

Abstract
Rapid polymerization of actin filament barbed ends generates protrusive forces at the cell edge, leading to cell migration. Two important regulators of free barbed ends, cofilin and Arp2/3, have been shown to work in synergy (net effect greater than additive). To explore this synergy, we model the dynamics of F-actin at the leading edge, motivated by data from EGF-stimulated mammary carcinoma cells. We study how synergy depends on the localized rates and relative timing of cofilin and Arp2/3 activation at the cell edge. The model incorporates diffusion of cofilin, membrane protrusion, F-actin capping, aging, and severing by cofilin and branch nucleation by Arp2/3 (but not G-actin recycling). In a well-mixed system, cofilin and Arp2/3 can each generate a large pulse of barbed ends on their own, but have little synergy; high synergy occurs only at low activation rates, when few barbed ends are produced. In the full spatially distributed model, both synergy and barbed-end production are significant over a range of activation rates. Furthermore, barbed-end production is greatest when Arp2/3 activation is delayed relative to cofilin. Our model supports a direct role for cofilin-mediated actin polymerization in stimulated cell migration, including chemotaxis and cancer invasion.
Introduction
In motile eukaryotic cells, actin filaments grow, push on the cell edge, and empower cell motility. New growing (barbed) ends of F-actin are formed by Arp2/3-mediated branching (1–4) and by cofilin severing of F-actin mother filaments. Barbed-end production by cofilin is observed experimentally (1,5–7), and complements cofilin’s other well-established role of recycling F-actin into monomers (8–10). Arp2/3 and cofilin have been shown to work in synergy to generate new actin barbed ends (11), motivating our model.
The control of actin filament dynamics by regulatory proteins such as cofilin and Arp2/3 is known to depend on the nucleotide-state (or age) of the actin. Arp2/3 forms a more stable branch on the side of ATP or ADP-Pi filament with a 10-fold increase in dissociation as the mother filament ages (12). Cofilin binds and severs almost 40× preferentially to ADP-F-actin (13). The barbed ends created by cofilin polymerize new ATP-F-actin, forming preferential binding sites for Arp2/3 complexes (12,14). Cofilin also accelerates the release of the phosphate (Pi) group and promotes debranching of filaments (14), as well as recycling actin filaments and replenishing the G-actin pool, roles we do not discuss here. The conversion from ATP to ADP F-actin state in vivo has been shown to occur within 10–30 s (3,13,15) (whereas, in vitro, Pi release occurs more slowly, at a timescale of minutes (16–18)).
Motivation for our model in this article stems from the experimentally observed actin-based protrusion after epidermal growth factor (EGF) stimulation in mammary carcinoma cells. In these cells, both cofilin and Arp2/3 are activated at the membrane. Active cofilin diffuses into the lamellipod (6), while active Arp2/3 is anchored to the WAVE2 complex (WASP-family verprolin homologous protein) at the leading edge (19–21). After EGF stimulation, cofilin is rapidly activated and released from the cell membrane. Here we focus on modeling early spatio-temporal actin dynamics after stimulation to characterize where, when, and how cofilin function could generate new barbed ends. The critical role of cofilin in regulating the spatiotemporal dynamics of actin cytoskeleton has been observed in a diverse array of processes from morphogenesis, receptor trafficking in synapses, and inflammation (22,23), further motivating our model.
In mammary carcinoma cells, a large peak of cofilin-dependent barbed ends is observed at 1 min after EGF stimulation (1,5,7). Given the ample availability of G-actin in the cytosol, growth of these barbed ends rapidly produces new F-actin, promoting Arp2/3 nucleation of a second peak of barbed ends ∼2 min later (24). To understand this synergy between cofilin and Arp2/3 (11), we use a mathematical model for actin dynamics at the leading edge of a motile cell. Specifically, we aim to address the following questions:
-
1.
How does cofilin-Arp2/3 synergy depend on biochemical parameters?
-
2.
How does the relative timing of Arp2/3 and cofilin stimulation affect synergy?
-
3.
How does spatial localization affect synergy and barbed-end production?
Mathematical Model
We consider a narrow transect of the lamellipod, a thin flat region ∼10 μm at the leading edge of the cell. We keep track of the length density of F-actin filaments and the corresponding barbed-end density, as well as the concentrations of active cofilin and Arp2/3 (see Fig. 1 for the geometry and schematic diagram). We discuss the model components first in a well-mixed null model and then in the fully spatio-temporal version.
Figure 1.
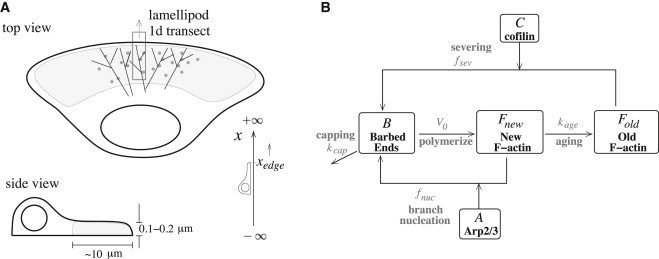
(A) Cell geometry showing a one-dimensional transect in top and side views (not to scale). Simulations span the region xL ≤ x ≤ xedge, where xedge is the cell edge and xL is 4 μm into the cell. (B) Schematic diagram of the model. Cofilin severs old ADP-F-actin and Arp2/3 binds to new ATP-F-actin to generate barbed ends. These lead to polymerization of new F-actin, which ages into old F-actin.
Well-mixed model
The assumptions and corresponding (well-mixed) model equations are as follows:
Assumption 1
ATP F-actin (length density Fnew) polymerizes from free barbed ends (number density B) at speed V0 (assumed constant) and ages into ADP F-actin, Fold, at rate kage. Jf is a small basal actin polymerization from other sources (e.g., formin (21,25)). Hence,
| (1) |
Assumption 2
We assume bulk turnover of old F-actin (rate kdeg), as in, for example (8,26,27)
| (2) |
This typical assumption replaces an older view of depolymerization at pointed ends.
Assumption 3
Cofilin, C, is transiently activated (step function JC(t)), and depleted by inactivation (rate kc), and by binding and severing old ADP-F-actin (rate fsev as in Tania et al. (28)). Hence,
| (3) |
We have shown that this nonlinear severing rate, possibly reflecting cooperativity (29), is needed for the large stimulus-induced amplification of barbed ends (28). Here, C0 is a typical cofilin concentration at which significant severing activity is observed and ℓ converts F-actin length density to a concentration. After severing, cofilin must be phosphorylated and then reactivated at the membrane on a slower timescale, a process not modeled here (but see Tania et al. (28)).
Assumption 4
Similarly, Arp2/3, A, is activated (step function JA(t)), and depleted by inactivation (rate ka), and by binding to ATP-F-actin, Fnew. This rate of branching, which nucleates barbed ends, is assumed proportional to Fnew and saturating in A (26). Thus,
| (4) |
At low Arp2/3, nucleation is roughly proportional to the product A × Fnew, whereas at high Arp2/3 it is proportional to binding sites along new F-actin.
Uncapped barbed ends, B, are generated by cofilin and Arp2/3 and disappear by capping (at rate kcap, assumed constant), yielding
| (5) |
The constant κ is used for unit conversion from concentration to barbed-end density.
Spatially extended model
The geometry of our model consists of a thin and narrow transect as illustrated in Fig. 1 A. The thickness of the lamellipod (<<1 μm) and the assumed narrow width of the transect imply that there are no significant gradients in either of these dimensions. Thus, we reduce the problem to a one-dimensional domain x ≤ xedge, where xedge denotes the location of the cell edge. We assume that both cofilin and Arp2/3 are locally activated at xedge. We model the transient behavior seen in Mouneimne et al. (24), where an initially static cell starts to move after an EGF stimulation. Formulating a consistent moving boundary problem, i.e., correctly posing the boundary conditions to satisfy conservation laws, is nontrivial (see below and the Supporting Material). Furthermore, the accelerating cell edge makes simulation more challenging than that with the steady-state motion (8,30).
The equation for uncapped barbed-end density (units of numbers/μm2) parallels Eq. 5, but with a term for motion toward the cell edge at the free-polymerization speed, V0, assumed constant:
| (6) |
Uncapped barbed ends accumulating at the cell edge become pushing barbed ends, Bp(t), that power cell protrusion by a thermal ratchet like mechanism (31). In the Supporting Material, we use conservation to derive the Bp(t) equation (and boundary conditions),
| (7) |
where Vmb is the cell-edge protrusion velocity. In turn, Vmb is determined by the pushing barbed ends. After Lacayo et al. (27), we assume that
| (8) |
Equation 8 means that motility is initiated only when barbed ends have built up sufficiently, and that Vmb saturates to the free-polymerization speed V0 at the high barbed-end density limit. The cell edge, xedge, moves (dragging cytosol with it) according to
| (9) |
Free cofilin C is thereby transported by bulk flow toward the edge at velocity Vmb. It also diffuses (diffusion coefficient Dc), and is depleted by inactivation (rate kc), and severing:
| (10) |
F-actin (units of length density μm/μm2 = n/μm) is tethered to the substrate and does not diffuse. It satisfies Eqs. 1 and 2, which are now partial differential equations for Fnew(x,t) and Fold(x,t). Active Arp2/3 is highly localized at and moves with the cell edge, bound to the WAVE2 complex. Arp2/3 is depleted as it nucleates barbed ends. Hence,
| (11) |
To avoid numerical issues, we implemented Eq. 11 with numerical diffusion (ϵAxx), where ϵ is so small that active Arp2/3 is restricted to a thin region well within 0.1 μm of the cell edge, a compromise to allow for our continuum approximation model. (We also tested even smaller ϵ, where simulations are prohibitively slow; see the Supporting Material.) Very close to the membrane, active Arp2/3 can bind to new F-actin and nucleate barbed ends, according to fnuc(A,Fnew) from Eq. 4. A basal rate of Arp2/3 inactivation, ka is included.
Local cofilin and Arp2/3 activation
At the cell membrane, cofilin is activated by PIP2 hydrolysis after stimulation (6,24,28). We model this boundary condition with a transient inward edge flux (with JC(t) = 0 for a resting cell). Arp2/3 is bound to the WAVE2 complex, and activated at the cell edge. Our boundary conditions are thus
| (12) |
(here, JC(t), Aedge(t) = 0 at rest, > 0 during stimulation). In summary, EGF stimulation is depicted as a transient flux of active cofilin released into the cell interior, and a significant elevation of Arp2/3 at the edge. The relative times at which cofilin and Arp2/3 are activated might not coincide. We later investigate the effect of possible delay between these times.
Summaries of variables and functions are given in Table 1. Parameter values (see Table S1 in the Supporting Material) were taken from the literature or previous models (8,26,30), with sensitivity analysis discussed below. See also details in the Supporting Material.
Table 1.
List of variables and functions used in the spatially extended model
| Definitions | Units | Equation | |
|---|---|---|---|
| Variables | |||
| x | Position | μm | |
| t | Time | s | |
| Fnew(x,t) | New F-actin filament length density | μm/μm2 | 1 |
| Fold(x,t) | Old F-actin filament length density | μm/μm2 | 2 |
| B(x,t) | Barbed-end density | numbers/μm2 | 6 |
| C(x,t) | Cofilin concentration | μM | 10 |
| A(x,t) | Arp2/3 concentration | μM | 11 |
| Bp(x,t) | Number of pushing barbed ends per μm of cell edge | numbers/μm | 7 |
| xedge(t) | Position of cell edge | μm | 9 |
| Functions | |||
| Fsev(C,Fold) | Cofilin severing function | μM/s | 3 |
| Fnuc(A,Fnew) | Arp2/3 nucleation function | μM/s | 4 |
| Vmb(Bp) | Membrane protrusion rate | μm/s | 8 |
| Bprod | Total barbed-end production per μm of cell edge | numbers/μm | 15 |
| S | Synergy between cofilin and Arp2/3 | — | 14 |
Results
Synergy in the well-mixed model
We first consider the well-mixed model (Eqs. 1–5) with basic parameter values (see Table S1) and resting/steady state as initial conditions. After a stimulus, the total number of barbed ends, Bprod, produced by cofilin and Arp2/3 integrated over time is
| (13) |
Barbed ends will be capped at rate kcap (Eq. 5). For numerical simulations, we computed up to 60 s poststimulus, after which there is no further severing or nucleation (see Fig. S1 in the Supporting Material). Given cofilin and Arp2/3 stimuli amplitudes JC and JA, synergy is defined as in Ichetovkin et al. (5) and DesMarais et al. (11),
| (14) |
where Bprod(X,Y) is total barbed ends generated by the given (cofilin, Arp2/3) stimuli. If cofilin and Arp2/3 act independently, then S ≈ 1, whereas significant synergy implies S > 1.
Varying the step function heights JC and JA during the 10 s stimulus, we find that cofilin and Arp2/3 can each generate a large pulse of barbed ends (Fig. 2 A). The barbed-end production curves have two regimes:
-
1.
A lower stimulus range with high sensitivity (a slight increase in activation leads to a much larger response), and
-
2.
A higher stimulus range with low sensitivity (additional input results in a very modest further increase in barbed-end production).
Figure 2.
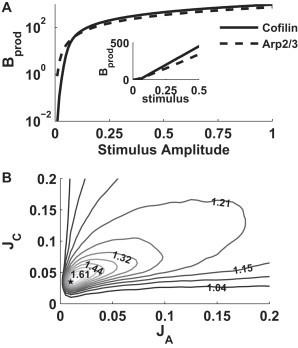
The well-mixed model (Eqs. 1–5). (A) Total number of barbed ends produced as a function of stimulus amplitude for cofilin alone (solid), and Arp2/3 alone (dashed). (Inset) Same plot on a linear scale. (B) Synergy S of cofilin and Arp2/3 as in Eq. 14. Maximum synergy (star) at (JC, JA) = (0.036, 0.012) μM/s. Parameter values as in Table S1 in the Supporting Material. Steady-state initial conditions: A(0) = 0, C(0) = 0, B(0) = 0, Fnew(0) = Jf/kage, and Fold = Jf/kdeg. Cofilin and Arp2/3 were activated simultaneously for 10 s.
In the presence of both cofilin and Arp2/3 (Fig. 2 B), a high synergy of ∼1.8 is only observed in the low stimulus range with high sensitivity. In this regime, cofilin and Arp2/3 produce very few barbed ends on their own but the total barbed-end production is very sensitive to additional stimulation, so that, by acting together, cofilin and Arp2/3 synergistically produce many more barbed ends. However, even with synergy, the level of barbed ends produced was still far too low (maximum value of <2/μm2 at maximal synergy, graph not shown) corresponding to a protrusion rate close to zero in the spatially extended model (see Eq. 8). Higher stimulation yields a low level of synergy (∼1.2 or less), much too low to account for in vitro experimental observation of 2× or higher synergy during simultaneous cofilin and Arp2/3 activation (5). We later show that this limitation is not observed in the spatially extended version of the model, highlighting the importance of spatial localization.
Results of the spatially extended model
Spatiotemporal dynamics in response to simultaneous cofilin and Arp2/3 activation are shown in Fig. 3 and Movie S1 (see the Supporting Material). Close to the cell edge, the concentrations of active cofilin and Arp2/3 increase rapidly within the first 5 s of stimulation, then remain relatively fixed until t = 10 s (see Movie S1) at the cell edge. During this period, active cofilin is elevated throughout the domain (4 μm). With ∼2 s delay, barbed ends accumulate (Fig. 3 C) with peak density at the cell edge. The pushing barbed ends, Bp, reach their maximal density at ∼8 s poststimulus. Edge protrusion, at a rate Vmb (Eq. 8 and Fig. 3 D), leaves behind F-actin (Fig. 3, A and B) whose density peaks ∼1 μm from the cell edge, consistent with recent data in Bravo-Cordero et al. (32). After the end of stimulation, cofilin and Arp2/3 decay, whereas F-actin polymerization continues for 10–20 s. Because capping eliminates barbed ends, the system gradually returns to its basal steady state.
Figure 3.
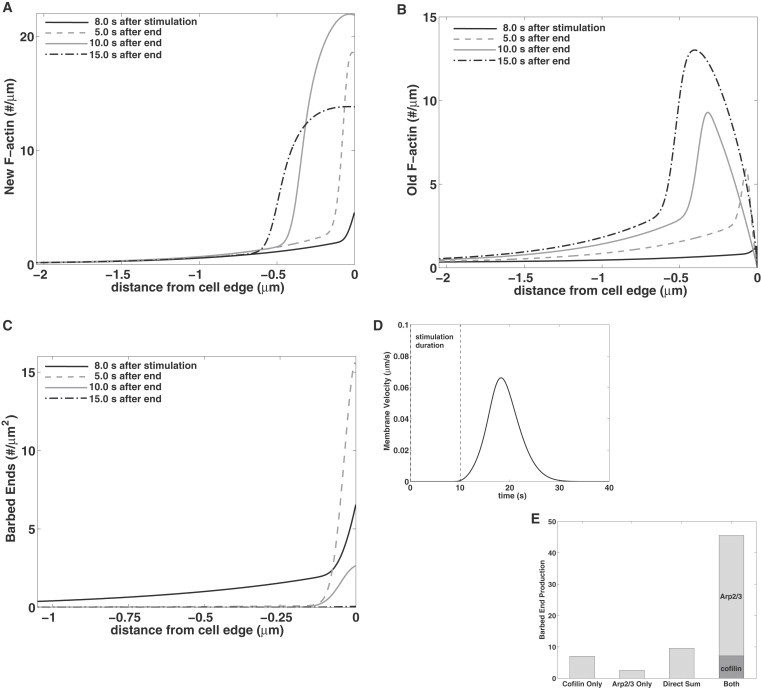
The spatial model with simultaneous activation of cofilin and Arp2/3 for 10 s (JC = 1.6 μM × μm/s, Aedge = 44 μM for 0.1 ≤ t ≤ 10.1 s; see boundary conditions in Eq. 12). (A–C) Profiles of new F-actin, and old F-actin, and barbed ends during stimulation (at 8 s) and at 5, 10, and 15 s after the end of stimulus. (D) The protrusion rate Vmb as a function of time. (E) Total barbed-end production obtained using cofilin alone; Arp2/3 alone; the expected barbed ends without synergy (direct sum); and in the presence of both. The barbed ends produced by cofilin (∫∫Fsevdx dt) and by Arp2/3 (∫∫Fnucdx dt) are shown.
In Fig. 3 E, we show total barbed ends that are produced by cofilin and Arp2/3 each acting alone, then the direct sum, and then the synergistic production. When both cofilin and Arp2/3 are present, their respective contribution is computed by integrating fsev and fnuc, respectively. Spatial distribution of severing (fsev) and nucleation (fnuc) rates over time are shown in Fig. S2. The spatial extent of cofilin (∼1 μm) severing is an order-of-magnitude larger than that of Arp2/3 nucleation in our model, though both peak at the cell edge. We find that ∼84% of total barbed ends are produced by Arp2/3 nucleation, with only ∼16% by cofilin. Importantly, cofilin primes the system by initially generating barbed ends from old filaments.
High synergy and large barbed ends peak in the spatial model
The total barbed-end production in the spatial model is computed by integrating over space and time,
| (15) |
Synergy is then computed according to Eq. 14 as before, with Aedge replacing JA in Eq. 14.
Barbed-end production in the presence of either cofilin or Arp2/3 alone is shown in Fig. 4 A. Although the Arp2/3 curve resembles that of the well-mixed system (Fig. 2 A), the cofilin production curve increases more gradually for low and mid-range cofilin stimulation. We attribute this to the fact that cofilin is activated at the cell edge, but severs old F-actin that is concentrated farther away (∼1 μm from the edge). There is a wide stimulus range over which barbed-end production is both sufficiently large and highly sensitive to additional stimulation. This then allows for simultaneously high synergy as well as large production of barbed ends as shown in Fig. 4 B.
Figure 4.
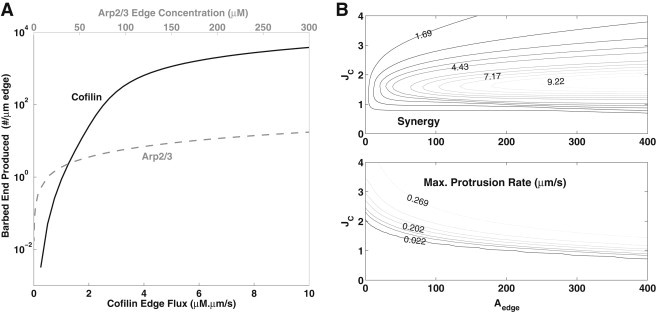
Barbed ends and synergy in the spatial model. (A) Total barbed ends (Eq. 15) produced by cofilin (solid) or Arp2/3 (gray) alone. (B) Synergy (top) and maximum protrusion rate Vmax (bottom) for varying cofilin flux, JC, and Arp2/3 edge concentration, Aedge (10 s simultaneous activation as in Fig. 3).
The dependence of synergy and maximal protrusion rate on both cofilin (JC) and Arp2/3 (Aedge) activation at the cell edge is shown in Fig. 4 B. High synergy (up to ∼9.9) occurs for 1 < JC < 3 μM × μm/s, provided Aedge is high (250 < Aedge < 400 μM). In the high synergy regime, a large pulse of barbed ends occurs, leading to fast protrusion (Vmb ∼ 0.28 μm/s at maximum synergy).
High synergy is obtained when barbed ends due to cofilin grow into new F-actin, which facilitates Arp2/3 nucleation. DesMarais et al. (11) found that the ratio of barbed ends produced by cofilin and by Arp2/3 (when each acts alone) is ∼2:1. This experimental result constrains the values of JC and Aedge to 1.6 < JC < 1.8 μM⋅μm/s and 40 < Aedge < 60 μM. We use this range in the rest of the article and find that this leads to model predictions that are most consistent with the experimental finding of DesMarais et al. (11). Within this range, a sufficiently high protrusion rate (0.05–0.2 μm/s), synergy of 4–5, and barbed-end production ratio of 2.5–3 (cofilin to Arp2/3 when each acts alone) were obtained (also see Fig. 3 E).
Relative timing of cofilin and Arp2/3 activation
Experiments on EGF stimulation of mammary carcinoma cells indicate that Arp2/3 activation occurs ∼10–20 s later than cofilin activation (11,24). The active cofilin is released from membrane lipid PIP2 after hydrolysis by PLC (6,33), whereas Arp2/3 is activated by the Cdc42-regulated WAVE2 (19,34,35), so distinct signaling pathways are at play. In the simulations so far, we assumed that cofilin and Arp2/3 are activated simultaneously. Now, we consider the effect of this relative Arp2/3 delay on our predictions.
In Fig. 5 A, we show how synergy changes as Arp2/3 activation is delayed relative to cofilin activation. During a delay τ, cofilin-generated barbed ends extend by V0τ, forming new filament density (V0τB) for Arp2/3 to bind. Capping eventually eliminates these barbed ends, resulting in the nonmonotonic relationship shown. Synergy (solid curve) nearly doubles from S = 4.7 (no delay) to a maximum of S = 7.1, when Arp2/3 activation is delayed by tarp ∼ 7.5 s. The maximal protrusion rate (dashed gray curve) increases by >50% over the same interval. This indicates that the ultimate output—cell protrusion—is affected by the relative timing of cofilin and Arp2/3 activation.
Figure 5.
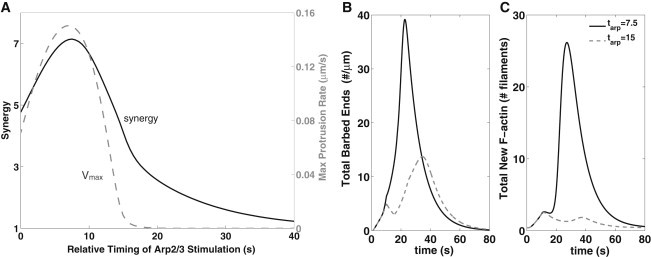
A delay in Arp2/3 activation affects cofilin-Arp2/3 synergy. (A) Synergy (solid) obtained as the relative timing of Arp2/3 activation is varied. The corresponding maximum protrusion rate is also shown (gray curve, axis on the right). (B and C) Comparison of barbed-end production when Arp2/3 is introduced at ta = 7.5 s (dashed gray curve) and at 15 s (gray curve), after cofilin activation. (B) Actin barbed ends (total barbed ends ∫B(x,t)dx + Bp(t)); (C) new F-actin (∫Fnewdx). (JC = 1.6 μM × μm/s; Aedge = 44 μM, as in Fig. 3; 10 s cofilin (0 < t < 10) and Arp2/3 stimuli with variation in the timing of Arp2/3 activation.)
In Fig. 5, B and C, we plot the total barbed ends and new F-actin over time, for two values of tarp. For tarp = 7.5 s, barbed ends created by cofilin severing have generated enough new F-actin to optimally prime the system for Arp2/3 action. As a result, we see a larger peak of barbed ends being generated, accompanied by a higher protrusion rate. For tarp = 15 s, some of that F-actin has already aged, so a smaller burst of barbed ends and lower protrusion rate is obtained. However, this yields two distinct peaks of barbed ends, as previously observed by Mouneimne et al. (24).
Filament protection by tropomyosin
Tropomyosin density increases from the cell edge inwards (36), and competes with cofilin for actin binding (37). We asked how this competition would affect our model results. To avoid significant expansion of the minimal model, we simply assumed that tropomyosin binding removes available cofilin binding sites on old F-actin. To do so, we modified the removal term, kdeg in Eq. 2 to kdeg(x) = kdeg – dT(x), with dT(x) a linear gradient, as in DesMarais et al. (36). We also included a class of tropomyosin-protected filaments, Ftm (details in the Supporting Material). We found that tropomyosin decreases barbed-end production by cofilin and restricts its activity more sharply, within ∼0.5 μm, toward the cell edge (see Fig. S3). Inclusion of tropomyosin does not qualitatively change our synergy results. We still obtain a cofilin barbed-end production curve with wide sensitive region, as before.
Binding to old versus new filaments
Recent data from Ti et al. (38) suggests that Arp2/3 binds to old (ADP-Pi and ADP) F-actin with similar kinetics, and higher affinity than to new (ATP) F-actin (although possibly without branching). To investigate how this would affect synergy, we modified the previous Arp2/3 binding term to
| (16) |
for 0 ≤ α ≤ 1 as the preferential binding of Arp2/3 to new (α ∼ 0) versus old (α ∼ 1) F-actin.
As shown in Fig. 6, α = 0 results in many barbed ends and a large protrusion rate. Larger α leads to lower Arp2/3 nucleation (and lower protrusion rate, dashed curve, Fig. 6 A). Larger α also lowers the synergy (solid curve, Fig. 6 A) and the peak of barbed ends produced (Fig. 6 B) but does not eliminate it: barbed ends created by cofilin still accelerate the substrate on which Arp2/3 can act. These results suggest that Arp2/3 binding to old F-actin has at most minor effect, because the narrow localization of Arp2/3 at the cell edge provides little overlap with old F-actin further into the cell. Overall, this agrees with results of a well-mixed model by Carlsson (39), who found Arp2/3-cofilin synergy only if Arp2/3 binds exclusively to new filaments.
Figure 6.
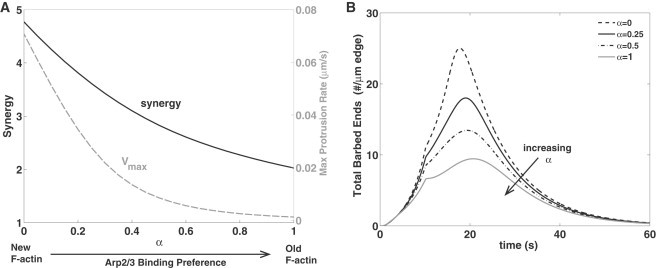
Effect of Arp2/3 binding preference to new ATP-F-actin versus old ADP-F-actin, depicted by α in Eq. 16. (A) Maximum protrusion rate Vmax (in gray dashed line) and synergy (in solid line) versus α. As in Fig. 3, JC = 1.6 μM × μm/s and Aedge = 44 μM, simultaneous cofilin and Arp2/3 activation for 10 s, but with Arp2/3 barbed-end production rate defined in Eq. 16. (B) Time course of total number of barbed ends (∫B(x,t)dx + Bp(t)) for α = 0, 0.25, 0.5, and 1.
We similarly considered the effect of cofilin binding (at low affinity) and severing of new F-actin, as in Blanchoin et al. (14) and Chan et al. (40). In the Supporting Material, we show that this model modification leads to scarcely any change: only a slight increase in barbed-end production and a drop in synergy. Note that we did not consider other putative cofilin roles such as accelerating phosphate release from F-actin, which promotes Arp2/3 dissociation and resultant filament debranching (14).
Parameter sensitivity
To check the sensitivity of our results, we varied key model parameters. Results are detailed extensively in the Supporting Material and briefly summarized here.
Filament aging
Some in vitro studies report slower rates of transition to ADP-actin (16–18), though other studies estimate the transition to ADP-actin to be 10 times faster in vivo, due to cofilin (3,13,14). We asked how varying the filament aging rate, kage, would affect our results. Briefly, when cofilin and Arp2/3 act together, barbed-end production increases monotonically with kage (see the Supporting Material). However, maximal synergy occurs when kage ≈ 0.12/s. Interestingly, this closely coincides with the value of kage reported in vivo.
Barbed-end capping and rate of growth
We found that faster capping (larger kcap) leads to a decrease in both barbed-end production and synergy whereas faster polymerization (higher V0) leads to a larger burst of barbed-end production as well as an increase in synergy. In the presence of cofilin and Arp2/3 alone, an increase in kcap leads to little change in barbed-end production but lead to a larger difference when cofilin and Arp2/3 are both present and working synergistically (see the Supporting Material for details).
Discussion
In this article, we explored synergy between cofilin and Arp2/3 in creating new barbed ends. Our model includes F-actin aging, capping, and severing by cofilin, as in Ditlev et al. (10) and Stuhrmann et al. (41) but not monomer recycling, nor the rare spontaneous nucleation of actin filaments from monomers. The model was motivated by data for transient responses of EGF-stimulated mammary carcinoma cells (11). There, cofilin is released from the membrane and its activity is focused in a zone ∼1 μm of the cell edge. Our model would apply (with suitable modifications) to other cells in which such release takes place, provided the cell-edge environment leads to a dominant filament-severing role of cofilin. Arp2/3 activity is restricted to a much smaller zone at the cell membrane (<0.1 μm). The maximum barbed-end density at the cell edge emerges as a model prediction that depends on biochemical rates, spatial localization of Arp2/3 and cofilin, and their relative timing.
Our model goes beyond Tania et al. (28), where a coarse spatial representation (two well-mixed compartments) was used. There, we considered multiple cofilin states in finer detail, but not the interaction with Arp2/3. Even so, we already noted that the peak of barbed ends depends on events occurring close to the cell membrane. Here we reduced the detail of the cofilin cycle, but included its interplay with Arp2/3 in both well-mixed and spatially distributed settings. In the well-mixed model, synergy is significant only at low cofilin and Arp2/3 activities and does not correlate well with barbed ends generated (Fig. 2). We note that correct formulation of the moving boundary problem for the cell edge, a challenging modeling issue, is an important feature of our model (see the Supporting Material).
Our spatially distributed model demonstrates a wide range of cofilin and Arp2/3 activity consistent with synergy and significant production of barbed ends. Synergy is accentuated if cofilin activity precedes Arp2/3 activity, because cofilin primes the system with new F-actin on which Arp2/3 can act. Hence, our model points to the fact that both spatial distribution and relative timing of cofilin and Arp2/3 activation are important determinants of synergy.
We carried out parameter sensitivity analysis, and modified several key model assumptions. For example, we showed that if cofilin severs new (ATP) F-actin then barbed-end production increases, but synergy decreases. In contrast, the binding of Arp2/3 to old F-actin has little effect, because the two have widely divergent spatial localizations.
We showed that by removing old F-actin binding sites, tropomyosin competes with and localizes cofilin activity (36,37) (see the Supporting Material). In this round of modeling, we did not, however, include a fully dynamic tropomyosin variable, nor its effect on Arp2/3 (42,43). Our preliminary results on tropomyosin suggest that this could be a fruitful future study.
We here assumed constant polymerization rate, V0 = 0.3 μm/s, for barbed ends away from the membrane, neglecting G-actin availability. Cofilin is known to also depolymerize actin and disassemble old filaments, allowing for G-actin monomers to be recycled (44–46). This monomer-recycling would increase available ATP-G-actin, speeding polymerization of ATP-F-actin, and enhancing the synergy between cofilin and Arp2/3. Using a realistic three-dimensional geometry, Novak et al. (9) found that F-actin disassembly at the rear and rapid polymerization at the front creates a concentration gradient that transports G-actin to the cell edge where it is being used up, thereby sustaining polymerization. G-actin recycling/sequestration by cofilin and other regulators (e.g., profilin and thymosin) has been previously considered in detail (8,10,47). Here, we focused on the complementary hypothesis of synergy via barbed-end production, which has yet to receive modeling attention. Incorporating G-actin into future versions of the model could allow us to study longer periods of activation where G-actin depletion becomes more significant.
Whereas some recent studies focused on the physical details of cofilin binding, individual filament geometry, filament bending (48), and severing (29,49,50) as well as its effect on nucleotide state of F-actin (50,51), here we focused solely on its role in creating new barbed ends by severing preexisting filaments (5,11,52). We have not considered the roles of other proteins such as Aip1 and coronin in regulating cofilin activity (53), nor profilin or thymosin that sequester G-actin or compete for binding. Finally, we simulated only low cofilin concentrations where severing, rather than de novo actin nucleation, occurs (54). Such omissions are limitations of our simplified continuum model, but keep its complexity manageable.
Our models make predictions about detailed spatial distributions of barbed-end and F-actin density that are experimentally testable. Although the cofilin-Arp2/3 synergy may well exist in a variety of cell types, the most extensive data sets are found in mammary carcinoma cells (11,24,36). The profiles we obtained agree qualitatively with spatial measurements in these articles. So far, such data were gathered at very coarse temporal resolution. Better time-resolution experiments would allow more direct quantitative comparisons. Experiments in which either Arp2/3 or cofilin are selectively inhibited would allow comparison to our predictions for synergy. Further, tests that manipulate monomer availability (hence V0), capping rates (hence kcap), or filament aging (kage) could be compared with predictions we have made above.
Acknowledgments
We are grateful to Eric Cytrynbaum, Jun Allard, and participants of a Banff International Research Station workshop, August 2011, for discussions.
This research was supported by a subcontract (to L.E.-K.) from the National Institutes of Health (grant No. R01 GM086882) to Anders Carlsson, Washington University, and by a Natural Sciences and Engineering Research Council of Canada discovery grant (to L.E.-K.). J.C. is funded by National Institutes of Health grant No. CA150344.
Supporting Material
References
- 1.Bailly M., Macaluso F., Condeelis J.S. Relationship between Arp2/3 complex and the barbed ends of actin filaments at the leading edge of carcinoma cells after epidermal growth factor stimulation. J. Cell Biol. 1999;145:331–345. doi: 10.1083/jcb.145.2.331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Mullins R.D., Heuser J.A., Pollard T.D. The interaction of Arp2/3 complex with actin: nucleation, high affinity pointed end capping, and formation of branching networks of filaments. Proc. Natl. Acad. Sci. USA. 1998;95:6181–6186. doi: 10.1073/pnas.95.11.6181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Pollard T.D., Blanchoin L., Mullins R.D. Molecular mechanisms controlling actin filament dynamics in nonmuscle cells. Annu. Rev. Biophys. Biomol. Struct. 2000;29:545–576. doi: 10.1146/annurev.biophys.29.1.545. [DOI] [PubMed] [Google Scholar]
- 4.Pollard T.D., Borisy G.G. Cellular motility driven by assembly and disassembly of actin filaments. Cell. 2003;112:453–465. doi: 10.1016/s0092-8674(03)00120-x. [DOI] [PubMed] [Google Scholar]
- 5.Ichetovkin I., Grant W., Condeelis J. Cofilin produces newly polymerized actin filaments that are preferred for dendritic nucleation by the Arp2/3 complex. Curr. Biol. 2002;12:79–84. doi: 10.1016/s0960-9822(01)00629-7. [DOI] [PubMed] [Google Scholar]
- 6.van Rheenen J., Song X., Condeelis J.S. EGF-induced PIP2 hydrolysis releases and activates cofilin locally in carcinoma cells. J. Cell Biol. 2007;179:1247–1259. doi: 10.1083/jcb.200706206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Wang W., Eddy R., Condeelis J. The cofilin pathway in breast cancer invasion and metastasis. Nat. Rev. Cancer. 2007;7:429–440. doi: 10.1038/nrc2148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Mogilner A., Edelstein-Keshet L. Regulation of actin dynamics in rapidly moving cells: a quantitative analysis. Biophys. J. 2002;83:1237–1258. doi: 10.1016/S0006-3495(02)73897-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Novak I.L., Slepchenko B.M., Mogilner A. Quantitative analysis of G-actin transport in motile cells. Biophys. J. 2008;95:1627–1638. doi: 10.1529/biophysj.108.130096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ditlev J.A., Vacanti N.M., Loew L.M. An open model of actin dendritic nucleation. Biophys. J. 2009;96:3529–3542. doi: 10.1016/j.bpj.2009.01.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.DesMarais V., Macaluso F., Bailly M. Synergistic interaction between the Arp2/3 complex and cofilin drives stimulated lamellipod extension. J. Cell Sci. 2004;117:3499–3510. doi: 10.1242/jcs.01211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Mahaffy R.E., Pollard T.D. Kinetics of the formation and dissociation of actin filament branches mediated by Arp2/3 complex. Biophys. J. 2006;91:3519–3528. doi: 10.1529/biophysj.106.080937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Blanchoin L., Pollard T.D. Mechanism of interaction of Acanthamoeba actophorin (ADF/Cofilin) with actin filaments. J. Biol. Chem. 1999;274:15538–15546. doi: 10.1074/jbc.274.22.15538. [DOI] [PubMed] [Google Scholar]
- 14.Blanchoin L., Pollard T.D., Mullins R.D. Interactions of ADF/cofilin, Arp2/3 complex, capping protein and profilin in remodeling of branched actin filament networks. Curr. Biol. 2000;10:1273–1282. doi: 10.1016/s0960-9822(00)00749-1. [DOI] [PubMed] [Google Scholar]
- 15.Blanchoin L., Pollard T.D. Hydrolysis of ATP by polymerized actin depends on the bound divalent cation but not profilin. Biochemistry. 2002;41:597–602. doi: 10.1021/bi011214b. [DOI] [PubMed] [Google Scholar]
- 16.Carlier M.F., Pantaloni D. Direct evidence for ADP-Pi-F-actin as the major intermediate in ATP-actin polymerization. Rate of dissociation of Pi from actin filaments. Biochemistry. 1986;25:7789–7792. doi: 10.1021/bi00372a001. [DOI] [PubMed] [Google Scholar]
- 17.Melki R., Fievez S., Carlier M.-F. Continuous monitoring of Pi release after nucleotide hydrolysis in actin or tubulin assembly using 2-amino-6-mercapto-7-methylpurine ribonucleoside and purine-nucleoside phosphorylase as an enzyme-linked assay. Biochemistry. 1996;35:12038–12045. doi: 10.1021/bi961325o. [DOI] [PubMed] [Google Scholar]
- 18.Jégou A., Niedermayer T., Romet-Lemonne G. Individual actin filaments in a microfluidic flow reveal the mechanism of ATP hydrolysis and give insight into the properties of profilin. PLoS Biol. 2011;9:e1001161. doi: 10.1371/journal.pbio.1001161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Sarmiento C., Wang W., Condeelis J. WASP family members and formin proteins coordinate regulation of cell protrusions in carcinoma cells. J. Cell Biol. 2008;180:1245–1260. doi: 10.1083/jcb.200708123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Lai F.P., Szczodrak M., Rottner K. Arp2/3 complex interactions and actin network turnover in lamellipodia. EMBO J. 2008;27:982–992. doi: 10.1038/emboj.2008.34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Welch M.D., Mullins R.D. Cellular control of actin nucleation. Annu. Rev. Cell Dev. Biol. 2002;18:247–288. doi: 10.1146/annurev.cellbio.18.040202.112133. [DOI] [PubMed] [Google Scholar]
- 22.Bamburg J.R., Wiggan O.P. ADF/cofilin and actin dynamics in disease. Trends Cell Biol. 2002;12:598–605. doi: 10.1016/s0962-8924(02)02404-2. [DOI] [PubMed] [Google Scholar]
- 23.Bravo-Cordero J.J., Magalhaes M.A.O., Condeelis J. Functions of cofilin in cell locomotion and invasion. Nat. Rev. Mol. Cell Biol. 2013;14:405–415. doi: 10.1038/nrm3609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Mouneimne G., Soon L., Condeelis J. Phospholipase C and cofilin are required for carcinoma cell directionality in response to EGF stimulation. J. Cell Biol. 2004;166:697–708. doi: 10.1083/jcb.200405156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Zigmond S.H. Formin-induced nucleation of actin filaments. Curr. Opin. Cell Biol. 2004;16:99–105. doi: 10.1016/j.ceb.2003.10.019. [DOI] [PubMed] [Google Scholar]
- 26.Marée A.F., Jilkine A., Edelstein-Keshet L. Polarization and movement of keratocytes: a multiscale modeling approach. Bull. Math. Biol. 2006;68:1169–1211. doi: 10.1007/s11538-006-9131-7. [DOI] [PubMed] [Google Scholar]
- 27.Lacayo C.I., Pincus Z., Theriot J.A. Emergence of large-scale cell morphology and movement from local actin filament growth dynamics. PLoS Biol. 2007;5:e233. doi: 10.1371/journal.pbio.0050233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Tania N., Prosk E., Edelstein-Keshet L. A temporal model of cofilin regulation and the early peak of actin barbed ends in invasive tumor cells. Biophys. J. 2011;100:1883–1892. doi: 10.1016/j.bpj.2011.02.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.De La Cruz E.M., Sept D. The kinetics of cooperative cofilin binding reveals two states of the cofilin-actin filament. Biophys. J. 2010;98:1893–1901. doi: 10.1016/j.bpj.2010.01.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Dawes A.T., Bard Ermentrout G., Edelstein-Keshet L. Actin filament branching and protrusion velocity in a simple 1D model of a motile cell. J. Theor. Biol. 2006;242:265–279. doi: 10.1016/j.jtbi.2006.02.017. [DOI] [PubMed] [Google Scholar]
- 31.Mogilner A., Oster G. Cell motility driven by actin polymerization. Biophys. J. 1996;71:3030–3045. doi: 10.1016/S0006-3495(96)79496-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Bravo-Cordero J.J., Sharma V.P., Hodgson L. Spatial regulation of RhoC activity defines protrusion formation in migrating cells. J. Cell Sci. 2013;126:3356–3369. doi: 10.1242/jcs.123547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.van Rheenen J., Condeelis J., Glogauer M. A common cofilin activity cycle in invasive tumor cells and inflammatory cells. J. Cell Sci. 2009;122:305–311. doi: 10.1242/jcs.031146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Millard T.H., Sharp S.J., Machesky L.M. Signaling to actin assembly via the WASP (Wiskott-Aldrich syndrome protein) family proteins and the Arp2/3 complex. Biochem. J. 2004;380:1–17. doi: 10.1042/BJ20040176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.El-Sibai M., Nalbant P., Backer J.M. Cdc42 is required for EGF-stimulated protrusion and motility in MTLn3 carcinoma cells. J. Cell Sci. 2007;120:3465–3474. doi: 10.1242/jcs.005942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.DesMarais V., Ichetovkin I., Hitchcock-DeGregori S.E. Spatial regulation of actin dynamics: a tropomyosin-free, actin-rich compartment at the leading edge. J. Cell Sci. 2002;115:4649–4660. doi: 10.1242/jcs.00147. [DOI] [PubMed] [Google Scholar]
- 37.Ono S., Ono K. Tropomyosin inhibits ADF/cofilin-dependent actin filament dynamics. J. Cell Biol. 2002;156:1065–1076. doi: 10.1083/jcb.200110013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Ti S.C., Jurgenson C.T., Pollard T.D. Structural and biochemical characterization of two binding sites for nucleation-promoting factor WASP-VCA on Arp2/3 complex. Proc. Natl. Acad. Sci. USA. 2011;108:E463–E471. doi: 10.1073/pnas.1100125108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Carlsson A.E. Stimulation of actin polymerization by filament severing. Biophys. J. 2006;90:413–422. doi: 10.1529/biophysj.105.069765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Chan C., Beltzner C.C., Pollard T.D. Cofilin dissociates Arp2/3 complex and branches from actin filaments. Curr. Biol. 2009;19:537–545. doi: 10.1016/j.cub.2009.02.060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Stuhrmann B., Huber F., Käs J. Robust organizational principles of protrusive biopolymer networks in migrating living cells. PLoS ONE. 2011;6:e14471. doi: 10.1371/journal.pone.0014471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Blanchoin L., Pollard T.D., Hitchcock-DeGregori S.E. Inhibition of the Arp2/3 complex-nucleated actin polymerization and branch formation by tropomyosin. Curr. Biol. 2001;11:1300–1304. doi: 10.1016/s0960-9822(01)00395-5. [DOI] [PubMed] [Google Scholar]
- 43.Cooper J.A. Actin dynamics: tropomyosin provides stability. Curr. Biol. 2002;12:R523–R525. doi: 10.1016/s0960-9822(02)01028-x. [DOI] [PubMed] [Google Scholar]
- 44.Carlier M.-F., Laurent V., Pantaloni D. Actin depolymerizing factor (ADF/cofilin) enhances the rate of filament turnover: implication in actin-based motility. J. Cell Biol. 1997;136:1307–1322. doi: 10.1083/jcb.136.6.1307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Bamburg J.R. Proteins of the ADF/cofilin family: essential regulators of actin dynamics. Annu. Rev. Cell Dev. Biol. 1999;15:185–230. doi: 10.1146/annurev.cellbio.15.1.185. [DOI] [PubMed] [Google Scholar]
- 46.Pantaloni D., Le Clainche C., Carlier M.-F. Mechanism of actin-based motility. Science. 2001;292:1502–1506. doi: 10.1126/science.1059975. [DOI] [PubMed] [Google Scholar]
- 47.Rubinstein B., Jacobson K., Mogilner A. Multiscale two-dimensional modeling of a motile simple-shaped cell. Multiscale Model Simul. 2005;3:413–439. doi: 10.1137/04060370X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Murrell M.P., Gardel M.L. F-actin buckling coordinates contractility and severing in a biomimetic actomyosin cortex. Proc. Natl. Acad. Sci. USA. 2012;109:20820–20825. doi: 10.1073/pnas.1214753109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Roland J., Berro J., Martiel J.-L. Stochastic severing of actin filaments by actin depolymerizing factor/cofilin controls the emergence of a steady dynamical regime. Biophys. J. 2008;94:2082–2094. doi: 10.1529/biophysj.107.121988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.McCullough B.R., Grintsevich E.E., De La Cruz E.M. Cofilin-linked changes in actin filament flexibility promote severing. Biophys. J. 2011;101:151–159. doi: 10.1016/j.bpj.2011.05.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Suarez C., Roland J., Blanchoin L. Cofilin tunes the nucleotide state of actin filaments and severs at bare and decorated segment boundaries. Curr. Biol. 2011;21:862–868. doi: 10.1016/j.cub.2011.03.064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Du J., Frieden C. Kinetic studies on the effect of yeast cofilin on yeast actin polymerization. Biochemistry. 1998;37:13276–13284. doi: 10.1021/bi981117r. [DOI] [PubMed] [Google Scholar]
- 53.Kueh H.Y., Charras G.T., Brieher W.M. Actin disassembly by cofilin, coronin, and Aip1 occurs in bursts and is inhibited by barbed-end cappers. J. Cell Biol. 2008;182:341–353. doi: 10.1083/jcb.200801027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Andrianantoandro E., Pollard T.D. Mechanism of actin filament turnover by severing and nucleation at different concentrations of ADF/cofilin. Mol. Cell. 2006;24:13–23. doi: 10.1016/j.molcel.2006.08.006. [DOI] [PubMed] [Google Scholar]
- 55.Svitkina T.M., Borisy G.G. Arp2/3 complex and actin depolymerizing factor/cofilin in dendritic organization and treadmilling of actin filament array in lamellipodia. J. Cell Biol. 1999;145:1009–1026. doi: 10.1083/jcb.145.5.1009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.De La Cruz E.M., Mandinova A., Pollard T.D. Polymerization and structure of nucleotide-free actin filaments. J. Mol. Biol. 2000;295:517–526. doi: 10.1006/jmbi.1999.3390. [DOI] [PubMed] [Google Scholar]
- 57.Nishida E., Maekawa S., Sakai H. Cofilin, a protein in porcine brain that binds to actin filaments and inhibits their interactions with myosin and tropomyosin. Biochemistry. 1984;23:5307–5313. doi: 10.1021/bi00317a032. [DOI] [PubMed] [Google Scholar]
- 58.Abraham V.C., Krishnamurthi V., Lanni F. The actin-based nanomachine at the leading edge of migrating cells. Biophys. J. 1999;77:1721–1732. doi: 10.1016/S0006-3495(99)77018-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Wegner A., Ruhnau K. Rate of binding of tropomyosin to actin filaments. Biochemistry. 1988;27:6994–7000. doi: 10.1021/bi00418a049. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.