

Abstract
It has been shown that dysregulation of IGF-1 signaling is associated with tumor incidence and progression, whereas blockade of the signaling can effectively inhibit carcinogenesis. Although several mechanisms of anticancer activity of quercetin were proposed, molecular targets of quercetin have not been identified yet. Hence, we assessed the effect of quercetin on IGF-1 signaling inhibition in BK5.IGF-1 transgenic (Tg) mice, which over-expresses IGF-1 in the skin epidermis. A quercetin diet (0.02% wt/wt) for 20 weeks remarkably delayed the incidence of skin tumor by 2 weeks and reduced tumor multiplicity by 35% in a 7,12-dimethylbenz(a)anthracene (DMBA)-tetradecanoyl phorbol-13-acetate (TPA) two stage mouse skin carcinogenesis protocol. Moreover, skin hyperplasia in Tg mice was significantly inhibited by a quercetin supplementation. Further analysis of the MT1/2 skin papilloma cell line showed that a quercetin treatment dose dependently suppressed IGF-1 induced phosphorylation of the IGF-1 receptor (IGF-1R), insulin receptor substrate (IRS)-1, Akt and S6K; however, had no effect on the phosphorylation of PTEN. Additionally, the quercetin treatment inhibited IGF-1 stimulated cell proliferation in a dose dependent manner. Taken together, these data suggest that quercetin has a potent anticancer activity through the inhibition of IGF-1 signaling.
Keywords: Quercetin, IGF-1, carcinogenesis, mouse skin cancer
Introduction
Insulin-like growth factor (IGF)-1 plays an important role in the regulation of cell proliferation, survival, and growth. Its primary action is mediated by binding to the insulin-like growth factor-1 receptor (IGF-1R). Activated IGF-1R via IGF-1 phosphorylates and activates the insulin receptor substrate-1 (IRS-1) and leads to the phosphorylation of Akt [1]. Akt subsequently activates the mammalian target of rapamycin (mTOR) and S6 kinase (S6K), which is involved in protein synthesis and cell growth. Autophosphorylated IGF-1R also activates Ras, Raf, MEK and ERK signaling pathways. The elevated level of plasma IGF-1 is associated with the risk of several cancers in multiple organs, such as the colon, prostate, and breast [2-4]. Moreover, the elevated tissue levels of IGF-1 enhance susceptibility to chemically induced skin tumorigenesis, and inhibition of IGF-1 signaling decreases tumor formation [5].
Many epidemiological and experimental studies have reported that natural dietary compounds found in fruits and vegetables reduce cancer incidences, as well as tumor multiplicity [6-8]. Quercetin is one of the most common flavonoids, and has potent anti-carcinogenic effects derived from its anti-inflammatory and anti-oxidative activities [6]. The amount of quercetin intake was negatively correlated with the incidence of colorectal [7] and lung cancers in clinical studies [8]. It has also been reported that quercetin inhibits hydrogen peroxide-induced DNA damage and enhances DNA repair in the colon cancer cell line [9]. Quercetin also induces apoptosis by activating the pro-apoptotic proteins, such as caspase 3 and caspase 9, and inducing cell cycle arrest in several cancer cells [10]. Most studies thus far, however, reported anticancer activities of quercetin in the context of its antioxidative properties, and only a limited number of studies have focused on its modulation of signaling pathways, such as NFκB, MAPK, p53, androgen receptor and estrogen receptor in tumor growth [11-14]. In this study, quercetin effectively inhibited the development of skin tumor in BK5. IGF-1 transgenic (Tg) mice and decreased the phosphorylation of IGF-1R, IRS-1, Akt and S6K in skin cancer cells, which indicates that quercetin has potent anticarcinogenic activity through the inhibition of IGF-1 signaling pathway. To the best of our knowledge, this is the first study to demonstrate that quercetin specifically blocks IGF-1R activation in vivo.
Materials and Methods
Materials
7,12-Dimethylbenz(a)anthracene (DMBA) and 5-bromo-2'-deoxyuridine (BrdU) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Tetradecanoyl phorbol-13-acetate (TPA) was purchased from Alexis Biochemicals (San Diego, CA, USA). All other chemicals were obtained from Sigma-Aldrich, unless otherwise indicated.
Animals and diet
BK5.IGF-1 Tg mouse were imported from Dr. John DiGiovanni's Lab (University of Texas M.D. Anderson Cancer Center, TX, USA) and bred in Catholic University of Daegu. All animals were kept in controlled conditions of humidity (50 ± 10%), light (12-hour light/dark cycle), and temperature (23 ± 2℃). Animal protocols utilized in this study were approved by the Animal Care and Use Committee at Catholic University of Daegu (IACUC-2010-8). The composition of AIN-93G diet (Control) or quercetin diet (AIN-93G diet containing 0.02% quercetin) was indicated in Table 1. Dose of quercetin was chosen based on a previous animal study, which has shown a decreased intestinal polyp multiplicity with 0.02% quercetin diet [15]. Food intakes were recorded daily and body weights were recorded once a week.
Table 1.
Composition of experimental diet
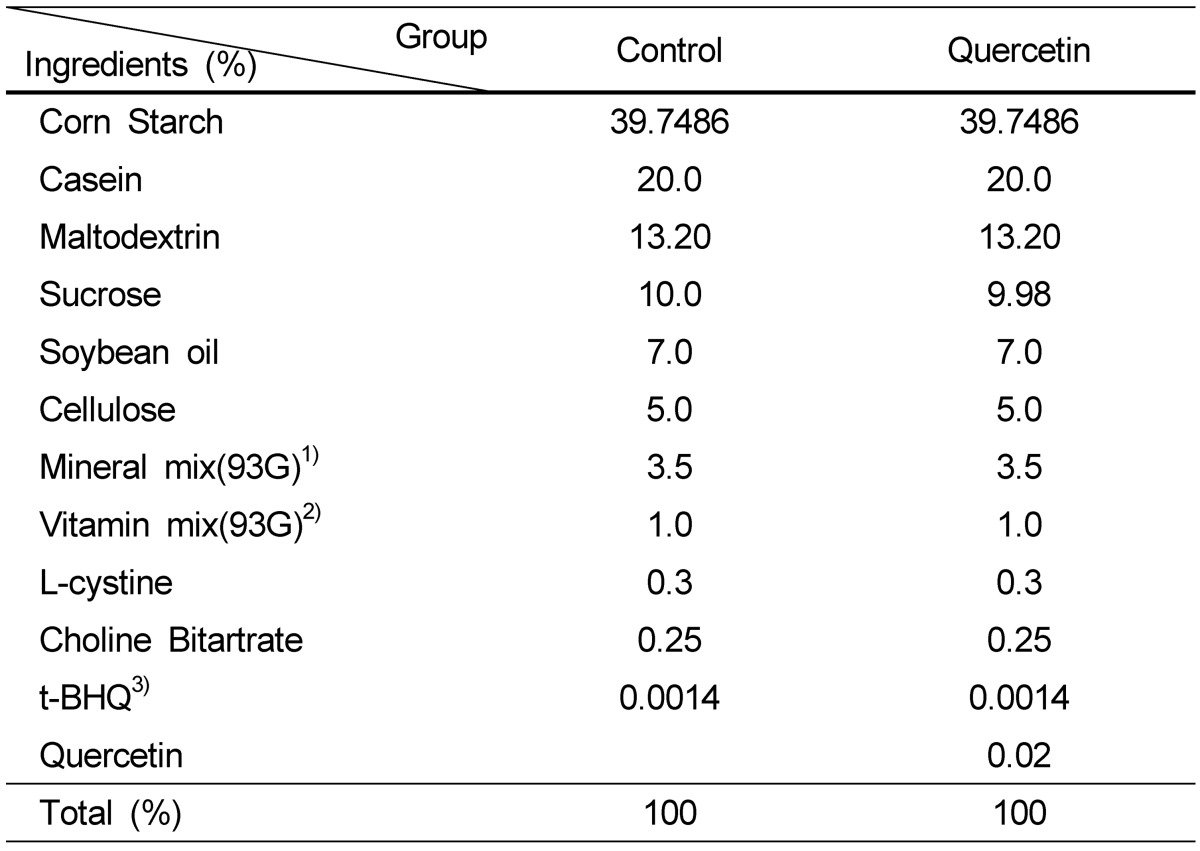
1)Composition of AIN-93G mineral Mix (g/kg) : Calcium carbonate, anhydrous 357.00; Potassium phosphate, monobasic 196.00; Potassium citrate, tri-potassium, monohydrate 70.78; Sodium chloride 74.00; Potassium sulfate 46.60; Magnesium oxide 24.00; Ferric citrate 6.06; Zinc carbonate 1.65; Manganous carbonate 0.63; Cupric carbonate 0.30; Potassium iodate 0.01; Sodium selenate, anhydrous 0.01025; Ammonium paramolybdate 4 hydrate 0.00795; Sodium meta-silicate, 9 hydrate 1.45; Chromium potassium sulfate, 12 hydrate 0.275; Lithium chloride 0.0174; Boric acid 0.0815; Sodium fluoride 0.0635; Nickel carbonate 0.0318; Ammonium vanadate 0.0066; Powdered sucrose 221.026
2)Composition of AIN-93G vitamin Mix (g/kg) : Nicotinic acid 3.000; Ca pantothenate 1.600; Pyridoxine-HCl 0.700; Thiamin-HCl 0.600; Riboflavin 0.600; Folic acid 0.200; D-Biotin 0.020; Vitamin B-12(cyanocobalamin) 2.500; Vitamin E (all-rac-α-tocopheryl acetate, 500 IU/g) 15.00; Vitamin A (all-trans-retinyl palmitate, 500.000 IU/g) 0.800; Vitamin D3 (cholecalciferol, 400.000IU/g) 0.250;Vitamin K (phylloquinone) 0.075; Powdered sucrose 974.655
3)t-BHQ : tert-Butylhydroquinone
Animal Experiment 1 (Two-stage mouse skin carcinogenesis)
The first animal study was performed to investigate the consequences of quercetin diet on skin carcinogenesis. Seventeen male BK5.IGF-1 Tg mice were randomly divided into two groups; control (n = 8) and quercetin (n = 9) group. At 8 weeks of age, dorsal skins of the mice were shaved two days before tumor initiation. Initiation was accomplished by a single topical application with 200 nmol (50 µg) DMBA under a subdued light. At 2 weeks after initiation, the mice were treated with 6.5 nmol (4 µg) of TPA in 200 µl of acetone twice a week for 20 weeks. Tg mice were fed AIN-93G diet (control) or quercetin diet (AIN-93G diet containing 0.02% quercetin) at 10 weeks of age with TPA promotion until the end of the experiment. Tumor incidence (number of mice bearing tumors/total number of mice) and tumor multiplicity (number of papillomas/mouse) were manually counted and recorded weekly. Tumor size was measured in diameters and length of each tumor by a caliper once a week.
Animal Experiment 2 (Histological analysis of Epidermal cell proliferation)
Eight-week old twelve male BK5.IGF-1 Tg mice were divided into the control (n = 6) and quercetin (n = 6) groups to investigate the effects of quercetin diet on epidermal cell proliferation of the Tg mice. Animals were fed ad libitum with either a control or quercetin diet for 4 weeks. In each diet group, dorsal skins of the mice were shaved and treated with either acetone (n = 3) or TPA (6.5 nmol (4 µg)/200 µl acetone/mouse, n = 3) twice a week for 2 weeks before sacrificing the animals. For measuring the rate of epidermal cell proliferation, 100 mg/kg body weight of BrdU was injected i.p. into mice 1 hr before sacrifice. Dorsal skin was isolated from the mice, fixed in formalin, and processed for paraffin embedding. Tissue sections were stained with hematoxylin and eosin (H&E) solution and immunostained with antibodies against BrdU (BD Biosciences, San Jose, CA, USA). The number of BrdU-positive cells in the interfollicular epidermis was counted in five random tissue sections in each mouse. The labeling index was calculated as the percentage of BrdU-positive basal cells to the total number of basal cells in the interfollicular epidermis.
Cell Culture
The murine papilloma cell line, the MT1/2 cells [16], were kindly provided by Dr. Susan M. Fischer's Lab (University of Texas M.D. Anderson Cancer Center, TX, USA), and were maintained in Minimum essential medium (GIBCO, Carlsbad, CA, USA) containing 10% FBS and 1% penicillin-streptomycin. All cell lines used in this study were maintained in a 37℃ humidified incubator with 5% CO2.
MTT Assay
To assess cell viability and cell proliferation, the MT1/2 cells (1.25 × 104 cells/well) were plated in 96-well plates. The cells were treated with various concentrations of quercetin (10, 20, 30, 40 and 50 µg/ml) or IGF-1 (20 ng/ml) for an indicated time. Then, the medium was aspirated from the wells, and 100 µl fresh serum-free medium containing MTT reagent (1 mg/ml) was added to each well. The cells were incubated for 4hr at 37℃ and lysed by addition of 100 µl DMSO, and the optical density (OD) at 540 nm was then measured.
Western Blotting Analysis
Cells were harvested in a mild-lysis buffer (10 mM Tris-HCl (pH7.5), 100 mM NaCl, 1% NP-40, 50 mM NaF, 2 mM EDTA (pH 8.0), 1 mM DTT, 1 mM PMSF, 10 µg/ml leupeptin and 10 µg/ml aprotonin). Cell lysates were subjected to 10% SDS-polyacrylamide gel electrophoresis, and subsequently, transferred onto the polyvinylidene difluoride membrane. After transfer, the membrane was blocked with a 5% non-fat dried milk in 0.1% Tween-20-Tris-buffered saline (TTBS) and probed with antibody against phospho Akt (S473), Akt, phospho S6K (T389), S6K, phospho Erk1/2 (Thr202/Tyr204), Erk, pIGF-1R (T1131), IGF-1R, pIRS-1 (S307), IRS-1, pPTEN (S380/T382/383), and PTEN (Cell Signaling Technology, Danvers, CO, USA) at 1:1000 dilution for O/N at 4℃. After three times of washing with a TTBS buffer, the membrane was incubated with a horse radish peroxidase-linked secondary antibody (1:1000) and visualized by ECL kit (Bio Science Technology, Pohang, Korea) according to the recommended procedure.
Statistical Analysis
All statistical analyses were performed by the SPSS program (Ver. 17), and the data were analyzed by a t-test and ANOVA analysis. The differences between the experimental groups were evaluated at the P < 0.05.
Results
Dietary quercetin supplementation delayed the time of tumor incidence and suppressed tumor multiplicity in BK5.IGF-1 Tg mice
The effect of quercetin on skin carcinogenesis was investigated in vivo using BK5.IGF-1 Tg mice in the first animal experiment (experiment 1). The mice were grouped into the control and quercetin diet groups, and fed with each diet for 20 weeks with TPA promotion. During the experimental period, food intakes of the control and quercetin groups were 7.3 and 7.4 g/day, respectively, and there was no significant difference between the two groups (Table 2). The body weight was gradually elevated without significant differences between the control and quercetin groups (Fig. 1). In DMBA-TPA two stage skin carcinogenesis protocol, animals in the control group developed skin papillomas at 2 weeks and reached up to 100% tumor incidence at 4 weeks of TPA promotion (Fig. 2A). In the quercetin group, skin papillomas were developed at 4 weeks and 100% of tumor incidence was recorded at 6 weeks of TPA promotion. This data indicates that the incidence of tumors was delayed by 2 weeks by quercetin supplementation. Quercetin also decreased mouse skin tumor multiplicity by 35% compared to that of the control group (Fig. 2B). It has been reported that BK5.IGF-1 Tg mice spontaneously developed skin papillomas due to highly activated IGF-1 signaling pathways [5]. We observed that spontaneous tumor development in the DMBA-TPA untreated region of the skin was also reduced by quercetin supplementation (Fig. 2C, 2D).
Table 2.
Daily diet intakes in BK5.IGF-1 transgenic mice

In experiment 1, BK5.IGF-1 transgenic mice in each group were fed either control diet (AIN-93G) or quercetin (AIN-93G + 0.02% quercetin) diet for 20 weeks. Food intake was recorded daily. Values are the mean ± SE.
Fig. 1.

Body weight change. In experiment 1, BK5.IGF-1 Tg mice in the control (n = 8) and quercetin (n = 9) diet groups were initiated with 200 nmol (50 µg) of DMBA and promoted with 6.5 nmol (4 µg) of TPA in 200 µl of acetone twice a week for 20 weeks. The body weight of BK5.IGF-1 Tg mice was measured once a week during a 20-week experimental period. Values are the mean ± SE.
Fig. 2.

Inhibition of skin tumor multiplicity by quercetin supplementation. A) Tumor incidence and B) tumor multiplicity of the control (n = 8) and quercetin (n = 9) diet groups were recorded weekly in experiment 1. C) Spontaneous tumor development was recorded. Three mice of the control group and one mouse in the quercetin group developed a total of 9 and 2 spontaneous tumors, respectively. D) Photographs of spontaneous tumor development in the non-TPA treated area in BK5.IGF-1 Tg mice. Values are the mean ± SE. *P < 0.05, **P < 0.01 and ***P < 0.001 when compared to the control group.
Quercetin decreased total number and size of tumors
In animal experiment 1, we further investigated how quercetin changed the number and size of the tumors. Quercetin supplementation showed a significant decrease in the tumor diameter (length and width) compared to that of the control group. At the end of the experimental period, the average number of tumors < 0.5 cm was 97 and 67.2, 0.5-1 cm were 32.7 and 18, and > 1 cm were 2.8 and 0.8, in the control and quercetin groups, respectively (Table 3). Quercetin supplementation also decreased the total number of tumors in the quertcetin group compared to that of the control group (Table 3).
Table 3.
Effect of quercetin supplementation on tumor size in BK5. IGF-1 transgenic mice

In experiment 1, the tumor size (length and width) were measured every week as described in the Materials and Methods section. Data shown is the record of tumor numbers according to the size of tumors at the end of chemical carcinogenesis protocol. Those numeric symbols < 0.5, 0.5-1, > 1 indicate the size of tumors which are less than 0.5 cm, or between 0.5-1 cm or bigger than 1 cm, respectively. Values are the mean ± SE. *P < 0.05, **P < 0.01 and ***P < 0.001 when compared to control group.
Quercetin reduced TPA-induced epidermal cell proliferation in BK5.IGF-1 Tg mice
To see the effect of quercetin on epidermal cell proliferation, we conducted a BrdU incorporation experiment (Experiment 2). TPA treatment induced skin hyperplasia in the control group, but quercetin supplementation remarkably reduced TPA-induced skin hyperplasia (Fig. 3A). BrdU labeling index was also reduced by quercetin supplementation, both in acetone and TPA treated animals (Fig. 3B).
Fig. 3.

Effect of quercetin on skin epidermal hyperplasia and basal cell proliferation. A) Histological analysis of acetone and TPA-treated BK5.IGF-1 Tg mouse skin. At the end of experiment 2, tissue sections were immunostained with an antibody against BrdU and photographed at 200X. The dorsal skins of BK5.IGF-1 Tg mice were fed with a) control diet treated with acetone, b) quercetin diet treated with acetone, c) control diet treated with TPA, and d) quercetin diet treated with TPA. B) Labeling index of acetone or TPA-treated mouse skin in each diet group. The index represents the percentage of the BrdU positive cells relative to the total number of the basal cells in the interfollicular epidermis. Each value is the mean ± SE of labeling indices from 3 mice/group. *P < 0.05
Quercetin suppressed IGF-1 signaling by inhibition of IGF-1R phosphorylation in skin cancer cells
To investigate a mechanism on the inhibition of skin tumor development in BK5.IGF-1 Tg mice by quercetin, we analyzed the phosphorylation of protein components in the IGF-1 signaling pathway by a quercetin treatment in a murine papilloma cell line, the MT1/2 cells. As shown in Fig. 4A, IGF-1 stimulation significantly increased the phosphorylation of IGF-1R, IRS-1, Akt, and S6K, and 1 hr treatment of quercetin effectively suppressed the phosphorylation of these proteins in a dose dependent manner without affecting the cell viability (Fig. 4B). Phosphorylation of PTEN, a negative regulator of IGF-1 signaling [19], however, was not changed by the quercetin treatment. We further investigated whether quercetin could abrogate the IGF-1 induced cell proliferation. Simultaneous incubation of quercetin (10-50 µM) and IGF-1 for 48 hrs significantly inhibited the IGF-1 stimulated cell proliferation, and especially > 30 µM of the quercetin inhibited cell proliferation as low as the cells in a serum free media (Fig. 4C).
Fig. 4.

Inhibition of IGF-1R activation and downstream effectors by quercetin in MT1/2 cells. A) Serum starved MT1/2 cells were treated with indicated doses (10 µM - 50 µM) of quercetin for 1 hr and then stimulated with IGF-1 (20 ng/ml) for 30 min. The phosphorylation and protein levels were determined by immunoblotting with the appropriate antibodies, as indicated. B) Cell viability was assessed by MTT assay. C) Cells were grown in serum free media and treated with IGF-1 (20 ng/ml) and indicated doses (10-50 µM) of quercetin for 48 hr. Proliferation of MT1/2 cells were measured by an MTT assay. Values are expressed as the means ± SE. Significant differences (P < 0.05) are indicated by different letters. SFM: serum free media, NS: not significant.
Discussion
Dysregulation of the IGF-1/IGF-1R system has been implicated in the proliferation of numerous cancer cells, including the colon, gastric, esophageal, hepatocytes and pancreatic cells, and a blockade of IGF-1 signaling pathways effectively inhibits tumor progression [2,3,17]. Commercially available compounds, inhibiting the IGF-1 signaling pathway (e.g.,wortmannin, picropodophyllin, NVP-AEW541), have been extensively investigated as a therapeutic option for people with malignant cancer [18,19]; however, it would be beneficial to offer natural therapies that could be easily incorporated into the diet and would act on the same target to reduce the risk of cancer. Particularly, quercetin highly abundant in plant food, such as onion, kale and broccoli [20,21], has been shown to have potent anti-carcinogenic and anti-tumorigenic properties in many organs, and suppress the growth and metastasis of the melanoma cells [12,13,22]. Although many studies reported the effects of quercetin in regulation of key elements in the cellular signal transduction pathways linked to the cell survival and proliferation, its molecular action in direct modulation of the IGF-1 signaling pathway remains to be answered.
The present study investigated a mechanism underlying the anticarcinogenic effect of quercetin on skin cancer in the context of IGF-1 signaling. BK5.IGF-1 Tg mice, which overexpress IGF-1 in the basal cells of skin epidermis, were utilized to confirm the effect of quercetin on the suppression of the IGF-1 signaling pathway. Quercetin supplementation decreased the DMBA-TPA induced tumor formation, as well as tumor size, and decreased TPA induced skin hyperplasia in Tg mice. Similar findings with epigallocatechin gallate (EGCG) have also been reported to decrease TPA induced skin tumor formation [23] and tumor size in lung cancer animal models [24]. BK5.IGF-1 transgenic mice also spontaneously induce tumors without TPA treatment due to spontaneous stimulation of IGF-1 signaling, and quercetin supplementation even suppressed spontaneous tumor formation. Although this study did not show the level of circulating IGF-1 in transgenic mice, IGF-1 has known for activating IGF-1 signaling pathway and inducing cell proliferation by autocrine or in a paracrine manner [25]. These data indicate the direct modulation of the IGF-1 signaling pathway by quercetin.
In addition to using an animal study to confirm the suppression of IGF-1 signaling induced skin tumor by quercetin, we analyzed the components of the IGF-1 signaling pathway in skin cancer cells to identify an actual target responsible for the chemopreventive effects of quercetin and elucidate a molecular mechanism of action. Quercetin treatment dose dependently decreased IGF-1 stimulated IGF-1R activation, as well as phosphorylation of IRS-1, Akt and S6K in the MT1/2 cells in this study. However, protein expression and phosphorylation of PTEN was not affected by a quercetin treatment. IGF-1R is a glycoprotein complex consisting of two extracellular α subunits, which confer binding specificity, and two transmembrane β subunits, which contain tyrosine kinase activity [26] and mediates action of IGF-1 on cellular differentiation and proliferation in cancer cells [27]. A ratio of IGF-1R and IRS-1 is mainly associated with cellular response to IGF-1 due to the inhibitory action of IRS-1 on IGF-1R induced cell differentiation [28]. The tumor suppressor gene, PTEN, is a negative regulator of the PI3K-Akt cell survival pathway, and activation of PI3K-Akt signaling through a loss of PTEN function is common in many other types of tumors [29-31]. However, protein expression and phosphorylation of PTEN was not affected by quercetin treatment, suggesting that suppression of Akt phosphorylation by quercetin is a PI3K cascade response of the suppression of upstream molecule of IGF-1 signaling. Based on the results of this study, IGF-1R appears to be the main target of quercetin in the blocking of IGF-1 signaling. It will be needed to further investigate whether quercetin directly binds to IGF-1R and inhibits its kinase activity.
On the other hand, how much of quercetin is absorbed and utilized at the physiological level is not consistent in the previously reported studies, since the bioavailability and metabolic range of quercetin (3-50 µg/L)[32] are varied by the type of supplements and food sources [33-35]. We used the doses of quercetin in the range of 10-50 µM in cell culture of this study and expect that these amounts can be achieved by eating plant based foods as a concentrated form, such as tea, tablet, and fruit skin extracts.
Signal transduction pathways in the downstream of IGF-1R are complex and have been studied extensively in cultured cells. Moreover, quercetin has been shown to inhibit a variety of enzymes involved in the cell proliferation and signal transduction pathways, including protein kinase C [36], tyrosine kinase [37], PI3K [38] and JNK [39]. Quercetin also has a binding activity to the type II estrogen binding sites [40] or aryl hydrocarbon receptors [41]. Therefore, other possible mechanisms of anticancer activity of quercetin cannot be excluded.
In this study, we identified that quercetin directly inhibited IGF-1R phosphorylation in the MT1/2 cells and suppressed skin cancer development in BK5. IGF-1 Tg mice. These results suggest a possible mechanism on how quercetin prevented cancer progression in the previous clinical and sub-clinical animal studies.
Footnotes
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (NRF-2008-331-C00308).
References
- 1.Kim E, Goraksha-Hicks P, Li L, Neufeld TP, Guan KL. Regulation of TORC1 by Rag GTPases in nutrient response. Nat Cell Biol. 2008;10:935–945. doi: 10.1038/ncb1753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Harrington LS, Findlay GM, Gray A, Tolkacheva T, Wigfield S, Rebholz H, Barnett J, Leslie NR, Cheng S, Shepherd PR, Gout I, Downes CP, Lamb RF. The TSC1-2 tumor suppressor controls insulin-PI3K signaling via regulation of IRS proteins. J Cell Biol. 2004;166:213–223. doi: 10.1083/jcb.200403069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Remacle-Bonnet MM, Garrouste FL, Heller S, André F, Marvaldi JL, Pommier GJ. Insulin-like growth factor-I protects colon cancer cells from death factor-induced apoptosis by potentiating tumor necrosis factor alpha-induced mitogen-activated protein kinase and nuclear factor kappaB signaling pathways. Cancer Res. 2000;60:2007–2017. [PubMed] [Google Scholar]
- 4.Shah OJ, Wang Z, Hunter T. Inappropriate activation of the TSC/Rheb/mTOR/S6K cassette induces IRS1/2 depletion, insulin resistance, and cell survival deficiencies. Curr Biol. 2004;14:1650–1656. doi: 10.1016/j.cub.2004.08.026. [DOI] [PubMed] [Google Scholar]
- 5.DiGiovanni J, Bol DK, Wilker E, Beltrán L, Carbajal S, Moats S, Ramirez A, Jorcano J, Kiguchi K. Constitutive expression of insulin-like growth factor-1 in epidermal basal cells of transgenic mice leads to spontaneous tumor promotion. Cancer Res. 2000;60:1561–1570. [PubMed] [Google Scholar]
- 6.Serafini M, Bellocco R, Wolk A, Ekström AM. Total antioxidant potential of fruit and vegetables and risk of gastric cancer. Gastroenterology. 2002;123:985–991. doi: 10.1053/gast.2002.35957. [DOI] [PubMed] [Google Scholar]
- 7.Kyle JA, Sharp L, Little J, Duthie GG, McNeill G. Dietary flavonoid intake and colorectal cancer: a case-control study. Br J Nutr. 2010;103:429–436. doi: 10.1017/S0007114509991784. [DOI] [PubMed] [Google Scholar]
- 8.Neuhouser ML. Dietary flavonoids and cancer risk: evidence from human population studies. Nutr Cancer. 2004;50:1–7. doi: 10.1207/s15327914nc5001_1. [DOI] [PubMed] [Google Scholar]
- 9.Duthie SJ, Dobson VL. Dietary flavonoids protect human colonocyte DNA from oxidative attack in vitro. Eur J Nutr. 1999;38:28–34. doi: 10.1007/s003940050043. [DOI] [PubMed] [Google Scholar]
- 10.Granado-Serrano AB, Martín MA, Bravo L, Goya L, Ramos S. Quercetin induces apoptosis via caspase activation, regulation of Bcl-2, and inhibition of PI-3-kinase/Akt and ERK pathways in a human hepatoma cell line (HepG2) J Nutr. 2006;136:2715–2721. doi: 10.1093/jn/136.11.2715. [DOI] [PubMed] [Google Scholar]
- 11.Phan TT, See P, Tran E, Nguyen TT, Chan SY, Lee ST, Huynh H. Suppression of insulin-like growth factor signalling pathway and collagen expression in keloid-derived fibroblasts by quercetin: its therapeutic potential use in the treatment and/or prevention of keloids. Br J Dermatol. 2003;148:544–552. doi: 10.1046/j.1365-2133.2003.05174.x. [DOI] [PubMed] [Google Scholar]
- 12.Vijayababu MR, Kanagaraj P, Arunkumar A, Ilangovan R, Aruldhas MM, Arunakaran J. Quercetin-induced growth inhibition and cell death in prostatic carcinoma cells (PC-3) are associated with increase in p21 and hypophosphorylated retinoblastoma proteins expression. J Cancer Res Clin Oncol. 2005;131:765–771. doi: 10.1007/s00432-005-0005-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Senthilkumar K, Elumalai P, Arunkumar R, Banudevi S, Gunadharini ND, Sharmila G, Selvakumar K, Arunakaran J. Quercetin regulates insulin like growth factor signaling and induces intrinsic and extrinsic pathway mediated apoptosis in androgen independent prostate cancer cells (PC-3) Mol Cell Biochem. 2010;344:173–184. doi: 10.1007/s11010-010-0540-4. [DOI] [PubMed] [Google Scholar]
- 14.Giuliani C, Noguchi Y, Harii N, Napolitano G, Tatone D, Bucci I, Piantelli M, Monaco F, Kohn LD. The flavonoid quercetin regulates growth and gene expression in rat FRTL-5 thyroid cells. Endocrinology. 2008;149:84–92. doi: 10.1210/en.2007-0618. [DOI] [PubMed] [Google Scholar]
- 15.Murphy EA, Davis JM, McClellan JL, Carmichael MD. Quercetin's effects on intestinal polyp multiplicity and macrophage number in the Apc (Min/+) mouse. Nutr Cancer. 2011;63:421–426. doi: 10.1080/01635581.2011.535954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Conti CJ, Fries JW, Viaje A, Miller DR, Morris R, Slaga TJ. In vivo behavior of murine epidermal cell lines derived from initiated and noninitiated skin. Cancer Res. 1988;48:435–439. [PubMed] [Google Scholar]
- 17.Satyamoorthy K, Li G, Vaidya B, Patel D, Herlyn M. Insulin-like growth factor-1 induces survival and growth of biologically early melanoma cells through both the mitogen-activated protein kinase and beta-catenin pathways. Cancer Res. 2001;61:7318–7324. [PubMed] [Google Scholar]
- 18.Takeuchi K, Ito F. Receptor tyrosine kinases and targeted cancer therapeutics. Biol Pharm Bull. 2011;34:1774–1780. doi: 10.1248/bpb.34.1774. [DOI] [PubMed] [Google Scholar]
- 19.Gallagher EJ, LeRoith D. Minireview: IGF, insulin, and cancer. Endocrinology. 2011;152:2546–2551. doi: 10.1210/en.2011-0231. [DOI] [PubMed] [Google Scholar]
- 20.Vasanthi HR, Mukherjee S, Das DK. Potential health benefits of broccoli- a chemico-biological overview. Mini Rev Med Chem. 2009;9:749–759. doi: 10.2174/138955709788452685. [DOI] [PubMed] [Google Scholar]
- 21.Slimestad R, Fossen T, Vågen IM. Onions: a source of unique dietary flavonoids. J Agric Food Chem. 2007;55:10067–10080. doi: 10.1021/jf0712503. [DOI] [PubMed] [Google Scholar]
- 22.Caltagirone S, Rossi C, Poggi A, Ranelletti FO, Natali PG, Brunetti M, Aiello FB, Piantelli M. Flavonoids apigenin and quercetin inhibit melanoma growth and metastatic potential. Int J Cancer. 2000;87:595–600. doi: 10.1002/1097-0215(20000815)87:4<595::aid-ijc21>3.0.co;2-5. [DOI] [PubMed] [Google Scholar]
- 23.Patel R, Krishnan R, Ramchandani A, Maru G. Polymeric black tea polyphenols inhibit mouse skin chemical carcinogenesis by decreasing cell proliferation. Cell Prolif. 2008;41:532–553. doi: 10.1111/j.1365-2184.2008.00528.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lu G, Liao J, Yang G, Reuhl KR, Hao X, Yang CS. Inhibition of adenoma progression to adenocarcinoma in a 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone-induced lung tumorigenesis model in A/J mice by tea polyphenols and caffeine. Cancer Res. 2006;66:11494–11501. doi: 10.1158/0008-5472.CAN-06-1497. [DOI] [PubMed] [Google Scholar]
- 25.Eming SA, Snow RG, Yarmush ML, Morgan JR. Targeted expression of insulin-like growth factor to human keratinocytes: modification of the autocrine control of keratinocyte proliferation. J Invest Dermatol. 1996;107:113–120. doi: 10.1111/1523-1747.ep12298351. [DOI] [PubMed] [Google Scholar]
- 26.Werner H, Le Roith D. New concepts in regulation and function of the insulin-like growth factors: implications for understanding normal growth and neoplasia. Cell Mol Life Sci. 2000;57:932–942. doi: 10.1007/PL00000735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Reiss K, Wang JY, Romano G, Furnari FB, Cavenee WK, Morrione A, Tu X, Baserga R. IGF-I receptor signaling in a prostatic cancer cell line with a PTEN mutation. Oncogene. 2000;19:2687–2694. doi: 10.1038/sj.onc.1203587. [DOI] [PubMed] [Google Scholar]
- 28.Valentinis B, Romano G, Peruzzi F, Morrione A, Prisco M, Soddu S, Cristofanelli B, Sacchi A, Baserga R. Growth and differentiation signals by the insulin-like growth factor 1 receptor in hemopoietic cells are mediated through different pathways. J Biol Chem. 1999;274:12423–12430. doi: 10.1074/jbc.274.18.12423. [DOI] [PubMed] [Google Scholar]
- 29.Hyun T, Yam A, Pece S, Xie X, Zhang J, Miki T, Gutkind JS, Li W. Loss of PTEN expression leading to high Akt activation in human multiple myelomas. Blood. 2000;96:3560–3568. [PubMed] [Google Scholar]
- 30.Cantley LC, Neel BG. New insights into tumor suppression: PTEN suppresses tumor formation by restraining the phosphoinositide 3-kinase/AKT pathway. Proc Natl Acad Sci U S A. 1999;96:4240–4245. doi: 10.1073/pnas.96.8.4240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Wang SI, Puc J, Li J, Bruce JN, Cairns P, Sidransky D, Parsons R. Somatic mutations of PTEN in glioblastoma multiforme. Cancer Res. 1997;57:4183–4186. [PubMed] [Google Scholar]
- 32.Erlund I, Freese R, Marniemi J, Hakala P, Alfthan G. Bioavailability of quercetin from berries and the diet. Nutr Cancer. 2006;54:13–17. doi: 10.1207/s15327914nc5401_3. [DOI] [PubMed] [Google Scholar]
- 33.Rupasinghe HP, Ronalds CM, Rathgeber B, Robinson RA. Absorption and tissue distribution of dietary quercetin and quercetin glycosides of apple skin in broiler chickens. J Sci Food Agric. 2010;90:1172–1178. doi: 10.1002/jsfa.3944. [DOI] [PubMed] [Google Scholar]
- 34.Koli R, Erlund I, Jula A, Marniemi J, Mattila P, Alfthan G. Bioavailability of various polyphenols from a diet containing moderate amounts of berries. J Agric Food Chem. 2010;58:3927–3932. doi: 10.1021/jf9024823. [DOI] [PubMed] [Google Scholar]
- 35.Santos MR, Rodríguez-Gómez MJ, Justino GC, Charro N, Florencio MH, Mira L. Influence of the metabolic profile on the in vivo antioxidant activity of quercetin under a low dosage oral regimen in rats. Br J Pharmacol. 2008;153:1750–1761. doi: 10.1038/bjp.2008.46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Agullo G, Gamet-Payrastre L, Manenti S, Viala C, Rémésy C, Chap H, Payrastre B. Relationship between flavonoid structure and inhibition of phosphatidylinositol 3-kinase: a comparison with tyrosine kinase and protein kinase C inhibition. Biochem Pharmacol. 1997;53:1649–1657. doi: 10.1016/s0006-2952(97)82453-7. [DOI] [PubMed] [Google Scholar]
- 37.Hagiwara M, Inoue S, Tanaka T, Nunoki K, Ito M, Hidaka H. Differential effects of flavonoids as inhibitors of tyrosine protein kinases and serine/threonine protein kinases. Biochem Pharmacol. 1988;37:2987–2992. doi: 10.1016/0006-2952(88)90286-9. [DOI] [PubMed] [Google Scholar]
- 38.Gamet-Payrastre L, Manenti S, Gratacap MP, Tulliez J, Chap H, Payrastre B. Flavonoids and the inhibition of PKC and PI 3-kinase. Gen Pharmacol. 1999;32:279–286. doi: 10.1016/s0306-3623(98)00220-1. [DOI] [PubMed] [Google Scholar]
- 39.Yoshizumi M, Tsuchiya K, Kirima K, Kyaw M, Suzaki Y, Tamaki T. Quercetin inhibits Shc- and phosphatidylinositol 3-kinase-mediated c-Jun N-terminal kinase activation by angiotensin II in cultured rat aortic smooth muscle cells. Mol Pharmacol. 2001;60:656–665. [PubMed] [Google Scholar]
- 40.Ranelletti FO, Ricci R, Larocca LM, Maggiano N, Capelli A, Scambia G, Benedetti-Panici P, Mancuso S, Rumi C, Piantelli M. Growth-inhibitory effect of quercetin and presence of type-II estrogen-binding sites in human colon-cancer cell lines and primary colorectal tumors. Int J Cancer. 1992;50:486–492. doi: 10.1002/ijc.2910500326. [DOI] [PubMed] [Google Scholar]
- 41.Ashida H, Fukuda I, Yamashita T, Kanazawa K. Flavones and flavonols at dietary levels inhibit a transformation of aryl hydrocarbon receptor induced by dioxin. FEBS Lett. 2000;476:213–217. doi: 10.1016/s0014-5793(00)01730-0. [DOI] [PubMed] [Google Scholar]