

Abstract
The production of host-selective toxins by the necrotrophic fungus Alternaria alternata is essential for the pathogenesis. A. alternata infection in citrus leaves induces rapid lipid peroxidation, accumulation of hydrogen peroxide (H2O2), and cell death. The mechanisms by which A. alternata avoids killing by reactive oxygen species (ROS) after invasion have begun to be elucidated. The ability to coordinate of signaling pathways is essential for the detoxification of cellular stresses induced by ROS and for pathogenicity in A. alternata. A low level of H2O2, produced by the NADPH oxidase (NOX) complex, modulates ROS resistance and triggers conidiation partially via regulating the redox-responsive regulators (YAP1 and SKN7) and the mitogen-activated protein (MAP) kinase (HOG1) mediated pathways, which subsequently regulate the genes required for the biosynthesis of siderophore, an iron-chelating compound. Siderophore-mediated iron acquisition plays a key role in ROS detoxification because of the requirement of iron for the activities of antioxidants (e.g., catalase and SOD). Fungal strains impaired for the ROS-detoxifying system severely reduce the virulence on susceptible citrus cultivars. This paper summarizes the current state of knowledge of signaling pathways associated with cellular responses to multidrugs, oxidative and osmotic stress, and fungicides, as well as the pathogenicity/virulence in the tangerine pathotype of A. alternata.
1. Introduction
Alternaria species have different lifestyles, ranging from saprophytes to endophytes to pathogens [1]. Alternaria species are a highly successful group of fungal pathogens that cause diseases in a wide variety of economically important crops, including apple, broccoli, cauliflower, carrot, citrus, pear, rice, strawberry, tomato, potato, and tobacco, as well as many ornamental and weed species. Due to their wide host range and worldwide distribution, Alternaria species cause severe economic problems. Alternaria species have been reported to cause diseases in nearly 400 plant species; A. alternata alone can infect more than 100 plant species [2–4]. One reason for the success of these pathogens may be attributed to their production of diverse phytotoxins [5, 6]. The host-selective toxins (HSTs) produced by many members of the genus Alternata have unique modes of action and toxicity to their respective host plants. Production of HST is critical for successful pathogenesis because HST-deficient mutants are incapable of attacking their host plants [7–11]. In addition to HSTs, many Alternaria species produce nonhost selective phytotoxins, such as brefeldin A, altertoxin, and tentoxin [1]. Others can produce mycotoxins that are harmful to humans and other animals [12]. Several Alternaria species can also cause upper respiratory tract infections and asthma in humans [13].
Alternaria alternata (Fr.) Keissler has several pathogenic variants, each producing a unique HST and causing disease in different host plants [5, 9, 10, 14, 15]. HSTs produced by Alternaria pathotypes are chemically diverse, ranging from low-molecular-weight compounds to cyclic peptides. The genes encoding polypeptides for biosynthesis of Alternaria HSTs have been shown to reside on a dispensable chromosome [9]. In citrus, A. alternata has two major pathotypes—the tangerine pathotype and the rough lemon type [16]. The citrus pathotypes are morphologically similar and can be differentiated only by pathological and genetic analyses [17]. The rough lemon pathotype, producing the host-selective ACRL toxin, is pathogenic exclusively to lemon (Citrus jambhiri Lush) and Rangpur lime (Citrus x limonia Osbeck). ACRL toxin affects mitochondrial function, disrupting posttranscriptional RNA splicing and causing metabolite leakage and malfunction of oxidative phosphorylation in susceptible host cells [18, 19]. In contrast, the tangerine pathotype of A. alternata produces the host-selective ACT toxin with a core 9,10-epoxy-8-hydroxy-9-methyl-decatrienoic acid structure [20] and causes brown spots on citrus leaves and fruit. ACT toxin is highly toxic to tangerines (C. reticulata Blanco) and grapefruit (C. paradisi Macfad.), as well as hybrids from grapefruit and tangerine, or tangerine and sweet orange (C. sinensis Osbeck). ACT toxin does not affect rough lemon or Rangpur lime [20]. The toxin is quickly translocated outward through the vascular system, causing rapid electrolyte leakage and necrotic lesions along the veins (Figure 1). A. alternata infection in citrus leaves induces rapid lipid peroxidation and accumulation of hydrogen peroxide (H2O2) [21]. Studies show that A. alternata has evolved a dramatic flexibility and uniqueness in the signaling pathways in order to respond to diverse environmental stimuli and to thrive within host plants. This paper discusses signaling pathways related to oxidative and osmotic stress resistance, fungicide sensitivity, conidia formation, and pathogenesis of A. alternata.
Figure 1.
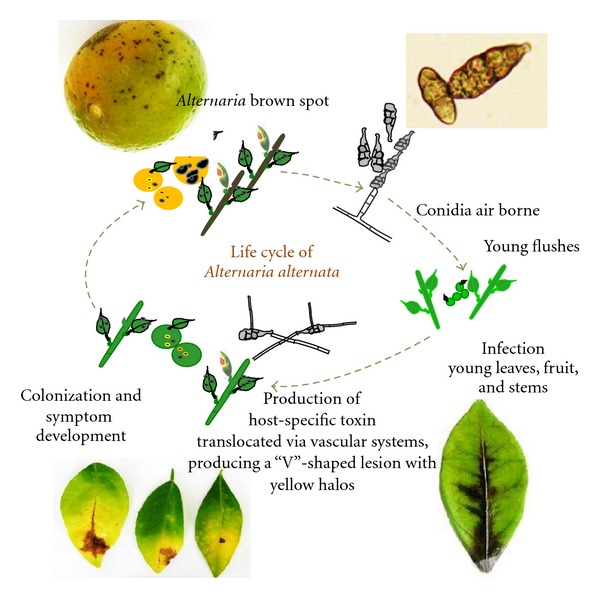
Life cycle of Alternaria alternata, the causal agent of citrus brown spot. ACT toxin produced by the tangerine pathotype of A. alternata is transported via the vascular system and formation of necrotic lesions on a detached calamondin leaf (bottom right).
2. Roles of Reactive Oxygen Species in Plant-Fungal Interactions
All organisms with an aerobic lifestyle inevitably generate toxic reactive oxygen species (ROS), primarily superoxide (O2 −), and hydrogen peroxide (H2O2) during physiological metabolisms [22–26]. During the course of host colonization, fungal pathogens of plants need to overcome a wide range of potentially harmful environmental challenges, particularly an oxidative burst, which could result in the production and accumulation of highly toxic ROS. In addition to the direct toxicity of ROS to cells, when produced in abundance, ROS can also serve as secondary messengers in the pathogen-response signal transduction pathways [23, 27]. Among ROS, H2O2 is relatively stable and able to pass freely through membranes, serving as a signaling cue for defense responses in surrounding cells and as a substrate for oxidative cross-linking in the plant cell wall [27–32]. Hydrogen peroxide can react with O2 − via the Haber-Weiss reaction or with metal ions via the Fenton pathway [33–35] to generate the extremely toxic hydroxyl radical. It has been well known that plants produce toxic ROS as a defense against pathogens [36–41]. In response to the microbe invasion, plant cells often produce excessive amounts of H2O2 by a specific plasma membrane NADPH oxidase, termed as the hypersensitive reaction (HR), which leads to programmed cell death and cellular defense against pathogen attack [42–46]. The HR plays a vital role in plant defenses against saprophytes and noncompatible or biotrophic pathogens; however, HR has been shown less effective against necrotrophic phytopathogens [47–51]. ROS have been shown to be involved in nonhost resistance in plants as well [52, 53].
The burst of the HR may ironically provide an advantage for necrotrophic phytopathogens, as they acquire nutrients exclusively from dead cells [54]. ROS have been thought to enhance plant colonization by necrotrophic pathogens such as Botrytis cinerea and Sclerotinia sclerotiorum [50, 51, 55]. Because many necrotrophic pathogens are able to produce a wide array of HSTs or cell-wall-degrading enzymes that kill host cells before colonization, leading to the accumulation of ROS, the pathogens must have evolved effective mechanisms to cope with the toxicity of ROS [47, 49].
3. YAP1-Mediated ROS Detoxification in A. alternata
ROS damage a wide range of biological molecules, including fatty acids, proteins/enzymes, sugars, and nucleic acids; thus, exposure to ROS may result in cell death [56–58]. The relative sensitivity of the fungal pathogen to ROS is likely determined by the effectiveness of its own ROS detoxification ability. In order to survive under aerobic conditions, fungi must have detoxification systems that can effectively scavenge ROS, maintain reduced redox states within subcellular microenvironments, and repair ROS-triggered damage [22, 59, 60]. Molecular and genetic studies aimed at understanding the mechanisms by which cells cope with the oxidative stresses and are protected from the deleterious effects of ROS have been intensively studied in both prokaryotes and eukaryotes. In the budding yeast Saccharomyces cerevisiae, the YAP1 transcription regulator plays a central role in the cellular pathways associated with the oxidative stress response [61, 62]. YAP1 is responsible for transcriptional activation of genes involved in multidrug resistance as well. YAP1, resembling mammalian AP-1, has a basic leucine zipper (bZIP) domain and has been shown to be activated by H2O2 and various ROS-generating oxidants, as well as heavy metals [63–66]. In the absence of oxidative challenges, YAP1 can be found in the cytoplasm at low levels. Upon perceiving oxidative or chemical stimuli, YAP1 quickly forms disulfide bonds, changes conformation, and is translocated into the nucleus where YAP1 regulates the expression of genes responsible for stress alleviation [67–71]. Conserved cysteine residues in both the amino and carboxyl terminal domains are essential for the formation of the disulfide bonds, nuclear relocalization and transcriptional regulation of YAP1 [72].
Although all microorganisms employ complex mechanisms, both enzymatic and nonenzymatic to avoid ROS toxicity [47, 61, 73], the pathological roles of oxidative stress mitigation remain uncertain in pathogenic fungal species. The role of ROS in host resistance and pathogen invasion is likely dictated by the physiological conditions of the host, the lifestyle of the pathogen, and the combination of different stimuli [38, 41, 74–76]. Hence, ROS produced by plants may have different effects against different pathogens [49, 75]. YAP1-mediated detoxification of ROS is an essential virulence determinant in the opportunistic human pathogen Candida albicans and the biotrophic maize pathogen Ustilago maydis [77, 78]. However, YAP1 is not required for virulence in the plant pathogens Cochliobolus heterostrophus and B. cinerea and in the animal pathogen Aspergillus fumigatus, even though the disrupted mutants exhibit increased sensitivity to H2O2 [79, 80]. The necrotrophic fungus Sc. sclerotiorum produces oxalic acid that suppresses host-generated ROS, and thus facilitates fungal evasion [73]. The Magnaporthe oryzae MoHYR1 gene encoding a glutathione peroxidase (GSHPx) is required for detoxifying plant-generated ROS and full virulence [81]. In S. cerevisiae, HYR1 forms a disulfide bond with YAP1, inducing a conformational alteration and nuclear localization of YAP1 upon exposure to ROS [82].
To thrive within host plants, A. alternata must be able to detoxify or obviate the ROS-mediated plant defense barriers. Our studies have demonstrated that cellular detoxification of ROS regulated by the redox-responsive YAP1 transcription regulator is important for pathogenesis of A. alternata to citrus [83, 84]. Inactivation of the A. alternata AP1 gene (designated AaAP1), encoding a YAP1-like transcription factor, resulted in fungal mutants that are hypersensitive to H2O2, menadione, and potassium superoxide (KO2). The promoter of AaAP1 contains a putative stress responsive element (STRE: AGAGGGG). Upon activation by H2O2, the AaAP1::sGFP fusion protein became localized in the nucleus. Fungal mutants lacking AaAP1 (Δyap1) are weakly virulent on susceptible citrus cultivars even though they synthesize HST toxins normally. However, Δyap1 mutant is not sensitive to osmotic and salt stress-related compounds (e.g., sorbitol, mannitol, NaCl, and KCl) (Figure 2). Δyap1 mutant produces wild-type level of conidia that germinate at a rate and magnitude similar to the wild-type strain. The nonpathogenic phenotype of Δyap1 mutant was accompanied with reduced activities of fungal antioxidants, including catalase, peroxidase, superoxide dismutase (SOD), and glutathione reductase. The inability of Δyap1 mutants to incite necrotic lesions is likely due to the mutants' inability to detoxify ROS because coapplication of Δyap1 mutants with the NADPH oxidase inhibitor, apocynin, or diphenylene iodonium, partially restored lesion-forming capability to the mutants. Δyap1 mutant is impaired in the penetration and colonization stages because the impaired mutant did not cause any visible necrotic lesions on wounded or unwounded leaves of the citrus cultivar Minneola. All mutant phenotypes were completely restored to the wild type in fungal strains expressing a functional copy of AaAP1. Δyap1 mutant displayed severe defects in antioxidant activities and was unable to detoxify H2O2 effectively. Our studies concluded that effective detoxification of ROS via the AaAP1-mediated pathway is absolutely required for successful colonization of citrus by A. alternata [83, 84]. The tobacco pathotype of A. alternata impaired for the biosynthesis of mannitol, an antioxidant and quencher of the hydroxyl radical, also greatly reduces virulence [85, 86], consistent with the importance of ROS detoxification in the pathogenesis of A. alternata.
Figure 2.
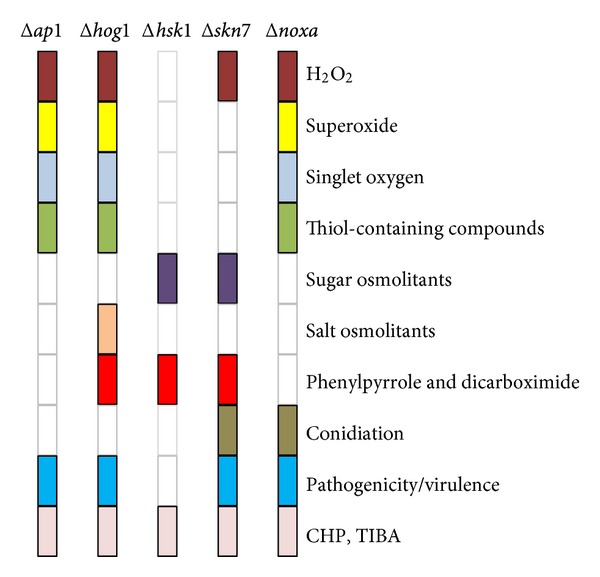
Phenotypic changes in A. alternata mutants lacking the transcription regulator (YAP1), the MAP kinase (HOG1), the “two-component” histidine kinase (HSK1), the response regulator (SKN7), or the NADPH oxidase (NOXA). Open rectangles denote wild-type phenotypes.
Furthermore, AaAP1 was found to be required for full resistance to 2,3,5-triiodobenzoic acid (TIBA), 2-chloro-5-hydroxypyridine (CHP), diethyl maleate (DEM), and many pyridine-containing compounds [87]. Diethyl maleate is a glutathione-depleting agent that has been shown to generate a nonreversible modification of cysteine residues in the Schizosaccharomyces pombe Pap1 protein. As a result, Pap1 is constitutively localized in nucleus and activates the genes required for ROS tolerance [88]. TIBA is often used as herbicides or as an inhibitor of indole-3-acetic acid (IAA) transportation [89, 90]. Pyridine is a heteroaromatic compound composed of five carbons and one nitrogen atom. Pyridine could accelerate the production of superoxide and hydroxyl radicals when Cu2+ and H2O2 are present [91, 92]. Pyridine and its derivatives serve as constituents of RNA and DNA, as electron carriers such as NADP/NADPH and flavin nucleotides (FAD/FADH) and as energy storage molecules such as ATP and GTP.
4. The “Two-Component” Histidine Kinase (HSK) Signaling Pathway
All living cells have a complicated yet well-regulated network often comprising different signaling transduction pathways to perceive changes in their environments and to adjust physiological and developmental processes [93–99]. “Two-component” histidine kinase (HSK) signaling transfer systems are commonly present in bacteria, slime molds, fungi, and plants; however, these systems have not yet been identified in animals [100, 101]. In bacteria, HSK signaling systems contain a histidine kinase (HSK) and a response regulator (RR); each is encoded by a separate gene [102, 103]. In contrast, all fungal HSKs have both the HSK and RR domains [101, 104, 105]. In response to environmental changes, a series of phosphate transfers between histidine (His) and aspartate (Asp) residues occurs in a pattern of His-Asp-His-Asp to regulate downstream signaling pathways such as mitogen-activated protein kinase (MAPK) cascades and eventually leads to a change in gene expression [100, 106, 107].
The S. cerevisiae histidine kinase, designated SLN1p, is required for osmotic adaption via the SLN1p-YPD1p (a protein containing a His phosphotransfer domain)-SSK1p or SKN7p cascade [108, 109]. SSK1p is the major regulator for osmolarity response; SKN7p plays only a minor role in osmosensing. Under normal osmolarity, the SLN1p kinase is phosphorylated and able to activate YPD1p and SSK1p with a phosphorelay mechanism (Figure 3). The phosphorylated SSK1p is inactive and incapable of activating the High Osmolarity-Glycerol 1 (HOG1) MAP kinase pathway (see below for details). In contrast, SLN1p is not phosphorylated under conditions of high osmolarity; therefore SSK1p is able to activate the HOG1-signaling cascade. The activated HOG1 pathway is responsible for glycerol accumulation, allowing the yeast to cope with the high osmolarity. S. cerevisiae also utilizes a non-HSK-related protein SHO1p to cope with osmotic stress [108, 110]. However, deletion of an SHO1 homolog in A. alternata did not impact cellular tolerance to oxidative and osmotic stress, fungicide sensitivity or fungal virulence (L.-H. Chen, unpublished).
Figure 3.
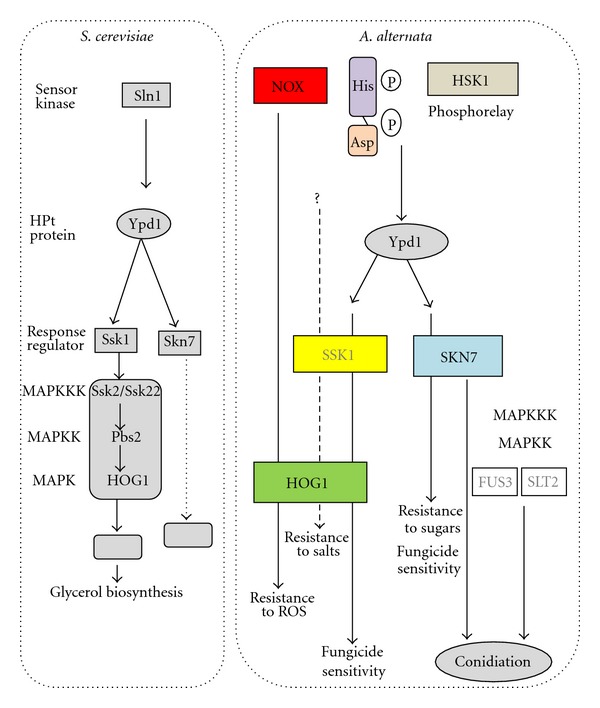
Schematic illustration and comparison of signaling pathways leading to osmotic stress resistance in the budding yeast, S. cerevisiae, and the pathways leading to ROS resistance, osmotic stress response, fungicide sensitivity, and conidia formation in the tangerine pathotype of A. alternata.
The budding yeast S. cerevisiae has only one HSK; all other fungi have multiple HSK signaling genes [111]. Fungal HSKs are divided into 11 groups based on phylogenetic relationships inferred from the conserved HSK and RR domains. Among them, Group III HSK is one of the best characterized HSKs in the filamentous fungi. Collectively, Group III HSK has been implicated in osmotic and oxidative responses, toxin biosynthesis, hyphal development, conidia formation, and virulence, as well as sensitivity to dicarboximide and phenylpyrrole fungicides in different fungal species [112–121].
Signals sensed by HSK are often transduced down to the HOG1 MAP kinase pathway. Fungi lacking Group III HSK or HOG1 often became resistant to dicarboximide and phenylpyrrole fungicides and exhibited an elevated sensitivity to osmotic stress [117, 122, 123]. Although the HSK-HOG1 signaling pathway is conserved, it may be recruited for divergent functions in different fungal species. As discussed above, the S. cerevisiae SLN1p negatively regulates HOG1 phosphorylation under conditions of high osmolarity [100, 106, 107]. The filamentous fungus Co. heterostrophus Group III HSK (Dic-1) positively regulates phosphorylation of the HOG1 MAP kinase, which subsequently activates expression of genes responsible for osmotic resistance and fungicide sensitivity [117]. In B. cinerea, the HOG1-like MAP kinase is not required for fungicide sensitivity even though it is negatively regulated by the “two-component” HSK. Furthermore, the salt-tolerant yeast species, Hortaea werneckii, copes with osmotic stress using a Group VII HSK-HOG1 pathway [124]. Those studies indicate that the HSK-HOG1 signaling pathways can be operated in very different regulatory mechanisms in various species.
The AaHSK1 gene, encoding a putative histidine kinase, was cloned from the tangerine pathotype of A. alternata [87]. AaHSK1, containing no transmembrane regions, is required for adaption to osmotic stress induced by sugars but not by salts (Figure 2). Δhsk1 displayed increased sensitivity to glucose, sucrose, sorbitol, or mannitol, but not to H2O2, KCl, or NaCl [125]. Similarly, the M. grisea HSK is required for resistance to sugar, but not salt, osmotic stress [116]. The F. oxysporum histidine kinase Fhk1 is responsible for resistance to osmotic stress, menadione, but not H2O2 [126]. Similar to Δyap1 mutant, AaHSK1 disruption mutants displayed an elevated sensitivity to TIBA and CHP, suggesting a possible link between YAP1 and HSK. Δhsk1 mutants displayed an elevated resistance to dicarboximide (iprodione and vinclozolin) and phenylpyrrole (fludioxonil) fungicides, suggesting that AaHSK1 is one of the primary targets of these fungicides. Similarly, resistance to dicarboximide and phenylpyrrole fungicides has been demonstrated to be associated with a mutation within the gene encoding a Group III HSK and/or an HOG1 MAP kinase in a number of filamentous fungi [113–118, 121, 122, 126, 127]. However, HSK is not involved in dicarboximide susceptibility in A. longipes.
The A. alternata HSK1 is not required for response to oxidative stress. The AaHSK1 gene product is not involved in pathogenicity or virulence because the AaHSK1-impaired mutants (Δhsk1) induced necrotic lesions at rates and magnitudes similar to the wild-type strain or the genetically reverted strain on wounded or unwounded leaves of citrus. In contrast, Group III HSK is a virulence determinant in the phytopathogenic fungi B. cinerea, Claviceps purpurea, and Fusarium oxysporum and in the human pathogen Cryptococcus neoformans [118, 126, 128, 129].
5. The HOG1 Mitogen-Activated Protein kinase- (MAPK-) Mediated Signaling Pathway
The HOG1 MAPK-mediated signaling cascades in eukaryotic cells are vital for sensing environmental stimuli and for transmitting these signals to the nucleus to modulate gene expression [130, 131]. MAPK-mediated cascade pathways are composed of three serine/threonine protein kinases—MAP kinase kinase kinase (MAPKKK), MAP kinase kinase (MAPKK), and MAP kinase (MAPK). This signal transduction pathway, in conjunction with HSK, is well conserved in all eukaryotes and functions in perceiving environmental stimuli via phosphorylation and gene activation [132, 133]. The phosphorylated MAPK activates a set of genes via regulating appropriate transcription factors.
The A. alternata ortholog (AaHOG1) contains a distinct phosphorylation motif (TGY) involved in the osmotic stress response [134]. Inactivation of the HOG1 ortholog by targeted gene disruption in the tangerine pathotype of A. alternata resulted in mutants that are highly sensitive to the oxidants tert-butyl-hydroxyperoxide, H2O2, and menadione, salts (Figure 2), as well as TIBA and CHP [87]. Because fungal strain lacking the FUS3 MAP kinase (Δfus3) grew faster than wild type in the presence of KCl or NaCl [125]. Thus, AaHOG1 and FUS3 play an opposite role in KCl and NaCl tolerance. HOG1 has been shown to suppress the FUS3/KSS1 signaling cascade during hyperosmotic stress in S. cerevisiae [135, 136]. A. alternata strains impaired at AaHOG1 (Δhog1) displayed wild-type levels of sensitivity to high concentrations of glucose, sucrose, sorbitol, or mannitol (Figure 2) even though sugar osmoticants increased AaHOG1 phosphorylation and subsequently nuclear localization in the Δhsk1 mutant background. In the wild-type background, sugar osmoticant had less effect on AaHOG1 phosphorylation and did not facilitate nuclear localization of AaHOG1.
The wild-type isolate of A. alternata is extremely sensitive to dicarboximide and phenylpyrrole fungicides, whereas fungal strain lacking AaHSK1 is highly resistant to them. Compared to the resistance seen in mutants defective at AaHSK1, Δhog1 mutant displayed only slightly increased resistance to these fungicides. In the wild-type strain of A. alternata, the AaHOG1 protein was phosphorylated at low levels under normal conditions. Exposure to iprodione or fludioxonil fungicide, NaCl, or H2O2 elevated AaHOG1 phosphorylation to varying degrees. Although impairment of AaHSK1 reduces AaHOG1 phosphorylation, A. alternata apparently recruits AaHSK1 and AaHOG1 to exert a unique function in resistance to sugar osmoticants and salt stress, respectively (Figure 3).
Under unchallenged conditions, expression of AaHOG1::sGFP fusion protein under control of the endogenous AaHOG1 promoter in the wild-type strain resulted in green fluorescence uniformly diffused along the hyphal cytoplasm. However, the green fluorescence became dense patches after exposure to H2O2, iprodione and fludioxonil fungicides, or NaCl. Thus, nuclear localization is important for proper functions of HOG1. Compared to Δhsk1 or Δyap1 mutant, Δhog1 mutant was highly resistant to cell wall-degrading enzymes (lyticase, driselase, β-D-glucanase, and β-glucuronidase), thereby failing to generate any protoplasts. As judged from distinct phenotypes in fungal mutants impaired in the AaHSK1 or AaHOG1 gene in A. alternata, it appears that AaHSK1 functions in osmotic tolerance and fungicide sensitivity via AaHOG1 and other gene (e.g., SKN7) activation branches.
AaHSK1 plays no role in pathogenesis of A. alternata. On the other hand, pathogenicity assays revealed that the AaHOG1-impaired mutants are nonpathogenic, producing no necrotic lesions on Minneola leaves that were unwounded or prewounded before inoculation. Similar to AaAP1 disruption, the AaHOG1-impaired mutant is defective at the penetration and colonization steps. Inactivation of the AaHOG1 gene did not impact the production of host-selective ACT toxin by A. alternata. HOG1 is required for virulence/pathogenicity in various fungal pathogens. These include Co. heterostrophus, Cryphonectria parasitica, B. cinerea, Mycosphaerella graminicola, Ca. albicans, and C. neoformans [137–142]. However, HOG1 is not a virulence determinant in M. grisea, Colletotrichum lagenarium, Bipolaris oryzae and As. fumigatus [122, 143–145]. Again, a conserved protein may have very different functions in fungi.
6. SKN7-Mediated ROS Detoxification
“Two-component” HSK-mediated signal transduction is vital for sensing and adapting to environmental changes in microorganisms. In S. cerevisiae, SLN1 histidine kinase transmits signals via a phosphotransfer process down to two response regulators, SSK1p and SKN7p, in response to osmotic stress. However, SKN7p is not regulated by the SLN1p-mediatd phosphorylation in the oxidative stress response [146], indicating that there are two different activation mechanisms in response to osmotic and oxidative stress. Under oxidative stress, SKN7p is phosphorylated at serine or threonine residue, forming a heterodimer with YAP1; together they transcriptionally activate the genes involved in the oxidative stress response [146–152]. YAP1 regulates cadmium resistance independent of SKN7p [153]. SKN7p can also form a heterodimer with the heat-shock transcription factor, the cell cycle transcription regulator, the calcium responsive activator, or the Rho1 GTPase [154–157].
In filamentous fungi, SKN7 is required for oxidative stress adaptation, hypoosmotic stress response, cell cycle, sexual mating, sporulation, cell wall biosynthesis, and fungicide sensitivity [120, 146, 155, 158–163]. The A. alternata SKN7 homolog (AaSKN7) was cloned and characterized in the tangerine pathotype [164]. The promoter of AaSKN7 contains a putative stress responsive element (STRE: AGAGGGG) that is often present in genes induced by various stresses such as oxidative damage in yeasts. AaSKN7 has a heat-shock transcription factor- (HSF-) type helix-turn-helix DNA-binding domain signature and a response regulatory (RR) domain. Genetic mutation analysis revealed that AaSKN7 is required for resistance to osmotic and oxidative stress and fungicide sensitivity, as well as conidiation and conidia morphology. AaSKN7 is primarily localized in the nucleus, whereas YAP1 and HOG1 are quickly transported into the nucleus upon sensing oxidative stress. AaSKN7 may interact directly with AaAP1 in nucleus in response to oxidative stress as demonstrated in the budding yeast [151]. Both A. alternata AP1 and HOG1 are required for resistance to different types of ROS including hydrogen peroxide, superoxide, and singlet oxygen. On the other hand, AaSKN7 is required for resistance to H2O2, tert-butyl hydroperoxide, and cumyl peroxide, but not to superoxide-generating compounds— diamide, menadione and potassium superoxide (Figure 2). It appears that AaSKN7 and AaHOG1 contribute independently to oxidative stress in A. alternata. Because AaHSK1 is not required for ROS resistance [87]; activation of AaSKN7 and AaHOG1 in response to ROS is likely mediated by unknown regulatory sensors other than AaHSK1. Furthermore, A. alternata HSK1 apparently can recruit SKN7 and HOG1 to deal with sugar and salt osmoticants, respectively. Although AaHSK1 and AaSKN7 play no roles in resistance to salt-induced stress, AaSKN7 is involved in resistance to sugar osmoticants likely via the AaHSK1—mediated signaling pathway (Figure 3). Δskn7/Δhog1 double mutants were hypersensitive to both salts and sugars, indicating that A. alternata is capable of sensing different environmental stimuli using distinct or shared signaling pathways. Furthermore, AaSKN7, independent of AaHSK1, is involved in conidia formation. Our studies also revealed that formation of conidia by A. alternata is closely regulated by the FUS3 and SLT2 MAP kinases-mediated signaling pathways, as well as by the G-protein and the NOX complex [125, 165–167]. However, recent studies revealed that cAMP-dependent protein kinase A (PKA) suppresses conidia formation by the tangerine pathotype of Alternaria alternata [168]. It remains uncertain if these signaling pathways actually interact during conidia formation.
A. alternata strains deleted for HSK1 or HOG1 showed an elevated resistance to dicarboximide and phenylpyrrole fungicides. Δskn7 mutant displayed an elevated resistance to those fungicides at levels between Δhsk1 and Δhog1 mutant strains, indicating that the involvement of SKN7 in fungicide sensitivity is likely mediated by the HSK1 signaling pathway. Fungal strain carrying skn7/hog1 double mutations exhibited fungicide resistance, similar to the strain carrying a single AaHSK1 gene mutation. The results indicated that the signals associated with fungicide sensitivity are transduced from AaHSK1 simultaneously down to both AaSKN7- and AaHOG-mediated pathways. The HSK-HOG signaling pathways are associated with fungicide susceptibility in A. brassicicola and N. crassa [121]; yet SKN7 is not involved in fungicide sensitivity in N. crassa. HSK governs Ssk1p (an upstream regulator of HOG1) and Skn7p for osmolarity adaption and fungicide sensitivity in the phytopathogenic fungus Co. heterostrophus and the human pathogen C. neoformans [128, 169, 170].
The roles of SKN7 in pathogenicity/virulence vary among fungal pathogens. A. alternata SKN7 is required for fungal colonization and lesion development in susceptible cultivars of citrus. Similar to Δyap1 and Δhog1, mutational inactivation of AaSKN7 in A. alternata resulted in reduced activities of catalase, SOD, and peroxidase, confirming further that the ability to detoxify host-generating H2O2 by A. alternata is crucial for successful pathogenesis in citrus. The SKN7 homologs are required for virulence in the human pathogens C. neoformans, Ca. albicans, and Ca. glabrata [150, 171, 172]. Again, in contrast, SKN7 is not a virulence determinant in the plant pathogens Co. heterostrophus and M. oryzae and in the human pathogen As. fumigatus [116, 162, 173].
7. The NADPH Oxidase (NOX): Mediated Signaling Pathway
The NADPH-dependent oxidase transfers electrons from NADPH to the oxygen molecule, leading to the production of a superoxide that is further metabolized to H2O2 by SOD [174, 175]. NOX complex is commonly found in animals, plants, and many multicellular microorganisms, but completely absent in prokaryotes [176]. Functionally, NOX complex plays a crucial role in cellular differentiation and defence response. In humans, the phagocytic NOX complex, involved in the production of superoxide and immunity, contains two major catalytic components gp22phox and gp91phox and multiple regulatory subunits Rac (a small GTPase), p40phox, p47phox, and p67phox [177]. NOX complex is also required for the regulation of hormone responses, cell proliferation, and apoptosis in animals ([178–180]. Activation of gp91phox is primarily regulated by p67phox and Rac2 [181]. Plants also have oxidases analogous to gp91phox, designated respiratory burst homologs (Rboh), which are required for physiological metabolisms and for ROS generation in response to pathogen invasion [175, 182, 183].
Many fungi have NADPH oxidase orthologs, NOXA, NOXB and NOXC that have been documented by genetic analysis to be required for developmental, physiological and pathological functions [25, 76, 184, 185]. Both NOXA, and NOXB are analogous with mammalian gp91phox. Expression of the NOXA/NOXB coding genes is closely regulated by the regulatory subunit, NOXR (p67phox homolog), and the small GTPase (Rac homolog) [185]. Fungal NOXC contains a calcium-binding EF-hand motif and is analogous to the mammalian NOX5 and the plant Rboh enzymes.
The functions of NOX complex in the regulation of multicellular development and pathogenicity vary markedly among fungal species that possess it [186]. Both NOXA and NOXB are involved in the regulation of sclerotia formation in both B. cinerea and Sc. sclerotiorum [141, 187]. In B. cinerea, only NOXB is required for the formation of the penetration structure, even though both NOX isoforms have a role in pathogenicity. In M. grisea, NOXA, and NOXB play a role in pathogenesis because both isoforms are required for the formation of penetration peg under the appressorium [188]. While NOXA is required for the development of sexual fruiting body in fungi, only NOXB is required for ascospore germination in N. crassa and Podospora anserina [189–191]. NOXA, but not NOXB, is required for establishing the mutualistic association between the fungal endophyte Epichloë festucae and perennial ryegrass [76]. When inoculated into its grass host, E. festucae strain lacking NOXA or NOXR becomes pathogenic, showing increased branching and causing severe stunting and premature senescence of the host [192, 193]. NOXA is coordinately regulated by the small GTPase Rac and NOXR as evidenced by yeast two-hybrid and pull-down analyses [194]. Furthermore, the yeast polarity protein orthologs, Bem1 and Cdc24, have recently been proven to be parts of fungal NADPH oxidase complex [195].
The tangerine pathotype of A. alternata has NOXA, NOXB, and NOXR homologs. A. alternata NOXA (AaNOXA) contains a NADPH-binding domain and six transmembrane domains and a ferredoxin synthase-type FAD-binding domain, commonly found in the NOXA-like family. Genetic analysis revealed that AaNOXA is responsible for producing superoxide and H2O2. ΔnoxA mutants accumulated less ROS within hyphae than the wild type, as judged by nitroblue tetrazolium (NBT), 3,3′-diaminobenzidine (DAB), and dichlorodihydrofluorescein diacetate (H2DCFDA) staining for the presence of superoxide and H2O2. Moreover, deletion of AaNOXA in A. alternata resulted in an elevated sensitivity to H2O2, superoxide-generating compounds (menadione and KO2), diamide, SDS, CHP, TIBA, and potent singlet oxygen-generating compounds (hematoporphyrin and rose Bengal) (Figure 2). These deficiencies are similar to the phenotypes previously seen for Δyap1 or Δhog1 mutant. Expression of the AaAP1 and AaHOG1 genes is likely regulated by AaNOXA, as deletion of AaNOXA decreased the accumulation of the AaAP1 and AaHOG1 gene transcripts. Reintroducing and expressing a wild-type AaNOXA in a ΔnoxA mutant restored ROS resistance and expression of both AaAP1 and AaHOG1 genes. ΔnoxA mutants also displayed increased sensitivity to NADPH oxidase inhibitors [diphenylene iodonium (DPI) and apocynin], NO•-generating compounds [sodium nitroprusside (SNP) and hydroxyl amine HCl (HAD)], NO• synthase substrate (L-arginine) and NO• synthase inhibitor [nitroarginine methyl ester (nitro-arg)]. Similar to Δyap1 and Δhog1, ΔnoxA mutants, producing normal ACT toxin, induced significantly smaller and fewer necrotic lesions than the wild type on detached Minneola or calamondin leaves 3 days postinoculation, indicating that NOXA is an important virulence determinant in A. alternata.
NOXA, NOXB, and NOXR are core components of the NOX complex, responsible for the production of H2O2. All three NOX components are required for vegetative growth, conidiation, resistance to oxidative and nitrosative stress, and full virulence. However, each isoform may independently and cooperatively interact with other yet unidentified components under different environmental conditions and during different developmental stages because the degree of impairment varied considerably among individual Δnox mutants. ΔnoxA mutant was more sensitive to H2O2, KO2, and diamide than ΔnoxB or ΔnoxR. In contrast, ΔnoxB or ΔnoxR mutation strain was more sensitive to cumyl H2O2 and SDS than ΔnoxA. The elevated sensitivity of ΔnoxB and ΔnoxR mutants to ROS was also accompanied by a reduced expression of two redox-responsive genes AaAP1 and AaHOG1. Although expression of AaAP1 and HOG1 was upregulated by the NOX system; both AaAP1 and HOG1 negatively regulate the expression of NoxB and NoxR. This transcriptional feedback loop might allow fungus to avoid excessive production of toxic ROS. In A. nidulans, NOXA is regulated by an HOG1 homolog [189]. Expression of the NOX complex coding genes has been shown to be regulated by FUS3/KSS1 and SLT2 MAP kinases in N. crassa and B. cinerea [141, 191]. Mammalian p47phox and p67phox are phosphorylated by the p38 HOG1 MAP kinase.
In fungi, expression of NOXA and NOXB is regulated by NOXR and Rac [141, 191, 194]. However, A. alternata NOXR negatively regulated the expression of NOXA and had no effects for the expression of NOXB. ΔnoxB mutant was highly resistant to calcofluor white, Congo red and dicarboximide and phenylpyrrole fungicides compared to the wild type or the mutant strain lacking NOXA and NOXR. NOXB seemingly plays a negatively regulatory role in the biosynthesis of chitin because ΔnoxB, but not ΔnoxA and ΔnoxR, had higher chitin content than the wild type (S.L. Yang, personal communication). As stated above, fungal strains disrupted at any of the AaAP1, AaHSK1, and AaHOG1 genes were all hypersensitive to CHP and TIBA. ΔnoxA, ΔnoxB, and ΔnoxR mutants also displayed increased sensitivity to these two compounds, suggesting the existence of essential cross-talks between different signaling pathways in the context of multidrug resistance.
The NOX complex has been shown to be required for pathogenicity/virulence in a number of fungal species [141, 167, 188, 196]. Pathogenicity assays revealed that ΔnoxB or ΔnoxR mutant is unable to produce necrotic lesions on unwounded citrus leaves. Both mutants induced wild-type lesions on citrus leaves that were wounded before inoculation, indicating that Δnox mutants are primarily arrested in the penetration stage. Δyap1 and Δhog1 mutants are blocked in both penetration and colonization stages.
8. Nonribosomal Peptide Synthetase- (NPS-) Mediated ROS Detoxification
Deletion of an A. alternata gene (AaNPS6), encoding a polypeptide analogous to fungal nonribosomal peptide synthetases (NPSs) resulted in fungi that reduced accumulation of host-selective toxin and melanin and displayed increased sensitivity to H2O2, superoxide-generating compounds (KO2 and menadione), and iron depletion (L.-H. Chen, personal communication). Δnps6 failed to produce siderophore, a low-molecular organic compound involved in acquiring iron from the environment [197, 198]. In nearly all living cells, iron required for numerous metabolic functions and electron transfer processes plays a vital role for cell proliferation and survival [199]. When starved for iron, microorganisms secrete siderophores to solubilize and extract iron. All fungal siderophores (rhodotorulic acid, fusarinines, coprogens, and ferrichromes) that have been identified contain hydroxamates and are synthesized from an unconventional amino acid, L-ornithine [197, 200, 201]. NPSs function to synthesize linear or cyclic peptides without the aid of ribosomes, adding D- or L-amino acids, proteins, nonproteins, hydroxyl acids and ornithine into nonribosomal peptides. Many of these peptides have medicinal, pharmaceutical, or agricultural values [202]. AM-toxin produced by the apple pathotype of A. alternata, HC-toxin produced by race 1 of the maize pathogen, Co. carbonum, and enniatin produced by Fusarium spp. are all nonribosomal peptides [7, 203, 204].
Many Alternaria species produce and excrete dimethyl coprogen siderophores [205–208]. Coprogen contains a diketopiperazine ring (dimerium acid), in which two N 5-acyl-N 5-hydroxy-ornithine units are joined by a peptide bond [198]. The third acyl ornithine unit is linked to the ring via an ester bond. NPSs are involved in assembling three N 5-acyl-N 5-hydroxy-ornithine units, which are the immediate precursors of hydroxamate siderophores. Fungi often have multiple NPS genes; each encodes a polypeptide with discrete domains—AMP-binding adenylation (A), thiolation (T) or peptidyl carrier protein (ACP), and condensation (C) domains that are organized as a module [209–212]. The adenylation domain is required for recognition and activation of amino acid substrate. The thiolation domain is involved in 4′-phosphopantheine binding. The condensation domain is involved in the formation of a peptide bond and elongation and release of the newly synthesized peptide. The number and order of modules in an NPS affect the length and structure of nonribosomal peptide.
Δyap1 and Δnps6 mutants of A. alternata displayed varying levels of hypersensitivity to H2O2 and superoxide-generating compounds. Δnps6 mutant is less sensitive to ROS than Δyap1, displaying an increased sensitivity to the test oxidants only when applied at higher concentrations: H2O2 (≥0.2%), KO2 (≥20 mM), and menadione (≥5 mM). However, elevated sensitivity to H2O2 seen in Δnps6 or Δyap1 mutant was alleviated by adding ferric iron into the medium, implicating an important role of iron and siderophore-mediated iron acquisition in the ROS resistance. We have observed that expression of the AaNPS6 gene was significantly downregulated in fungal mutant lacking YAP1. Deletion of YAP1 in A. alternata also reduced the production of siderophores. Moreover, the rescued strain expressing a functional copy of YAP1 accumulated wild-type level of siderophores and AaNPS6 gene transcript. Expression of AaNPS6 and production of siderophores were also down-regulated in fungal strain lacking HOG1 or NOXA (L.-H. Chen and S.L. Yang, personal communication), confirming a close linkage between iron acquisition and ROS resistance. The wild-type strain of A. alternata, when grown under iron-depleted conditions, produced barely measurable catalase and SOD activities (L.-H. Chen, personal communication). Both antioxidant activities were detectable in A. alternata cultured under iron-rich conditions. Hence, we concluded that the increased sensitivity to oxidative stress and the reduced pathogenicity seen in Δyap1, Δhog1, ΔnoxA, or Δnps6 were seemingly due to the decreased ability of oxidative stress-detoxifying enzymes.
Genetic analyses revealed that siderophore produced by NPS6 is required for full virulence of the tangerine pathotype of A. alternata. This is likely due to the inability of Δnps6 mutant to detoxify toxic ROS efficiently. Siderophores are also required for fungal pathogenesis in A. brassicicola, As. fumigatus, Co. heterostrophus, Co. miyabeanus, F. graminearum, and M. grisea [208, 210, 213, 214]. However, siderophore is not required for pathogenesis of the basidiomycete maize pathogen, U. maydis [215]. It will be of great interest to determine if NPS6 is also regulated by YAP1 and HOG1 in other fungal species.
9. Conclusions
Based on the observed phenotypes derived from mutants lacking YAP1, HOG1, SKN7, NOX, or NPS6, a regulatory network is assembled to underscore the intricate interplays among these signaling pathways in A. alternata (Figure 4). The NOX complex appears to have an important role in the production of ROS, which may act as secondary messages to regulate various metabolic processes in A. alternata. The NOX complex is required for transcriptional activation of two important regulators, YAP1 and HOG1, which subsequently regulate the expression of genes encoding the nonribosomal peptide synthetase (NPS6) and perhaps other enzymes involved in the biosynthesis of siderophores as well. SKN7 physically interacts with YAP1, regulating the genes involved in ROS detoxification. Maintaining iron homeostasis is critical for ROS detoxification because of the requirement of iron for antioxidant enzymatic activities. Impairment of the NOX complex, the YAP1 regulator, the HOG1 kinase, or the siderophore-mediated iron acquisition in A. alternata impacts its ability to detoxify ROS and to colonize host plant, implicating the importance of ROS detoxification in the successful pathogenesis of A. alternata. In addition to detoxifying ROS, A. alternata utilizes specialized or synergistically regulated signaling pathways, involved in HSK1, HOG1, and/or SKN7, in response to osmotic stress, fungicides, and other toxic compounds. This cross-interaction between different signaling pathways may have eccentric advantages for integrating cellular responses to a broader spectrum of environmental stimuli.
Figure 4.
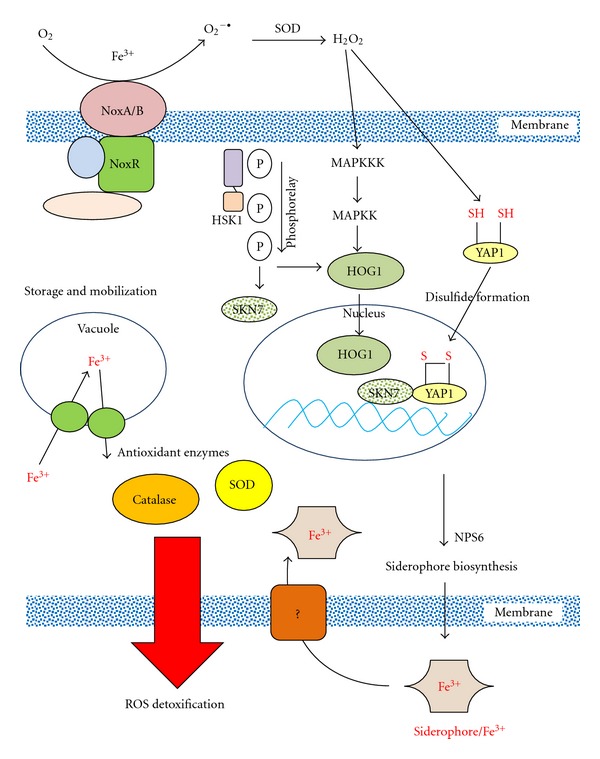
Schematic illustration of signaling pathways leading to ROS detoxification in the tangerine pathotype of A. alternata. H2O2 produced by the membrane-bound NADPH oxidase (NOX) complex plays a central role in the activation of genes responsible for ROS resistance. Upon exposure to ROS, YAP1 forms disulfide bonds between two conserved cysteine residues, undergoes conformation changes, and is transported into the nucleus where YAP1 regulates the expression of numerous genes associated with environmental stress. The YAP1 and SKN7 redox-responsive regulators, the HOG1 mitogen-activated protein (MAP) kinase, the NPS6-mediated siderophore biosynthesis, and the NOX complex are required for ROS detoxification. NPS6 encoding a nonribosomal peptide synthetase is required for the biosynthesis of siderophores, which can extract environmental iron. Iron is stored in vacuoles. Siderophore-mediated iron acquisition plays a critical role if ROS resistance because iron is a major cofactor for the activities of catalase and SOD. HOG1, in cooperation with unknown regulators, is also required for salt resistance. The two-component histidine kinase (HSK1), likely interacting with SKN7, is primarily used for cellular resistance to sugars. Fungicide sensitivity involves HSK1, HOG1, and SKN7.
Acknowledgments
The author would like to thank the current and former Chung Laboratory Members Siwy Ling Yang, Li-Hung Chen, Hsieh-Chin Tsai, and Dr. Ching-Hsuan Lin for their contributions to this work and Wen-Chun Wu for her assistance in taking photographs. This paper was supported by the Florida Agricultural Experiment Station.
References
- 1.Thomma BPHJ. Alternaria spp.: from general saprophyte to specific parasite. Molecular Plant Pathology. 2003;4(4):225–236. doi: 10.1046/j.1364-3703.2003.00173.x. [DOI] [PubMed] [Google Scholar]
- 2.Simmons EG. Alternaria taxonomy: current status, viewpoint, change. In: Chelkowski J, Visconti A, editors. Alternaria Biology, Plant Disease and Metabolites. Amsterdam, The Netherlands: Elsevier; 1992. pp. 1–35. [Google Scholar]
- 3.Kusaba M, Tsuge T. Phylogeny of Alternaria fungi known to produce host specific toxins on the basis of variation in internal transcribed spacers of ribosomal DNA. Current Genetics. 1995;28(5):491–498. doi: 10.1007/BF00310821. [DOI] [PubMed] [Google Scholar]
- 4.Rotem J. The Genus Alternaria: Biology, Epidemiology and Pathogenicity. St. Paul, Minn, USA: APS Press; 1994. [Google Scholar]
- 5.Nishimura S, Kohmoto K. Host-specific toxins and chemical structures from Alternaria species. Annual Review of Phytopathology. 1983;21:87–116. doi: 10.1146/annurev.py.21.090183.000511. [DOI] [PubMed] [Google Scholar]
- 6.Montemurro N, Visconti A. Alternaria metabolites—chemical and biological data. In: Chelkowski J, Visconti A, editors. Alternaria Biology, Plant Disease and Metabolites. Amsterdam, The Netherlands: Elsevier; 1992. pp. 449–557. [Google Scholar]
- 7.Johnson RD, Johnson L, Itoh Y, Kodama M, Otani H, Kohmoto K. Cloning and characterization of a cyclic peptide synthetase gene from Alternaria alternata apple pathotype whose product is involved in AM-toxin synthesis and pathogenicity. Molecular Plant-Microbe Interactions. 2000;13(7):742–753. doi: 10.1094/MPMI.2000.13.7.742. [DOI] [PubMed] [Google Scholar]
- 8.Johnson LJ, Johnson RD, Akamatsu H, et al. Spontaneous loss of a conditionally dispensable chromosome from the Alternaria alternata apple pathotype leads to loss of toxin production and pathogenicity. Current Genetics. 2001;40(1):65–72. doi: 10.1007/s002940100233. [DOI] [PubMed] [Google Scholar]
- 9.Hatta R, Ito K, Hosaki Y, et al. A conditionally dispensable chromosome controls host-specific pathogenicity in the fungal plant pathogen Alternaria alternata . Genetics. 2002;161(1):59–70. doi: 10.1093/genetics/161.1.59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ito K, Tanaka T, Hatta R, Yamamoto M, Akimitsu K, Tsuge T. Dissection of the host range of the fungal plant pathogen Alternaria alternata by modification of secondary metabolism. Molecular Microbiology. 2004;52(2):399–411. doi: 10.1111/j.1365-2958.2004.04004.x. [DOI] [PubMed] [Google Scholar]
- 11.Harimoto Y, Hatta R, Kodama M, Yamamoto M, Otani H, Tsuge T. Expression profiles of genes encoded by the supernumerary chromosome controlling AM-toxin biosynthesis and pathogenicity in the apple pathotype of Alternaria alternata . Molecular Plant-Microbe Interactions. 2007;20(12):1463–1476. doi: 10.1094/MPMI-20-12-1463. [DOI] [PubMed] [Google Scholar]
- 12.Orvehed M, Haggblom P, Soderhall K. Nitrogen inhibition of mycotoxin production by Alternaria alternata . Applied and Environmental Microbiology. 1988;54(10):2361–2364. doi: 10.1128/aem.54.10.2361-2364.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bush RK, Prochnau JJ. Alternaria-induced asthma. Journal of Allergy and Clinical Immunology. 2004;113(2):227–234. doi: 10.1016/j.jaci.2003.11.023. [DOI] [PubMed] [Google Scholar]
- 14.Kohmoto K, Akimitsu K, Otani H. Correlation of resistance and susceptibility of citrus to Alternaria alternata with sensitivity to host-specific toxins. Phytopathology. 1991;81:719–722. [Google Scholar]
- 15.Otani H, Kohmoto K, Kodama M. Alternaria toxins and their effects on host plants. Canadian Journal of Botany. 1996;73:S453–S458. [Google Scholar]
- 16.Peever TL, Su G, Carpenter-Boggs L, Timmer LW. Molecular systematics of citrus-associated Alternaria species. Mycologia. 2004;96(1):119–134. [PubMed] [Google Scholar]
- 17.Peever TL, Canihos Y, Olsen L, Ibañez A, Liu YC, Timmer LW. Population genetic structure and host specificity of Alternaria spp. causing brown spot of Minneola tangelo and rough lemon in Florida. Phytopathology. 1999;89(10):851–860. doi: 10.1094/PHYTO.1999.89.10.851. [DOI] [PubMed] [Google Scholar]
- 18.Akimitsu K, Kohmoto K, Otani H, Nishimura S. Host-specific effect of toxin from the rough lemon pathotype of Alternaria alternata on mitochondria. Plant Physiology. 1989;89:925–931. doi: 10.1104/pp.89.3.925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ohtani K, Yamamoto H, Akimitsu K. Sensitivity to Alternaria alternata toxin in citrus because of altered mitochondrial RNA processing. Proceedings of the National Academy of Sciences of the United States of America. 2002;99(4):2439–2444. doi: 10.1073/pnas.042448499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kohmoto K, Itoh Y, Shimomura N, et al. Isolation and biological activities of two host-specific toxins from tangerine pathotype of Alternaria alternata . Phytopathology. 1993;83:495–502. [Google Scholar]
- 21.Lin CH, Yang SL, Chung KR. Cellular responses required for oxidative stress tolerance, colonization, and lesion formation by the necrotrophic fungus Alternaria alternata in citrus. Current Microbiology. 2011;62(3):807–815. doi: 10.1007/s00284-010-9795-y. [DOI] [PubMed] [Google Scholar]
- 22.Mittler R. Oxidative stress, antioxidants and stress tolerance. Trends in Plant Science. 2002;7(9):405–410. doi: 10.1016/s1360-1385(02)02312-9. [DOI] [PubMed] [Google Scholar]
- 23.Neill S, Desikan R, Hancock J. Hydrogen peroxide signalling. Current Opinion in Plant Biology. 2002;5(5):388–395. doi: 10.1016/s1369-5266(02)00282-0. [DOI] [PubMed] [Google Scholar]
- 24.Apel K, Hirt H. Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annual Review of Plant Biology. 2004;55:373–399. doi: 10.1146/annurev.arplant.55.031903.141701. [DOI] [PubMed] [Google Scholar]
- 25.Aguirre J, Ríos-Momberg M, Hewitt D, Hansberg W. Reactive oxygen species and development in microbial eukaryotes. Trends in Microbiology. 2005;13(3):111–118. doi: 10.1016/j.tim.2005.01.007. [DOI] [PubMed] [Google Scholar]
- 26.Herrero E, Ros J, Bellí G, Cabiscol E. Redox control and oxidative stress in yeast cells. Biochimica et Biophysica Acta. 2008;1780(11):1217–1235. doi: 10.1016/j.bbagen.2007.12.004. [DOI] [PubMed] [Google Scholar]
- 27.Veal EA, Day AM, Morgan BA. Hydrogen peroxide sensing and signaling. Molecular Cell. 2007;26(1):1–14. doi: 10.1016/j.molcel.2007.03.016. [DOI] [PubMed] [Google Scholar]
- 28.Orozco-Cardenas M, Ryan CA. Hydrogen peroxide is generated systemically in plant leaves by wounding and systemin via the octadecanoid pathway. Proceedings of the National Academy of Sciences of the United States of America. 1999;96(11):6553–6557. doi: 10.1073/pnas.96.11.6553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Branco MR, Marinho HS, Cyrne L, Antunes F. Decrease of H2O2 plasma membrane permeability during Adaptation to H2O2 in Saccharomyces cerevisiae . Journal of Biological Chemistry. 2004;279(8):6501–6506. doi: 10.1074/jbc.M311818200. [DOI] [PubMed] [Google Scholar]
- 30.Stone JR, Yang S. Hydrogen peroxide: a signaling messenger. Antioxidants and Redox Signaling. 2006;8(3-4):243–270. doi: 10.1089/ars.2006.8.243. [DOI] [PubMed] [Google Scholar]
- 31.D’Autréaux B, Toledano MB. ROS as signalling molecules: mechanisms that generate specificity in ROS homeostasis. Nature Reviews Molecular Cell Biology. 2007;8(10):813–824. doi: 10.1038/nrm2256. [DOI] [PubMed] [Google Scholar]
- 32.Ślesak I, Libik M, Karpinska B, Karpinski S, Miszalski Z. The role of hydrogen peroxide in regulation of plant metabolism and cellular signalling in response to environmental stresses. Acta Biochimica Polonica. 2007;54(1):39–50. [PubMed] [Google Scholar]
- 33.Halliwell B, Gutteridge JMC. Biologically relevant metal ion-dependent hydroxyl radical generation. An update. FEBS Letters. 1992;307(1):108–112. doi: 10.1016/0014-5793(92)80911-y. [DOI] [PubMed] [Google Scholar]
- 34.Wink DA, Nims RW, Saavedra JE, Utermahlen WE, Jr., Ford PC. The fenton oxidation mechanism: reactivities of biologically relevant substrates with two oxidizing intermediates differ from those predicted for the hydroxyl radical. Proceedings of the National Academy of Sciences of the United States of America. 1994;91(14):6604–6608. doi: 10.1073/pnas.91.14.6604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Imlay JA. Pathways of oxidative damage. Annual Review of Microbiology. 2003;57:395–418. doi: 10.1146/annurev.micro.57.030502.090938. [DOI] [PubMed] [Google Scholar]
- 36.Mehdy MC. Active oxygen species in plant defense against pathogens. Plant Physiology. 1994;105(2):467–472. doi: 10.1104/pp.105.2.467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lam E, Kato N, Lawton M. Programmed cell death, mitochondria and the plant hypersensitive response. Nature. 2001;411(6839):848–853. doi: 10.1038/35081184. [DOI] [PubMed] [Google Scholar]
- 38.Mellersh DG, Foulds IV, Higgins VJ, Heath MC. H2O2 plays different roles in determining penetration failure in three diverse plant-fungal interactions. Plant Journal. 2002;29(3):257–268. doi: 10.1046/j.0960-7412.2001.01215.x. [DOI] [PubMed] [Google Scholar]
- 39.Dangl JL, Jones JDG. Plant pathogens and integrated defence responses to infection. Nature. 2001;411(6839):826–833. doi: 10.1038/35081161. [DOI] [PubMed] [Google Scholar]
- 40.Mahalingam R, Fedoroff N. Stress response, cell death and signalling: the many faces of reactive oxygen species. Physiologia Plantarum. 2003;119(1):56–68. [Google Scholar]
- 41.Shetty NP, Mehrabi R, Lütken H, et al. Role of hydrogen peroxide during the interaction between the hemibiotrophic fungal pathogen Septoria tritici and wheat. New Phytologist. 2007;174(3):637–647. doi: 10.1111/j.1469-8137.2007.02026.x. [DOI] [PubMed] [Google Scholar]
- 42.Doke N, Miura Y, Sanchez LM, et al. The oxidative burst protects plants against pathogen attack: mechanism and role as an emergency signal for plant bio-defence—a review. Gene. 1996;179(1):45–51. doi: 10.1016/s0378-1119(96)00423-4. [DOI] [PubMed] [Google Scholar]
- 43.Lamb C, Dixon RA. The oxidative burst in plant disease resistance. Annual Review of Plant Biology. 1997;48:251–275. doi: 10.1146/annurev.arplant.48.1.251. [DOI] [PubMed] [Google Scholar]
- 44.Greenberg JT. Programmed cell death in plant-pathogen interactions. Annual Review of Plant Biology. 1997;48:525–545. doi: 10.1146/annurev.arplant.48.1.525. [DOI] [PubMed] [Google Scholar]
- 45.Wojtaszek P. Oxidative burst: an early plant response to pathogen infection. Biochemical Journal. 1997;322(3):681–692. doi: 10.1042/bj3220681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Greenberg JT, Yao N. The role of regulation of programmed cell death in plant-pathogen interactions. Cellular Microbiology. 2004;6(3):201–211. doi: 10.1111/j.1462-5822.2004.00361.x. [DOI] [PubMed] [Google Scholar]
- 47.Mayer AM, Staples RC, Gil-ad NL. Mechanisms of survival of necrotrophic fungal plant pathogens in hosts expressing the hypersensitive response. Phytochemistry. 2001;58(1):33–41. doi: 10.1016/s0031-9422(01)00187-x. [DOI] [PubMed] [Google Scholar]
- 48.Able AJ. Role of reactive oxygen species in the response of barley to necrotrophic pathogens. Protoplasma. 2003;221(1-2):137–143. doi: 10.1007/s00709-002-0064-1. [DOI] [PubMed] [Google Scholar]
- 49.Glazebrook J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annual Review of Phytopathology. 2005;43:205–227. doi: 10.1146/annurev.phyto.43.040204.135923. [DOI] [PubMed] [Google Scholar]
- 50.Unger C, Kleta S, Jandl G, Tiedemann AV. Suppression of the defence-related oxidative burst in bean leaf tissue and bean suspension cells by the necrotrophic pathogen Botrytis cinerea . Journal of Phytopathology. 2005;153(1):15–26. [Google Scholar]
- 51.Walz A, Zingen-Sell I, Theisen S, Kortekamp A. Reactive oxygen intermediates and oxalic acid in the pathogenesis of the necrotrophic fungus Sclerotinia sclerotiorum . European Journal of Plant Pathology. 2008;120(4):317–330. [Google Scholar]
- 52.Trujillo M, Kogel KH, Hückelhoven R. Superoxide and hydrogen peroxide play different roles in the nonhost interaction of barley and wheat with inappropriate formae speciales of Blumeria graminis . Molecular Plant-Microbe Interactions. 2004;17(3):304–312. doi: 10.1094/MPMI.2004.17.3.304. [DOI] [PubMed] [Google Scholar]
- 53.Narusaka Y, Narusaka M, Seki M, et al. Cytological and molecular analyses of non-host resistance of Arabidopsis thaliana to Alternaria alternata . Molecular Plant Pathology. 2005;6(6):615–627. doi: 10.1111/j.1364-3703.2005.00310.x. [DOI] [PubMed] [Google Scholar]
- 54.Divon HH, Fluhr R. Nutrition acquisition strategies during fungal infection of plants. FEMS Microbiology Letters. 2007;266(1):65–74. doi: 10.1111/j.1574-6968.2006.00504.x. [DOI] [PubMed] [Google Scholar]
- 55.Govrin EM, Levine A. The hypersensitive response facilitates plant infection by the necrotrophic pathogen Botrytis cinerea . Current Biology. 2000;10(13):751–757. doi: 10.1016/s0960-9822(00)00560-1. [DOI] [PubMed] [Google Scholar]
- 56.Daub ME, Ehrenshaft M. The photoactivated Cercospora toxin cercosporin: contributions to plant disease and fundamental biology. Annual Review of Phytopathology. 2000;38:461–490. doi: 10.1146/annurev.phyto.38.1.461. [DOI] [PubMed] [Google Scholar]
- 57.Thannickal VJ, Fanburg BL. Reactive oxygen species in cell signaling. American Journal of Physiology—Lung Cellular and Molecular Physiology. 2000;279(6):L1005–L1028. doi: 10.1152/ajplung.2000.279.6.L1005. [DOI] [PubMed] [Google Scholar]
- 58.Daub ME, Herrero S, Chung KR. Photoactivated perylenequinone toxins in fungal pathogenesis of plants. FEMS Microbiology Letters. 2005;252(2):197–206. doi: 10.1016/j.femsle.2005.08.033. [DOI] [PubMed] [Google Scholar]
- 59.Miller RA, Britigan BE. Role of oxidants in microbial pathophysiology. Clinical Microbiology Reviews. 1997;10(1):1–18. doi: 10.1128/cmr.10.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Thammavongs B, Denou E, Missous G, Guéguen M, Panoff JM. Response to environmental stress as a global phenomenon in biology: the example of microorganisms. Microbes and Environments. 2008;23(1):20–23. doi: 10.1264/jsme2.23.20. [DOI] [PubMed] [Google Scholar]
- 61.Moye-Rowley WS. Regulation of the transcriptional response to oxidative stress in fungi: similarities and differences. Eukaryotic Cell. 2003;2(3):381–389. doi: 10.1128/EC.2.3.381-389.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Toledano MB, Delaunay A, Monceau L, Tacnet F. Microbial H2O2 sensors as archetypical redox signaling modules. Trends in Biochemical Sciences. 2004;29(7):351–357. doi: 10.1016/j.tibs.2004.05.005. [DOI] [PubMed] [Google Scholar]
- 63.Delaunay A, Isnard AD, Toledano MB. H2O2 sensing through oxidation of the Yap1 transcription factor. The EMBO Journal. 2000;19(19):5157–5166. doi: 10.1093/emboj/19.19.5157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Toone WM, Jones N. Stress-activated signalling pathways in yeast. Genes to Cells. 1998;3(8):485–498. doi: 10.1046/j.1365-2443.1998.00211.x. [DOI] [PubMed] [Google Scholar]
- 65.Toone WM, Jones N. AP-1 transcription factors in yeast. Current Opinion in Genetics and Development. 1999;9(1):55–61. doi: 10.1016/s0959-437x(99)80008-2. [DOI] [PubMed] [Google Scholar]
- 66.Toone WM, Morgan BA, Jones N. Redox control of AP-1-like factors in yeast and beyond. Oncogene. 2001;20(19):2336–2346. doi: 10.1038/sj.onc.1204384. [DOI] [PubMed] [Google Scholar]
- 67.Yan C, Lee LH, Davis LI. Crm1p mediates regulated nuclear export of a yeast AP-1-like transcription factor. The EMBO Journal. 1998;17(24):7416–7429. doi: 10.1093/emboj/17.24.7416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Coleman ST, Epping EA, Steggerda SM, Moye-Rowley WS. Yap1p activates gene transcription in an oxidant-specific fashion. Molecular and Cellular Biology. 1999;19(12):8302–8313. doi: 10.1128/mcb.19.12.8302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Wood MJ, Andrade EC, Storz G. The redox domain of the Yap1p transcription factor contains two disulfide bonds. Biochemistry. 2003;42(41):11982–11991. doi: 10.1021/bi035003d. [DOI] [PubMed] [Google Scholar]
- 70.Gulshan K, Moye-Rowley WS. Multidrug resistance in fungi. Eukaryotic Cell. 2007;6(11):1933–1942. doi: 10.1128/EC.00254-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Okazaki S, Tachibana T, Naganuma A, Mano N, Kuge S. Multistep disulfide bond formation in Yap1 Is required for sensing and transduction of H2O2 stress signal. Molecular Cell. 2007;27(4):675–688. doi: 10.1016/j.molcel.2007.06.035. [DOI] [PubMed] [Google Scholar]
- 72.Kuge S, Arita M, Murayama A, et al. Regulation of the yeast Yap1p nuclear export signal is mediated by redox signal-induced reversible disulfide bond formation. Molecular and Cellular Biology. 2001;21(18):6139–6150. doi: 10.1128/MCB.21.18.6139-6150.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Cessna SG, Sears VE, Dickman MB, Low PS. Oxalic acid, a pathogenicity factor for Sclerotinia sclerotiorum, suppresses the oxidative burst of the host plant. Plant Cell. 2000;12(11):2191–2199. doi: 10.1105/tpc.12.11.2191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Hückelhoven R, Kogel KH. Reactive oxygen intermediates in plant-microbe interactions: Who is who in powdery mildew resistance? Planta. 2003;216(6):891–902. doi: 10.1007/s00425-003-0973-z. [DOI] [PubMed] [Google Scholar]
- 75.Spoel SH, Johnson JS, Dong X. Regulation of tradeoffs between plant defenses against pathogens with different lifestyles. Proceedings of the National Academy of Sciences of the United States of America. 2007;104(47):18842–18847. doi: 10.1073/pnas.0708139104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Takemoto D, Tanaka A, Scott B. NADPH oxidases in fungi: diverse roles of reactive oxygen species in fungal cellular differentiation. Fungal Genetics and Biology. 2007;44(11):1065–1076. doi: 10.1016/j.fgb.2007.04.011. [DOI] [PubMed] [Google Scholar]
- 77.Enjalbert B, MacCallum DM, Odds FC, Brown AJP. Niche-specific activation of the oxidative stress response by the pathogenic fungus Candida albicans . Infection and Immunity. 2007;75(5):2143–2151. doi: 10.1128/IAI.01680-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Molina L, Kahmann R. An Ustilago maydis gene involved in H2O2 detoxification is required for virulence. Plant Cell. 2007;19(7):2293–2309. doi: 10.1105/tpc.107.052332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Lev S, Hadar R, Amedeo P, Baker SE, Yoder OC, Horwitz BA. Activation of an AP1-like transcription factor of the maize pathogen Cochliobolus heterostrophus in response to oxidative stress and plant signals. Eukaryotic Cell. 2005;4(2):443–454. doi: 10.1128/EC.4.2.443-454.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Lessing F, Kniemeyer O, Wozniok I, et al. The Aspergillus fumigatus transcriptional regulator AfYap1 represents the major regulator for defense against reactive oxygen intermediates but is dispensable for pathogenicity in an intranasal mouse infection model. Eukaryotic Cell. 2007;6(12):2290–2302. doi: 10.1128/EC.00267-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Huang K, Czymmek KJ, Caplan JL, Sweigard JA, Donofrio NM. HYR1-mediated detoxification of reactive oxygen species is required for full virulence in the rice blast fungus. PLoS Pathogens. 2011;7(4) doi: 10.1371/journal.ppat.1001335.e1001335 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Kim KH, Willger SD, Park SW, et al. TmpL, a transmembrane protein required for intracellular redox homeostasis and virulence in a plant and an animal fungal pathogen. PLoS Pathogens. 2009;5(11) doi: 10.1371/journal.ppat.1000653.e1000653 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Lin CH, Yang SL, Chung KR. The YAP1 homolog-mediated oxidative stress tolerance is crucial for pathogenicity of the necrotrophic fungus Alternaria alternata in citrus. Molecular Plant-Microbe Interactions. 2009;22(8):942–952. doi: 10.1094/MPMI-22-8-0942. [DOI] [PubMed] [Google Scholar]
- 84.Yang SL, Lin CH, Chung KR. Coordinate control of oxidative stress tolerance, vegetative growth, and fungal pathogenicity via the AP1 pathway in the rough lemon pathotype of Alternaria alternata . Physiological and Molecular Plant Pathology. 2009;74(2):100–110. [Google Scholar]
- 85.Vélëz H, Glassbrook NJ, Daub ME. Mannitol metabolism in the phytopathogenic fungus Alternaria alternata . Fungal Genetics and Biology. 2007;44(4):258–268. doi: 10.1016/j.fgb.2006.09.008. [DOI] [PubMed] [Google Scholar]
- 86.Vélëz H, Glassbrook NJ, Daub ME. Mannitol biosynthesis is required for plant pathogenicity by Alternaria alternata . FEMS Microbiology Letters. 2008;285(1):122–129. doi: 10.1111/j.1574-6968.2008.01224.x. [DOI] [PubMed] [Google Scholar]
- 87.Lin CH, Chung KR. Specialized and shared functions of the histidine kinase- and HOG1 MAP kinase-mediated signaling pathways in Alternaria alternata, a filamentous fungal pathogen of citrus. Fungal Genetics and Biology. 2010;47(10):818–827. doi: 10.1016/j.fgb.2010.06.009. [DOI] [PubMed] [Google Scholar]
- 88.Castillo EA, Ayté J, Chiva C, et al. Diethylmaleate activates the transcription factor Pap1 by covalent modification of critical cysteine residues. Molecular Microbiology. 2002;45(1):243–254. doi: 10.1046/j.1365-2958.2002.03020.x. [DOI] [PubMed] [Google Scholar]
- 89.Tsurumi S, Ohwaki Y. Transport of 14C-lableled indoleacetic acid in Vicia root segments. Plant and Cell Physiology. 1978;19(7):1195–1206. [Google Scholar]
- 90.Lahey KA, Yuan R, Burns JK, Ueng PP, Timmer LW, Chung KR. Induction of phytohormones and differential gene expression in citrus flowers infected by the fungus Colletotrichum acutatum . Molecular Plant-Microbe Interactions. 2004;17(12):1394–1401. doi: 10.1094/MPMI.2004.17.12.1394. [DOI] [PubMed] [Google Scholar]
- 91.Watanabe T, Koller K, Messner K. Copper-dependent depolymerization of lignin in the presence of fungal metabolite, pyridine. Journal of Biotechnology. 1998;62(3):221–230. doi: 10.1016/s0168-1656(98)00063-7. [DOI] [PubMed] [Google Scholar]
- 92.Nerud F, Baldrian P, Gabriel J, Ogbeifun D. Decolorization of synthetic dyes by the Fenton reagent and the Cu/pyridine/H2O2 system. Chemosphere. 2001;44(5):957–961. doi: 10.1016/s0045-6535(00)00482-3. [DOI] [PubMed] [Google Scholar]
- 93.Kunkel BN, Brooks DM. Cross talk between signaling pathways in pathogen defense. Current Opinion in Plant Biology. 2002;5(4):325–331. doi: 10.1016/s1369-5266(02)00275-3. [DOI] [PubMed] [Google Scholar]
- 94.Martindale JL, Holbrook NJ. Cellular response to oxidative stress: signaling for suicide and survival. Journal of Cellular Physiology. 2002;192(1):1–15. doi: 10.1002/jcp.10119. [DOI] [PubMed] [Google Scholar]
- 95.Vranová E, Inzé D, Van Breusegem F. Signal transduction during oxidative stress. Journal of Experimental Botany. 2002;53(372):1227–1236. [PubMed] [Google Scholar]
- 96.Harding HP, Zhang Y, Zeng H, et al. An integrated stress response regulates amino acid metabolism and resistance to oxidative stress. Molecular Cell. 2003;11(3):619–633. doi: 10.1016/s1097-2765(03)00105-9. [DOI] [PubMed] [Google Scholar]
- 97.Ikner A, Shiozaki K. Yeast signaling pathways in the oxidative stress response. Mutation Research. 2005;569(1-2):13–27. doi: 10.1016/j.mrfmmm.2004.09.006. [DOI] [PubMed] [Google Scholar]
- 98.Chauhan N, Latge JP, Calderone R. Signalling and oxidant adaptation in Candida albicans and Aspergillus fumigatus . Nature Reviews Microbiology. 2006;4(6):435–444. doi: 10.1038/nrmicro1426. [DOI] [PubMed] [Google Scholar]
- 99.Pitzschke A, Forzani C, Hirt H. Reactive oxygen species signaling in plants. Antioxidants and Redox Signaling. 2006;8(9-10):1757–1764. doi: 10.1089/ars.2006.8.1757. [DOI] [PubMed] [Google Scholar]
- 100.Thomason P, Kay R. Eukaryotic signal transduction via histidine-aspartate phosphorelay. Journal of Cell Science. 2000;113(18):3141–3150. doi: 10.1242/jcs.113.18.3141. [DOI] [PubMed] [Google Scholar]
- 101.Wolanin PM, Thomason PA, Stock JB. Histidine protein kinases: key signal transducers outside the animal kingdom. Genome Biology. 2002;3(10, article 3013):3013.1–3013.8. doi: 10.1186/gb-2002-3-10-reviews3013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Parkinson JS, Kofoid EC. Communication modules in bacterial signaling proteins. Annual Review of Genetics. 1992;26:71–112. doi: 10.1146/annurev.ge.26.120192.000443. [DOI] [PubMed] [Google Scholar]
- 103.Hoch JA. Two-component and phosphorelay signal transduction. Current Opinion in Microbiology. 2000;3(2):165–170. doi: 10.1016/s1369-5274(00)00070-9. [DOI] [PubMed] [Google Scholar]
- 104.Santos JL, Shiozaki K. Fungal histidine kinases. Science's STKE. 2001;2001(98):p. re1. doi: 10.1126/stke.2001.98.re1. [DOI] [PubMed] [Google Scholar]
- 105.West AH, Stock AM. Histidine kinases and response regulator proteins in two-component signaling systems. Trends in Biochemical Sciences. 2001;26(6):369–376. doi: 10.1016/s0968-0004(01)01852-7. [DOI] [PubMed] [Google Scholar]
- 106.Wurgler-Murphy SM, Saito H. Two-component signal transducers and MAPK cascades. Trends in Biochemical Sciences. 1997;22(5):172–176. doi: 10.1016/s0968-0004(97)01036-0. [DOI] [PubMed] [Google Scholar]
- 107.Kruppa M, Calderone R. Two-component signal transduction in human fungal pathogens. FEMS Yeast Research. 2006;6(2):149–159. doi: 10.1111/j.1567-1364.2006.00024.x. [DOI] [PubMed] [Google Scholar]
- 108.Maeda T, Wurgler-Murphy SM, Saito H. A two-component system that regulates an osmosensing MAP kinase cascade in yeast. Nature. 1994;369(6477):242–245. doi: 10.1038/369242a0. [DOI] [PubMed] [Google Scholar]
- 109.Posas F, Wurgler-Murphy SM, Maeda T, Witten EA, Thai TC, Saito H. Yeast HOG1 MAP kinase cascade is regulated by a multistep phosphorelay mechanism in the SLN1-YPD1-SSK1 ’two-component’ osmosensor. Cell. 1996;86(6):865–875. doi: 10.1016/s0092-8674(00)80162-2. [DOI] [PubMed] [Google Scholar]
- 110.Westfall PJ, Ballon DR, Thorner J. When the stress of your environment makes you go HOG wild. Science. 2004;306(5701):1511–1512. doi: 10.1126/science.1104879. [DOI] [PubMed] [Google Scholar]
- 111.Catlett NL, Yoder OC, Turgeon BG. Whole-genome analysis of two-component signal transduction genes in fungal pathogens. Eukaryotic Cell. 2003;2(6):1151–1161. doi: 10.1128/EC.2.6.1151-1161.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Alex LA, Borkovich KA, Simon MI. Hyphal development in Neurospora crassa: involvement of a two-component histidine kinase. Proceedings of the National Academy of Sciences of the United States of America. 1996;93(8):3416–3421. doi: 10.1073/pnas.93.8.3416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Cui W, Beever RE, Parkes SL, Weeds PL, Templeton MD. An osmosensing histidine kinase mediates dicarboximide fungicide resistance in Botryotinia fuckeliana (Botrytis cinerea) Fungal Genetics and Biology. 2002;36(3):187–198. doi: 10.1016/s1087-1845(02)00009-9. [DOI] [PubMed] [Google Scholar]
- 114.Dry IB, Yuan KH, Hutton DG. Dicarboximide resistance in field isolates of Alternaria alternata is mediated by a mutation in a two-component histidine kinase gene. Fungal Genetics and Biology. 2004;41(1):102–108. doi: 10.1016/j.fgb.2003.09.002. [DOI] [PubMed] [Google Scholar]
- 115.Avenot H, Simoneau P, Iacomi-Vasilescu B, Bataillé-Simoneau N. Characterization of mutations in the two-component histidine kinase gene AbNIK1 from Alternaria brassicicola that confer high dicarboximide and phenylpyrrole resistance. Current Genetics. 2005;47(4):234–243. doi: 10.1007/s00294-005-0568-2. [DOI] [PubMed] [Google Scholar]
- 116.Motoyama T, Kadokura K, Ohira T, et al. A two-component histidine kinase of the rice blast fungus is involved in osmotic stress response and fungicide action. Fungal Genetics and Biology. 2005;42(3):200–212. doi: 10.1016/j.fgb.2004.11.002. [DOI] [PubMed] [Google Scholar]
- 117.Yoshimi A, Kojima K, Takano Y, Tanaka C. Group III histidine kinase is a positive regulator of HOG1-type mitogen-activated protein kinase in filamentous fungi. Eukaryotic Cell. 2005;4(11):1820–1828. doi: 10.1128/EC.4.11.1820-1828.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Viaud M, Fillinger S, Liu W, et al. A class III histidine kinase acts as a novel virulence factor in Bortrytis cinerea . Molecular Plant-Microbe Interactions. 2006;19(9):1042–1050. doi: 10.1094/MPMI-19-1042. [DOI] [PubMed] [Google Scholar]
- 119.Ochiai N, Tokai T, Nishiuchi T, Takahashi-Ando N, Fujimura M, Kimura M. Involvement of the osmosensor histidine kinase and osmotic stress-activated protein kinases in the regulation of secondary metabolism in Fusarium graminearum . Biochemical and Biophysical Research Communications. 2007;363(3):639–644. doi: 10.1016/j.bbrc.2007.09.027. [DOI] [PubMed] [Google Scholar]
- 120.Vargas-Pérez I, Sánchez O, Kawasaki L, Georgellis D, Aguirre J. Response regulators SrrA and SskA are central components of a phosphorelay system involved in stress signal transduction and asexual sporulation in Aspergillus nidulans . Eukaryotic Cell. 2007;6(9):1570–1583. doi: 10.1128/EC.00085-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Dongo A, Bataillé-Simoneau N, Campion C, et al. The group III two-component histidine kinase of filamentous fungi is involved in the fungicidal activity of the bacterial polyketide ambruticin. Applied and Environmental Microbiology. 2009;75(1):127–134. doi: 10.1128/AEM.00993-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Kojima K, Takano Y, Yoshimi A, Tanaka C, Kikuchi T, Okuno T. Fungicide activity through activation of a fungal signalling pathway. Molecular Microbiology. 2004;53(6):1785–1796. doi: 10.1111/j.1365-2958.2004.04244.x. [DOI] [PubMed] [Google Scholar]
- 123.Furukawa K, Hoshi Y, Maeda T, Nakajima T, Abe K. Aspergillus nidulans HOG pathway is activated only by two-component signalling pathway in response to osmotic stress. Molecular Microbiology. 2005;56(5):1246–1261. doi: 10.1111/j.1365-2958.2005.04605.x. [DOI] [PubMed] [Google Scholar]
- 124.Lenassi M, Plemenitaš A. Novel group VII histidine kinase HwHhk7B from the halophilic fungi Hortaea werneckii has a putative role in osmosensing. Current Genetics. 2007;51(6):393–405. doi: 10.1007/s00294-007-0131-4. [DOI] [PubMed] [Google Scholar]
- 125.Lin CH, Yang SL, Wang NY, Chung KR. The FUS3 MAPK signaling pathway of the citrus pathogen Alternaria alternata functions independently or cooperatively with the fungal redox-responsive AP1 regulator for diverse developmental, physiological and pathogenic processes. Fungal Genetics and Biology. 2010;47(4):381–391. doi: 10.1016/j.fgb.2009.12.009. [DOI] [PubMed] [Google Scholar]
- 126.Rispail N, di Pietro A. The two-component histidine kinase Fhk1 controls stress adaptation and virulence of Fusarium oxysporum . Molecular Plant Pathology. 2010;11(3):395–407. doi: 10.1111/j.1364-3703.2010.00612.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Eaton CJ, Jourdain I, Foster SJ, Hyams JS, Scott B. Functional analysis of a fungal endophyte stress-activated MAP kinase. Current Genetics. 2008;53(3):163–174. doi: 10.1007/s00294-007-0174-6. [DOI] [PubMed] [Google Scholar]
- 128.Bahn YS, Kojima K, Cox GM, Heitman J. A unique fungal two-component system regulates stress responses, drug sensitivity, sexual development, and virulence of Cryptococcus neoformans . Molecular Biology of the Cell. 2006;17(7):3122–3135. doi: 10.1091/mbc.E06-02-0113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Nathues E, Jörgens C, Lorenz N, Tudzynski P. The histidine kinase CpHK2 has impact on spore germination, oxidative stress and fungicide resistance, and virulence of the ergot fungus Claviceps purpurea . Molecular Plant Pathology. 2007;8(5):653–665. doi: 10.1111/j.1364-3703.2007.00421.x. [DOI] [PubMed] [Google Scholar]
- 130.Gustin MC, Albertyn J, Alexander M, Davenport K. Map kinase pathways in the yeast Saccharomyces cerevisiae . Microbiology and Molecular Biology Reviews. 1998;62(4):1264–1300. doi: 10.1128/mmbr.62.4.1264-1300.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Qi M, Elion EA. MAP kinase pathways. Journal of Cell Science. 2005;118(16):3569–3572. doi: 10.1242/jcs.02470. [DOI] [PubMed] [Google Scholar]
- 132.Pelech SL, Sanghera JS. MAP kinases: charting the regulatory pathways. Science. 1992;257(5075):1355–1356. doi: 10.1126/science.1382311. [DOI] [PubMed] [Google Scholar]
- 133.Robinson MJ, Cobb MH. Mitogen-activated protein kinase pathways. Current Opinion in Cell Biology. 1997;9(2):180–186. doi: 10.1016/s0955-0674(97)80061-0. [DOI] [PubMed] [Google Scholar]
- 134.Kültz D. Phylogenetic and functional classification of mitogen—and stress—activated protein kinases. Journal of Molecular Evolution. 1998;46(5):571–588. doi: 10.1007/pl00006338. [DOI] [PubMed] [Google Scholar]
- 135.O’Rourke SM, Herskowitz I. The HOG1 MAPK prevents cross talk between the HOG and pheromone response MAPK pathways in Saccharomyces cerevisiae . Genes and Development. 1998;12(18):2874–2886. doi: 10.1101/gad.12.18.2874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Shock TR, Thompson J, Yates JR, Madhani HD. HOG1 mitogen-activated protein kinase (MAPK) interrupts signal transduction between the Kss1 MAPK and the Tec1 transcription factor to maintain pathway specificity. Eukaryotic Cell. 2009;8(4):606–616. doi: 10.1128/EC.00005-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Park SM, Choi ES, Kim MJ, Cha BJ, Yang MS, Kim DH. Characterization of HOG1 homologue, CpMK1, from Cryphonectria parasitica and evidence for hypovirus-mediated perturbation of its phosphorylation in response to hypertonic stress. Molecular Microbiology. 2004;51(5):1267–1277. doi: 10.1111/j.1365-2958.2004.03919.x. [DOI] [PubMed] [Google Scholar]
- 138.Bahn YS, Kojima K, Cox GM, Heitman J. Specialization of the HOG pathway and its impact on differentiation and virulence of Cryptococcus neoformans . Molecular Biology of the Cell. 2005;16(5):2285–2300. doi: 10.1091/mbc.E04-11-0987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Mehrabi R, Zwiers LH, de Waard MA, Kema GHJ. MgHOG1 regulates dimorphism and pathogenicity in the fungal wheat pathogen Mycosphaerella graminicola . Molecular Plant-Microbe Interactions. 2006;19(11):1262–1269. doi: 10.1094/MPMI-19-1262. [DOI] [PubMed] [Google Scholar]
- 140.Menon V, Li D, Chauhan N, et al. Functional studies of the Ssk1p response regulator protein of Candida albicans as determined by phenotypic analysis of receiver domain point mutants. Molecular Microbiology. 2006;62(4):997–1013. doi: 10.1111/j.1365-2958.2006.05438.x. [DOI] [PubMed] [Google Scholar]
- 141.Segmüller N, Kokkelink L, Giesbert S, Odinius D, van Kan J, Tudzynski P. NADPH oxidases are involved in differentiation and pathogenicity in Botrytis cinerea . Molecular Plant-Microbe Interactions. 2008;21(6):808–819. doi: 10.1094/MPMI-21-6-0808. [DOI] [PubMed] [Google Scholar]
- 142.Igbaria A, Lev S, Rose MS, et al. Distinct and combined roles of the MAP kinases of Cochliobolus heterostrophus in virulence and stress responses. Molecular Plant-Microbe Interactions. 2008;21(6):769–780. doi: 10.1094/MPMI-21-6-0769. [DOI] [PubMed] [Google Scholar]
- 143.Dixon KP, Xu JR, Smirnoff N, Talbot NJ. Independent signaling pathways regulate cellular turgor during hyperosmotic stress and appressorium-mediated plant infection by Magnaporthe grisea . Plant Cell. 1999;11(10):2045–2058. doi: 10.1105/tpc.11.10.2045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Du C, Sarfati J, Latge JP, Calderone R. The role of the sakA (HOG1) and tcsB (SLN1) genes in the oxidant adaptation of Aspergillus fumigatus . Medical Mycology. 2006;44(3):211–218. doi: 10.1080/13693780500338886. [DOI] [PubMed] [Google Scholar]
- 145.Moriwaki A, Kihara J, Mori C, Arase S. A MAP kinase gene, BMK1, is required for conidiation and pathogenicity in the rice leaf spot pathogen Bipolaris oryzae . Microbiological Research. 2007;162(2):108–114. doi: 10.1016/j.micres.2006.01.014. [DOI] [PubMed] [Google Scholar]
- 146.Morgan BA, Banks GR, Toone WM, Raitt D, Kuge S, Johnston LH. The Skn7 response regulator controls gene expression in the oxidative stress response of the budding yeast Saccharomyces cerevisiae . The EMBO Journal. 1997;16(5):1035–1044. doi: 10.1093/emboj/16.5.1035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Lu JMY, Deschenes RJ, Fassler JS. Saccharomyces cerevisiae histidine phosphotransferase Ypd1p shuttles between the nucleus and cytoplasm for SLN1-dependent phosphorylation of Ssk1p and Skn7p. Eukaryotic Cell. 2003;2(6):1304–1314. doi: 10.1128/EC.2.6.1304-1314.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Lu JMY, Deschenes RJ, Fassler JS. Role for the ran binding protein, Mog1p, in Saccharomyces cerevisiaeSLN1-SKN7 signal transduction. Eukaryotic Cell. 2004;3(6):1544–1556. doi: 10.1128/EC.3.6.1544-1556.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.He XJ, Fassler JS. Identification of novel Yap1p and Skn7p binding sites involved in the oxidative stress response of Saccharomyces cerevisiae . Molecular Microbiology. 2005;58(5):1454–1467. doi: 10.1111/j.1365-2958.2005.04917.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Wormley FL, Jr., Heinrich G, Miller JL, Perfect JR, Cox GM. Identification and characterization of an SKN7 homologue in Cryptococcus neoformans . Infection and Immunity. 2005;73(8):5022–5030. doi: 10.1128/IAI.73.8.5022-5030.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.He XJ, Mulford KE, Fassler JS. Oxidative stress function of the Saccharomyces cerevisiae skn7 receiver domain. Eukaryotic Cell. 2009;8(5):768–778. doi: 10.1128/EC.00021-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Fassler JS, West AH. Fungal Skn7 stress responses and their relationship to virulence. Eukaryotic Cell. 2011;10(2):156–167. doi: 10.1128/EC.00245-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Lee J, Godon C, Lagniel G, et al. Yap1 and Skn7 control two specialized oxidative stress response regulons in yeast. Journal of Biological Chemistry. 1999;274(23):16040–16046. doi: 10.1074/jbc.274.23.16040. [DOI] [PubMed] [Google Scholar]
- 154.Alberts AS, Bouquin N, Johnston LH, Treisman R. Analysis of RhoA-binding proteins reveals an interaction domain conserved in heterotrimeric G protein β subunits and the yeast response regulator protein Skn7. Journal of Biological Chemistry. 1998;273(15):8616–8622. doi: 10.1074/jbc.273.15.8616. [DOI] [PubMed] [Google Scholar]
- 155.Bouquin N, Johnson AL, Morgan BA, Johnston LH. Association of the cell cycle transcription factor Mbp1 with the Skn7 response regulator in budding yeast. Molecular Biology of the Cell. 1999;10(10):3389–3400. doi: 10.1091/mbc.10.10.3389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Raitt DC, Johnson AL, Erkine AM, et al. The Skn7 response regulator of Saccharomyces cerevisiae interacts with Hsf1 in vivo and is required for the induction of heat shock genes by oxidative stress. Molecular Biology of the Cell. 2000;11(7):2335–2347. doi: 10.1091/mbc.11.7.2335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Williams KE, Cyert MS. The eukaryotic response regulator Skn7p regulates calcineurin signaling through stabilization of Crz1p. The EMBO Journal. 2001;20(13):3473–3483. doi: 10.1093/emboj/20.13.3473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Brown JL, North S, Bussey H. SKN7, a yeast multicopy suppressor of a mutation affecting cell wall β- glucan assembly, encodes a product with domains homologous to prokaryotic two-component regulators and to heat shock transcription factors. Journal of Bacteriology. 1993;175(21):6908–6915. doi: 10.1128/jb.175.21.6908-6915.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Brown JL, Bussey H, Stewart RC. Yeast Skn7p functions in a eukaryotic two-component regulatory pathway. The EMBO Journal. 1994;13(21):5186–5194. doi: 10.1002/j.1460-2075.1994.tb06849.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Tao W, Deschenes RJ, Fassler JS. Intracellular glycerol levels modulate the activity of SLN1p, a Saccharomyces cerevisiae two-component regulator. Journal of Biological Chemistry. 1999;274(1):360–367. doi: 10.1074/jbc.274.1.360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Nakamichi N, Yanada H, Aiba H, Aoyama K, Ohmiya R, Mizuno T. Characterization of the Prr1 response regulator with special reference to sexual development in Schizosaccharomyces pombe . Bioscience, Biotechnology and Biochemistry. 2003;67(3):547–555. doi: 10.1271/bbb.67.547. [DOI] [PubMed] [Google Scholar]
- 162.Oide S, Liu J, Yun SH, et al. Histidine kinase two-component response regulator proteins regulate reproductive development, virulence, and stress responses of the fungal cereal pathogens Cochliobolus heterostrophus and Gibberella zeae . Eukaryotic Cell. 2010;9(12):1867–1880. doi: 10.1128/EC.00150-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Hagiwara D, Mizuno T, Abe K. Characterization of the conserved phosphorylation site in the Aspergillus nidulans response regulator SrrA. Current Genetics. 2011;57(2):103–114. doi: 10.1007/s00294-010-0330-2. [DOI] [PubMed] [Google Scholar]
- 164.Chen L-H, Lin C-H, Chung K-R. Roles for SKN7 response regulator in stress resistance, conidiation and virulence in the citrus pathogen Alternaria alternata . Fungal Genetics and Biology. 2012;49(10):802–813. doi: 10.1016/j.fgb.2012.07.006. [DOI] [PubMed] [Google Scholar]
- 165.Wang NY, Lin CH, Chung KR. A Gα subunit gene is essential for conidiation and potassium efflux but dispensable for pathogenicity of Alternaria alternata on citrus. Current Genetics. 2010;56(1):43–51. doi: 10.1007/s00294-009-0278-2. [DOI] [PubMed] [Google Scholar]
- 166.Yago JI, Lin CH, Chung KR. The SLT2 mitogen-activated protein kinase-mediated signalling pathway governs conidiation, morphogenesis, fungal virulence and production of toxin and melanin in the tangerine pathotype of Alternaria alternata . Molecular Plant Pathology. 2011;12(7):653–665. doi: 10.1111/j.1364-3703.2010.00701.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Yang SL, Chung KR. The NADPH oxidase-mediated production of H2O2and resistance to oxidative stress in the necrotrophic pathogen Alternaria alternata of citrus. Molecular Plant Pathology. 2012;13:900–914. doi: 10.1111/j.1364-3703.2012.00799.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Tsai H-C, Yang SL, Chung K-R. Cyclic AMP-dependent protein kinase A negatively regulates conidia formation by the tangerine pathotype of Alternaria alternata . doi: 10.1007/s11274-012-1182-3. World Journal of Microbiology and Biotechnology. In press. [DOI] [PubMed] [Google Scholar]
- 169.Izumitsu K, Yoshimi A, Tanaka C. Two-component response regulators Ssk1p and Skn7p additively regulate high-osmolarity adaptation and fungicide sensitivity in Cochliobolus heterostrophus . Eukaryotic Cell. 2007;6(2):171–181. doi: 10.1128/EC.00326-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Izumitsu K, Yoshimi A, Hamada S, Morita A, Saitoh Y, Tanaka C. Dic2 and Dic3 loci confer osmotic adaptation and fungicidal sensitivity independent of the HOG pathway in Cochliobolus heterostrophus . Mycological Research. 2009;113(10):1208–1215. doi: 10.1016/j.mycres.2009.08.005. [DOI] [PubMed] [Google Scholar]
- 171.Singh P, Chauhan N, Ghosh A, Dixon F, Calderone R. SKN7 of Candida albicans: mutant construction and phenotype analysis. Infection and Immunity. 2004;72(4):2390–2394. doi: 10.1128/IAI.72.4.2390-2394.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Saijo T, Miyazaki T, Izumikawa K, et al. Skn7p is involved in oxidative stress response and virulence of Candida glabrata . Mycopathologia. 2010;169(2):81–90. doi: 10.1007/s11046-009-9233-5. [DOI] [PubMed] [Google Scholar]
- 173.Lamarre C, Ibrahim-Granet O, Du C, Calderone R, Latgé JP. Characterization of the SKN7 ortholog of Aspergillus fumigatus . Fungal Genetics and Biology. 2007;44(7):682–690. doi: 10.1016/j.fgb.2007.01.009. [DOI] [PubMed] [Google Scholar]
- 174.Lambeth JD. NOX enzymes and the biology of reactive oxygen. Nature Reviews Immunology. 2004;4(3):181–189. doi: 10.1038/nri1312. [DOI] [PubMed] [Google Scholar]
- 175.Fluhr R. Reactive oxygen-generating HADPH oxidases in plants. In: del Río LA, Puppo A, editors. Reactive Oxygen Species in Plant Signaling, Signaling and Communication in Plants. Berlin, Germany: Springer; 2009. pp. 1–23. [Google Scholar]
- 176.Kawahara BT, Quinn MT, Lambeth JD. Molecular evolution of the reactive oxygen-generating NADPH oxidase (Nox/Duox) family of enzymes. BMC Evolutionary Biology. 2007;7, article 109 doi: 10.1186/1471-2148-7-109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Diekmann D, Abo A, Johnston C, Segal AW, Hall A. Interaction of Rac with P67phox and regulation of phagocytic NADPH oxidase activity. Science. 1994;265(5171):531–533. doi: 10.1126/science.8036496. [DOI] [PubMed] [Google Scholar]
- 178.Suh YA, Arnold RS, Lassegue B, et al. Cell transformation by the superoxide-generating oxidase Mox1. Nature. 1999;401(6748):79–82. doi: 10.1038/43459. [DOI] [PubMed] [Google Scholar]
- 179.Lambeth JD, Cheng G, Arnold RS, Edens WA. Novel homologs of gp91phox . Trends in Biochemical Sciences. 2000;25(10):459–461. doi: 10.1016/s0968-0004(00)01658-3. [DOI] [PubMed] [Google Scholar]
- 180.Lardy B, Bof M, Aubry L, et al. NADPH oxidase homologs are required for normal cell differentiation and morphogenesis in Dictyostelium discoideum . Biochimica et Biophysica Acta. 2005;1744(2):199–212. doi: 10.1016/j.bbamcr.2005.02.004. [DOI] [PubMed] [Google Scholar]
- 181.Diebold BA, Bokoch GM. Molecular basis for Rac2 regulation of phagocyte NADPH oxidase. Nature Immunology. 2001;2(3):211–215. doi: 10.1038/85259. [DOI] [PubMed] [Google Scholar]
- 182.Torres MA, Dangl JL, Jones JDG. Arabidopsis gp91phox homologues AtrbohD and AtrbohF are required for accumulation of reactive oxygen intermediates in the plant defense response. Proceedings of the National Academy of Sciences of the United States of America. 2002;99(1):517–522. doi: 10.1073/pnas.012452499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Yoshioka H, Numata N, Nakajima K, et al. Nicotiana benthamianag p91phox homologs NbrbohA and NbrbohB participate in H2O2 accumulation and resistance to Phytophthora infestans . Plant Cell. 2003;15(3):706–718. doi: 10.1105/tpc.008680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Lalucque H, Silar P. NADPH oxidase: an enzyme for multicellularity? Trends in Microbiology. 2003;11(1):9–12. doi: 10.1016/s0966-842x(02)00007-0. [DOI] [PubMed] [Google Scholar]
- 185.Scott B, Eaton CJ. Role of reactive oxygen species in fungal cellular differentiations. Current Opinion in Microbiology. 2008;11(6):488–493. doi: 10.1016/j.mib.2008.10.008. [DOI] [PubMed] [Google Scholar]
- 186.Heller J, Tudzynski P. Reactive oxygen species in phytopathogenic fungi: signaling, development, and disease. Annual Review of Phytopathology. 2011;49:369–390. doi: 10.1146/annurev-phyto-072910-095355. [DOI] [PubMed] [Google Scholar]
- 187.Kim H-J, Chen C, Kabbage M, Dickman MB. Identification and characterization of Sclerotinia sclerotiorum NADPH oxidases. Applied and Environmental Microbiology. 2011;77(21):7721–7729. doi: 10.1128/AEM.05472-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Egan MJ, Wang ZY, Jones MA, Smirnoff N, Talbot NJ. Generation of reactive oxygen species by fungal NADPH oxidases is required for rice blast disease. Proceedings of the National Academy of Sciences of the United States of America. 2007;104(28):11772–11777. doi: 10.1073/pnas.0700574104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Lara-Ortíz T, Riveros-Rosas H, Aguirre J. Reactive oxygen species generated by microbial NADPH oxidase NoxA regulate sexual development in Aspergillus nidulans . Molecular Microbiology. 2003;50(4):1241–1255. doi: 10.1046/j.1365-2958.2003.03800.x. [DOI] [PubMed] [Google Scholar]
- 190.Malagnac F, Lalucque H, Lepère G, Silar P. Two NADPH oxidase isoforms are required for sexual reproduction and ascospore germination in the filamentous fungus Podospora anserina . Fungal Genetics and Biology. 2004;41(11):982–997. doi: 10.1016/j.fgb.2004.07.008. [DOI] [PubMed] [Google Scholar]
- 191.Cano-Domínguez N, Álvarez-Delfín K, Hansberg W, Aguirre J. NADPH oxidases NOX-1 and NOX-2 require the regulatory subunit NOR-1 to control cell differentiation and growth in Neurospora crassa . Eukaryotic Cell. 2008;7(8):1352–1361. doi: 10.1128/EC.00137-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Takemoto D, Tanaka A, Scott B. A P67phox-like regulator is recruited to control hyphal branching in a fungal-grass mutualistic symbiosis. Plant Cell. 2006;18(10):2807–2821. doi: 10.1105/tpc.106.046169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Tanaka A, Christensen MJ, Takemoto D, Park P, Scott B. Reactive oxygen species play a role in regulating a fungus-perennial ryegrass mutualistic interaction. Plant Cell. 2006;18(4):1052–1066. doi: 10.1105/tpc.105.039263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Tanaka A, Takemoto D, Hyon GS, Park P, Scott B. NoxA activation by the small GTPase RacA is required to maintain a mutualistic symbiotic association between Epichloë festucae and perennial ryegrass. Molecular Microbiology. 2008;68(5):1165–1178. doi: 10.1111/j.1365-2958.2008.06217.x. [DOI] [PubMed] [Google Scholar]
- 195.Takemoto D, Kamakura S, Saikia S, et al. Polarity proteins Bem1 and Cdc24 are components of the filamentous fungal NADPH oxidase complex. Proceedings of the National Academy of Sciences of the United States of America. 2011;108(7):2861–2866. doi: 10.1073/pnas.1017309108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Giesbert S, Schürg T, Scheele S, Tudzynski P. The NADPH oxidase Cpnox1 is required for full pathogenicity of the ergot fungus Claviceps purpurea . Molecular Plant Pathology. 2008;9(3):317–327. doi: 10.1111/j.1364-3703.2008.00466.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Haas H. Molecular genetics of fungal siderophore biosynthesis and uptake: the role of siderophores in iron uptake and storage. Applied Microbiology and Biotechnology. 2003;62(4):316–330. doi: 10.1007/s00253-003-1335-2. [DOI] [PubMed] [Google Scholar]
- 198.Haas H, Eisendle M, Turgeon BG. Siderophores in fungal physiology and virulence. Annual Review of Phytopathology. 2008;46:149–187. doi: 10.1146/annurev.phyto.45.062806.094338. [DOI] [PubMed] [Google Scholar]
- 199.Kaplan CD, Kaplan J. Iron acquisition and transcriptional regulation. Chemical Reviews. 2009;109(10):4536–4552. doi: 10.1021/cr9001676. [DOI] [PubMed] [Google Scholar]
- 200.Renshaw JC, Robson GD, Trinci APJ, et al. Fungal siderophores: structures, functions and applications. Mycological Research. 2002;106(10):1123–1142. [Google Scholar]
- 201.Stintzi A, Raymond KN. Siderophore chemistry. In: Templeton DM, editor. Molecular and Cellular Iron Transport. New York, NY, USA: Marcel Dekker; 2002. pp. 273–319. [Google Scholar]
- 202.Kleinkauf H, von Döhren H. A nonribosomal system of peptide biosynthesis. European Journal of Biochemistry. 1996;236(2):335–351. doi: 10.1111/j.1432-1033.1996.00335.x. [DOI] [PubMed] [Google Scholar]
- 203.Panaccione DG, Scott-Craig JS, Pocard JA, Walton JD. A cyclic peptide synthetase gene required for pathogenicity of the fungus Cochliobolus carbonum on maize. Proceedings of the National Academy of Sciences of the United States of America. 1992;89(14):6590–6594. doi: 10.1073/pnas.89.14.6590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Haese A, Schubert M, Herrmann M, Zocher R. Molecular characterization of the enniatin synthetase gene encoding a multifunctional enzyme catalysing N-methyldepsipeptide formation in Fusarium scirpi . Molecular Microbiology. 1993;7(6):905–914. doi: 10.1111/j.1365-2958.1993.tb01181.x. [DOI] [PubMed] [Google Scholar]
- 205.Jalal MAF, Love SK, van der Helm D. Nα-Dimethylcoprogens Three novel trihydroxamate siderophores from pathogenic fungi. Biology of Metals. 1988;1(1):4–8. doi: 10.1007/BF01128011. [DOI] [PubMed] [Google Scholar]
- 206.Jalal MAF, van der Helm D. Siderophores of highly phytopathogenic Alternaria longipes: structures of hydroxycoprogens. Biology of Metals. 1989;2(1):11–17. doi: 10.1007/BF01116195. [DOI] [PubMed] [Google Scholar]
- 207.Ohra J, Morita K, Tsujino Y, et al. Production of two phytotoxic metabolites by the fungus Alternaria cassia . Bioscience, Biotechnology, and Biochemistry. 1995;59:1782–1783. doi: 10.1271/bbb.59.113. [DOI] [PubMed] [Google Scholar]
- 208.Oide S, Moeder W, Krasnoff S, et al. NPS6, encoding a nonribosomal peptide synthetase involved in siderophore-mediated iron metabolism, is a conserved virulence determinant of plant pathogenic ascomycetes. Plant Cell. 2006;18(10):2836–2853. doi: 10.1105/tpc.106.045633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.von Döhren H. Biochemistry and general genetics of nonribosomal peptide synthetases in fungi. Advances in Biochemical Engineering/Biotechnology. 2004;88:217–264. doi: 10.1007/b99262. [DOI] [PubMed] [Google Scholar]
- 210.Lee BN, Kroken S, Chou DYT, Robbertse B, Yoder OC, Turgeon BG. Functional analysis of all nonribosomal peptide synthetases in Cochliobolus heterostrophus reveals a factor, NPS6, involved in virulence and resistance to oxidative stress. Eukaryotic Cell. 2005;4(3):545–555. doi: 10.1128/EC.4.3.545-555.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Bushley KE, Ripoll DR, Turgeon BG. Module evolution and substrate specificity of fungal nonribosomal peptide synthetases involved in siderophore biosynthesis. BMC Evolutionary Biology. 2008;8(1, article 328) doi: 10.1186/1471-2148-8-328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Bushley KE, Turgeon BG. Phylogenomics reveals subfamilies of fungal nonribosomal peptide synthetases and their evolutionary relationships. BMC Evolutionary Biology. 2010;10(1, article 26) doi: 10.1186/1471-2148-10-26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Hissen AHT, Wan ANC, Warwas ML, Pinto LJ, Moore MM. The Aspergillus fumigatus siderophore biosynthetic gene sidA, encoding L-ornithine N 5-oxygenase, is required for virulence. Infection and Immunity. 2005;73(9):5493–5503. doi: 10.1128/IAI.73.9.5493-5503.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Schrettl M, Bignell E, Kragl C, et al. Distinct roles for intra- and extracellular siderophores during Aspergillus fumigatus infection. PLoS Pathogens. 2007;3(9, article e128) doi: 10.1371/journal.ppat.0030128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Mei B, Budde AD, Leong SA. sid1, a gene initiating siderophore biosynthesis in Ustilago maydis: molecular characterization, regulation by iron, and role in phytopathogenicity. Proceedings of the National Academy of Sciences of the United States of America. 1993;90(3):903–907. doi: 10.1073/pnas.90.3.903. [DOI] [PMC free article] [PubMed] [Google Scholar]