates mutagenic lesions in cytosines of ssDNA21,22. In addition, a ura3–29 reporter23, which can revert to Ura+ via three different base substitutions at one C-G pair (Fig. 2c), was inserted in the donor chromosome in two different orientations (Ori1 and Ori2). We expected that MMS will specifically elevate the level of BIR-associated mutagenesis in Ori2, where cytosine is located in the mutant position of the leading (ssDNA) strand, but not in Ori1, which contains guanine instead (Fig. 2d). Indeed, we observed that even though BIR dramatically stimulated base substitutions in ura3–29 irrespectively of its orientations, the effect of MMS was orientation-dependent (Fig. 2e; Extended Data Table 1). Specifically, MMS highly amplified BIR-induced mutagenesis in cells containing ura3–29 in Ori2, while its effect on BIR-mutagenesis in Ori1was relatively modest. This observation supports the conjecture that ssDNA accumulated behind the BIR bubble is the cause of BIR-associated mutagenesis. Additionally, the spectrum of BIR-induced mutations was also orientation dependent, supporting our conclusion (Extended Data Fig. 2b).
Since the Pif1 helicase is a key component of the BIR machinery24 (see also accompanying manuscript by Wilson et al.), we hypothesized that Pif1 is essential for long-range BIR. We observed that, even though BIR-sized products were formed in pif1Δ mutant (Extended Data Fig. 1a,b), no extended BrdU tracts were observed in either the donor or recipient chromosomes (Fig. 3d, e). In addition, approximately 22% of recipient molecules contained short (<20 kb) BrdU patches that co-localized with probe P2 (Fig. 3d, e; Extended Data Fig. 2a) and likely represented DNA synthesis that was prematurely terminated. Therefore, it is likely that the majority of outcomes in pif1Δ mutants formed during the time frame of these experiments were half-crossovers (Extended Data Fig. 1c), supporting our hypothesis that Pif1 is required for BIR-associated DNA synthesis. The low amount of BIR precluded 2D analysis of BIR intermediates in pif1Δ. We investigated whether Pif1 may be necessary for BIR-induced frameshift mutations. Strikingly, we observed that all BIR-induced frameshift mutations were eliminated in the pif1Δ mutant at the 36 kb position, and there was a 20-fold reduction in frameshift mutations at the 16 kb position (Fig. 2b; Extended Data Table 2). Thus, while BIR may initiate in the absence of Pif1, these data support that Pif1 is required for long-range synthesis during BIR. Therefore, Pif1 can be added to the list of other previously identified proteins, including Polδ, Polζ, Msh2, Mlh1, Dun1, and others that are involved in BIR and associated mutagenesis2,9,10,15.
Overall, the results of this study demonstrate that BIR is carried out by a migrating bubble driven by Pif1 with asynchronous synthesis of leading and lagging strands resulting in a mutation-prone accumulation of ssDNA and leads to conservative inheritance of the new genetic material. The bubble migration mechanism and associated mutagenesis may be relevant to cellular processes where BIR has been implicated such as alternative telomere lengthening, and mitochondria maintenance10,25–29, where Pif1 plays an important role. An intriguing possibility is that the burst of mutations recently linked to replication stress/fork collapse in pre-cancerous cells30 may be linked to conservative synthesis initiated by BIR.
Methods
Media, strains, and plasmids
All yeast strains (Extended Data Table 3) were isogenic to AM10032, which is a Chr III disome with the following genotype: hmlΔ::ADE1/hmlΔ::ADE3 MATa-LEU2-tel/MATα-inc hmrΔ::HPH FS2Δ::NAT/FS2 leu2/leu2-3,112 thr4 ura3–52 ade3::GAL::HO ade1 met13.
AM1291 and AM1482 are derivatives of AM1003 and were created by deleting LYS2 from its native location, and inserting lys2-Ins A4(A4) at different positions of Chr III9. AM2191 and AM 2198 were constructed from AM1291 and AM1482 by replacement of PIF1 with KANMX module31. Control strains AM1449, AM1649, AM 2247, AM2257, which contained no HO cut site in the recipient Chr III, were obtained from AM1291, AM1482, AM2191, and AM2198 as previously described9. AM2439 and AM2438 were created by integrating three and two copies of TEF1/BSD-snt1 into SNT1 of AM1291 and AM1482 respectively. The TEF1/BSD-snt1 plasmid was constructed by cloning of a PCR-amplified 1 kb region of SNT1 (from 185626 to 186589 positions of Chr III) into the BamHI/HindIII fragment of TEF1/BSD (Invitrogen). The resulting plasmid was linearized by SnaBI and integrated at SNT1 to introduce a donor-specific region into the MATα-inc containing copy of Chr III. The selection of transformants with integration of multiple copies of the plasmid was achieved by PFGE followed by Southern hybridization with TEF1/BSD used as a probe. AM2118 was isogenic to AM12479, but contained KANMX module at Chr II between PTC4 and TPS1.
AM2110 is a derivative of AM1003, and was created by deleting URA3 (using delitto perfetto approach) and replacing hmr::HPH with hmr::KANMX. In addition, it contains lys2::InsA4 inserted at SED4 (36 kb centromere distal to MATα-inc). AM2161 and AM2820 were derivatives of AM2110 where ura3–29-HPH fragments (Ori1 and Ori2 respectively) were inserted 16 kb centromere distal to MATα-inc between RSC6 and THR4. The ura3–29-HPH cassettes containing ura3–29 allele23 in two orientations were a gift from Youri Pavlov. The insertion of ura3–29-HPH 16 kb centromere distal to MATα-inc was achieved by transformation of AM2110 with DNA fragments generated by PCR amplification of ura3–29-HPH using the following primers with targeting tails (uppercase) and ura3–29-HPH amplification sequence (lower case): 5’TCTTTCTGCAATTATTGCACGCCTCCTCGTGAGTAGTGACCGTGCGAACAAAAGAGTCATTACAACGAGGAAATAGAAGA agtcagtgagcgaggaagc3’ and 5’ATATTTGCTGCTATACTACCAAATGGAAAAATATAAGATACACAATATAGATAGTATTAAAAAAACGTGTATACGTTATT attgtactgagagtgcacc3’ Control strains AM2442, AM2259, and AM2842 which contained no HO cut site in the recipient Chr III, were obtained from AM2118, AM2161, and AM2820 as previously described9.
AM2406 is a derivative of AM1003 that was constructed by inserting BrdU cassette (with the human equilibrative nucleoside transporter (hENT1) and the herpes simplex virus thymidine kinase)14 into URA3 to facilitate efficient BrdU incorporation in yeast. In particular, the p306-BrdU plasmid14 was linearized with StuI and inserted by transformation into the URA3 gene (Chr V). In addition, AM2406 contained insertion of three tandem arrays of the TEF1/BSD-snt1 at SNT1, and replacement of TPS1 with a KANMX module. TPS1 was deleted to reduce accumulation of trehalose, which interfered with DNA purification.
Rich medium (yeast extract-peptone-dextrose [YEPD]) and synthetic complete medium, with bases and amino acids omitted as specified, were made as described32. YEP-raffinose, YEP-lactate and YEP-galactose were made as described9,24. Cultures were grown at 30°C.
Analysis of BIR efficiency
DSBs were initiated by HO induction by addition of galactose2. BIR efficiency was determined genetically and by physical analysis in time-course experiments using PFGE as previously described2. The average efficiency of BIR at each time point was calculated based on results of four independent experiments.
2D analysis of molecular intermediates of BIR
Cells were grown overnight in synthetic leucine drop-out media, transferred to YEP-raffinose, and incubated for ~16 hours, until cell density reached ~1 × 107 cells/ml. An aliquot was taken for analysis of the S-phase replication fork, and 2% galactose was added to induce HO endonuclease in the remainder of the culture. In these experiments, the efficiency of BIR was (80±15)%, as determined by PFGE analysis2 10 hours following DSB (Extended Data Fig. 5c). DSB induction led to G2/M arrest ~3 hours after galactose addition as cells were in the process of completing BIR repair (Extended Data Fig. 5d). At this point, nocodazole was added to the culture to a final concentration of 0.015 mg/ml to maintain the arrest. Cells were collected at different intervals following the break and subjected to psoralen crosslinking that allowed to constrain branch migration during DNA purification as previously described33. Chromosomal DNA was extracted and neutral/neutral 2D analysis was carried out according to34. PstI-digested DNA was separated in the first dimension on a 0.4% gel without ethidium bromide in 1X TBE buffer at 1 V/cm for 22 hours. The second dimension was run at 6 V/cm in 1X TBE buffer containing 0.3 µg/ml ethidium bromide for 12 hours.
Alternatively, to guarantee that the observed intermediates do not result from mechanical stress during genomic DNA preparation, we conducted 2D-gel electrophoresis using chromosomal DNA embedded in agarose plugs. In particular, cells collected at different intervals after induction of BIR were treated with psoralen as described in33. The cells were then resuspended in 750 µl solution of 1 M Sorbitol, 0.5 M EDTA (pH 8) and treated with 0.2 mg/ml lyticase for 1 hour at 37°C. The spheroplasts were washed in a solution of 50 mM Tris, 50 mM EDTA and 100 mM NaCl. The spheroplasts were then embedded in 0.8% low melt agarose at a concentration of 1.5 × 1010 cells/ml. The chromosomal DNA embedded in agarose was digested with BglII, and 2D-gel electrophoresis was carried out as described for the 2D analysis of PstI-digested chromosomal DNA.
To identify regions of single-stranded DNA, a PstI or BglII digest was preceded by pre-incubation of genomic DNA with oligonucleotides that were complimentary to the PstI or BglII sites flanking the LYS2 gene and had the following sequences: GGTCGCCCTGCAGCACAAGC (PstO3), GTCCTTTCCAGATCTTGGCAACTTT (BglO3), GCTTGTGCTGCAGGGCGACC (PstO5), AAAGTTGCCAAGATCTGGAAAGGAC (BglO5), where “O3 and “O5” indicate oligos that are complimentary to the Watson and Crick strands at the centromere-proximal site, respectively; and TAGATGGCTGCAGAACCAGT (PstO4), TGGATCTGGTAGATCTGTAAACTTGG (BglO4), ACTGGTTCTGCAGCCATCTA (PstO6), CCAAGTTTACAGATCTACCAGATCCA (BglO6), where “O4” and “O6” indicate oligos that are complimentary to the Watson and Crick strands at the telomere-proximal site, respectively.
Southern hybridization was performed using LYS2 fragment obtained by PCR amplification of a 0.6-kb region of LYS2 (from 471835 to 472443 kb positions of chromosome II ) or using HPH-hybridizing fragment obtained by PCR amplification of HPH from the pAG32 plasmid 35.
Along with analysis of BIR intermediates, cell cycle distribution was analyzed by flow cytometry4 and BIR kinetics were analyzed by PFGE. For PFGE, chromosome plugs were prepared4 with genomic DNA embedded in plugs of 1% low-melting agarose and separated at 6 V/cm for 40 hours using the CHEF DRII apparatus. PFGE was followed by Southern analysis with an ADE1-specific probe labeled with P32. Images were analyzed using a Molecular Dynamics PhosphorImager.
DNA Combing and Fluorescent In Situ Hybridization
Cells were grown overnight in synthetic leucine drop-out media , transferred to YEP-lactate, and incubated for ~20 hours, until cell density reached ~1 × 107 cells/ml. Cells were arrested by nocodazole added to 0.015 mg/ml, and DSBs were induced 2.5 hours later by addition of galactose to the final concentration of 2%. When experiments were performed according to this protocol, the efficiency of BIR was (54.0±9.8)%, as determined by PFGE analysis2 11 hours following DSB induction (Extended Data Fig. 1a, b). BrdU was added to the culture 3.5 hours following DSB induction by galactose to the final concentration of 0.4mg/ml after all normal DNA replication was completed but prior to the beginning of BIR. Aliquots were removed to embed cells into agarose plugs prior to and 11 hours after induction of DSBs with galactose. In experiments involving pif1Δ strains, the analysis was performed 13 hours following DSB induction due to slower kinetics of DSB repair in pif1Δ (data not shown). The uniform arrest of cells at G2/M was confirmed by the absence of BrdU incorporation in any chromosomes other than chromosome III, which was assayed by PFGE analysis of yeast chromosomes extracted from samples taken prior to the addition of BrdU and 11 or 13 h following DSB induction and probing with anti-BrdU antibodies.
Genomic DNA preparation and molecular combing were performed as described36. Color hybridization of Chr III molecules was performed using three fluorescent probes. P1 probe was prepared using the TEF1/BSD plasmid (Invitrogen) and hybridized to the 15 kb region containing three tandem repeats of the TEF1/BSD-snt1 plasmid inserted into the donor copy of Chr III at position 186535. P2 probe marked the position close to strand invasion during BIR and was comprised of a set of four 5 kb fragments that corresponded to the following positions on the donor Chr III: 200205 to 205140, 205117 to 210385, 210361 to 215385, and 215361 to 220337. The P3 probe highlights the region close to the telomeric end of Chr III and is made up of three 5-kb fragments corresponding to the following positions on the donor chromosome: 274778 to 279801, 279778 to 284814 and 284791 to 289782. The probes were made by PCR amplification of genomic DNA from AM2406. Nucleotide sequences of the primers used to generate fragments for labeling are available upon request. Probes were labeled with biotin-dUTP. Hybridization and fluorescent detection of combed DNA molecules were achieved according to protocols described36 with a few modifications. Successive layers of fluorophore-conjugated antibodies diluted in 1× PBST (1× PBS + 0.05% Tween) were used. For the biotin-conjugated probes, the following series was used at a dilution of 1:4000: 1) Alexa-488-Streptavidin (Molecular Probes; Life Technologies, Cat. #32354) 2) biotinylated antistreptavidin (From Vector Lab, Cat. # BA-0500), 3) Alexa-488-streptavidin, 4) biotinylated anti-streptavidin and 5) Alexa-488-Streptavidin. To detect BrdU incorporation, the following series were used at the indicated dilutions: 1) 1:20 dilution of mouse anti-BrdU (BD Biosciences, Cat. #347580), 2) 1:50 dilution of Cy3-coupled rat anti-mouse (Jackson ImmunoResearch Lab, PA; Cat. # 415-165-166) and (3) 1:50 dilution of Cy3-mouse anti-rat (Jackson ImmunoResearch Lab, PA; Cat. #212-165-168). All images were acquired using the Zeiss LSM 510 Confocal Microscope with 100× objective. The lengths of the fluorescent stretches were calculated by comparison with the length of P1, P2 and P3 hybridization signals.
The statistical comparison between donor and recipient chromosomes in respect to BrdU incorporation was performed using the Chi-square test. For each experiment, the frequency of semiconservative BIR (F) was calculated as follows: F = A/N × f × b, where A represents the number of donor molecules with long BrdU tracts; N represents the total number of analyzed donor molecules; f represents the efficiency of BIR in the experiment (calculated by physical analysis as a % of the truncated chromosome III converted in the BIR product4) ; and b represents the fraction of recipient molecules containing full and long interrupted BIR tracts.
Mutagenesis Associated with DSB Repair
To determine mutation frequency associated with BIR, yeast strains were grown from individual colonies with agitation in liquid synthetic media lacking leucine for approximately 20 h, diluted 20-fold with fresh YEP-Lac, and grown to logarithmic phase for approximately 16 h. Next, 20% galactose was added to the culture to a final concentration of 2%, and cells were incubated with agitation for 7 h. Samples from each culture were plated at appropriate concentrations on adenine drop-out media and on media omitting lysine and adenine before (0 h) and 7 h after the addition of galactose (7 h) to measure the frequency of Lys+ cells. To measure the frequency of Ura+ cells, samples were plated at appropriate concentrations on adenine drop-out media and on media omitting uracil and adenine before (0 h) and 7 h after the addition of galactose (7 h). To determine spontaneous mutation frequencies, no-DSB strains were grown similarly to the DSB-containing strains. Because spontaneous mutation frequencies were calculated based on the number of mutations accumulated during many cell generations, the rate of spontaneous mutagenesis in no-DSB control strains was calculated using the following modification of Drake equation: μ=0.4343 f/log(Nμ), where μ= the rate of spontaneous mutagenesis, f= mutation frequency, and N= the number of cells in yeast culture. The rate of mutations after galactose treatment (μ7) was determined using a simplified version of the Drake equation: μ7= (f7 − f0), where f7 and f0 are the mutation frequencies among Ade+ cells at times 7 h and 0 h, respectively. This modification was necessary because experimental strains did not divide or underwent ≤ 1 division between 0h and 7h.
MMS was added at 1.5 mM 30 minutes after galactose addition. Cells were incubated with agitation for 7 hours, treated with 10% sodium thiosulfate to inactivate MMS, diluted and plated. The loss of viability following MMS treatment was barely detectable and never exceeded 40% independently of ura3–29 orientation. The rate of mutations following MMS treatment was determined using simplified version of the Drake equation: μ7= (f7 − f0), where f7 and f0 are the mutation frequencies among Ade+ cells at times 7 h (following MMS treatment) and 0 h, respectively. This modification was necessary because experimental strains did not divide or underwent ≤ 1 division between 0 h and 7 h in the presence of MMS.
Rates are reported as the median value (Fig. 2b,e; Extended Data Tables 1 and 2), and the 95% confidence limits for the median are calculated for the strains with a minimum of six individual experiments. For strains with 4–5 individual experiments, the range of the median was calculated. Statistical comparisons between median mutation rates were performed using the Mann-Whitney U test 37.
Analysis of BIR-induced Lys+ mutants
Lys+ revertants were obtained in BIR mutagenesis experiments9. After phenotypic examination, cultures were grown from mutants for chromosome analysis by PFGE using 1% low-melting agarose at 6 V/cm for 48 hours. DNA bands corresponding to the donor and repaired recipient Chr III were excised, equilibrated in β-agarase buffer (NEB), melted at 65°C, and subjected to β-agarase treatment for 1 hour at 40°C. The obtained DNA was PCR amplified using LYS2-specific DNA primers9, followed by sequencing analysis.
Analysis of mutation spectra of ura3–29 Ura+ reversions
To determine the spectrum of Ura+ in individual experiments, a portion of the URA3 gene from independent Ura+ was PCR amplified using URA3-specific primers: 5’GTGTGCTTCATTGGATGTTCGTAC3’ and 5’AAAAGGCCTCTAGGTTCCTTTGTT3’ followed by sequencing analysis using 5’CTGGAGTTAGTTGAAGCATTAGG3’ as a primer.
For experimental strains undergoing BIR repair, 7h Ura+ BIR events (confirmed as Ade+Leu− on selective media) were sequenced. Because these cells underwent ≤1 divisions between the 0h and 7h timepoints and the Ura+ frequency at 7h significantly exceeded that at 0h, all Ura+ events resulting from DSB repair were considered independent.
Supplementary Material
a, BIR efficiency was analyzed by PFGE from samples used for dynamic molecular combing analysis (Fig. 3d). DNA was prepared from cells containing truncated chromosome III (Trunc Chr III) before DSB induction and 11 hours or 13 hours following DSB induction from wt (PIF1) and pif1Δ cells respectively. In pif1Δ, a later time point (13 hours) was analyzed due to slower kinetics of DSB repair in pif1Δ as compared to PIF1. Chromosomes were separated by PFGE followed by Southern hybridization with an ADE1-specific probe. b, Quantification of DSB repair efficiency (BIR, or other recombination pathways) based on the results of 3-5 individual experiments. c, Schematic of the BIR assay. Interruption of BIR leads to the resolution of BIR intermediates resulting in half-crossover formation.
{kind=link}
BIR intermediates were analyzed by 2D gel electrophoresis of BglII-digested intact chromosomal DNA embedded in agarose plugs. a, D-loop migration in 2D gels (hybridized to LYS2, black rectangle) during coordinated (i) and uncoordinated (ii, iii) leading- and lagging-strand synthesis. b, Schematic of 2D gel separation of replication and BIR intermediates. Annealing to oligonucleotides (BglO3 and BglO4) restores BglII sites (B) in ssDNA (see a(ii)) and changes migration of the intermediate as shown by 2’ (red). c, 2D analysis of Y-arc during normal replication (0 Hr) and bubble-like structures at time points following BIR induction. d, High molecular-weight tails (arrows) disappear following simultaneous addition of BglO3 and BglO4 (BIR/BglO3+BglO4). The addition of each of these oligos individually (BIR/BglO3 or BIR/BglO4) failed to eliminate the tail.
{kind=link}
aRates calculated at 0h based on 0h frequencies using the Drake equation (see Methods for details). At 7h, rates were calculated as (7h frequency − 0h frequency); differences <0 are reported as "0". bFor strains with ≥6 experiments, the 95% CI of the median is given. cStatistically significant elevation of 7h mutation rate in strains in the presence of MMS over 7h mutation rate in the absence of MMS. dPercent of BIR (average ± STD) calculated based on 3–6 experiments among DSB repair outcomes collected at 7h on either adenine dropout media (Ade+) or on adenine/uracil dropout media (Ade+Ura+).
{kind=link}
{kind=link}
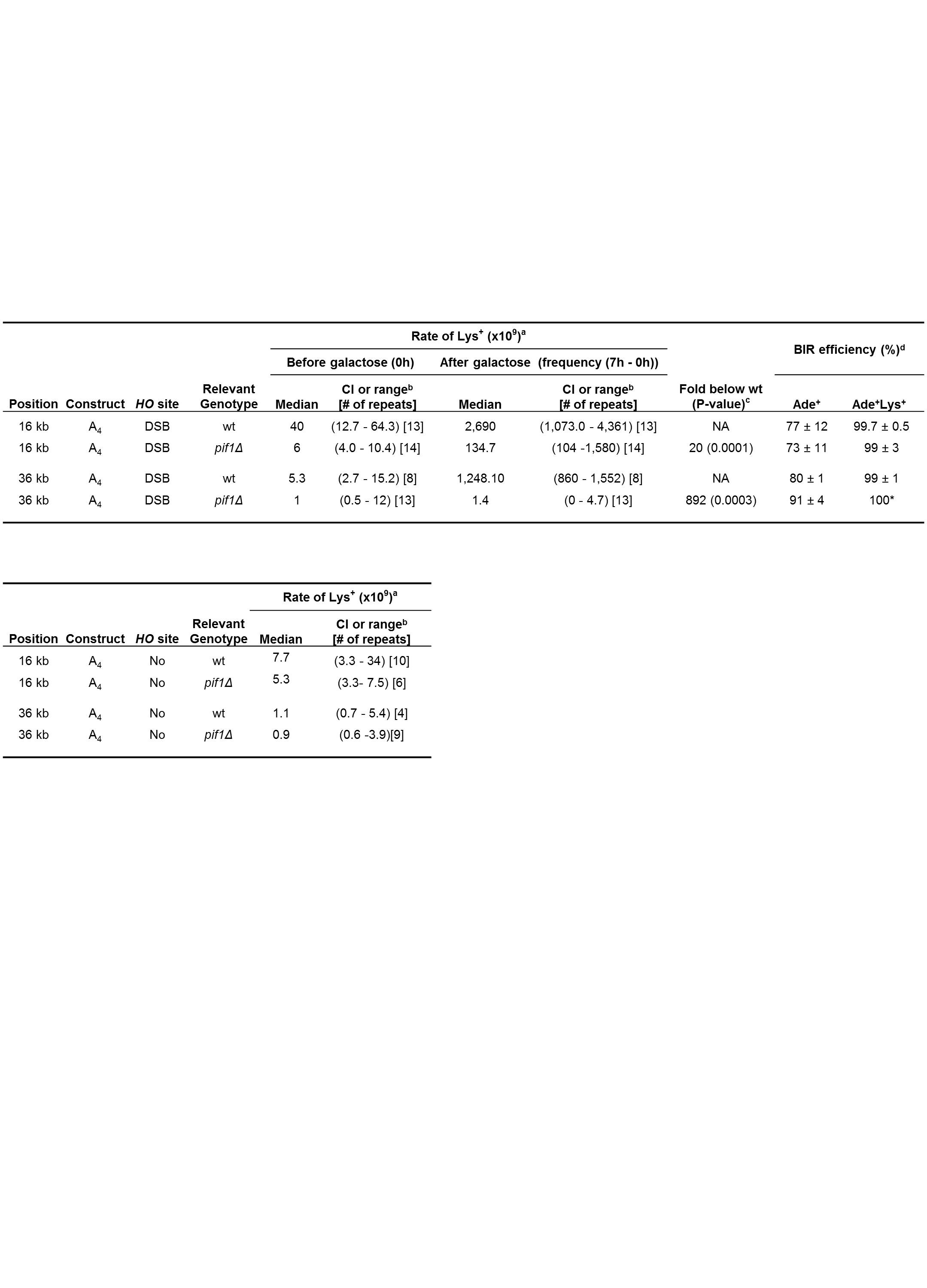
aRates calculated at 0h based on 0h frequencies using the Drake equation (see Methods for details). At 7h, rates were calculated as (7h frequency − 0h frequency); differences <0 are reported as "0". bFor strains with ≥6 experiments, the 95% CI of the median is given. For the strains with <6 experiments, the median range is given. cStatistically significant decrease of median rate at 7h in pif1Δ compared to wt. dPercent of BIR (average ± STD) calculated based on 4–8 experiments among DSB repair outcomes collected at 7h on either adenine dropout media (Ade+) or on adenine/lysine dropout media (Ade+Lys+). * No STD could be calculated because of a very low number of Lys+ (between 1 and 5) in each experiment.
{kind=link}
a, The summary of molecular combing analysis presented in Fig. 3 and in Extended Data Figure 3 is shown. b, Mutation spectra of BIR-induced base substitutions in ura3–29 in the presence or absence of 1.5 mM MMS is shown.
{kind=link}
a, Left: Interrupted BrdU tract in recipient may result from half-crossover. Right: an example of wt (PIF1) recipient with interrupted BrdU tract hybridized to P1, P2, P3 probes (green) and treated with anti-BrdU antibody (red). b, Left: BIR initiated by strand invasion between FS2 (inverted repeat of Ty1 located 30 kb centromere proximal to MAT) and P1 results in formation of recipients hybridizing to P1, P2, P3 and BrdU. Right: an example of wt (PIF1) recipient. Top: hybridization to P1, P2, P3; middle: treatment with anti-BrdU antibody; bottom: merge. c, Left: BrdU incorporation in the recipient resulting from BIR (red) and from filling-in synthesis (pink) following extensive resection. Right: an example of wt (PIF1) recipient. Top: hybridization to P1, P2, P3; middle: treatment with anti-BrdU antibody; bottom: merge. d, Left: HJ resolution at the end of BIR progression leads to switch from conservative to semiconservative BIR resulting in a short patch of BrdU overlapping with P3 in the donor. Right: an example of BrdU incorporation in the donor from wt (PIF1) strain hybridized to P1, P2, P3 and treated with anti-BrdU antibody.
{kind=link}
Results from a series of experiments where only P1, P2 and anti-BrdU antibody were used. a, BrdU incorporation in the recipient is expected from conservative BIR (i; red) and from filling-in synthesis (pink) following extensive resection (ii). Analysis of the donor (D) and repaired recipient (R) chromosomes extracted following PFGE (b) and hybridization with probes (green tract) and treatment with anti-BrdU antibodies (red tract) (c). No BrdU tracts are visible in more than 97% of donors. The repaired recipient contains long stretches of BrdU overlapping with the P2 region.
{kind=link}
a, BIR kinetics was analyzed by PFGE from samples used to determine the structure of BIR intermediates by 2D electrophoresis (Fig. 4c, d). DNA was prepared for PFGE at intervals after induction of DSBs at MATa and separated by PFGE (a) followed by Southern hybridization with an ADE1-specific probe (b). c, Quantification of BIR efficiency based on the results of four individual experiments including the one shown in Fig. 4. d, Flow cytometery of DNA analysis of cells undergoing BIR repair.
{kind=link}
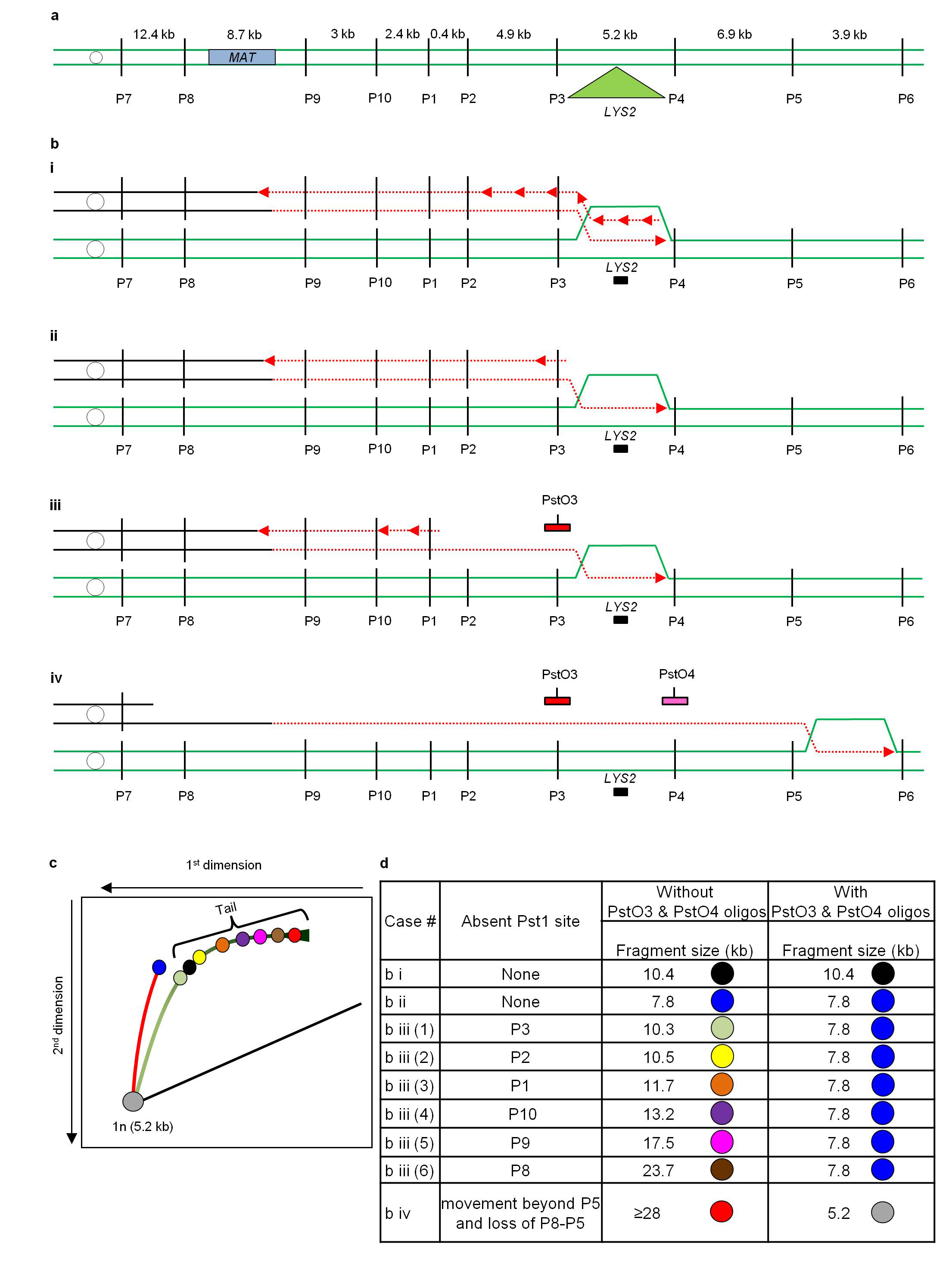
a, The structure of the Chr III region with LYS2 inserted 16 kb centromere distal to MATα-inc. P1, P2, P3, etc. designate positions of PstI sites flanking LYS2. b, The structure of replication bubbles migrating through LYS2 (with black rectangle designating LYS2-specific probe). (i), Replication bubble with synchronous leading and lagging strands (double-stranded). (ii), Replication bubble with delayed initiation of the lagging strand with respect to the leading strand (partially single-stranded bubble). (iii), A partially single-stranded bubble with one or several PstI sites behind the bubble inactivated due to accumulation of single-stranded DNA. Red and pink rectangles represent oligonucleotides PstO3 and PstO4 respectively. (iv), A single-stranded bubble that has passed beyond the P3-P4 region. c, Theoretical bubble-migration curves for the intermediates shown in b. d, Calculation of parameters of the bubble-like structures for the intermediates shown in b.
{kind=link}
Acknowledgements
We thank Oscar Aparicio for the gift of BrdU cassette plasmid. We thank Youri Pavlov for providing ura3–29 cassette plasmid and Paul Chastain and Anni Hangaard Andersen for providing us with reagents. We are thankful to Dmitry Gordenin and all members of AM, KL, GI, and JH laboratories for fruitful discussions throughout the work. This work was funded by grants from the US National Institutes of Health and National Science Foundation (MCB-0818122 from NSF to KL, and NIH grants R01GM084242 to AM, R01GM082950 to KL, R03ES016434 to AM and KL, GM76020 to JEH, and GM080600 to GI).
Footnotes
Supplementary Information is available in the online version of the paper.
Author Contributions
SR, NS, KL and AM designed experiments. SR and AD constructed the experimental system. NS, SR, and RE carried out 2D experiments. KL, SR, AM, NS, YZ carried out molecular combing experiments. SA, RE and AD carried out experiments involving sequencing of BIR-induced mutations. RE and SA carried out experiments aimed to determine the effect of BIR on base substitutions. JEH provided key expertise. GI contributed to the studies of the role of Pif1 in BIR. NS, SR, AD, JEH, KL, and AM wrote the paper. NS and SR contributed equally to this work.
The authors declare no competing financial interests.
References
- 1.Smith CE, Llorente B, Symington LS. Template switching during break-induced replication. Nature. 2007;447:102–105. doi: 10.1038/nature05723. [DOI] [PubMed] [Google Scholar]
- 2.Deem A, et al. Defective break-induced replication leads to half-crossovers in Saccharomyces cerevisiae. Genetics. 2008;179:1845–1860. doi: 10.1534/genetics.108.087940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Malkova A, Haber JE. Mutations arising during repair of chromosome breaks. Annual review of genetics. 2012;46:455–473. doi: 10.1146/annurev-genet-110711-155547. [DOI] [PubMed] [Google Scholar]
- 4.Malkova A, Naylor ML, Yamaguchi M, Ira G, Haber JE. RAD51-dependent break-induced replication differs in kinetics and checkpoint responses from RAD51-mediated gene conversion. Molecular and cellular biology. 2005;25:933–944. doi: 10.1128/MCB.25.3.933-944.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Davis AP, Symington LS. RAD51-dependent break-induced replication in yeast. Molecular and cellular biology. 2004;24:2344–2351. doi: 10.1128/MCB.24.6.2344-2351.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bosco G, Haber JE. Chromosome break-induced DNA replication leads to nonreciprocal translocations and telomere capture. Genetics. 1998;150:1037–1047. doi: 10.1093/genetics/150.3.1037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hastings PJ, Ira G, Lupski JR. A microhomology-mediated break-induced replication model for the origin of human copy number variation. PLoS Genet. 2009;5:e1000327. doi: 10.1371/journal.pgen.1000327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Payen C, Koszul R, Dujon B, Fischer G. Segmental duplications arise from Pol32-dependent repair of broken forks through two alternative replication-based mechanisms. PLoS Genet. 2008;4:e1000175. doi: 10.1371/journal.pgen.1000175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Deem A, et al. Break-induced replication is highly inaccurate. PLoS Biol. 2011;9:e1000594. doi: 10.1371/journal.pbio.1000594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lydeard JR, Jain S, Yamaguchi M, Haber JE. Break-induced replication and telomerase-independent telomere maintenance require Pol32. Nature. 2007;448:820–823. doi: 10.1038/nature06047. [DOI] [PubMed] [Google Scholar]
- 11.Lydeard JR, et al. Break-induced replication requires all essential DNA replication factors except those specific for pre-RC assembly. Genes & development. 2010;24:1133–1144. doi: 10.1101/gad.1922610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Llorente B, Smith CE, Symington LS. Break-induced replication: what is it and what is it for? Cell Cycle. 2008;7:859–864. doi: 10.4161/cc.7.7.5613. [DOI] [PubMed] [Google Scholar]
- 13.Malkova A, Ira G. Break-induced replication: functions and molecular mechanism. Current opinion in genetics & development. 2013 doi: 10.1016/j.gde.2013.05.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Viggiani CJ, Aparicio OM. New vectors for simplified construction of BrdU-Incorporating strains of Saccharomyces cerevisiae. Yeast. 2006;23:1045–1051. doi: 10.1002/yea.1406. [DOI] [PubMed] [Google Scholar]
- 15.Smith CE, Lam AF, Symington LS. Aberrant double-strand break repair resulting in half crossovers in mutants defective for Rad51 or the DNA polymerase delta complex. Molecular and cellular biology. 2009;29:1432–1441. doi: 10.1128/MCB.01469-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Vanhulle K, et al. Inverted DNA repeats channel repair of distant double-strand breaks into chromatid fusions and chromosomal rearrangements. Molecular and cellular biology. 2007;27:2601–2614. doi: 10.1128/MCB.01740-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Fangman WL, Brewer BJ. Activation of replication origins within yeast chromosomes. Annu Rev Cell Biol. 1991;7:375–402. doi: 10.1146/annurev.cb.07.110191.002111. [DOI] [PubMed] [Google Scholar]
- 18.Formosa T, Alberts BM. DNA synthesis dependent on genetic recombination: characterization of a reaction catalyzed by purified bacteriophage T4 proteins. Cell. 1986;47:793–806. doi: 10.1016/0092-8674(86)90522-2. [DOI] [PubMed] [Google Scholar]
- 19.Ferguson DO, Holloman WK. Recombinational repair of gaps in DNA is asymmetric in Ustilago maydis and can be explained by a migrating D-loop model. Proceedings of the National Academy of Sciences of the United States of America. 1996;93:5419–5424. doi: 10.1073/pnas.93.11.5419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Yang Y, Sterling J, Storici F, Resnick MA, Gordenin DA. Hypermutability of damaged single-strand DNA formed at double-strand breaks and uncapped telomeres in yeast Saccharomyces cerevisiae. PLoS Genet. 2008;4:e1000264. doi: 10.1371/journal.pgen.1000264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Yang Y, Gordenin DA, Resnick MA. A single-strand specific lesion drives MMS-induced hyper-mutability at a double-strand break in yeast. DNA repair. 2010;9:914–921. doi: 10.1016/j.dnarep.2010.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Roberts SA, et al. Clustered mutations in yeast and in human cancers can arise from damaged long single-strand DNA regions. Molecular cell. 2012;46:424–435. doi: 10.1016/j.molcel.2012.03.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Shcherbakova PV, Pavlov YI. 3'-->5' exonucleases of DNA polymerases epsilon and delta correct base analog induced DNA replication errors on opposite DNA strands in Saccharomyces cerevisiae. Genetics. 1996;142:717–726. doi: 10.1093/genetics/142.3.717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Chung WH, Zhu Z, Papusha A, Malkova A, Ira G. Defective resection at DNA double-strand breaks leads to de novo telomere formation and enhances gene targeting. PLoS Genet. 2010;6:e1000948. doi: 10.1371/journal.pgen.1000948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Hashimoto Y, Costanzo V. Studying DNA replication fork stability in Xenopus egg extract. Methods Mol Biol. 2011;745:437–445. doi: 10.1007/978-1-61779-129-1_25. [DOI] [PubMed] [Google Scholar]
- 26.Pohjoismaki JL, Goffart S. Of circles, forks and humanity: Topological organisation and replication of mammalian mitochondrial DNA. BioEssays : news and reviews in molecular, cellular and developmental biology. 2011;33:290–299. doi: 10.1002/bies.201000137. [DOI] [PubMed] [Google Scholar]
- 27.Le S, Moore JK, Haber JE, Greider CW. RAD50 and RAD51 define two pathways that collaborate to maintain telomeres in the absence of telomerase. Genetics. 1999;152:143–152. doi: 10.1093/genetics/152.1.143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kreuzer KN, Saunders M, Weislo LJ, Kreuzer HW. Recombination-dependent DNA replication stimulated by double-strand breaks in bacteriophage T4. Journal of bacteriology. 1995;177:6844–6853. doi: 10.1128/jb.177.23.6844-6853.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kuzminov A. Collapse and repair of replication forks in Escherichia coli. Mol Microbiol. 1995;16:373–384. doi: 10.1111/j.1365-2958.1995.tb02403.x. [DOI] [PubMed] [Google Scholar]
- 30.Halazonetis TD, Gorgoulis VG, Bartek J. An oncogene-induced DNA damage model for cancer development. Science. 2008;319:1352–1355. doi: 10.1126/science.1140735. [DOI] [PubMed] [Google Scholar]
- 31.Wach A, Brachat A, Pohlmann R, Philippsen P. New heterologous modules for classical or PCR-based gene disruptions in Saccharomyces cerevisiae. Yeast. 1994;10:1793–1808. doi: 10.1002/yea.320101310. [DOI] [PubMed] [Google Scholar]
- 32.Guthrie C, Fink G. Guide to Yeast Genetics and Molecular Biology. San Diego: Academic Press; 1991. [Google Scholar]
- 33.Oh SD, et al. Stabilization and electrophoretic analysis of meiotic recombination intermediates in Saccharomyces cerevisiae. Methods Mol Biol. 2009;557:209–234. doi: 10.1007/978-1-59745-527-5_14. [DOI] [PubMed] [Google Scholar]
- 34.Friedman KL, Brewer BJ. Analysis of replication intermediates by two-dimensional agarose gel electrophoresis. Methods in enzymology. 1995;262:613–627. doi: 10.1016/0076-6879(95)62048-6. [DOI] [PubMed] [Google Scholar]
- 35.Goldstein AL, McCusker JH. Three new dominant drug resistance cassettes for gene disruption in Saccharomyces cerevisiae. Yeast. 1999;15:1541–1553. doi: 10.1002/(SICI)1097-0061(199910)15:14<1541::AID-YEA476>3.0.CO;2-K. [DOI] [PubMed] [Google Scholar]
- 36.Conti C, Caburet S, Schurra C, Bensimon A. Molecular combing. Curr Protoc Cytom. 2001;Chapter 8(Unit 8 10) doi: 10.1002/0471142956.cy0810s16. [DOI] [PubMed] [Google Scholar]
- 37.Mann H, Whitney D. On a test of whether one of 2 random variables is stochastically larger than the other. Annals of Mathematical Statistics. 1947;18:50–60. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
a, BIR efficiency was analyzed by PFGE from samples used for dynamic molecular combing analysis (Fig. 3d). DNA was prepared from cells containing truncated chromosome III (Trunc Chr III) before DSB induction and 11 hours or 13 hours following DSB induction from wt (PIF1) and pif1Δ cells respectively. In pif1Δ, a later time point (13 hours) was analyzed due to slower kinetics of DSB repair in pif1Δ as compared to PIF1. Chromosomes were separated by PFGE followed by Southern hybridization with an ADE1-specific probe. b, Quantification of DSB repair efficiency (BIR, or other recombination pathways) based on the results of 3-5 individual experiments. c, Schematic of the BIR assay. Interruption of BIR leads to the resolution of BIR intermediates resulting in half-crossover formation.
BIR intermediates were analyzed by 2D gel electrophoresis of BglII-digested intact chromosomal DNA embedded in agarose plugs. a, D-loop migration in 2D gels (hybridized to LYS2, black rectangle) during coordinated (i) and uncoordinated (ii, iii) leading- and lagging-strand synthesis. b, Schematic of 2D gel separation of replication and BIR intermediates. Annealing to oligonucleotides (BglO3 and BglO4) restores BglII sites (B) in ssDNA (see a(ii)) and changes migration of the intermediate as shown by 2’ (red). c, 2D analysis of Y-arc during normal replication (0 Hr) and bubble-like structures at time points following BIR induction. d, High molecular-weight tails (arrows) disappear following simultaneous addition of BglO3 and BglO4 (BIR/BglO3+BglO4). The addition of each of these oligos individually (BIR/BglO3 or BIR/BglO4) failed to eliminate the tail.
aRates calculated at 0h based on 0h frequencies using the Drake equation (see Methods for details). At 7h, rates were calculated as (7h frequency − 0h frequency); differences <0 are reported as "0". bFor strains with ≥6 experiments, the 95% CI of the median is given. cStatistically significant elevation of 7h mutation rate in strains in the presence of MMS over 7h mutation rate in the absence of MMS. dPercent of BIR (average ± STD) calculated based on 3–6 experiments among DSB repair outcomes collected at 7h on either adenine dropout media (Ade+) or on adenine/uracil dropout media (Ade+Ura+).
aRates calculated at 0h based on 0h frequencies using the Drake equation (see Methods for details). At 7h, rates were calculated as (7h frequency − 0h frequency); differences <0 are reported as "0". bFor strains with ≥6 experiments, the 95% CI of the median is given. For the strains with <6 experiments, the median range is given. cStatistically significant decrease of median rate at 7h in pif1Δ compared to wt. dPercent of BIR (average ± STD) calculated based on 4–8 experiments among DSB repair outcomes collected at 7h on either adenine dropout media (Ade+) or on adenine/lysine dropout media (Ade+Lys+). * No STD could be calculated because of a very low number of Lys+ (between 1 and 5) in each experiment.
a, The summary of molecular combing analysis presented in Fig. 3 and in Extended Data Figure 3 is shown. b, Mutation spectra of BIR-induced base substitutions in ura3–29 in the presence or absence of 1.5 mM MMS is shown.
a, Left: Interrupted BrdU tract in recipient may result from half-crossover. Right: an example of wt (PIF1) recipient with interrupted BrdU tract hybridized to P1, P2, P3 probes (green) and treated with anti-BrdU antibody (red). b, Left: BIR initiated by strand invasion between FS2 (inverted repeat of Ty1 located 30 kb centromere proximal to MAT) and P1 results in formation of recipients hybridizing to P1, P2, P3 and BrdU. Right: an example of wt (PIF1) recipient. Top: hybridization to P1, P2, P3; middle: treatment with anti-BrdU antibody; bottom: merge. c, Left: BrdU incorporation in the recipient resulting from BIR (red) and from filling-in synthesis (pink) following extensive resection. Right: an example of wt (PIF1) recipient. Top: hybridization to P1, P2, P3; middle: treatment with anti-BrdU antibody; bottom: merge. d, Left: HJ resolution at the end of BIR progression leads to switch from conservative to semiconservative BIR resulting in a short patch of BrdU overlapping with P3 in the donor. Right: an example of BrdU incorporation in the donor from wt (PIF1) strain hybridized to P1, P2, P3 and treated with anti-BrdU antibody.
Results from a series of experiments where only P1, P2 and anti-BrdU antibody were used. a, BrdU incorporation in the recipient is expected from conservative BIR (i; red) and from filling-in synthesis (pink) following extensive resection (ii). Analysis of the donor (D) and repaired recipient (R) chromosomes extracted following PFGE (b) and hybridization with probes (green tract) and treatment with anti-BrdU antibodies (red tract) (c). No BrdU tracts are visible in more than 97% of donors. The repaired recipient contains long stretches of BrdU overlapping with the P2 region.
a, BIR kinetics was analyzed by PFGE from samples used to determine the structure of BIR intermediates by 2D electrophoresis (Fig. 4c, d). DNA was prepared for PFGE at intervals after induction of DSBs at MATa and separated by PFGE (a) followed by Southern hybridization with an ADE1-specific probe (b). c, Quantification of BIR efficiency based on the results of four individual experiments including the one shown in Fig. 4. d, Flow cytometery of DNA analysis of cells undergoing BIR repair.
a, The structure of the Chr III region with LYS2 inserted 16 kb centromere distal to MATα-inc. P1, P2, P3, etc. designate positions of PstI sites flanking LYS2. b, The structure of replication bubbles migrating through LYS2 (with black rectangle designating LYS2-specific probe). (i), Replication bubble with synchronous leading and lagging strands (double-stranded). (ii), Replication bubble with delayed initiation of the lagging strand with respect to the leading strand (partially single-stranded bubble). (iii), A partially single-stranded bubble with one or several PstI sites behind the bubble inactivated due to accumulation of single-stranded DNA. Red and pink rectangles represent oligonucleotides PstO3 and PstO4 respectively. (iv), A single-stranded bubble that has passed beyond the P3-P4 region. c, Theoretical bubble-migration curves for the intermediates shown in b. d, Calculation of parameters of the bubble-like structures for the intermediates shown in b.