

Abstract
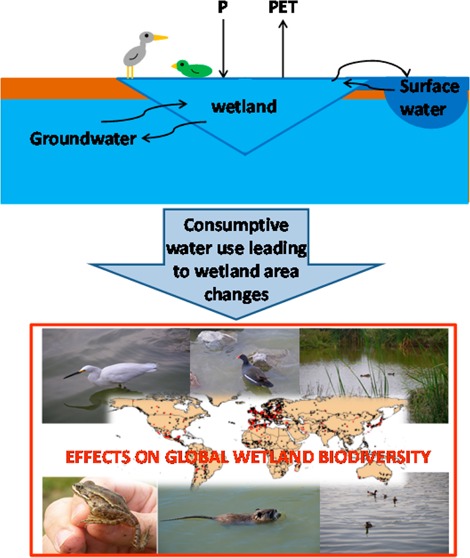
Wetlands are complex ecosystems that harbor a large diversity of species. Wetlands are among the most threatened ecosystems on our planet, due to human influences such as conversion and drainage. We assessed impacts from water consumption on the species richness of waterbirds, nonresidential birds, water-dependent mammals, reptiles and amphibians in wetlands, considering a larger number of taxa than previous life cycle impact assessment methods. Effect factors (EF) were derived for 1184 wetlands of international importance. EFs quantify the number of global species-equivalents lost per m2 of wetland area loss. Vulnerability and range size of species were included to reflect conservation values. Further, we derived spatially explicit characterization factors (CFs) that distinguish between surface water and groundwater consumption. All relevant watershed areas that are contributing to feeding the respective wetlands were determined for CF applications. In an example of rose production, we compared damages of water consumption in Kenya and The Netherlands. In both cases, the impact was largest for waterbirds. The total impact from water consumption in Kenya was 67 times larger than in The Netherlands, due to larger species richness and species’ vulnerability in Kenya, as well as more arid conditions and larger amounts of water consumed.
Introduction
Wetlands cover an area larger than 1280 million hectares worldwide1 and are among the most complex ecosystems in the world, due to a combination of different aquatic and terrestrial conditions. Many species have adapted to these mosaic ecosystems, leading to high varieties of all major groups of animals and plants.2 Wetlands can contribute massively to biodiversity within a landscape, often exhibiting high levels of alpha and beta diversity (i.e., species diversity and turnover between habitats).3 They are frequently used by migratory birds for resting and can be important drinking water sources, especially in semiarid and arid regions.
At the same time, wetlands are among the most threatened ecosystems on our planet.4 They are degraded and converted to human uses more rapidly than any other ecosystem, and the status of freshwater species is deteriorating faster than for other species.1,5 Globally, more than 50% of wetland areas were lost during the 20th century,1 mostly due to conversion and drainage.6 Since wetlands are essentially characterized by hydrologic conditions, changes in water volumes and timing of flows are major threats.7
Life cycle assessment (LCA) is a methodology for assessing the total environmental impacts of a product or service through its entire life cycle.8 LCA has rapidly developed over the last years, and life cycle impact assessment (LCIA) methods have started to include impacts from water and land use. LCIA methods are available globally for assessing the impact of water use on ecosystems in general,9,10 for the impact of surface water consumption on aquatic ecosystems (fish species) below 42° North,11 and regionally for impacts of groundwater use on plant species in The Netherlands.12 Yet, no global methodology currently exists in LCA to determine the effects of surface and groundwater consumption specifically on wetland ecosystems. Furthermore, impacts on ecosystems and biodiversity are commonly calculated in PDF (potentially disappeared fraction of species).10−15 However, PDF does not account for absolute variations in species richness. The same relative impact (e.g., a PDF of 0.5) is considered equivalent in a species poor ecosystem and a species rich ecosystem, although in the latter case more species are lost. Moreover, PDF does not take vulnerabilities and distribution ranges of species into consideration, treating all species equally whether critically threatened and endemic or widespread and common. For a review of existing practices and shortcomings in the assessment of biodiversity in LCA, see ref (16). The aim of this paper is to develop a new approach for quantifying impacts on biodiversity due to anthropogenic water consumption. In this approach, effect factors (EF) measure the absolute loss of species due to wetland area loss, including the species’ vulnerability. The existing fate factors17 that quantify the loss of wetland area due to water consumption are combined with the EFs in order to calculate characterization factors.
Materials and Methods
Biodiversity Data and Maps
We took into account all inland wetlands which were classified under the Ramsar convention on 17 August 2012 (1184 wetlands).18 73% of these wetlands were considered to be important for waterbirds for different life stages, and 67% were important for birds in general. 25%, 26% and 49% were important for amphibians, reptiles and mammals, respectively (Supporting Information (SI), section S1). Bird distribution data were available from BirdLife & NatureServe,19 data for amphibians and reptiles were from IUCN20−22 (see SI S2 for data sources and species numbers). For each species, map-files of the individual geographic extent of its distribution were available, including information about the presence, origin and season. Waterbirds were chosen for their obvious connection with water and were defined as birds whose primary habitat was, according to BirdLife, “wetland (inland)” or “artificial landscapes (aquatic)”.23 Residential birds whose habitat was not “wetland (inland)” or “artificial landscapes (aquatic)” were excluded since there is limited connection to wetlands and they are therefore considered irrelevant for estimating species loss in wetlands. Nonresidential birds (excluding seasonal category “resident”) were chosen because among them are migratory birds, which require staging and resting grounds during their migration. We assumed that wetland area loss would be a severe drawback for them along their migration routes. Nonresidential waterbirds are included in the waterbird category to avoid double counting. We included all amphibian species and those reptile species whose habitat was defined from IUCN as “wetlands (inland)”20 and also contained data. For waterbirds, reptiles and amphibians all seasonal categories (SI, S3) were included. The origin of species, that is, whether they are native or introduced was not considered when calculating current species richness values. With the software Matlab,24 species with presence categories “extant”, “probably extant” and “possibly extant” (see SI, S3) were identified, and the maps were transformed to raster files with a 0.05 decimal degree resolution with input datum WGS84. These were added up, resulting in global species richness maps for waterbirds, nonresidential birds, reptiles and amphibians.
For mammals, habitat suitability models are available25 and were used to refine their IUCN range. We considered only water-dependent mammals for calculating species-richness maps with 5 km resolution using the WGS84 datum. Other taxa (e.g., plants, fish) are not considered.
Calculation of Effect Factors
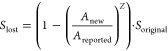
We assume that the aim of biodiversity assessment in LCA is to quantify and minimize the risk of global extinction of species of different taxa. We developed effect factors (EF) that quantify the contribution to potential global extinction of species due to a loss in wetland area that is caused by water consumption. The EF combines three parts based on global maps: (1) potential species loss, (2) vulnerability of present species communities, and (3) habitat loss risk (graphical summary in SI, S9). The potential species loss in numbers of species-equivalents lost (Slost) per area changed is derived from the species-area relationship that has been used in LCA before,13,26,27 based on an original area Areported and a new area Anew (both from ref (17)), as well as the original species richness Soriginal from global species richness maps (eq 1, further explanations in SI, S8).
 |
1 |
The exponent z is the slope of the species-area relationship and was derived from Drakare et al.28 for birds (0.37), mammals (0.34), amphibians (0.2), and reptiles (0.33) as described in SI (S8). Soriginal was derived from current species richness maps (SI S3–S6) and therefore does not represent pristine species richness, but is appropriate because we were also using current wetland area data as a reference.
The vulnerability of species communities is quantified with a vulnerability score (VS) as an indicator for global extinction risk. VS is a function of the area of the extent of occurrence (EOO, encompassing the outermost geographic limits of all areas where a species occurs)29 as a predictor for the susceptibility to anthropogenic disturbance (because species with a small range are intrinsically rare) and a threat level (TL) indicating already occurring threats. VS is calculated as global maps for each species i in taxon p, and each pixel j (0.05° × 0.05°, for mammals 5 km × 5 km) as the area of the respective pixel (EOOi,j) where species i occurs divided by the total EOO of the species and multiplied with TLi,j (eq 2). The EOO may include discontinuous areas and areas which may be unsuitable as habitat.30 Thus, it is larger than the actual area of occupancy of a species. TL represents discrete values ranging from 1 to 5 on a linear scale (1-least concern, 2-near threatened, 3-vulnerable, 4-endangered, 5-critically endangered) from the IUCN Red List of threatened species.31 Total VSp,j of each taxon p per pixel is obtained by summing all values for all species i which occur in pixel j and dividing by the number of species of the taxon present in pixel j (Sp,j, eq 2).
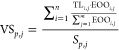 |
2 |
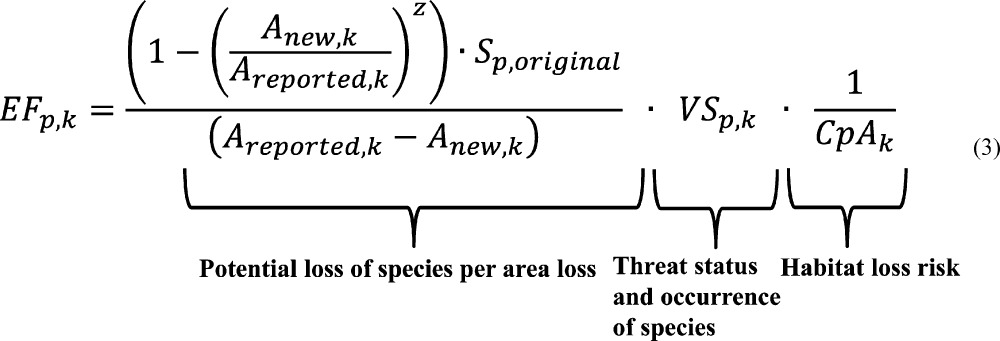
As third part for the EF calculation the habitat loss risk index (CpA) was calculated on a global map (SI, S6). CpA is the weighted and scaled waterbody count per area, which was used as a weighting factor for the density of the network of waterbodies (as proxy of habitat rarity in the region). It accounts for the number of river sections (one point per section)32 and the number of data points from the global lakes and wetland database33 in each subwatershed.32 This was divided by the area of subwatersheds and weighted with an aridity index (precipitation P(34)/potential evapotranspiration PET35), resulting in an index per pixel. The aridity index was included because in wet regions with relatively high P and low PET, wetlands are less relevant since alternative and temporary water supplies are more frequent. The CpA was scaled, so that 1 was the global maximum value in order to have an index that is relative to the smallest habitat loss risk. This led to lower values in more arid regions, that is, the habitat type “wetland” was scarcer and thus more critical. Thus, the reciprocal of CpA is used in the EF calculation (eq 3).
 |
3 |
The aforementioned parts are combined to one formula for calculating the effect factor (EF) for each wetland k and each taxon p. The unit of the EF is species-eq/m2, that is, global species-equivalents lost per square meter of wetland lost. For wetlands which are smaller than a pixel, parameters Soriginal, VS, and CpA were extracted from global maps. In case the wetland was larger than one pixel, the parameter values were extracted from all relevant pixels and averaged over the Ramsar or waterbody area (Areported) of wetland k. Anew is the calculated new area after water consumption (calculated as described in ref17).
Calculation of Characterization Factors
Characterization factors (CFs) were calculated as a combination of effect (EF) and fate factor (FF, relating water consumption to wetland area loss)17 (eq 4) for each taxon p and for each wetland k. The unit of the CF is species-eq·yr/m3 water consumed and shows the loss of species-eq. due to water consumption.
| 4 |
For each SW-fed wetland, we assumed that water consumption upstream would reduce the inflow into the wetland and thus cause biodiversity damage. This upstream area was determined in Matlab24 by selecting, on a hydrologically corrected digital elevation model (DEM)36 (resolution 0.05° × 0.05°), the parts of a watershed above and at the same altitude as the center of the wetland. Isolated areas with no connection to the wetland were removed from this selection. Where catchments of different wetlands overlapped, CFs were summed (SI Figure S22). Therefore, consuming water in the upper part of a watershed is potentially worse than at the mouth, since consumed water impacts all wetlands located downstream but has no impact on upstream areas.
For GW-fed wetlands CFs were applicable on the respective area of relevance (AoR). The AoR surrounds the wetland and is the area from which water infiltrates into the wetland (see ref (17) for further explanations). Due to geohydrological conditions, these conceptual areas can be large, and it is possible that the whole watershed is regarded as the AoR. For some cases this is realistic, for example, in the Great Artesian Basin in Australia, where pumping of water reduces spring flows which are large distances away.37 Due to a lack of good global maps of aquifer presence, we used the surface watershed borders to define the maximal size of AoRs. In areas with overlapping CFs from different GW-fed wetlands, values were summed since multiple wetlands are influenced if pumping occurred in that region (SI Figure S23).
Sensitivity Analysis
For the sensitivity analysis of EFs of birds, amphibians and reptiles, the presence category “possibly extinct” (category 4) was included in the species richness maps, since the respective species may still occur in very low numbers in those areas. Furthermore, high number of potentially extinct species might be an indicator of increased vulnerability of the region.
Additionally, the Ramsar area was changed to the reported waterbody area (as done in the FF calculations)17 in order to test the sensitivity of EFs and CFs toward area differences. As discussed in ref (17), the aim of the suggested CF is to take complete wetland areas into account and not only open waterbodies. Some wetlands (water-logged soil, waterbodies overgrown with vegetation) are invisible on satellite pictures, making the Ramsar area, which takes into account invisible waterbodies important to biodiversity, more suitable as the base area. For water-dependent mammals, extent of suitable habitat was used as a proxy for area of occupancy (AOO). The AOO is the area within the EOO where species actually occur.38 In order to test the sensitivity of the EF for water-dependent mammals, we calculated it once with EOO and once with AOO. The influence of other relevant parameters from the FFs,17 such as amount of water consumed, hydraulic conductivity, water depth, and SW flow volumes, were integrated into the sensitivity analysis of the CFs.
We also checked using the Spearman’s rank-order correlation whether there were correlations between CpA, S, and VS and how they correlate to the EF. Since both FF and CpA contain information about precipitation and potential evapotranspiration, we also tested their correlation.
For each taxon, we calculated CFs with the unit PDF·yr/m3 in order to compare them to the values with species-eq·yr/m3. For calculating CFs with PDF units, we omitted VS and CpA from the EF calculation.
Application Example
We calculated the impact from water consumption for the production of a bunch of 10 roses in Kenya and in The Netherlands. Roses are the dominant cut flower in Kenya, accounting for almost 88% of cultivation.39 One of the largest production areas (1911 ha in 2006)40 is close to lake Naivasha (SW-fed), which is listed under the Ramsar Convention as being internationally important. One average rose (25 g) consumed 4.1 L of irrigation water: 3.4 L surface water, and 0.7 L groundwater.40 Rose production in The Netherlands (region of Bleiswijk, South Holland) was assumed to require only 1.6 L/stem irrigation water from surface water, in addition to 1.6 L/stem precipitation.41,42 CFSW and CFGW were extracted from the CF-maps at the location of lake Naivasha and Bleiswijk. This example serves as an illustration and does not aim at calculating a full LCA.
Results
Biodiversity
Maps of species richness and VS for all taxa are shown in SI, S3–S6 for presence categories 1–3 and 1–4. Maximum species richness was 284, 112, 27, 134, 15 for waterbirds, nonresidential birds, reptiles, amphibians, and water-dependent mammals, respectively. Differences between bird richness maps and VS with presence 1–3 and 1–4 were zero or very small. The maximal differences in wetland regions were five species for amphibians, three for waterbirds and one for nonresidential birds, reptiles and mammals. For reptiles, mammals, and amphibians, maximum species richness and overall global distribution is smaller than for birds, and, in contrast to birds, they were not present in all wetlands (SI, S10).
Effect Factors
EFs of wetlands vary over many orders of magnitude, underlining the importance of spatial differentiation in LCIA of water consumption (see individual EFs in SI Excel sheet). The mean, minimal and maximal EFs for all wetlands are displayed in Table 1, and the averages for SW and GW are often similar. The dominant EFs were in the majority of cases calculated for waterbirds, being often the most species-rich taxon, closely followed by nonresidential birds and amphibians. EFs for reptiles and water-dependent mammals were often the smallest, since species richness was often the smallest of all taxa. EFs for reptiles, amphibians and water-dependent mammals may be zero, since not all wetlands harbor these taxa. 121, 168, and 43 SW-fed wetlands show absences of mammals, reptiles and amphibians, respectively. In groundwater-fed wetlands, there are no mammals, reptiles and amphibians in 28, 25, and 6 wetlands, respectively. Reasons for the difference between EFs of SW-fed and GW-fed wetlands are related to the different locations of the wetlands, as well as the different changes in area that are influencing the nonlinear species-area relationship.
Table 1. Summary of Non-Zero Effect Factors (EF) [Species-eq/m2] and Characterization Factors (CF) [Species-eq·yr/m3] for Waterbirds, Non-Residential Birds, Amphibians, Reptiles (All with Presence Categories 1–3) and Water-Dependent Mammals (Based on EOO), Based on the Area of the Ramsar Sitesa.
| EF [species-eq/m2] |
CF [species-eq·yr/m3] |
|||
|---|---|---|---|---|
| SW | GW | SW | GW | |
| waterbirds min | 3.2 × 10–14 | 3.1 × 10–13 | 1.6 × 10–15 | 5.0 × 10–15 |
| waterbirds max | 1.6 × 10–05 | 1.8 × 10–06 | 1.1 × 10–05 | 2.1 × 10–07 |
| waterbirds mean | 3.6 × 10–08 | 1.8 × 10–08 | 1.2 × 10–08 | 4.2 × 10–09 |
| CV | 19 | 8 | 28 | 5 |
| nonresidents min | 1.5 × 10–15 | 1.4 × 10–13 | 5.3 × 10–17 | 6.9 × 10–15 |
| nonresidents max | 9.1 × 10–06 | 1.5 × 10–06 | 2.2 × 10–07 | 3.0 × 10–06 |
| nonresidents mean | 1.9 × 10–08 | 1.6 × 10–08 | 1.2 × 10–09 | 2.1 × 10–08 |
| CV | 18 | 7 | 8 | 11 |
| water-dep. mammals min | 8.9 × 10–16 | 5.0 × 10–15 | 3.5 × 10–17 | 1.2 × 10–16 |
| water-dep. mammals max | 2.0 × 10–06 | 2.0 × 10–07 | 3.3 × 10–08 | 5.3 × 10–08 |
| water-dep. mammals mean | 2.4 × 10–09 | 2.5 × 10–09 | 1.3 × 10–10 | 4.8 × 10–10 |
| CV | 26 | 7 | 10 | 9 |
| wetland reptiles min | 2.74 × 10–16 | 8.54 × 10–15 | 1.22 × 10–17 | 4.63 × 10–17 |
| wetland reptiles max | 4.37 × 10–07 | 1.13 × 10–05 | 5.11 × 10–07 | 9.53 × 10–07 |
| wetland reptiles mean | 8.69 × 10–10 | 7.69 × 10–08 | 6.18 × 10–10 | 7.58 × 10–09 |
| CV | 16 | 12 | 26 | 10 |
| amphibians min | 1.17 × 10–15 | 5.32 × 10–15 | 4.38 × 10–17 | 5.82 × 10–16 |
| amphibians max | 6.47 × 10–05 | 7.71 × 10–07 | 4.56 × 10–05 | 9.56 × 10–08 |
| amphibians mean | 8.50 × 10–08 | 1.23 × 10–08 | 4.58 × 10–08 | 1.75 × 10–09 |
| CV | 24 | 5 | 31 | 5 |
| combined taxa min | 1.58 × 10–13 | 9.34 × 10–13 | 2.33 × 10–15 | 1.57 × 10–14 |
| combined taxa max | 8.07 × 10–05 | 1.15 × 10–05 | 5.68 × 10–05 | 3.27 × 10–06 |
| combined taxa mean | 1.43 × 10–07 | 1.26 × 10–07 | 6.01 × 10–08 | 3.51 × 10–08 |
| CV | 19 | 8 | 29 | 8 |
The last lines show combined (summed) EFs and CFs, assuming the same weight for all species. CV is the coefficient of variation.
Spearman’s rank-order correlations ρ (SI Table S10) between S, VS, and EF are largest for amphibians and reptiles, indicating that VS has a large influence on EF. CpA is always negatively correlated to EF. Most mutual correlations between CpA and VS, as well as CpA and S were low or nonexistent, illustrating that they are complementary to each other. Correlations between S and VS were large, since VS is related to the number of species present and assigns them a weight according to their vulnerability. Histograms for species richness show that waterbirds are usually the most species-rich taxon and reptiles the taxon with lowest richness (SI Figure S37).
The median difference between EFs calculated with Ramsar area or waterbody area was a factor of 1.67 for all taxa in SW-fed and GW-fed wetlands (SI, S11). Differences in S, VS, and CpA were mostly low (less than 2%), since in many cases there was no or little difference between S, VS, and CpA on Ramsar or waterbody area. Nevertheless, the underlying area was very important due to the nonlinearity of the species-area relationship.
Characterization Factors
Characterization factors (CFs) were calculated for all wetlands (see SI Excel sheet) and result in global CF maps (Figure 1). Impacts for waterbirds were mostly larger than for other taxa. In the majority of the cases, magnitudes of impacts on nonresidential birds were next, closely followed by amphibians and then mammals and reptiles. Where CFnonresidential birds was larger than CFwaterbirds, it was mostly due to higher vulnerability scores (VS) and only in few cases due to higher species richness (S). CFmammals was never larger than CFwaterbirds and only in few cases larger than CFnonresidential birds, uniquely due to larger VS. CFamphibians and CFreptiles were larger than CFwaterbirds in 86 wetlands and 2 wetlands, respectively (out of 1184), and this was always accompanied by a larger VS. Not all wetlands had CFs for mammals, reptiles or amphibians (CF = 0) and therefore the coverage of CFs was smaller than that for the bird species (Figure 1). CFSWs covered 153 out of 238 major watersheds for birds, containing 76% of global land surface (LS). Analogously, CFSWs for mammals, reptiles and amphibians covered 135 (64% LS), 129 (61% LS) and 146 (67% LS) major watersheds, respectively. CFGWs of birds, mammals, reptiles, and amphibians covered 70 (37% LS), 57 (30% LS), 60 (28% LS), and 68 (35% LS) major watersheds, respectively.
Figure 1.

Global maps of CFs, with indications of wetland type (SW, surface water-fed, GW – groundwater-fed) and taxon. White areas are areas where no impact on a Ramsar wetland is perceived. Base map with country boundaries adapted from ref (43). (A) CFs for waterbirds and SW consumption in SW-fed wetlands. (B) CFs for waterbirds and GW consumption in GW-fed wetlands. (C) CFs for nonresidential birds and SW consumption in SW-fed wetlands. (D) CFs for nonresidential birds and GW consumption in groundwater-fed wetlands. (E) CFs for water-dependent mammals and SW consumption in SW-fed wetlands. (F) CFs for water-dependent mammals and GW consumption in GW-fed wetlands. (G) CFs for wetland reptiles and SW consumption in SW-fed wetlands. (H) CFs for wetland reptiles and GW consumption in GW-fed wetlands. (J) CFs for amphibians and SW consumption in SW-fed wetlands. (K) CFs for amphibians and GW consumption in GW-fed wetlands.( L) CFs for all taxa combined and SW consumption in SW-fed wetlands. (M) CFs for all taxa combined and GW consumption in GW-fed wetlands. Note that the CFs for groundwater-fed wetlands should only be used for marginal changes or cases studies with good data coverage, due to the uncertainty in the FFs.17
The variation of CFs is shown in Table 1. The large sensitivity of CFs to changes in underlying area (Ramsar area or waterbody area) was determined by the sensitivity of FFs17 and by the nonlinearity of the species-area relationship for the EFs (SI, S11). It is suggested to use Ramsar-area based CFs, since these underlying areas contain less uncertainty (see also ref (17)), even though the larger areas are not a conservative assumption.
Additionally, the CFs are sensitive to the amount of water consumed, surface water flows (SW-fed wetlands) and hydraulic conductivity (GW-fed wetlands). Including species which were “possibly extinct” or changing from EOO to AOO (mammals) had little influence on most CFs (SI Figure S39 and S40). We found no or small correlation between CpA and FF for SW and GW wetlands. Spearman’s rank order coefficients between CFSW and CFGW of different taxa were large (ρ > 0.5) between most taxa (SI Table S9).
CFSW and CFGW with the unit PDF·yr/m3 were strongly correlated to CFs (in species eq·yr/m3) of waterbirds and amphibians, and CFGW was also correlated to nonresidential birds. An example for the differences between CF in PDF and in species-eq. is given in SI S12, showing that species richness, vulnerability and CpA matter.
Application Example
We calculated the impact of water consumption during the cultivation of 10 roses in Kenya and The Netherlands (Table 2). The largest impact was caused in both cases for waterbirds. In Kenya, impacts from SW consumption were for all taxa larger than those from GW consumption. The impact in The Netherlands is only from SW consumption, since no groundwater is consumed and 50% of the required water for growing roses is from precipitation. Assuming the same weight for all taxa, we added the CFs for all taxa. In total, the impact in Kenya is 67 times larger than the one in The Netherlands due to larger species richness, larger VS, smaller CpA and a larger amount of irrigation water required (4.1 L/stem vs 1.6 L/stem).
Table 2. Calculated Impacts from Water Consumption for the Production of a Bunch of 10 Roses in Kenya and the Netherlandsa.
| Kenya
[species-eq·yr] |
The Netherlands [species-eq·yr] | ||
|---|---|---|---|
| species | SW | GW | SW |
| waterbirds | 4.75 × 10–12 | 1.29 × 10–14 | 3.30 × 10–14 |
| nonresidential birds | 2.80 × 10–13 | 8.65 × 10–16 | 5.23 × 10–14 |
| mammals | 7.46 × 10–14 | 2.20 × 10–16 | 2.14 × 10–15 |
| reptiles | 3.87 × 10–15 | 1.14 × 10–17 | 3.06 × 10–16 |
| amphibans | 1.06 × 10–12 | 9.25 × 10–15 | 4.79 × 10–15 |
| total | 6.17 × 10–12 | 2.33 × 10–14 | 9.25 × 10–14 |
Impacts are reported for each taxon separately, before summing them to total impacts due to SW consumption or GW consumption. There is no GW consumption at the case study site in the Netherlands.
We refrained from calculating a complete LCA because our units of biodiversity loss are not compatible to existing methods, which are mostly based on PDF.
Discussion
Effect Factors
The inclusion of ecosystem rarity (through CpA) as well as vulnerability of biodiversity (with VS) is a considerable improvement in comparison to existing LCIA methodologies for the assessment of ecosystem damages. A similar approach was presented by Michelsen44 for land use impacts on biodiversity from forestry operations in Norway. He defined ecosystem scarcity and ecosystem vulnerability factors which “give information on the intrinsic biodiversity value of an area”,44 as well as a criterion for including present conditions for biodiversity in an area. He thus addressed scarcity and vulnerability on an ecosystem level. With VS, we focus on vulnerability and rarity at species level, whereas CpA, being an ecosystem scarcity index, goes in a similar direction as Michelsen’s work.
The combination of TL and EOO into VS considers that impacts in some regions, where rare species reside, may be worse than in regions with more common species, despite the fact that the absolute loss of species in the area might be similar. For example, absolute species loss for waterbirds in “Reserva Natural Lagunas de Campillos” (Spain) and “Luknajno Lake Nature Reserve” (Poland) are similar, but VS and CpA are 1 order of magnitude larger and smaller, respectively (i.e., higher vulnerability and higher habitat loss risk) in the Spanish wetland. Waterbird species richness was 70 in the Spanish wetland and 91 in the Polish wetland, but the EF of the former was 30 times larger.
Both TL and EOO are necessary for complete vulnerability scores. The Red List criteria based on geographic distribution (i.e., EOO or AOO) are important since 21% of the threatened bird species are listed as threatened solely on the basis of geographic distribution and 40% of threatened bird species qualified as threatened under at least one criterion containing geographical distribution.29 Mammals were listed as threatened uniquely due to changes in geographic distribution in 35% of the cases, and qualified as threatened under at least one criterion of geographical distribution in 44% of the cases.29 Yet range size informs only one of the five Red List criteria, and it is not sufficient alone for a species to qualify as threatened (indicators of decline in addition to the small range are indeed necessary for this). The TL highlights the species’ imminent threats, as considered by the Red List, and reflects changes in EOO and population size. EOO itself gives only an indication about geographic distribution, not about threat. However, it provides additional information for nonthreatened, small range species which are more likely to become threatened in future due to fewer possibilities of evasion of impacted regions.
The reference situation commonly used in land use assessment is the potential natural vegetation, even though there are ongoing debates about appropriate reference states.45 Due to the lack of data availability, we used the current situation as reference. One might assume that most wetlands have historically decreased in size, but information about past developments is incomplete. Hence, it is more logical to apply current species richness values instead of historical values. Therefore, we excluded confirmed extinct species in the EF calculation.
Characterization Factors
CFs for waterbirds were largest because they were the most species-rich taxon. Spearman rank order correlations of CFs between different taxa are large for both SW-fed and GW-fed wetlands. This highlights that often several taxa are abundant in one wetland (some, like amphibians, are strongly dependent on water bodies for at least a section of their life cycle). Wetlands and ecosystems in general contain niches for different species that may interact or are dependent on each other. For many waterbirds, amphibians make up a part of their diet,46 and this might explain part of the species richness correlation.
Additionally, the destruction of a wetland has similar consequences on the different taxa because species-area relationship has a common effect, and z-factors are similar for most taxa. The maps of the CFs showed that there are areas with overlapping impacts. Regions with relatively low impacts for individual wetlands may receive large cumulative impacts, as for example, the Danube basin for CFSW with 94 Ramsar wetlands. It is an improvement to not only have CFs on a watershed or country level, but to have them also on individual wetland catchments within the relevant regions since SW consumption upstream of a series of wetlands can damage all of them. Average watershed values cannot take this situation into account and therefore our procedure is a step forward in the spatial refinement of LCIA methodologies (see further explanations in SI S9). The same applies for CFGW where consumption from a certain aquifer can damage multiple wetlands. However, with higher spatial resolution data uncertainty increases and determination of AoR also adds to uncertainty, also when matching inventory to CF.47 Another drawback of the high level of detail is the lack of global coverage (only ∼10% of wetlands considered). Therefore, low CF in some areas might be caused by lack of data and not be due to low species loss risk.
In general, using a combination of multiple taxa is advantageous for identifying priority areas, since the representation of the overall biodiversity is improved.48 In the application example, we considered all species-equivalents to have the same weight. Thus, there is a tendency for species-rich taxa to have higher influences on the impact scores than species-poor groups. It is debatable whether this is justified, since some ecosystem functions are covered by many species, while others are provided only by a few species of a taxon. Consequently, in addition to weighting according to vulnerability within a taxon, a weighting between taxa could be developed in order to adjust impacts from taxa with totally different species numbers. Another approach could be weighting according to trophic level (predators, herbivores, etc.) to reflect the dependence of higher order species on lower level trophic groups. Additionally, this could be reversed to reflect the disproportionate importance of higher trophic groups on ecosystem processes, especially apex predators.49 Keystone species, are another example of species with a disproportionately high influence on the ecosystem of which they are part.50 Their identification is difficult, partly because of differences in definitions and data availability.51 Finally, functional group weighting also represents a possibility to reflect species value in terms of ecosystem function regulation. We recommend a deeper, more extensive discussion on this issue within LCA drawing on available knowledge from the ecological literature. Such a discussion is beyond the scope of the present paper, and until a suitable weighting scheme is developed we suggest that all species should be regarded as having equal value.
Data Quality and Sensitivity
Taking the waterbody area instead of the Ramsar area had the most substantial influence on the EF. Most often, waterbody areas were smaller than Ramsar site areas, and the EF calculated with the smaller area was larger, showing the susceptibility of smaller areas to change. CFs reacted sensitively to changes in underlying area, which is mostly due to the sensitivity of the FF (discussed in ref (17)) and the nonlinear species-area relationship (see SI, S11). As discussed in ref (17), FFs depend strongly on the water consumption (especially for GW-fed wetlands) and this is propagated also to the CF (SI, S11). Hydraulic conductivity and surface water flows also contribute to the sensitivity of the CF.
Global data coverage was available for all taxa, but data quality (i.e., the accuracy and completeness of the range maps)52 differed within and between taxonomic groups. For example, there are 6285 amphibians species listed in the IUCN red list.20 Map and threat level data (both are needed) are available for 6021 species,20,22 that is, for 264 amphibian species no data was available. Reptile map-data is available for 3087 species.21 Only 348 reptile species live in wetlands,20 and of those 80 do not have map data, meaning we could consider 268 species. Many reptile species are also still unknown, in contrast to birds and mammals which have more complete records.53 There is presently no uncertainty information associated to the primary species richness data, impeding a quantification of the associated uncertainty of EF. However, we are confident that the error is small especially for birds and mammals, since most of their species are well-known. Differences between species richness maps with different presence categories were low, indicating basic robustness to data completeness and that the general patterns likely reflect reality. Taking EOO data for most species groups and assuming no seasonal influence on presence/absence (i.e., migration), especially for birds, is likely to have overestimated richness data. Therefore, it is more useful to think of S as the maximum potential species richness of recorded species. For water-dependent mammals there was little difference in species richness when the habitat suitability map (AOO) or the outermost geographical limits of the EOO were taken (SI, S6). AOOs for other species are not yet available, and sensitivities to changes in habitat area cannot be assessed. However, as data becomes available on AOO through distribution models,54 expert opinion or related procedures, the situation is likely to improve.52 Although we only considered a subset of all taxa (exclusion of e.g., plants, fish, insects) because of the global data availability the chosen taxa act as surrogates for the species community present. Still, comparisons to other methods that take other taxa into account have to be applied cautiously (e.g., land use).
Practical Implications
Required inventory information includes the amount of surface water and groundwater consumed at a certain location. For agriculture, for example, water requirement ratios (SI, S13) for determining consumptive water use are available, and the values of Döll et al.55 can be applied for estimating the share of groundwater and surface water consumption. For the background system, global databases can be used (e.g., available for most crops from ref (56)), although the spatial information given is currently rather coarse. If the region of water consumption is unknown, global average CFs and related variability are available (Table 1), but they should be used with caution and include uncertainty estimates. Matching spatially resolved LCI and LCIA data is still a challenge and not yet included in any standard software, but its feasibility has already been demonstrated in research software and publications.47 Therefore, we expect that it is only a matter of time until such analyses become standard practice.
Our approach is well-suited as a screening methodology on a global scale, but it will never replace modeling on local levels, where more details are included and analyzed. We suggest a tiered procedure, using the method of this paper for identifying potential hotspots in the life-cycle of products and processes which should be further investigated with local analysis.
Comparability with other currently existing LCA methodologies is difficult because of the different units used (PDF, species-eq). As mentioned, two wetlands with the same impact in PDF can have quite different species richness. Consequently, the absolute species loss is different and the overall CFs not the same (SI, S12). Research is ongoing to also derive compatible CFs for other ecosystem impact categories, such as the impacts of land use and eutrophication.
Another reason for difficulties in comparisons with existing methodologies is that these do not take species’ vulnerabilities into account. The introduction of TL give threatened species a weight that is up to 5 times larger than for a species of least concern. In ecology, it is common to use linear scales, such as the one proposed here.57 In the ReCiPe project, value choices are made to include either threatened species only (egalitarian perspective) or all species with equal importance (individualist and hierarchist perspectives).58 In our approach, threat status’ of species are taken into account with VS, incorporating the current threat level and the geographical distribution. In addition, species that are not yet threatened but have a small distribution area and are therefore potentially more vulnerable if their habitat is damaged are included, while they are currently not considered if only threatened species are considered. ReCiPe13 takes into account 1.5 million terrestrial species and 100 000 freshwater species to calculate global species densities of 1.38 × 10–8 L/m2 and 7.89 × 10–10 L/m3. We take 10 110 species into account that specifically occur in wetlands. The calculated wetland species density varies between 2.4 × 10–9 L/m2 and 4.8 × 10–2 L/m2 with an average of 1.1 × 10–4 L/m2, and is thus on average 4 orders of magnitude larger than the one from ReCiPe. ReCiPe13 converts PDF into the unit species·yr, but this unit is not directly comparable to our species-eq·yr, since we are targeting global extinction and include weighting according to the threat and distribution of species as well as the threat for habitat loss. As a result, straightforward comparisons are impossible at the moment. This shows the importance of taking local species richness (via richness maps) into account, specifically for wetlands, since they are hotspots of biodiversity. Species density is highly variable and cannot be represented by one global value.
Outlook
Impacts calculated are only impacts on Ramsar wetlands, keeping in mind the irregular distribution of these wetlands. Since most countries currently lack wetland inventories,7 taking the Ramsar database59 is ensuring currently the best global coverage of wetlands. We cover ∼10% of all inland wetlands globally (134 216 253 ha in August 2012),59 among them some of the most important ones. Improved coverage can be facilitated in the future by making use of national wetland inventories and applying the presented approach by using the maps of global species richness calculated here. This allows for deriving generic CFs for wetlands for a specific country or region. In order to more adequately represent biodiversity damages in the future, further research is planned for enhancing the comparability between our approach and other LCIA methodologies for the assessment of ecosystem damages and for a weighting scheme between taxa.
Acknowledgments
We thank BirdLife and IUCN for providing raw species richness data, C.Mutel and M.Curran for helpful comments and C. Raptis and J. Boucher for English proofreading. The research was funded by ETH Research Grant CH1-0208-3 and the European Commission under the 7th framework program on environment (ENV.2009.3.3.2.1: LC-IMPACT- Improved Life Cycle Impact Assessment methods (LCIA) for better sustainability assessment of technologies).
Supporting Information Available
Details and further results are available. This material is available free of charge via the Internet at http://pubs.acs.org.
The authors declare no competing financial interest.
Supplementary Material
References
- Millennium ecosystem assessment. In Ecosystems and Human Well-Being: Wetlands and Water. Synthesis; World Resources Institute: Washington, DC, 2005. [Google Scholar]
- Wetlands, Biodiversity and the Ramsar Convention: The Role of the Convention on Wetlands in the Conservatin and Wise Use of Biodiversity; Halls A. J., Ed.; Ramsar Convention Bureau: Gland, Switzerland, 1997. [Google Scholar]
- Flinn K. M.; Lechowicz M. J.; Waterway M. J. Plant species diversity and composition of wetlands within an upland forest. Am. J. Bot. 2008, 95101216–1224. [DOI] [PubMed] [Google Scholar]
- Lambert A.Economic Valuation of Wetlands: An Important Component of Wetland Management Strategies at the River Basin Scale. http://www.conservationfinance.org/guide/guide/images/18_lambe.pdf (accessed 24 May 2012).
- WWF. Living Planet Report 2012; WWF International: Gland, Switzerland, 2012. [Google Scholar]
- WWF. Threats to wetlands. http://wwf.panda.org/about_our_earth/about_freshwater/intro/threats/ (accessed 24 May 2012).
- Zedler J. B.; Kercher S. Wetland resources: Status, trends, ecosystem services, and restorability. Ann. Rev. Environ. Resour. 2005, 30, 39–74. [Google Scholar]
- ISO. Environmental Management - Life Cycle Assessment - Principles and Framework, International Standard ISO 14040; International Organisation for Standardisation: Geneva, Switzerland, 2006. [Google Scholar]
- Milà i Canals L.; Chenoweth J.; Chapagain A.; Orr S.; Antón A.; Clift R. Assessing freshwater use impacts in LCA: Part I—Inventory modelling and characterisation factors for the main impact pathways. int. J. Life Cycle Assess. 2009, 14128–42. [Google Scholar]
- Pfister S.; Koehler A.; Hellweg S. Assessing the environmental impacts of freshwater consumption in LCA. Environ. Sci. Technol. 2009, 43114098–4104. [DOI] [PubMed] [Google Scholar]
- Hanafiah M. M.; Xenopoulos M. A.; Pfister S.; Leuven R. S.; Huijbregts M. A. J. Characterization factors for water consumption and greenhouse gas emissions based on freshwater fish species extinction. Environ. Sci. Technol. 2011, 45125572–5278. [DOI] [PubMed] [Google Scholar]
- van Zelm R.; Schipper A. M.; Rombouts M.; Snepvangers J.; Huijbregts M. A. J. Implementing groundwater extraction in life cycle impact assessment: Characterization factors based on plant species richness for the Netherlands. Environ. Sci. Technol. 2011, 452629–635. [DOI] [PubMed] [Google Scholar]
- Goedkoop M.;Heijungs R.;Huijbregts M. A. J.;De Schryver A.;Struijs J.;van Zelm R.. ReCiPe 2008: A Life Cycle Impact Assessment Method Which Comprises Harmonised Category Indicators at the Midpoint and Endpoint Levels. First Edition. Report I: Characterization. Ruimte en Milieu, Ministerie van Volkshuisvesting, Ruimtelijke Ordening en Milieubeheer. 2009.
- Goedkoop M.;Spriensma R.. The Eco-Indicator 99. A damage oriented method for life cycle impact assessment. Methodology report and Annex. Pré Consultants, Amersfoort, The Netherlands. 1999. http://www.pre.nl/eco-indicator99/.
- Verones F.; Bartl K.; Pfister S.; Jiménez Vílchez R.; Hellweg S. Modeling the local biodiversity impacts of agricultural water use: Case study of a wetland in the coastal arid area of Peru. Environ. Sci. Technol. 2012, 4694966–4974. [DOI] [PubMed] [Google Scholar]
- Curran M.; De Baan L.; De Schryver A.; Van Zelm R.; Hellweg S.; Koellner T.; Sonnemann G.; Huijbregts M. A. J. Toward meaningful end points of biodiversity in life cycle assessment. Environ. Sci. Technol. 2011, 45170–79. [DOI] [PubMed] [Google Scholar]
- Verones F.; Pfister S.; Hellweg S. Quantifying area changes of internationally important wetlands due to water consumption in LCA. Environ. Sci. Technol. 2013, 10.1021/es400266v. [DOI] [PubMed] [Google Scholar]
- Ramsar Convention. Convention on Wetlands of International Importance especially as Waterfowl Habitat, The Convention on Wetlands text, as amended in 1982 and 1987. Director, Office of International Standards and Legal Affairs; United Nations Educational, Scientific and Cultural Organization (UNESCO): Paris, 1994. [Google Scholar]
- BirdLife International;Nature Serve. Bird Species Distribution Maps of the World; BirdLife International: Cambridge, UK: and NatureServe: Arlington, USA, 2011. [Google Scholar]
- IUCN, (International Union for Conservation of Nature and Natural Resources). IUCN Red List of Threatened Species. Version 2012.1. http://www.iucnredlist.org (accessed 26 November 2012).
- IUCN, (International Union for Conservation of Nature and Natural Resources). Spatial data for reptiles. http://www.iucnredlist.org/technical-documents/spatial-data#reptiles (accessed 26 November 2012).
- IUCN, (International Union for Conservation of Nature and Natural Resources). Spatial data for amphibians. http://www.iucnredlist.org/technical-documents/spatial-data#amphibians (accessed 26 November 2012).
- BirdLife International. BirdLife Data Zone - Species Search. http://www.birdlife.org/datazone/species/search (accessed 30 April 2012).
- MathWorks. Matlab Version 7.12.0.635 (R2011a), 2011.
- Rondinini C.; Di Marco M.; Chiozza F.; Santulli G.; Baisero D.; Visconti P.; Hoffmann M.; Schipper J.; Stuart S. N.; Tognelli M. F.; Amori G.; Falcuscci A.; Maiorano L.; Boitani L. Global habitat suitability models of terrestrial mammals. Phil. Trans. R. Soc. B 2011, 366, 2633–2641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koellner T.; Scholz W. R. Assessment of land use impacts on the natural environment. Part 2: Generic characterization factors for local species diversity in central europe. int. J. Life Cycle Assess. 2008, 13, 32–48. [Google Scholar]
- Schmidt J. Development of LCIA characterisation factors for land use impacts on biodiversity. J. Clean. Prod. 2008, 16, 1929–1942. [Google Scholar]
- Drakare S.; Lennon J.; Hillebrand H. The imprint of the geographical, evolutionary and ecological context on species-area relationships. Ecol. Lett. 2006, 9, 215–227. [DOI] [PubMed] [Google Scholar]
- Gaston K. J.; Fuller R. A. The size of species’ geographic range. J. Appl. Ecol. 2009, 46, 1–9. [Google Scholar]
- IUCN, (International Union for Conservation of Nature and Natural Resources). 2001 IUCN Red List Categories and Criteria version 3.1. http://www.iucnredlist.org/technical-documents/categories-and-criteria/2001-categories-criteria (accessed 9 October 2012).
- IUCN, (International Union for Conservation of Nature and Natural Resources). The IUCN Red List of Threatened Species. Version 2012.2. www.iucnredlist.org (accessed 25 October 2012).
- Lehner B.; Verdin K.; Jarvis A. New global hydrography derived from spaceborne elevation data. Eos, Transactions, AGU. 2008, 891093–94. [Google Scholar]
- Lehner B.; Döll P. Development and validation of a global database of lakes, reservoirs and wetlands. J. Hydrol. 2004, 2961–41–22. [Google Scholar]
- New M.; Lister D.; Hulme M.; Makin I. A high-resolution data set of surface climate over global land areas. Clim. Res. 2002, 2111–25. [Google Scholar]
- Trabucco A.;Zomer R. Global Potential Evapo-Transpiration (Global-PET) and Global Aridity Index (Global-Aridity) Geo-Database., CGIAR Consortium for Spatial Information, Editor 2009, Available online from the CGIAR-CSI GeoPortal. http://www.csi.cgiar.org.
- USGS, (US Geological Survey). GTOPO30. http://eros.usgs.gov/#/Find_Data/Products_and_Data_Available/gtopo30_info (accessed 21 May 2012).
- Fensham R. J.; Fairfax R. J. Spring wetlands of the Great Artesian Basin, Queensland, Australia. Wetlands Ecol. Manage. 2003, 115343–362. [Google Scholar]
- Gaston K. J. How large is a species’ geographic range?. Oikos. 1991, 61, 434–438. [Google Scholar]
- Kenya Flower Council. The flower industry in Kenya. http://www.kenyaflowercouncil.org/floricultureinkenya.php (accessed 19 October 2012).
- Mekonnen M. M.;Hoekstra A. Y.. Mitigating the Water Footprint of Export Cut Flowers from the Lake Naivasha Basin, Kenya. Values of Water Research Report Series No.45; UNESCO-IHE: Delft, the Netherlands, 2010. [Google Scholar]
- Torrellas M.; Antón A.; Ruijs M.; García Victoria N.; Stanghellini C.; Montero J. I. Environmental and economic assessment of protected crops in four European scenarios. J. Clean. Prod. 2012, 28, 45–55. [Google Scholar]
- Antón A.Personal communication 31 January 2013. 2013
- ESRI. ESRI Data&Maps. http://www.arcgis.com/home/group.html?owner=esri&title=ESRI%20Data%20%26%20Maps&content=all&focus=maps (accessed 14 December 2009).
- Michelsen O. Assessment of Land Use Impact on Biodiversity. Proposal of a new methodology exemplified with forestry operations in Norway. Int. J. Life Cycle Assess. 2008, 13122–31. [Google Scholar]
- Koellner T.;de Baan L.;Beck T.;Brandão M.;Civit B.;Margni M.;Milà i Canals L.;Saad R.;Maia de Souza D.;Müller-Wenk R.. UNEP-SETAC guideline on global land use impact assessment on biodiversity and ecosystem services in LCA. Int. J. Life Cycle Assess. in press, [Google Scholar]
- Horváth Z.; Ferenczi M.; Móra A.; Vad C. F.; Ambrus A.; Forró L.; Szövényi G.; Andrikovics S. Invertebrate food sources for waterbirds provided by the recostructed wetland of Nyirkai-Hany, nothwestern Hungary. Hydrobiologia. 2012, 697159–72. [Google Scholar]
- Mutel C. L.; Pfister S.; Hellweg S. GIS-based regionalized life cycle assessment: How big is small enough? Methodology and case study of electricity generation. Environ. Sci. Technol. 2012, 4621096–1103. [DOI] [PubMed] [Google Scholar]
- Larsen F. W.; Bladt J.; Balmford A.; Rahbek C. Birds as biodiversity surrogates: Will supplementing birds with other taxa improve effectiveness?. J. Appl. Ecol. 2012, 49, 349–356. [Google Scholar]
- Estes J. A.; Terborgh J.; Brashares J. S.; Power M. E.; Berger J.; Bond W. J.; Carpenter S. R. Trophic Downgrading of Planet Earth. Science. 2011, 3336040301–306. [DOI] [PubMed] [Google Scholar]
- Leader-Williams N.;Dublin H. T.. Charismatic megafauna as ‘flagship species’. In Priorities for the Conservation of Mammalian Diversity. Has the Panda Has Its Day?; Entwsistle A., Dunstone N., Eds.; Cambridge University Press: Cambridge, UK. 2000; p 53–81. [Google Scholar]
- Cottee-Jones H. E. W.; Whittaker R. J. The keystone species concept: A critical appraisal. Front. Biogeogr. 2012, 43117–127. [Google Scholar]
- Jetz W.; McPherson J. M.; Guralnick R. P. Integrating biodiversity distribution knowledge: Toward a global map of life. Trends Ecol. Evol. 2011, 273151–159. [DOI] [PubMed] [Google Scholar]
- Groombridge B.;Jenkins M. D.. World Atlas of Biodiversity, Prepared by the UNEP World Conservation Monitoring Centre; University of California Press: Berkeley, CA, 2002. [Google Scholar]
- Elith J.; Graham C. H.; Anderson R. P.; Dudik M.; Ferrier S.; Guisan A.; Hijmans R. J. Novel methods improve prediction of species’ distributions from occurrence data. Ecography. 2006, 292129–151. [Google Scholar]
- Döll P.; Hoffmann-Dobrev H.; Portmann F.; Siebert S.; Eicker A.; Rodell M.; Strassberg G.; Scanlon B. Impact of water withdrawals from groundwater and surface water on continental water storage variations. J. Geodyn. 2011, 59-60, 143–156. [Google Scholar]
- Pfister S.; Bayer P.; Koehler A.; Hellweg S. Environmental impacts of water use in global crop production: Hotspots and trade-offs with land use. Environ. Sci. Technol. 2011, 45135761–5768. [DOI] [PubMed] [Google Scholar]
- Purvis A.;Cardillo M.;Grenyer R.;Collen B.. Correlates of extinction risk: Phylogeny, biology, threat and scale. In Phylogeny and Conservation. Conservation Biology 8; Purvis A., Gittleman J. L., Brooks T., Eds.; Cambridge University Press: Cambridge, UK, 2005; p 448. [Google Scholar]
- De Schryver A. M.; Brakkee K. W.; Goedkoop M. J.; Huijbregts M. A. J. Characterization factors for global warming in life cycle assessment based on damages to humans and ecosystems. Environ. Sci. Technol. 2008, 4361689–1695. [DOI] [PubMed] [Google Scholar]
- Ramsar Sites Information Service. http://ramsar.wetlands.org/Database/AbouttheRamsarSitesDatabase/tabid/812/Default.aspx (accessed 16 May 2012).
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.