

Abstract
Aquaporins are channel proteins which transport water across cell membranes. We show that the bread wheat aquaporin gene TaTIP2;2 maps to the long arm of chromosome 7b and that its product localizes to the endomembrane system. The gene is expressed constitutively in both the root and the leaf, and is down-regulated by salinity and drought stress. Salinity stress induced an increased level of C-methylation within the CNG trinucleotides in the TaTIP2;2 promoter region. The heterologous expression of TaTIP2;2 in Arabidopsis thaliana compromised its drought and salinity tolerance, suggesting that TaTIP2;2 may be a negative regulator of abiotic stress. The proline content of transgenic A. thaliana plants fell, consistent with the down-regulation of P5CS1, while the expression of SOS1, SOS2, SOS3, CBF3 and DREB2A, which are all stress tolerance-related genes acting in an ABA-independent fashion, was also down-regulated. The supply of exogenous ABA had little effect either on TaTIP2;2 expression in wheat or on the phenotype of transgenic A. thaliana. The expression level of the ABA signalling genes ABI1, ABI2 and ABF3 remained unaltered in the transgenic A. thaliana plants. Thus TaTIP2;2 probably regulates the response to stress via an ABA-independent pathway(s).
Introduction
A range of abiotic stresses, including soil salinity, drought and extreme temperature, can compromise crop yield and quality. Improving tolerance to these stresses is thus a major priority in many crop breeding programmes. The effectiveness of water transporters, such as the aquaporins, is an important component of the plant response to stress [1]. The aquaporins belong to a highly conserved major intrinsic protein family, and combine with the cell membrane system to control the flow of water between and within the cell. Their structure is characterized by the formation of six transmembrane domains connected by five loops [2–4]. Based on their sub-cellular localization and sequence, plant aquaporins have been classified into four sub-families, namely the plasma membrane intrinsic proteins (PIPs), the tonoplast intrinsic proteins (TIPs), the nodulin 26-like intrinsic proteins (NIPs) and the small basic intrinsic proteins [5]. There are also three new subclasses of aquaporins in moss Physcomitrella patens, named GlpF-like intrinsic protein (GIP), hybrid intrinsic protein (HIP) and X intrinsic protein (XIP) [6].
The Panax ginseng aquaporin gene PgTIP1 [7], tomato (Solanum lycopersicum) SlTIP2;2 [8] and cotton (Gossypium hirsutum) GhPIP2;7 [9] have been shown to positively regulate salinity and drought tolerance, but negative effects on stress tolerance of certain aquaporins have also been documented [10]. The stress inducibility of aquaporin genes is variable, with examples including both their up- [11–15] and down-regulation [13,14,16,17] as well as their insensitivity [13–15]. Among the wheat aquaporins, it is known that certain TIPs show a marked affinity with ammonia [18], while the activity of particular NIP [19] and PIP genes [20–22] have been associated with an improved response to abiotic stress. Of a set of 35 wheat aquaporin genes, 24 were shown to be PIPs and 11 were TIPs that have diverse sequence characteristics [23]. However, as yet there is an insufficient understanding of the role of the aquaporins in the abiotic stress response of wheat.
The bread wheat cultivar Shanrong No. 3 (SR3) is a derivative of an asymmetric somatic hybrid between cv. Jinan 177 (JN177) and an accession of tall wheatgrass (Thinopyrum ponticum) [24]. SR3 has proven to show an enhanced level of both salinity and drought tolerance over JN177 [25]. A microarray-based gene expression study has shown that in drought and salinity stressed SR3 plants, TaTIP2 was down-regulated [26]. Here, we report the isolation of the TaTIP2;2 SR3 allele. We have determined its chromosomal location and confirmed that its expression is suppressed by both drought and salinity stress. We show that it encodes a protein deposited in the endomembrane, and that its heterologous expression in A. thaliana compromised the level of tolerance to salinity and drought stress. Finally, we demonstrate that the gene is involved in the down-regulation of proline synthesis and acts in an ABA-independent manner.
Materials and Methods
Plant materials and growing conditions
SR3 seedlings were raised in half-strength Hoagland's liquid medium [27] at 22°C under a 16h photoperiod with the light intensity of 3,000 lux. At the three leaf stage, a portion of the seedlings was exposed to abiotic stress by the addition to the medium of either 150mM NaCl, 18% w/v PEG or 100μM ABA. After 0h, 0.5h or 48h of this treatment, RNA was extracted from both the leaf and the root using an RNAiso plus kit (Takara, Dalian, China), following the protocol recommended by the manufacturer. A. thaliana plants were cultured on either half strength MS medium or soil under the same environmental conditions as the wheat seedlings.
Isolation of TaTIP2;2
The TaTIP2;2 cDNA sequence (GenBank accession number AY525640) was used to design a primer pair (TaTIP2, sequences given in Table 1) able to amplify the gene's open reading frame from a template of cDNA prepared from salinity stressed SR3 seedlings. For cloning purposes, an XbaI restriction site was included in the forward primer and a SacI site in the reverse primer. The PCR comprised an initial denaturation of 95°C/3min, followed by 35 cycles of 94°C/40s, 65°C/50s, 72°C/60s, ending with a final extension of 72°C/10min. The amplicons were gel-purified, digested with XbaI and SacI and ligated with XbaI/SacI digested pSTART [28]. The resulting construct was transferred into Agrobacterium tumefaciens strain EHA105 for the agroinfection of A. thaliana Col-0 via the floral dip method [29]. Homozygous transgenic segregants in the T3 generation were used for phenotypic and gene expression analysis. The same primer pair was used to recover the SR3 TaTIP2;2 genomic sequence, and the resulting amplicon was gel-purified and ligated with the pMD18-T vector (Takara, Dalian, China) to allow for its sequencing.
Table 1. Sequences of PCR primers used.
| Name | Forward sequence (5’-3’) | Reverse sequence (5’-3’) |
|---|---|---|
| TaTIP2 | GCTCTAGAATGCCGGGCTCCATCGCCTTCG | CGAGCTCTTAGTAGTCGTTGCCGGCGACGGA |
| TaTIP2;2 | CTCTCATCCTCCCAGTTCTGTTC | CACGTACCGGTAGACGACGC |
| Actin | GTTCCAATCTATGAGGGATACACGC | GAACCTCCACTGAGAACAACATTACC |
| GC island1 | TTGGTGGTTATATAATTTTGGAGGT | CAAAACAATTTTTCAAATCCAATAC |
| GC island2 | TGTTAAGGGGGAAGTTGATATTTA | AAAAAATACCATAACATACACCAAC |
| TaTIP2;2-GFP | GCTCTAGAATGCCGGGCTCCATCGCCTTCG | GCGGATCCGTAGTCGTTGCCGGCGACGGA |
A transmembrane domain prediction of the predicted TaTIP2;2 protein was obtained using the TMHMM tool provided at http://www.cbs.dtu.dk/services/TMHMM. The software package DNAMAN v6.0 (http://www.lynnon.com/) was employed to obtain the intron/exon structure of the genomic sequence. The phylogeny of TaTIP2;2 was investigated by comparing its protein sequence with those of TIP sequences from A. thaliana, rice, maize, barley and wheat represented in the GenBank database.
Chromosomal location of TaTIP2;2
The genomic DNA of a full set of wheat cv. Chinese Spring nulli-tetrasomic lines [30] and a partial set of ditelocentric lines [31] was used as a template for PCRs primed with TaTIP2. The PCR comprised an initial denaturation of 95°C/5min, followed by 35 cycles of 94°C/30s, 65°C/40s, 72°C/60s, ending with a final extension of 72°C/10min. The resulting amplicons were separated by agarose electrophoresis.
Analysis of TaTIP2;2 expression
The cDNA first strand was synthesized using a Tianscript RT kit (Tiangen, Beijing, China), and this was used as the template for a semi-quantitative RT-PCR (sqRT-PCR), primed by TaTIP2;2 (primer sequences given in Table 1). The wheat Actin gene (GenBank accession AB181991) was used as a reference (primer sequences given in Table 1). The PCR comprised an initial denaturation of 95°C/5min, followed by 25-30 cycles of 94°C/30s, 55°C/30s, 72°C/60s, ending with a final extension of 72°C/10min.
Methylation analysis of TaTIP2;2 promoter
The methylation status of the TaTIP2;2 promoter was investigated both before and after the imposition of salinity stress (150mM NaCl for 48h). The 2.5kbp sequence upstream of the TaTIP2;2 start codon was obtained from a draft assembly of the wheat cv. Chinese Spring genomic sequence (http://www.cerealsdb.uk.net/CerealsDB/Documents/DOC_search_reads.php). Bisulphate sequencing was applied to genomic DNA extracted from SR3 at the three leaf stage. The DNA was processed with a sodium bisulphate kit (Qiagen, Hilden, Germany) and then used as a PCR template. The detection of CG islands and the design of primers were facilitated by MethPrimer software [32], and the resulting primer sequences are given in Table 1. The PCR comprised an initial denaturation of 94°C/4min, followed by 35 cycles of 94°C/30s, 60°C/30s, 72°C/30s, ending with a final extension of 72°C/10min. The amplified fragments were gel-purified, ligated with the pEASY T1 vector (Transgene) and transformed into E. coli. A random selection of at least 15 clones per amplicon was sequenced. BiQ Analyzer software (http://biq-analyzer.bioinf.mpi-inf.mpg.de/) was used to calculate the the ratios of cytosine methylation at each CG/CNG/CNN site.
Sub-cellular localization of TaTIP2;2 in wheat protoplasts
The TaTIP2;2 gene without its stop codon was cloned into the XbaI and BamHI sites of the pUC-GFP vector to form a TaTIP2;2::GFP gene fusion. The sequence was amplified using the primer pair TaTIP2;2::GFP (primer sequences given in Table 1), and transferred into wheat cv. Yangmai158 protoplasts isolated from embryogenic calli to perform a transient expression analysis, following the methods described by Yoo et al. [33]. GFP fluorescence was monitored with a Leica TCS SP2 confocal laser scanning microscope under 488nm excitation.
Phenotyping of transgenic A. thaliana
Seed of both wild type and transgenic A. thaliana was surface-sterilized (0.1% w/v HgCl2, 15min), rinsed five times in water, and held at 4°C on half strength MS medium for 36h. To investigate the response to drought and salinity stress, a portion of the seeds was then held at 22°C for 3d, following which they were transferred onto half strength MS medium containing either 300mM mannitol (drought treatment) or 150mM NaCl (salinity treatment). The plates were orientated vertically and held for 10d at 22°C. A second portion of the seed was used to measure seedling proline content. For this purpose, three week old seedlings cultured on MS medium were processed as described by Troll and Lindsley [34].
The expression of stress-related genes in transgenic A. thaliana
RNA of four week old A. thaliana plants on half strength MS was extracted with the same method for SR3 and used as a template for quantitative RT-PCRs targeting the stress-related genes ABI1, ABI2, ABF3, SOS1, SOS2, SOS3, CBF3, DREB2A, DREB2B, MYB2, MYC2, RAB18, RD29B and P5CS1. Relevant primer sequences and the analytical method adopted were as reported elsewhere [35,36].
Results
The sequence characteristics of TaTIP2;2
The SR3 TaTIP2;2 ORF comprises a 747bp sequence, predicted to encode a 248 residue polypeptide. Its sequence is the same with the previously reported TaTIP2;2 (GenBank accession number AY525640) [18]. Alignment of its opening reading frame with the TaTIP2;2 genomic sequence showed that the genomic copy consists of three exons (130bp, 249bp and 367bp) and two introns (83bp and 92bp) (Figure 1A). TMHMM analysis indicated that the protein contains six transmembrane domains (Figure 1B), typical of the aquaporins. An alignment of TaTIP2;2 protein with other plant TIP proteins confirmed the conventional classification of the aquaporin gene family, the three wheat TIP2 proteins clustered closely with one another, and their sequence was ~89% homologous with that of the rice homologue OsTIP2;2 (Figure S1).
Figure 1. Characteristics of TaTIP2;2.
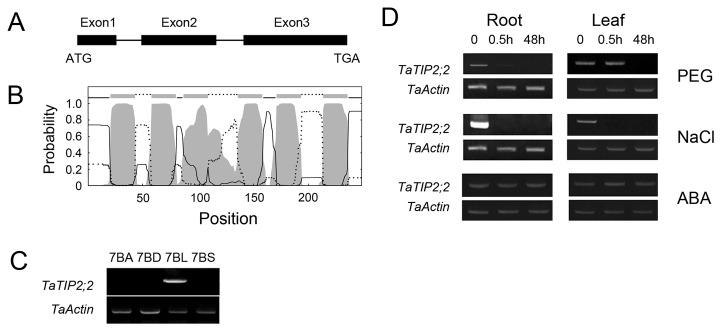
(A) The genomic copy consists of three exons (indicated by filled bars) and two introns (lines). (B) Domain prediction of the gene product. Internal residues indicated by full lines, external ones by dotted lines. The transmembrane domains are shaded grey. (C) Chromosomal location of TaTIP2;2 based on aneuploid stocks of cv. Chinese Spring. 7BA, 7BD are deficient for chromosome 7B, 7BS for chromosome arm 7BL and 7BL for chromosome arm 7BS. (D) sqRT-PCR analysis shows that TaTIP2;2 was expressed in both the root and leaf, and was down-regulated by drought (PEG) and salinity, but not by ABA.
When DNA of the Chinese Spring nulli-tetrasomic lines was amplified with the TaTIP2 primer pair, only lines deficient for chromosome 7B (7BA and 7BD) failed to amplify the expected product. The PCR profile of the ditelocentric line carrying the long arm but not the short arm of 7B (7BL) was the same as that of the euploid, while that of the line carrying the short arm but not the long arm (7BS) was the same as that of the lines deficient for chromosome 7B (Figure 1C). Thus the gene must be located on the long arm of chromosome 7B.
Expression of TaTIP2;2 in response to abiotic stress and exogenous ABA
The outcome of the sqRT-PCR experiments showed that TaTIP2;2 was expressed in both the leaf and root of SR3 (Figure 1D). In the root, the gene was down-regulated following a 0.5h exposure to either salinity or drought stress. In the leaf, it was also down-regulated following a 0.5h exposure to salinity, but its down-regulation in response to PEG treatment was delayed. There was no apparent effect of exogenous ABA on its expression (Figure 1D).
Methylation of the TaTIP2;2 promoter under stress treatment
The 2.5kbp region upstream of the SR3 TaTIP2;2 start codon contained two major CG islands, one located at positions -425 to -616, and the other at -1982 to -2,272. Bisulphate PCR analysis showed that almost all the cytosines present at CG dinucleotide sites were hyper-methylated irrespective of the presence of salinity stress (Figure 2A), while the methylation intensity at the CNG trinucleotide sites was increased by the imposition of salinity stress (Figure 2B).
Figure 2. Methylation status of the TaTIP2;2 promoter region as affected by salinity stress.

(A) CG methylation (filled circles show methylated and open circles unmethylated CG sites). (B) CNG methylation. The horizontal axis shows the positions of CNG sites, and the vertical axis the proportion of methylated sites.
Sub-cellular distribution of TaTIP2;2 protein
Both GFP on its own and the TaTIP2;2::GFP fusion protein were transiently expressed in wheat protoplasts. In the case of the construct containing only GFP, confocal microscopy identified signal throughout the cytoplasm and nucleus (Figure 3A-C). However, in the case of the TaTIP2;2::GFP construct, there was no fluorescence on the cytoplasm membrane; rather the signal was concentrated in the tonoplast of the central vacuole and throughout the endomembrane system (Figure 3D-F).
Figure 3. Sub-cellular localization of TaTIP2;2 protein in wheat protoplasts.

(A-C) A transgene encoding GFP alone generates signal throughout the protoplast. (A) Fluorescent image, (B) bright field image, (C) merger of A and B. (D-F) The TaTIP2;2-GFP fusion is deposited in the endomembrane system. (D) Fluorescent image, (E) bright field image, (F) merger of D and E. Bars, 10μM.
Heterologous expression of TaTIP2;2 in stressed A. thaliana
Two homozygous transgenic T3 A. thaliana selections (OE1 and OE2) carried TaTIP2;2 driven by the CaMV 35S promoter, and successfully expressed the gene (Figure 4A). The behaviour of OE1 and OE2 differed from that of the wild type control in response to exposure to ten days of salinity or osmotic (mannitol) stress. In the presence of NaCl, the growth of the transgenic plants ceased before the cotyledons had fully opened, and most of the seedlings were bleached; in contrast, the cotyledons of the wild type plants opened normally, and bleaching was less frequent (Figure 4B, C). The effect of mannitol stress was to shorten the length of the transgenics' primary roots compared to those of the wild type (Figure 4D, E). Thus the constitutive expression of TaTIP2;2 in A. thaliana compromised the level of drought and salinity. There was no differential response to the supply of 5μM ABA (Figure 4F). The proline content in OE1 and OE2 was lower than in the wild type (Figure 4G).
Figure 4. The behaviour of transgenic A. thaliana expressing.

TaTIP2;2. (A) Genomic PCR (upper panel) and RT-PCR (lower panel) analysis shows that the T3 selections OE1 and OE2 carry and express the transgene. (B) The growth of wild type and transgenic plants challenged with salinity. (C) The rate of bleached progeny from wild type and transgenic plants exposed to 150mM NaCl. (D) The growth of wild type and transgenic plants challenged with mannitol. (E) Root growth of wild type and transgenic plants challenged with mannitol. (F) The growth of wild type and transgenic plants challenged with ABA. (G) The proline content of wild type and transgenic A. thaliana. Col: wild type Col-0 ecotype. Standard deviation was calculated with STDEVP function of Microsoft Excel 2010. Asterisks indicate significant differences between means (Student’s t-test, P<0.05).
Effect of TaTIP2;2 heterologous expression on the expression of abiotic stress-related genes
As revealed by quantitative RT-PCR, the expression level of the ABA signalling genes ABI1, ABI2 and ABF3 was not obviously altered by the presence of the TaTIP2;2 transgene (Figure 5A), similar results were also produced from DREB2B, MYB2, MYC2, RAB18 and RD29B (data not shown), but that of SOS1, SOS2, SOS3, CBF3, DREB2A and P5CS1 was markedly lowered (Figure 5B,C).
Figure 5. The effect of the heterologous expression of TaTIP2;2 on the expression of abiotic stress-related genes in A. thaliana.
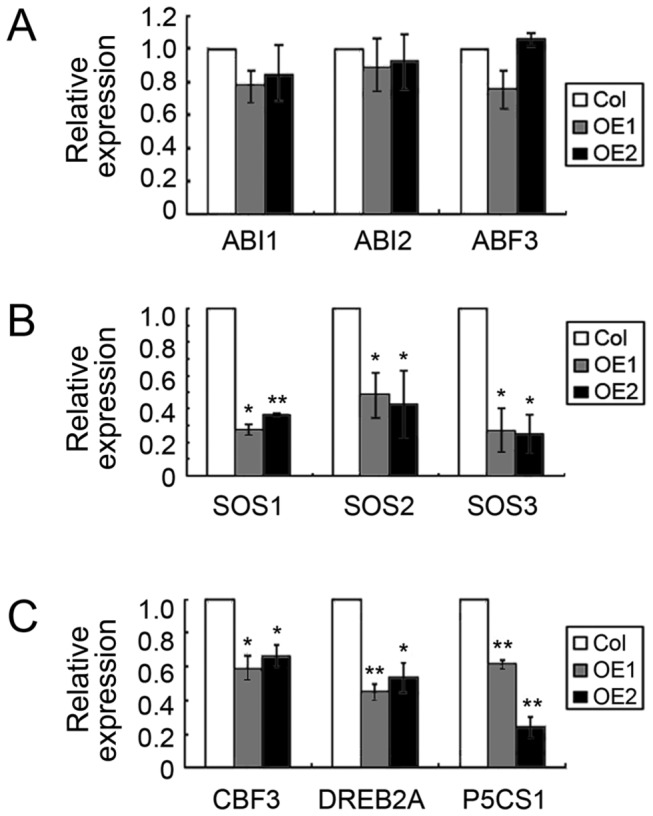
(A) ABI1, ABI2 and ABF3 expression was not affected. (B) SOS1, SOS2 and SOS3 were down-regulated. (C) CBF3, DREB2A and P5CS1 were down-regulated. Col: wild type Col-0 ecotype, OE1 and OE2: T3 selections expressing TaTIP2;2. Standard deviation was calculated with STDEVP function of Microsoft Excel 2010. Asterisks indicate significant differences between means (Student’s t-test, P<0.05 or 0.01).
Discussion
The participation of TaTIP2;2 in the determination of stress tolerance
Response of plant to drought and osmotic stress is intimately related with water transport, in which the aquaporins are involved [8]. Although a number of attempts have been made to characterize the expression profile of various aquaporin genes [3,37], their in vivo function has been much less well researched [7,10,19,38]. Only few examples have been presented which suggest a role for them in stress response. In wheat, Gao et al. [19] showed that TaNIP activity can promote stress tolerance by increasing the K+/Na+ ratio via the regulation of the SOS pathway; wheat PIP aquaporins genes TaAQP7 [21] and TaAQP8 [20], durum wheat PIP genes TdPIP1;1 and TdPIP2;1 [22], cotton PIP gene GhPIP2;7 [9] were also found to be beneficial to stress tolerance. However, many instances of variation in TIP aquaporin expression induced by abiotic stress have been documented. Thus, the down-regulation of both ZmTIP2;2 and TaTIP2;2 was induced by salinity [15,26,39] and that of OsTaTIP2;2 by low temperature [17]. Here, we have demonstrated that the response of TaTIP2;2 to the imposition of stress suggests that it acts as a negative regulator of salinity and drought stress. The observation that this response is independent of ABA is consistent with previous indications that TIP genes generally are not subject to hormonal regulation [14].
Gene expression can be regulated in a number of ways, but a prominent mechanism is represented by promoter methylation. We have shown that the TaTIP2;2 upstream sequence experienced an increase in C-methylation at CNG sites as a result of the imposition of salinity stress, and that TaTIP2;2 expression was reduced by this treatment. With respect to the regulation of gene expression, C-methylation at CNG sites is thought to be more important than at CG sites [40]. As a result, it is possible that these two events are causally related.
It has been suggested that the TIPs are deposited in the tonoplast. With the A. thaliana TIPs, AtTIP1;1 and AtTIP2;2 localized to the tonoplast of the root central vacuole and vacuolar bulbs [41]. Through an analysis of signal peptides or anchors, the site of TaTIP2;2 deposition has been bioinformatically predicted to lie within the plasma membrane [23], a conclusion inconsistent with the present experimental results, which showed that the protein was present not only in wheat protoplast tonoplasts, but also distributed throughout the endomembrane system (Figure 3D). The P. ginseng protein PgTIP1 appears to enhance the level of stress tolerance, but its sub-cellular location has not been determined [7]. A tomato TIP protein SlTIP2;2 was found localized to the tonoplast, and overexpression of it has increased stress tolerance [8]. Contradictory to this, TaTIP2;2 has a negative effect on stress response. Potentially the localization of a given TIP may be related to its effect on the stress response, but as yet there are insufficient relevant data available to reach any sensible conclusion on this issue.
TaTIP2;2 is a negative regulator of stress tolerance
Despite the aquaporins (including the TIPs) having been the focus of a substantial body of research, there is little understanding of how they contribute to the stress response. The A. thaliana proteins AtTIP1;1 and AtTIP1;2 have been characterized as hydrogen peroxide channels, suggesting their possible role in the signalling of stress induced by reactive oxygen species [42]. Salinity stressed A. thaliana lines heterologously expressing PgTIP1;1 can accumulate more Na+ and are more drought tolerant than the wild type [7]. SlTIP2;2 maintains the osmotic water permeability of tonoplast and extends the capacity of vacuole for osmotic buffering under stress [8]. Evidence for the involvement of TIPs in stress response signalling remains scanty.
Proline is frequently used by plants as an osmolyte, and its accumulation is a common response to a wide range of abiotic stresses [43,44]. Glutamate semialdehyde, the precursor of proline, is formed by the reduction of glutamate catalysed by pyrroline-5-carboxylate synthetase (P5CS) [43]. However, although the expression of TaNIP was found to be responsible for increasing the proline content of transgenic A. thaliana, it had no effect on the transcription of P5CS1 [17]. In the present experiments, in contrast, the heterologous expression of TaTIP2;2 did reduce P5CS1 expression (Figure 5C), which may explain the lower proline content of the transgenic lines (Figure 4G). The implication is that TaTIP2;2 expression reduced the osmotic tolerance of transgenic A. thaliana partially via the suppression of proline synthesis.
The SOS pathway genes SOS1, SOS2 and SOS3 are all positive regulators of salinity tolerance in A. thaliana [45]. SOS3 [46] is a calcium sensor, which activates the kinase activity of SOS2 [47]. The resulting complex up-regulates activity of SOS1 [48–50], a Na+/H+ antiporter located on the cell membrane that exports Na+ to the extracellular space and reduces the severity of the salinity stress [48,50]. Heterologous expression of TaNIP has been shown to reduce the level of SOS2 expression, although surprisingly, the phenotypic effect of this reduction was to enhance abiotic stress resistance [19]. All three SOS genes were down-regulated by the TaTIP2;2 transgene (Figure 5B), leading to our hypothesis that the negative impact of TaTIP2;2 expression on the salinity tolerance of transgenic A. thaliana operates via its regulation of the SOS pathway.
Both CBF3 [51] and DREB2A [52] are positive stress regulators acting independently of ABA. Their up-regulation has been shown to enhance the level of drought and salinity stress tolerance. Here, the heterologous expression of TaTIP2;2 down-regulated both genes (Figure 5C), the effect of which would be expected to inhibit plant growth in the presence of abiotic stress. ABA plays a central role in determining stress tolerance [53]. The products of ABI1 [54] and ABI2 [55] are both negative regulators of ABA signalling, while that of ABF3 [56] is a positive regulator. None of these three genes was transcriptionally affected by the heterologous expression of TaTIP2;2 (Figure 5A). Expression of other tested ABA responsive stress regulators MYB2, MYC2 [57,58], RAB18 [59,60] and RD29B [58] was also not changed by the overexpression of TaTIP2;2 (data not shown). These results suggest that the altered stress tolerance of the transgenic A. thaliana plants was not induced through an ABA-dependent pathway. Together with the response of TaTIP2;2 in ABA treated wheat is not obvious (Figure 1D), it can be concluded that TaTIP2;2 is ABA-independent. In the study of A. thaliana [13], rice [61], Brassica napus [12], Craterostigma plantagineum [62] and radish [14] aquaporin genes, ABA-dependent and independent members were both found, suggests that ABA-independent mechanism is commonly involved in the function of aquaporins.
As a channel protein gene, TaTIP2;2 is behooved to lie at the most downstream of the signalling pathways. How its overexpression regulated several upstream genes of these signalling pathways still needs to be revealed. Our results show that TaTIP2;2 localizes in the endomembrane system (Figure 3). Given that tonoplast aquaporins are important for maintaining the osmotic equilibration in the cytoplasm [63], it could be suggested that overexpression of TaTIP2;2 change the water balance inside the cells, and therefore lead to the change of the osmotic pressure in some compartments. This could be detected by osmosensors [57,58,64], which then triggers the signalling pathways and change the expression of stress responsive genes. On the other hand, auqaporins could transport solutes together with water, like H2O2, ammonia, glycerol, and so on [42,65–67]. Some of the solutes (for example, H2O2) are secondary messengers for stress response[68], which could also activate stress signalling and change the expression of the related genes.
In summary, it appears that TaTIP2;2 acts as a negative regulator of the abiotic stress response. It is down-regulated by salinity-induced C-methylation in its promoter region. A possible scenario for improving the stress tolerance of wheat and other crops could be to decrease its expression using a genetic engineering approach.
Supporting Information
Phylogeny of plant TIP proteins. AtTIPs from A. thaliana, HvTIPs from barley, OsTIPs from rice, TaTIPs from wheat, ZmTIPs from maize. TaTIP2;2 shown in bold type.
(TIF)
Funding Statement
This work was supported by the National Transgenic Project (20011ZX08002-002) of China, National Basic Research 973 Program of China (2012CB114200), the funds of the Major Program of the Natural Science Foundation of China (no. 30530480), and National Science Foundation of China (grant no. 91117006). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Kaldenhoff R, Fischer M (2006) Functional aquaporin diversity in plants. Biochim Biophys Acta 1758: 1134-1141. doi: 10.1016/j.bbamem.2006.03.012. PubMed: 16730645. [DOI] [PubMed] [Google Scholar]
- 2. Chaumont F, Barrieu F, Wojcik E, Chrispeels MJ, Jung R (2001) Aquaporins constitute a large and highly divergent protein family in maize. Plant Physiol 125: 1206-1215. doi: 10.1104/pp.125.3.1206. PubMed: 11244102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Forrest KL, Bhave M (2007) Major intrinsic proteins (MIPs) in plants: a complex gene family with major impacts on plant phenotype. Funct Integr Genomics 7: 263-289. doi: 10.1007/s10142-007-0049-4. PubMed: 17562090. [DOI] [PubMed] [Google Scholar]
- 4. Heymann JB, Engel A (1999) Aquaporins: phylogeny, structure, and physiology of water channels. News Physiol Sci 14: 187-193. PubMed: 11390849. [DOI] [PubMed] [Google Scholar]
- 5. Maurel C (2007) Plant aquaporins: novel functions and regulation properties. FEBS Lett 581: 2227-2236. doi: 10.1016/j.febslet.2007.03.021. PubMed: 17382935. [DOI] [PubMed] [Google Scholar]
- 6. Danielson JA, Johanson U (2008) Unexpected complexity of the aquaporin gene family in the moss Physcomitrella patens . BMC Plant Biol 8: 45. doi: 10.1186/1471-2229-8-45. PubMed: 18430224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Peng Y, Lin W, Cai W, Arora R (2007) Overexpression of a Panax ginseng tonoplast aquaporin alters salt tolerance, drought tolerance and cold acclimation ability in transgenic Arabidopsis plants. Planta 226: 729-740. doi: 10.1007/s00425-007-0520-4. PubMed: 17443343. [DOI] [PubMed] [Google Scholar]
- 8. Sade N, Vinocur BJ, Diber A, Shatil A, Ronen G et al. (2009) Improving plant stress tolerance and yield production: is the tonoplast aquaporin SlTIP2;2 a key to isohydric to anisohydric conversion? New Phytol 181: 651-661. doi: 10.1111/j.1469-8137.2008.02689.x. PubMed: 19054338. [DOI] [PubMed] [Google Scholar]
- 9. Zhang J, Li D, Zou D, Luo F, Wang X et al. (2013) A cotton gene encoding a plasma membrane aquaporin is involved in seedling development and in response to drought stress. Acta Biochim Biophys Sin (Shanghai) 45: 104-114. doi: 10.1093/abbs/gms096. [DOI] [PubMed] [Google Scholar]
- 10. Aharon R, Shahak Y, Wininger S, Bendov R, Kapulnik Y et al. (2003) Overexpression of a plasma membrane aquaporin in transgenic tobacco improves plant vigor under favorable growth conditions but not under drought or salt stress. Plant Cell 15: 439-447. doi: 10.1105/tpc.009225. PubMed: 12566583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Aroca R, Ferrante A, Vernieri P, Chrispeels MJ (2006) Drought, abscisic acid and transpiration rate effects on the regulation of PIP aquaporin gene expression and abundance in Phaseolus vulgaris plants. Ann Bot 98: 1301-1310. doi: 10.1093/aob/mcl219. PubMed: 17028296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Gao YP, Young L, Bonham-Smith P, Gusta LV (1999) Characterization and expression of plasma and tonoplast membrane aquaporins in primed seed of Brassica napus during germination under stress conditions. Plant Mol Biol 40: 635-644. doi: 10.1023/A:1006212216876. PubMed: 10480387. [DOI] [PubMed] [Google Scholar]
- 13. Jang JY, Kim DG, Kim YO, Kim JS, Kang H (2004) An expression analysis of a gene family encoding plasma membrane aquaporins in response to abiotic stresses in Arabidopsis thaliana . Plant Mol Biol 54: 713-725. doi: 10.1023/B:PLAN.0000040900.61345.a6. PubMed: 15356390. [DOI] [PubMed] [Google Scholar]
- 14. Suga S, Komatsu S, Maeshima M (2002) Aquaporin isoforms responsive to salt and water stresses and phytohormones in radish seedlings. Plant Cell Physiol 43: 1229-1237. doi: 10.1093/pcp/pcf148. PubMed: 12407203. [DOI] [PubMed] [Google Scholar]
- 15. Zhu C, Schraut D, Hartung W, Schäffner AR (2005) Differential responses of maize MIP genes to salt stress and ABA. J Exp Bot 56: 2971-2981. doi: 10.1093/jxb/eri294. PubMed: 16216844. [DOI] [PubMed] [Google Scholar]
- 16. Boursiac Y, Chen S, Luu DT, Sorieul M, van den Dries N et al. (2005) Early effects of salinity on water transport in Arabidopsis roots. Molecular and cellular features of aquaporin expression. Plant Physiol 139: 790-805. doi: 10.1104/pp.105.065029. PubMed: 16183846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Sakurai J, Ishikawa F, Yamaguchi T, Uemura M, Maeshima M (2005) Identification of 33 rice aquaporin genes and analysis of their expression and function. Plant Cell Physiol 46: 1568-1577. doi: 10.1093/pcp/pci172. PubMed: 16033806. [DOI] [PubMed] [Google Scholar]
- 18. Jahn TP, Møller AL, Zeuthen T, Holm LM, Klaerke DA et al. (2004) Aquaporin homologues in plants and mammals transport ammonia. FEBS Lett 574: 31-36. doi: 10.1016/j.febslet.2004.08.004. PubMed: 15358535. [DOI] [PubMed] [Google Scholar]
- 19. Gao Z, He X, Zhao B, Zhou C, Liang Y et al. (2010) Overexpressing a putative aquaporin gene from wheat, TaNIP, enhances salt tolerance in transgenic Arabidopsis . Plant Cell Physiol 51: 767-775. doi: 10.1093/pcp/pcq036. PubMed: 20360019. [DOI] [PubMed] [Google Scholar]
- 20. Hu W, Yuan Q, Wang Y, Cai R, Deng X et al. (2012) Overexpression of a wheat aquaporin gene, TaAQP8, enhances salt stress tolerance in transgenic tobacco. Plant Cell Physiol 53: 2127-2141. doi: 10.1093/pcp/pcs154. PubMed: 23161856. [DOI] [PubMed] [Google Scholar]
- 21. Zhou S, Hu W, Deng X, Ma Z, Chen L et al. (2012) Overexpression of the wheat aquaporin gene, TaAQP7, enhances drought tolerance in transgenic tobacco. PLOS ONE 7: e52439. doi: 10.1371/journal.pone.0052439. PubMed: 23285044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Ayadi M, Cavez D, Miled N, Chaumont F, Masmoudi K (2011) Identification and characterization of two plasma membrane aquaporins in durum wheat (Triticum turgidum L. subsp. durum) and their role in abiotic stress tolerance. Plant Physiol Biochem 49: 1029-1039. [DOI] [PubMed] [Google Scholar]
- 23. Forrest KL, Bhave M (2008) The PIP and TIP aquaporins in wheat form a large and diverse family with unique gene structures and functionally important features. Funct Integr Genomics 8: 115-133. doi: 10.1007/s10142-007-0065-4. PubMed: 18030508. [DOI] [PubMed] [Google Scholar]
- 24. Xia G, Xiang F, Zhou A, Wang H, Chen H (2003) Asymmetric somatic hybridization between wheat (Triticum aestivum L.) and Agropyron elongatum (Host) Nevishi. Theor Appl Genet 107: 299-305. doi: 10.1007/s00122-003-1247-7. PubMed: 12845444. [DOI] [PubMed] [Google Scholar]
- 25. Peng Z, Wang M, Li F, Lv H, Li C et al. (2009) A proteomic study of the response to salinity and drought stress in an introgression strain of bread wheat. Mol Cell Proteomics 8: 2676-2686. doi: 10.1074/mcp.M900052-MCP200. PubMed: 19734139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Liu C, Li S, Wang M, Xia G (2012) A transcriptomic analysis reveals the nature of salinity tolerance of a wheat introgression line. Plant Mol Biol 78: 159-169. doi: 10.1007/s11103-011-9854-1. PubMed: 22089973. [DOI] [PubMed] [Google Scholar]
- 27. Hoagland DR, Arnon DI (1938) The water-culture method for growing plants without soil. Berkeley, CA: Univ Calif Coll Agric Exp Sta Circ; p. 7. [Google Scholar]
- 28. De Amicis F, Patti T, Marchetti S (2007) Improvement of the pBI121 plant expression vector by leader replacement with a sequence combining a poly(CAA) and a CT motif. Transgenic Res 16: 731-738. doi: 10.1007/s11248-006-9063-x. PubMed: 17237982. [DOI] [PubMed] [Google Scholar]
- 29. Clough SJ, Bent AF (1998) Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana . Plant J 16: 735-743. doi: 10.1046/j.1365-313x.1998.00343.x. PubMed: 10069079. [DOI] [PubMed] [Google Scholar]
- 30. Sears ER (1954) The aneuploids of common wheat. MO Agri Exp Stn. Res Bull 572: 59. [Google Scholar]
- 31. Steinitz-Sears LM, Sears ER (1957) Ultraviolet and X-Ray induced chromosomal aberrations in wheat. Genetics 42: 623-630. PubMed: 17247722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Li LC, Dahiya R (2002) MethPrimer: designing primers for methylation PCRs. Bioinformatics 18: 1427-1431. doi: 10.1093/bioinformatics/18.11.1427. PubMed: 12424112. [DOI] [PubMed] [Google Scholar]
- 33. Yoo SD, Cho YH, Sheen J (2007) Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis. Nat Protoc 2: 1565-1572. doi: 10.1038/nprot.2007.199. PubMed: 17585298. [DOI] [PubMed] [Google Scholar]
- 34. Troll W, Lindsley J (1955) A photometric method for the determination of proline. J Biol Chem 215: 655-660. PubMed: 13242563. [PubMed] [Google Scholar]
- 35. Li C, Lv J, Zhao X, Ai X, Zhu X et al. (2010) TaCHP: a wheat zinc finger protein gene down-regulated by abscisic acid and salinity stress plays a positive role in stress tolerance. Plant Physiol 154: 211-221. doi: 10.1104/pp.110.161182. PubMed: 20639406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Li S, Xu C, Yang Y, Xia G (2010) Functional analysis of TaDi19A, a salt-responsive gene in wheat. Plant Cell Environ 33: 117-129. PubMed: 19895399. [DOI] [PubMed] [Google Scholar]
- 37. Tyerman SD, Niemietz CM, Bramley H (2002) Plant aquaporins: multifunctional water and solute channels with expanding roles. Plant Cell Environ 25: 173-194. doi: 10.1046/j.0016-8025.2001.00791.x. PubMed: 11841662. [DOI] [PubMed] [Google Scholar]
- 38. Lian HL, Yu X, Ye Q, Ding X, Kitagawa Y et al. (2004) The role of aquaporin RWC3 in drought avoidance in rice. Plant Cell Physiol 45: 481-489. doi: 10.1093/pcp/pch058. PubMed: 15111723. [DOI] [PubMed] [Google Scholar]
- 39. Kawaura K, Mochida K, Yamazaki Y, Ogihara Y (2006) Transcriptome analysis of salinity stress responses in common wheat using a 22k oligo-DNA microarray. Funct Integr Genomics 6: 132-142. doi: 10.1007/s10142-005-0010-3. PubMed: 16328439. [DOI] [PubMed] [Google Scholar]
- 40. Chan SW, Henderson IR, Jacobsen SE (2005) Gardening the genome: DNA methylation in Arabidopsis thaliana . Nat Rev Genet 6: 351-360. doi: 10.1038/nrn1665. PubMed: 15861207. [DOI] [PubMed] [Google Scholar]
- 41. Gattolin S, Sorieul M, Hunter PR, Khonsari RH, Frigerio L (2009) In vivo imaging of the tonoplast intrinsic protein family in Arabidopsis roots. BMC Plant Biol 9: 133. doi: 10.1186/1471-2229-9-133. PubMed: 19922653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Bienert GP, Møller AL, Kristiansen KA, Schulz A, Møller IM et al. (2007) Specific aquaporins facilitate the diffusion of hydrogen peroxide across membranes. J Biol Chem 282: 1183-1192. PubMed: 17105724. [DOI] [PubMed] [Google Scholar]
- 43. Szabados L, Savouré A (2010) Proline: a multifunctional amino acid. Trends Plant Sci 15: 89-97. doi: 10.1016/j.tplants.2009.11.009. PubMed: 20036181. [DOI] [PubMed] [Google Scholar]
- 44. Verbruggen N, Hermans C (2008) Proline accumulation in plants: a review. Amino Acids 35: 753-759. doi: 10.1007/s00726-008-0061-6. PubMed: 18379856. [DOI] [PubMed] [Google Scholar]
- 45. Zhu JK (2000) Genetic analysis of plant salt tolerance using Arabidopsis . Plant Physiol 124: 941-948. doi: 10.1104/pp.124.3.941. PubMed: 11080272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Liu J, Zhu JK (1997) An Arabidopsis mutant that requires increased calcium for potassium nutrition and salt tolerance. Proc Natl Acad Sci U S A 94: 14960-14964. doi: 10.1073/pnas.94.26.14960. PubMed: 9405721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Zhu JK, Liu J, Xiong L (1998) Genetic analysis of salt tolerance in Arabidopsis. Evidence for a critical role of potassium nutrition. Plant Cell 10: 1181-1191. doi: 10.1105/tpc.10.7.1181. PubMed: 9668136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Wu SJ, Ding L, Zhu JK (1996) SOS1, a genetic locus essential for salt tolerance and potassium acquisition. Plant Cell 8: 617-627. doi: 10.1105/tpc.8.4.617. PubMed: 12239394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Halfter U, Ishitani M, Zhu JK (2000) The Arabidopsis SOS2 protein kinase physically interacts with and is activated by the calcium-binding protein SOS3. Proc Natl Acad Sci U S A 97: 3735-3740. doi: 10.1073/pnas.97.7.3735. PubMed: 10725350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Shi H, Ishitani M, Kim C, Zhu JK (2000) The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na+/H+ antiporter. Proc Natl Acad Sci U S A 97: 6896-6901. doi: 10.1073/pnas.120170197. PubMed: 10823923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Oh SJ, Song SI, Kim YS, Jang HJ, Kim SY et al. (2005) Arabidopsis CBF3/DREB1A and ABF3 in transgenic rice increased tolerance to abiotic stress without stunting growth. Plant Physiol 138: 341-351. doi: 10.1104/pp.104.059147. PubMed: 15834008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Sakuma Y, Maruyama K, Osakabe Y, Qin F, Seki M et al. (2006) Functional analysis of an Arabidopsis transcription factor, DREB2A, involved in drought-responsive gene expression. Plant Cell 18: 1292-1309. doi: 10.1105/tpc.105.035881. PubMed: 16617101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Zhang J, Jia W, Yang J, Ismail AM (2006) Role of ABA in integrating plant responses to drought and salt stresses. Field Crops Res 97: 9. [Google Scholar]
- 54. Gosti F, Beaudoin N, Serizet C, Webb AA, Vartanian N et al. (1999) ABI1 protein phosphatase 2C is a negative regulator of abscisic acid signaling. Plant Cell 11: 1897-1910. doi: 10.1105/tpc.11.10.1897. PubMed: 10521520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Merlot S, Gosti F, Guerrier D, Vavasseur A, Giraudat J (2001) The ABI1 and ABI2 protein phosphatases 2C act in a negative feedback regulatory loop of the abscisic acid signalling pathway. Plant J 25: 295-303. doi: 10.1046/j.1365-313x.2001.00965.x. PubMed: 11208021. [DOI] [PubMed] [Google Scholar]
- 56. Kang JY, Choi HI, Im MY, Kim SY (2002) Arabidopsis basic leucine zipper proteins that mediate stress-responsive abscisic acid signaling. Plant Cell 14: 343-357. doi: 10.1105/tpc.010362. PubMed: 11884679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Boudsocq M, Laurière C (2005) Osmotic signaling in plants: multiple pathways mediated by emerging kinase families. Plant Physiol 138: 1185-1194. doi: 10.1104/pp.105.061275. PubMed: 16009994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Shinozaki K, Yamaguchi-Shinozaki K (2000) Molecular responses to dehydration and low temperature: differences and cross-talk between two stress signaling pathways. Curr Opin Plant Biol 3: 217-223. doi: 10.1016/S1369-5266(00)00067-4. PubMed: 10837265. [DOI] [PubMed] [Google Scholar]
- 59. Lang V, Palva ET (1992) The expression of a rab-related gene, rab18, is induced by abscisic acid during the cold acclimation process of Arabidopsis thaliana (L.) Heynh. Plant Mol Biol 20: 951-962. [DOI] [PubMed]
- 60. Jakab G, Ton J, Flors V, Zimmerli L, Métraux JP et al. (2005) Enhancing Arabidopsis salt and drought stress tolerance by chemical priming for its abscisic acid responses. Plant Physiol 139: 267-274. doi: 10.1104/pp.105.065698. PubMed: 16113213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Lian HL, Yu X, Lane D, Sun WN, Tang ZC et al. (2006) Upland rice and lowland rice exhibited different PIP expression under water deficit and ABA treatment. Cell Res 16: 651-660. doi: 10.1038/sj.cr.7310068. PubMed: 16773042. [DOI] [PubMed] [Google Scholar]
- 62. Mariaux JB, Bockel C, Salamini F, Bartels D (1998) Desiccation- and abscisic acid-responsive genes encoding major intrinsic proteins (MIPs) from the resurrection plant Craterostigma plantagineum . Plant Mol Biol 38: 1089-1099. doi: 10.1023/A:1006013130681. PubMed: 9869415. [DOI] [PubMed] [Google Scholar]
- 63. Barrieu F, Chaumont F, Chrispeels MJ (1998) High expression of the tonoplast aquaporin ZmTIP1 in epidermal and conducting tissues of maize. Plant Physiol 117: 1153-1163. doi: 10.1104/pp.117.4.1153. PubMed: 9701571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Nongpiur R, Soni P, Karan R, Singla-Pareek SL, Pareek A (2012) Histidine kinases in plants: cross talk between hormone and stress responses. Plant Signal Behav 7: 1230-1237. doi: 10.4161/psb.21516. PubMed: 22902699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Chaumont F, Barrieu F, Jung R, Chrispeels MJ (2000) Plasma membrane intrinsic proteins from maize cluster in two sequence subgroups with differential aquaporin activity. Plant Physiol 122: 1025-1034. doi: 10.1104/pp.122.4.1025. PubMed: 10759498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Beebo A, Thomas D, Der C, Sanchez L, Leborgne-Castel N et al. (2009) Life with and without AtTIP1;1, an Arabidopsis aquaporin preferentially localized in the apposing tonoplasts of adjacent vacuoles. Plant Mol Biol 70: 193-209. doi: 10.1007/s11103-009-9465-2. PubMed: 19229639. [DOI] [PubMed] [Google Scholar]
- 67. Schüssler MD, Alexandersson E, Bienert GP, Kichey T, Laursen KH et al. (2008) The effects of the loss of TIP1;1 and TIP1;2 aquaporins in Arabidopsis thaliana . Plant J 56: 756-767. doi: 10.1111/j.1365-313X.2008.03632.x. PubMed: 18643996. [DOI] [PubMed] [Google Scholar]
- 68. Xiong L, Schumaker KS, Zhu JK (2002) Cell signaling during cold, drought, and salt stress. Plant Cell 14 Suppl: S165-S183. doi: 10.1105/tpc.010278. PubMed: 12045276. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Phylogeny of plant TIP proteins. AtTIPs from A. thaliana, HvTIPs from barley, OsTIPs from rice, TaTIPs from wheat, ZmTIPs from maize. TaTIP2;2 shown in bold type.
(TIF)