

Summary
The plasma membrane in eukaryotic cells contains microdomains that are enriched in certain glycosphingolipids, gangliosides, and sterols (such as cholesterol) to form membrane/lipid rafts (MLR). These regions exist as caveolae, morphologically observable flask-like invaginations, or as a less easily detectable planar form. MLR are scaffolds for many molecular entities, including signaling receptors and ion channels that communicate extracellular stimuli to the intracellular milieu. Much evidence indicates that this organization and/or the clustering of MLR into more active signaling platforms depends upon interactions with and dynamic rearrangement of the cytoskeleton. Several cytoskeletal components and binding partners, as well as enzymes that regulate the cytoskeleton, localize to MLR and help regulate lateral diffusion of membrane proteins and lipids in response to extracellular events (e.g., receptor activation, shear stress, electrical conductance, and nutrient demand). MLR regulate cellular polarity, adherence to the extracellular matrix, signaling events (including ones that affect growth and migration), and are sites of cellular entry of certain pathogens, toxins and nanoparticles. The dynamic interaction between MLR and the underlying cytoskeleton thus regulates many facets of the function of eukaryotic cells and their adaptation to changing environments. Here, we review general features of MLR and caveolae and their role in several aspects of cellular function, including polarity of endothelial and epithelial cells, cell migration, mechanotransduction, lymphocyte activation, neuronal growth and signaling, and a variety of disease settings.
Keywords: membrane/lipid rafts, caveolae, caveolin, cytoskeleton, signaling receptors, ion channels
Introduction
The plasma membrane is a dynamic entity. It is both a barrier that separates the extracellular and intracellular environments and a structure composed of proteins and lipids that controls and is controlled by many biological processes. Hydrophobic moieties of lipids self-associate while their hydrophilic regions interact with the aqueous environment (in both the extracellular and intracellular milieus) to create the physical basis of the plasma membrane bilayer[1, 2]. This amphipathicity of lipids is essential for separating internal cellular structures from the external environment. Eukaryotic plasma membranes are composed of glycerophospholipids, sphingolipids, and sterols (in particular, cholesterol). The head group of the glycerophospholipids varies, as can the length and degree of saturation of the associated fatty acyl chains. Sphingolipids also vary in their ceramide backbones and the >500 different carbohydrate structures in the head groups. The introduction of cholesterol to the plasma membrane appears to have occurred later in evolution than that of certain other membrane lipids; its presence coincides with the increase in environmental oxygen concentration that occurred ~2.5 billion years ago[3]. The presence of sphingolipids and sterols increases the complexity of eukaryotic membranes and distinguishes them from the membranes of prokaryotes. Cholesterol increases the thickness and stiffness of lipid bilayers[4] and allows for protein sorting[5]. The hydrophobic effect of the amphipathic molecules on lipid shapes (i.e., inverted cones, cylindrical, or conical) forms lamellar, micellar, or cubic supramolecular structures. Certain lipids of eukaryotic cells are not cylindrical and therefore not predicted to support the formation of a membrane (lamellar) bilayer; however, the inclusion of proteins in membranes results in macromolecular assemblies and bilayer asymmetries that help promote the formation of the lamellar membrane bilayer.
Forty years ago Singer and Nicolson described the plasma membrane (PM) as having a `fluid mosaic' environment that randomly partitions proteins and lipids so as to achieve the lowest free energy[6]. Substantial subsequent evidence has revealed that this partitioning of proteins is not homogenous and random but instead consists of clusters of structural proteins (e.g., integrins and intracellular scaffolds), enzymes, signaling receptors, transporters and channels within lipid domains; some of these lipid domains are enriched in cholesterol and certain saturated acyl lipids and are termed lipid or membrane rafts[7–11]. These membrane domains and their unique protein and lipid content are critical for many cellular functions. Along with clathrin-coated pits, membrane/lipid rafts (MLR) are structurally and functionally distinct, important regions of the PM[10].
There are two major types of MLR: those that contain the cholesterol binding protein caveolin (Cav) and those that do not. Cav-containing MLR form morphologically distinct entities, caveolae (“little caves”), flask-like invaginations of the PM (detectable at the resolution of electron microscopy) while MLR that lack Cav are flat and not identifiable by electron microscopy. However, some cells, such as neurons and lymphocytes, that express Cav and contain lipid rafts do not have morphologically identifiable invaginated structures[12]. It remains unclear why such cells express Cav but do not form caveolae.
Interactions between MLR and cytoskeletal components can contribute to the regulation of MLR assembly/clustering and cytoskeletal dynamics[13, 14]. Although the association between cytoskeletal components and MLR/caveolae had been previously described[15, 16], recent evidence has extended the notion that cytoskeletal components (e.g., actin, tubulin, vinculin, filamin, and tau)[17, 18] can localize to MLR and be platforms for cytoskeletal tethering and for communication to the extracellular matrix (ECM) via integrins, cadherins, occludins, and other cellular adhesion molecules (CAMs). Moreover, MLR can cluster and this clustering may depend upon cholesterol and actin tethering to the membrane[19]. Kusumi and colleagues proposed a `picket-fence' model, whereby actin filaments anchored to MLR regulate lateral diffusion of membrane proteins and lipids[20, 21]. This transient anchoring of transmembrane proteins with actin filaments was hypothesized to resemble a row of `pickets' that regulate (slow) diffusion of adjacent proteins and lipids. Based on their additional work, Kusumi and colleagues have proposed that the transient `clustering' or coalescing of homodimer rafts forms hetero- and homo-GPI-anchored protein oligomeric rafts, within the inner leaflet, through raft-based lipid interactions that generate functional raft domains [22, 23]. Gowrishankar et al have demonstrated that this nanomicrodomain clustering depends upon cholesterol, sphingolipids and an active cortical actin meshwork[24]. These actin meshwork “snippets” are composed of 250 nm actin filaments cross-linked by myosin motors that facilitate lateral movement of GPI-anchored proteins in an energy-dependent manner. The combination of lateral membrane movement and transmembrane interaction among integrins, membrane bilayer lipids, and membrane proteins within MLR, along with interaction of the actin/myosin cytoskeleton and cytoskeletal tethering partners, can contribute to cellular migration, mechanotransduction, cell growth, endothelial and epithelial barrier formation, and immune cell activation—physiologically important responses and ones that can be altered in disease settings.
MLR thus serve as regulators of numerous cellular events, including: 1) cellular polarity and organization of trafficking and sorting mechanisms, 2) formation of platforms for ECM adhesion and intracellular cytoskeletal tethering to the PM (intracellular-extracellular skeletal linkage, in particular via integrins, and lateral membrane clustering), and 3) transduction of signaling cascades across the PM (“outside-in” signaling), which in turn can rearrange cytoskeletal architecture and alter cell growth, migration, and other functions and 4) entry of viruses, bacteria, toxins and nanoparticles. The remainder of this review focuses on MLR-cytoskeletal interactions that influence signaling receptors and channels within the PM and emphasizes articles published in the past 5 years (Table).
Table.
Examples of cell types, raft/cytoskeletal/receptor/channel interactions and raft-regulated cellular functions.
| Cell Type | Raft-associated Receptors, channels, and cytoskeletal components | Raft-mediated cell function | References |
|---|---|---|---|
| Endothelial cells | (1) eNOS, Src, P-Cav-1, p190RhoGAP, TRPC1, IP2R3, β1-integrin, VEGFR2,annexin, Ras, | (1) Mechanotransduction | [99–116] |
| (2) α(3)β(v)-integrins-VEGFR2 coupling | (2) EC adherence and angiogenesis: | ||
| (3) Cav-1, ICAM1, adherens; filamin A eNOS, dynamin | (3) Endothelial barrier function | ||
| (4) P- Cav-1, Src, filamin A PKCa, actin, GTPase dynamin | (4) Endocytosis and trafficking | ||
|
| |||
| Epithelial cells | (1) GM1, GM3, prominin, Cav-1, PTRF, Rho, TRPC1, AQP5 | (1) Apical membrane polarity/signaling | [117–128] |
| (2) Cav-1, PTRF (cavin), tetraspanin, claudin-2, occludin, cadherins, actin, connexins,, integrins, talins, paxillin, FAK, alpha actinin | (2) Lateral adherence and barrier function: tight (TJ), adherens (AJ), gap junctions (GJ) | ||
| (3) Cav-1, dynamin 2, NKCC2, Rab11a, tubulin | (3) Endocytosis and trafficking | ||
|
| |||
| Striated myocytes (cardiac, skeletal) | (1) Cav-3, β-AR, α-AR, P2X7R, G proteins, AC, RyR, TRPC, SAC, K channels, filamin A β-DG | (1) Mechanotransduction and contractility | [42, 136–153] |
| (2) AC, actin, Cav-3, filamin, tubulin | (2) Cytoskeletal tethering | ||
|
| |||
| Smooth muscle cells | (1) Cav-1, NCX1, SERCA2, β-DG, RhoA | (1) Mechanotransduction and contractility | [112–115] |
| (2) Cav-1, TRPV4, TPRC1, K_2.3, K_1.1, K selective channels | (2) Vasculartone | ||
| (3) Cav-1, TNFR-associated factor 6, p38 MAPK, p190RhoGAP, actin | (3) Migration/Invasion | ||
|
| |||
| Neurons | (1) TrkA GM1, cholesterol, tubulin, Cav-1, NMDAR, Src | (1) Neuritic growth/dendritic-axonal arborization | [14, 17, 76, 93, 154–178] |
| (2) β1-integrin, tetraspanin, Glu A2/3, AMPAR, PI4Ktype II, actin | (2) Post synaptic density composition | ||
| (3) Reggie/flottilins, ganglioside GD3, PKC, GAP43,, Rad1, Cadherins, Src, TRPC5, CaMKK, CaMKIg | (3) Growth cone formation/advancement | ||
| (4) Cav-1 and VGCC | (4) Membrane architecture and Ca2+ regulation | ||
| (5) Cav-1, Cav-3, ERa/ERb, mGlur1, mGluR2/3 | (5) Neuronal membrane receptor cross-talk and internalization | ||
|
| |||
| Lymphocytes | (1) actin | (1) Mechanotransduction | [10, 154, 179–186] |
| (2) α(4) α(5) β1-integrin, fibronectin, TCR, CCR6, tetraspanin, SFK, Rho, actin, GM1 ganglioside, myosin 1c, MHC-II | (2) Immunologic synapse formation | ||
|
| |||
| Erythrocytes, platelets | (1) Rac, tubulin | (1) Erythroblast enucleation | [187–189] |
| (2) Glycoprotein (GP) IB-IX-V, Lyn, filamin, prion, actin, | (2) platelet adherence to damaged endothelium | ||
|
| |||
| Pathologies | (1) TET, viral proton channel M2, actin, Dyrk3, tubulin | (1) Infectious agents | [166, 190–226] |
| (2) Src, Cav-1, PTRF (Cavin), Met/EGFR, b-integrins, CD24, P2Y2, VEGFR, KCC4, myosin | (2) Cancer | ||
| (3) Flotillin, Cav-1, ER, VDAC, γ-secretase, APP, Aβ | (3) APP processing and Alzheimer's Disease | ||
Methods for the Isolation and Enrichment of Raft Domains
A subset of MLR, termed caveolae (“little caves”), were first observed microscopically by Palade and Yamada[25, 26], however the concept of lipid domains was formalized by Karnovsky et al. [27]. These domains were subsequently termed glycolipid-enriched membranes by Parton and colleagues [28] and then known as detergent-resistant membranes (originally coined by Baird and colleagues [29] based on their insolubility to detergents, in particular nonionic detergents at 4°C[30]), the latter property being attributable to the high content of saturated acyl chains [31, 32]. MLR were termed lipid domains[27] and noted to be enriched in cholesterol, sphingolipids, and lipids with saturated acyl chains. These constituents result in MLR being resistant to membrane disruption by nonionic detergents and having a low density. As a result, one can isolate MLR based on detergent-resistance and subsequent fractionation on density gradients[31, 32]. Early experiments isolated MLR with 1% Triton X-100 followed by sucrose density gradient fractionation[31]; later studies employed other detergents including Lubrol, Brij, Nonidet, CHAPS, and octylglucoside at varying concentrations[14, 33]. However, concerns arose that use of such detergents may produce a `false' clustering of raft lipids with proteins, interactions that do not exist in cells[33].
Along with techniques used for cell lysis, the method by which rafts are enriched is just as critical. Song et al. introduced a non-detergent, high pH (sodium carbonate) isolation method[34], followed by a long duration (16–20 h) of centrifugation. However, this method introduced the possibility that “raft fractions” might also include non-MLR membranes. Another approach used an isotonic buffer (0.25 M sucrose) on a Percoll gradient to isolate PMs followed by a discontinous gradient of OptiPrep to generate a much cleaner MLR isolation[35]. Some have suggested that this methodology produces high variability between preparations and among cell types[36]. Other techniques include use of a silica-coating that involves anti-Cav-coated magnetic micropheres to purify Cav-positive PM vesicles from other detergent-insoluble membranes rich in GPI-achored proteins, but devoid of caveolae[37, 38]. Even with this approach, we (and others) caution against potential discrepancies between varying results based upon different fractionation techniques[30, 39] Membrane purification protocols may also isolate organelles that interact with MLR or the endoplasmic reticulum, whose extensive tubular network can survive cell disruption and therefore may be falsely considered as organelle impurity[40]. Evidence is emerging that MLR may reside intracellularly in Golgi-endosomal components and in vesicular cargos, although in the past, detection of such structures was considered evidence of lack of purity of MLR[41]. Lysis buffers that do not sufficiently disrupt the cytoskeleton to properly separate different species of MLR (non-caveolar versus caveolar) can also produce confounding results, especially when performed on cells highly abundant in cytoskeletal architeture, such as adult cardiac myocytes[36, 42]. Although many types of preparations have contributed to current understanding of MLR structure and function, their limitations provide a rationale for the development of improved methodologies that optimize membrane purification and enrichment strategies[33]. For example, improved fractionation methods in combination with morphological approaches, such as super-resolution microscopy that allows for fluorescence imaging with a precision near 20 nm, may prove useful for obtaining more consistent and accurate understanding of MLR organization and function[43].
A major issue related to MLR, and one that has arisen at least in part as a consequence of disagreements regarding methods for their isolation and characterization, concerns their existence and biological relevance[44]. Since most of the PM exists in an ordered phase[45], some suggest a `percolating phase', such that the disordered phase serves as the microdomain[46]. This latter theory, while contrary to the canonical raft hypothesis, may explain clustering of non-raft proteins in the PM and suggests a bridge between rafts and `protein islands'[47]. Additional studies are needed to help resolve the different notions regarding plasma membrane organization and MLR.
MLR lipid constituents: Sphingolipids and cholesterol
MLR are highly enriched in sterols and sphingolipids. Sphingolipids are derived from ceramide, a lipid involved in numerous cellular responses, including differentiation, adherence, migration and cell death[48–50]. Ceramide within the ER is synthesized into sphingolipids or shuttled to the Golgi complex to become gangliosides (in particular, GM1 and GM3). GM, which connotes ganglioside plus one (mono)sialic acid, or GD, wherein the D stands for disialic acid, are complex glycolipids with strong amphiphilic properties due to the saccharidic headgroup and a double-tailed hydrophobic moiety (i.e, ceramide[51]). The numbers 1, 2, and 3 (e.g., GM1, GM2, or GM3) denote the migration of the gangliosides on thin-layer chromatography. The presence of sialic acid distinguishes gangliosides from neutral glycosphingolipds and sulfatides. Although non-Cav-containing MLR exist in planar forms that cannot be readily identified morphologically, MLR can be labeled with fluorescent cholera toxin (CTX) B subunits, which bind to ganglioside GM1[52, 53]. Sphingolipids contain acyl chains that are longer than the width of a single leaflet of the PM[54] and thus, may help facilitate transmembrane events in the absence of transmembrane proteins.
Cholesterol is another major membrane component of MLR. Following its synthesis in the ER, cholesterol is trafficked to the Golgi where it binds sphingolipids or proteins such as Cav[55–59]. These sphingolipid- and cholesterol-enriched structures self-aggregate in the Golgi and generate vesicles that are transported to the PM to form MLR. In the formation of MLR, the sphingolipids associate via their head groups and cholesterol interdigitates between the sphingolipids[60, 61]. Sphingolipids and cholesterol, as well as GPI-anchored proteins, are predominantly found in the outer leaflets (exofacial domains) of the PM, while Cav and associated proteins are found in inner leaflet (cytofacial) domains. Cav and other inner leaflet proteins interact with membrane lipids via myristic and/or palmitic acid, which are present on a number of MLR-localized proteins (e.g., Src and heterotrimeric GTP-binding [G] proteins). Cholesterol binds certain membrane proteins, such as G-protein-coupled receptors (GPCRs), and can influence properties of GPCRs and presumably other membrane proteins[61].
MLR Scaffolds: Caveolin, Cavin, Flotillin/Reggie
Caveolin (Cav), an integral membrane protein that exists in 3 isoforms (Cav-1, -2, and -3), was first discovered as a tyrosine-phosphorylated substrate of Src in transformed chick fibroblasts[62]. This phosphorylation can be mediated by Src, Fyn, Yes, and c-Abl, certain growth factors, and integrin activation[63]. Cav-1 is also phosphorylated on serine 80, an event implicated in trafficking of cholesterol[64, 65]. Cav-1 contains an α-helical hairpin intramembrane domain (IMD; residues 102–134) and palmitoylated cysteines 133, 143, and 156, which are not required for membrane insertion but help facilitate binding of cholesterol and its transport to the membrane[66](Figure 1A). A cholesterol recognition/interaction amino acid consensus (CRAC) motif (residues 94–101, VTKYWFYR) within Cav-1 appears to mediate its binding to cholesterol[67](Figure 1B). Cav-1 also scaffolds and organizes a wide variety of signaling components via a β-stranded Cav scaffolding domain (CSD)[68, 69]. It has been proposed that this 20 amino acid (e.g., amino acid residues 82–101 for Cav-1) CSD is critical for the binding (via a Cav binding domain [CBD]) and regulation of the activity of signaling components[12, 70], although this proposal has been questioned[71, 72]. Originally thought to be negative regulators of signal transduction components[69, 73], Cavs may also activate cellular events, including various signaling proteins. Examples include responses mediated by insulin, Ras, and neurotrophin[74–76]. All three isoforms of Cav contain a conserved structural motif (FEDVIAEP) that allows for their oligomerization within a caveolar coat[77], typically consisting of 12–18 monomers of Cav-1/Cav-2[78, 79] to form a heteroligomer-filamentous structure[80]. Recent data suggest that 9 monomers can form a homoligomer-toroidal shape for Cav-3[81] and for the CSD(Figure 1C).
Figure 1. Schematic depicting proposed caveolin monomer structures and oligomer complexes.

A, Adapted from Fernandez et al. (2002), a model in which the caveolin-1 (Cav-1) scaffolding domain (CSD) is shown as an α-helix (AA 79–96) with Cav-1 oligomers composed of 7 monomers and an approximate diameter of 11 nm. This proposed heptamer forms because α-helical lateral interactions proximal to the cytofacial lipid bilayer give rise to a filamentous assembly 50 nm long. B, An alternative model by Hoop et al. (2012) in which the CSD is a β-strand (red/orange) separated by the wedged shaped α-helix (green barrels) within the cytofacial bilayer by cholesterol (yellow) interacting with a cholesterol recognition/interaction amino acid consensus (CRAC) motif (blue) with palmitoyl acids (brown strands) anchored to cysteine residues. C, Model by Whiteley et al. (2012) in which Cav-3 is arranged with 9 monomers assembled in a toroidal shape ~16.5 nm in diameter and 5.5 nm in height.
Since Cav exists in cells that lack morphological caveolae[12, 82], this raises the question: what causes these unique plasmalemmal invaginations? Recent evidence has shown that caveolar formation appears to depend upon the protein cavin (also known as polymerase I and transcript release factor, PTRF[83]). There are 4 isoforms of cavin: cavin-1 (PTRF), cavin-2 (aka serum deprivation protein response [SDPR]), cavin-3 (aka sdr-related gene product that binds C-kinase [SRBC]), and cavin-4 (aka muscle restricted coiled-coil protein [MURC])[84]. Cavin is a peripheral membrane protein that binds to phosphatidylserine within caveolae[85]. Cavin and Cav exist in a 1:1 stoichiometric ratio in caveolae in close proximity with each other[86]. Cavin is phosphorylated on serine residues 36, 40, 365, and 366[85], possibly by the serine/threonine kinase ARAF1[77]. Cavin appears to be essential for caveolar invagination, i.e. for the formation of morphologic caveolae. Another family of proteins that have been described in MLR are Flotillin-1/reggie-2 and flotillin-2/reggie-1, which were originally discovered as neuronal proteins in retinal ganglion cells during regeneration after injury[87, 88]. Flotillins/reggies oligomerize via their C-terminal tail domains[89] and have been implicated in the endocytosis of GPI-anchored proteins. The planar form, of MLR ranging 1–1000 nm in diameter, is enriched in GPI-anchored proteins and in flotillins. By contrast, caveolae have a diameter of 70–120 nm[25, 90].
Receptor Activation and MLR Clustering
Planar and caveolar MLR can bind proteins. Although no specific amino acid sequence or motif has been identified that targets a particular protein to MLR, certain post-translational modifications such as glycosylphosphatidylinositol (GPI)-anchors, palmitoylation and myristoylation appear to help localize proteins to MLR [91]. One observes a high variability of membrane domains and MLR abundance in different cell types and the MLR can vary in their content of cholesterol and expression of caveolin, which, in turn, influences MLR abundance and function, including in their localization/enrichment of certain membrane proteins, such as signaling receptors[92]. MLR can exist as clusters; alternative models propose that 1) non-MLR receptors translocate into MLR upon ligand binding leading to signal transduction or 2) spatially separate MLRs cluster to one another only after ligand bindng and receptor activation[93]. Other models propose that clustering can occur via inner leaflet interactions (facilitated by flotillins, Cavs, and annexins) or outer leaflet clustering through GPI-anchored proteins. Clustering of MLR can alter their function, for example, creating signaling platforms[10, 94, 95]. Caveolae can undergo endocytosis (via dynamin-II) to become intracellular organelles, termed caveosomes[96], which influence functional activities, such as in mechanotransduction to modulate blood flow and vascular tone[97]. Although the model proposed by Gowrishankar et al[24] demonstrates that lateral membrane clustering of microdomains depends upon cholesterol, sphingolipids, and an active actin/myosin network, this model can also account for MLR-mediated endocytosis and exocytosis, and membranes that lack sterols, such as in C. elegans[98].
Cellular polarity: Endothelium and Epithelium as examples
A monolayer of endothelial cells (endothelium) lines the vasculature as an initial barrier between bloodborne components (e.g., oxygen, nutrients, cells) and tissue parenchyma. Caveolae are higher in abundance in lung vascular endothelial cells than in any other mammalian cell type[99]. Caveolae concentrate proteins involved in vesicular trafficking (i.e., formation, fission/fusion, and docking), as well as signaling receptors that include receptor tyrosine kinases (RTK), GPCRs, G proteins and downtream enzymes such as adenylyl cyclases (ACs) and endothelial nitric oxide synthase (eNOS, NOS3), all of which co-localize with Cav-1 in caveolae of endothelial cells (Figure 2). In addition, endothelial caveolae sense alterations in shear stress and help mediate mechanotransduction in response to blood flow, i.e. changes in vascular contractile tone and reactivity to various stimul. Caveolae also contribute to intracellular calcium and redox signaling, and macromolecular transcytosis in endothelial cells[100].
Figure 2. Membrane/lipid rafts and the endothelium barrier.
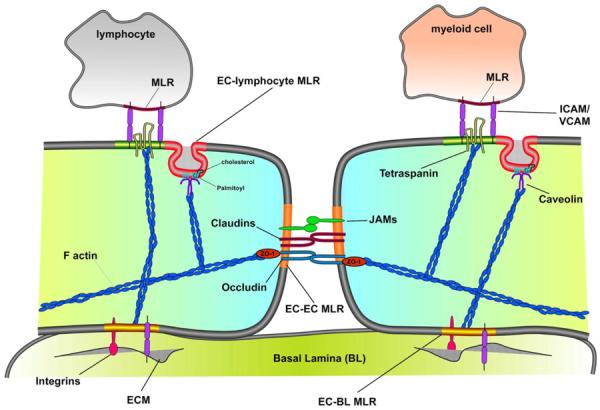
Schematic depicting different types of MLR and how they serve to regulate endothelial cell (EC) morphology, adherence, and function. Scaffolding rafts secure the ECs to their surrounding environment [i.e., EC-EC adherence, EC-BL (basal lamina)adherence, and EC-lymphocyte adherence]. Integrins and cellular adhesion molecules(CAM) work in concert to bind to the extracellular matrix (ECM) and establish the adhesion raft with underlying BL. Occludins, claudins, zona occludins 1(ZO-1), and junctional adhesion molecules (JAMs) form the scaffolding raft of the interendothelial environemnt. Tetraspanins, caveolins/caveolae, and ICAM/VCAM (inter/vascular CAMs) form a lymphocyte adhesion raft (LAR), which facilitates lymphocyte recruitment and migration across the endothelium.
Numerous processes contribute to lymphocyte adherence and migration across the endothelium[101]. In order to facilitate lymphoctye activation and migration, the endothelial PM works in concert with the underlying cytoskeleton[102, 103]. Many lines of evidence suggest that lipid reorganization occurs during this event, which affects MLR dynamics and in turn, the recruitment of key adhesion molecules, junctional proteins, and transporters. This apical PM reorganization works in concert with the underlying actin cytoskeleton to mediate the morphological changes needed for lymphocyte migration[15, 104]. Some studies suggest that this interaction occurs between adhesion molecules such as cadherins and the actin cytoskeleton via catenins after a series of events regulated by Rho GTPases[102, 103]. MLR also organize additional components that facilitate intercellular adhesion and integrity to initiate leukocyte transcellular transport and events such as AMPKa1 activation, cadherin signaling[105, 106] and transport of low-density lipoproteins and chemokines across the microvasculature[107, 108].
Endothelial cell Cav-1 in MLR regulate other events involved in endothelial cell polarity and barrier function[109], including nitric oxide production/signaling and Src phosphorylation[110], Cav-1 and eNOS-mediated activation of p190RhoGAP-1[111], and Cav-1 regulation of TRPC1 (transient receptor potential cation channel 1), IP3 receptor signaling and calcium entry (via Ca2+ store release-induced Ca2+ entry)[112]. In addition to NOS signaling pathways[113], other effector enzymes (e.g., AC) as well as actin binding proteins, such as filamin, contribute to endothelial barrier function in a Cav-1 dependent manner[105].
Interactions of caveolae with cytoskeletal components regulate endothelial trafficking and endocytosis. Caveolae are closely associated with actin fibers and can internalize membrane vesicles to Rab11 endosomes[114]. Actin-binding proteins such as filamin A facilitate Cav-1 internalization in a PKCα-dependent manner during cellular detachment. Other data show that caveolar endocytosis is controlled by actin; Src-induced phosphorylation of Cav-1 increases the association of filamin A with Cav-1 and subsequent caveolar-mediated transport and vesicular internalization[115]. Regulation of caveolar endocytosis can also occur by intersectin 2L, a guanine nucleotide exchange factor for the Rho GTPase Cdc42[116]. Thus, the regulation of actin cytoskeletal-mediated endocytosis depends upon Cav-1, at least in part, by its post-translational modifications.
Epithelial cells provide an interface between an organism and its outside environment and regulate entry of ions and nutrients. The apical membrane of epithelial cells contain MLR that are enriched in α-and β-integrins that adhere to components of the ECM (including collagen, laminins, fibronectin) and that communicate with the actin cytoskeleton via talins, paxillin, and focal adhesion kinase (FAK)[117, 118](Figure 3). MLR clustering of signaling complexes is crucial for formation of apical membrane microdomains that have GM1 within microvilli and in basal membranes and GM3 at the apical base[119]. Such MLR contain proteins that include prominin, a cholesterol-binding pentaspan glycoprotein[120] and Cav-1. Cav-1 is necessary for a variety of types of adherence in epithelial cells, including recruitment of and interaction with protein components of tight, gap, and adherent junctions (TJ, GJ, AJ)[121]. Cav-1 interacts with claudins and occludins in apical and lateral membrane MLR. These polarized membrane regions are necessary for restricting paracellular transport and depend upon actin-alpha actinin interactions. Within AJ zona occludins establish links to the tubulin-based cytoskeleton, as well as actin; Cav-1 appears to be necessary for retrieval of these junctional proteins[122]. Caveolar-type MLR are also involved in endocytosis by epithelial cells[123], which in some instances involves Rab11 and the tubulin-based cytoskeleton[124]. Additional findings indicate that basolateral membranes may contain different Cavs[125, 126] and lipid components[127, 128] than those of the apical membranes of epithelial cells. These differences in protein and/or lipid components may contribute in part to the distinct polarities within the same cell.
Figure 3. Membrane/lipid rafts and the epithelium barrier.
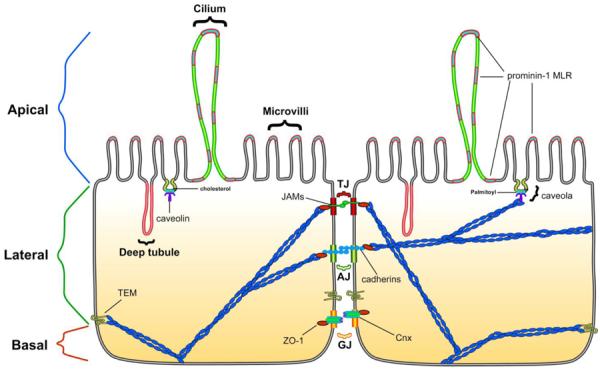
Schematic illustrating the role of membrane/lipid rafts (MLR) in epithelial apical-basal polarity. MLR form unique plasmalemma outward (cilium and microvilli) and invaginations (deep tubules). MLR and associated scaffolds also establish tight junctions (TJ), adherent junctions (AJ), gap junctions (GJ), tetraspaninenriched microdomains (TEM), caveolin-enriched microdomains/caveolar membranes. Scaffolding and cholesterol-binding proteins such as caveolin and prominin as well as junctional adhesion molecules (JAM), zona occludins (ZO), cadherins, and connexins all participate as signaling platforms, sites for intercellular adherence and for actin cytoskeletal tethering in order to create a barrier from the outside environment but also regions that deliver molecules, nutrients, and ions into cells and ultimately to host organisms.
MLR-cytoskeletal tethering: Migration and Mechanotransduction
PM signaling platforms facilitate ECM adhesion and cytoskeletal tethering to the membrane. These interactions are necessary for cell migration, especially in the setting of tissue remodeling, and require polarized signaling complexes within MLR and actin components[129]. For example, MLR-actin interactions lead to pseudopod protrusion and phagocyte migration following nucleotide-mediated chemotaxis[130]. Cav-1 is involved in endothelial migration, organizing pro-angiogenic signaling components with MAP kinases, Src and TNF receptor-associated factor 6[131]. In the brain, Cav-1 is phosphorylated in podosomes that permit microglial invasion, a migratory mechanism similar to that exhibited by invadosomes in peripheral tissues[132]. While some cells use MLR to faciliate signaling between the ECM and intracellular cytoskeletal components and as a result, induce morphological changes necessary for detachment and migration, other cells use such platforms to cause contraction. For example, lymphocyte mechanotransduction occurs through stretch-activated cation channels and subsequent actin rearrangement in MLR[133, 134], while endothelial mechanotransduction depends in part upon integrin/P-Cav-1 signalng events[63, 97, 135].
Localization of receptors and channels in MLR helps regulate cytoskeletal events necessary for contraction of smooth and striated muscle myocytes[136–138]. In the heart, contraction of cardiac myocytes leads to cardiac emptying and is initiated by the movement of ions across the plasma membrane (sarcolemma) via several types of ion channels [i.e., , K+, KATP, KCa, TRPC channels, voltage-gated Ca2+channels, VGCC,and ryanodine receptor (RyR)][136–139] that cluster within Cav-3-enriched MLR. Cav-3 scaffolds the β-adrenergic receptor-Gαs-AC signaling cascade, contributing to the generation of cAMP[42, 140]. In addition, Cav-3 interacts with the sarcoplasmic reticulum Ca2+channel, RyR, which upon activation, increases Ca2+ release into the cytosol[141]. Caveolae also organize other cardiac myocyte receptors, G proteins, channels, and actin binding components[13, 142–145] and proteins that mediate cell-cell adherence[146].
Skeletal muscle contraction depends upon Cav-3-enriched MLR and interations with cytoskeletal components. For example, depletion of cholesterol from skeletal myocyte membranes disrupts β-dystroglycan interaction with Cav-3 and weakens contraction[147]. Akin to what occurs in cardiac myocytes, Cav-3 forms microdomain complexes with RyR and VGCC in transverse tubule membranes in addition to ones with other Ca2+ channels [148], interactions that regulate Ca2+ flux and excitation-contraction coupling[137, 149]. Cavs also organize ion channels (e.g., TRPC, K+, Na+ and Ca2+ channels) and interact with cytosketal components involved in smooth muscle contraction and vascular tone[150–153].
Neurons: Implications for pro-growth signaling and neuronal membrane repair
Many studies have examined factors that promote neuronal development and regeneration, especially in the setting of injury or neurodegeneration. The neuronal membrane, in particular neuronal MLR, are critical for the initiation of growth in response to extracellular cues. This role derives from the organization in MLR of complexes involved in cellular polarity and pro-survival and pro-growth receptor signaling necessary for neuronal sprouting (Figure 4). Accumulating evidence indicates that pre- and post-synaptic proteins essential for neuronal communication localize to MLR [93]. The localization of receptors, proteins, and signaling molecules to MLR partially depends upon scaffolding and cholesterol binding proteins such as Cav-1, flotillin-2 (Flot-2), and tetraspanin-7 (TSPN-7)[76, 154, 155]. Examples include GPCRs, RTKs, ion channels, G proteins, ACs and Rho GTPase family members, which have been shown to be clustered together by Cav-1 and Flot-2, thereby facilitating high fidelity signaling involved in the regulation of neuronal responses. However, in addition to scaffolding proteins, the coalescence of smaller scale MLR into a larger rafts through lateral membrane distibution can also occur due to a lipid-lipid or protein-lipid interactions. These events induce cholesterol enrichment and also lead to recruitment of transmembrane, exofacial and cytofacial lipid-anchored proteins[156]. The lateral distribution of these rafts into clusters can be stablized due to their affinity for existing raft domains; such interactions can enhance the segration of those components with weak MLR affinity (i.e, transferrin receptor)[156].
Figure 4. Membrane/lipid rafts, growth cone advancement and axonal guidance.
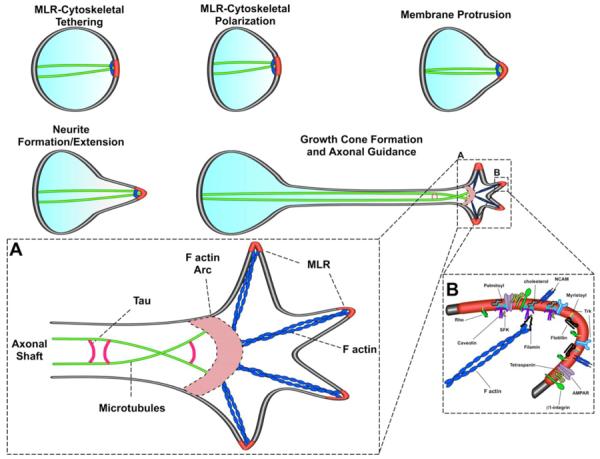
Series of schematics depicting MLR (red PM) at the leading edge of a neuronal growth cone. This polarity and forward migration of the axonal growth cone cannot occur without MLR and the underlying actin (blue) and tubulin (green) cytoskeleton. Inset A shows a closer illustration of the growth cone with MLR at the leading tips receiving extracellular guidance cues and also tethering and transducing those cues to the filamentous (F-) actin. The axonal microtubule (MT)-associated protein Tau cross-links adjacent MT to maintain axonal integrity and facilitate guidance. Inset B shows a close up of several important MLR signaling receptors and enzymes (AMPAR, Trk, SFK, Rho GTPases), scaffolds (caveolin, flotillin, tetraspanin), and adhesion molecules (integrins, NCAM or neuronal cellular adhesion molecule). F-actin binding proteins such as filamin anchor F-actin to MLR scaffold proteins.
MLR are at the leading edge of growth cones and promote neurite extension and subsequent axonal guidance[157–160]. Disruption of MLR perturbs their interaction with microtubule proteins and causes neurite retraction[17]. In addition to receptors and ion channels, cyclic nucleotides are involved in growth of neurites: cAMP promotes axonal growth and inhibits dendritic growth while cGMP promotes dendritic growth and inhibits axonal growth[161]. The cAMP effector Epac mediates cAMP-promoted axonal attraction, while cAMP acting via PKA induces axonal repulsion[162]. Although more work is needed to identify if distinct cyclic nucleotide `pools' promote neurite growth and axonal/dendritic guidance that orignate from neuronal MLR, AC8, a neuron-specific isoform, localizes to MLR, and can tether the actin cytoskeleton to rafts[163]. Evidence does not yet exist for Cav-1/AC 8 interactions in neuronal MLR but Cav-1 and membrane cholesterol can regulate a variety of neuronal receptors and channels (e.g.,VGCC that promote cAMP production and Ca2+ signaling[76, 164]).
Cav-1 has a direct effect on various neuronal receptors. Cav-1 enhances NMDAR-Src signaling and neuroprotection while loss of Cav-1 blunts the ability of NMDAR-Src to protect from oxygen/glucose deprivation and ischemic insult[165, 166]. Cav-1 also regulates AMPAR signaling and turnover[82] and mGluR signaling and internalization in hippocampal and striatal neurons[82, 167–169]. Cavs have also been implicated in the effects on membrane estrogen receptor (ER) signaling in neurons[168]. While there is evidence that Cav-1 promotes crosstalk between ERα and mGluR1, the “muscle isoform”, Cav-3, mediates ERα/ERβ crosstalk with mGLuR2/3[170]. It remains to be determined whether this cross-talk results from cytoskeletal-mediated clustering of MLR or from neurotransmitter signaling and changes in membrane conductance. Neurons lack morphologic caveolae but Cav isoforms and MLR seem to regulate synpatic receptors, channels, and components of the actin cytoskeleton, which in turn facilitate neuronal growth and arborization[76].
Besides Cav, other MLR scaffolding proteins (e.g., Flot-2 and TSPN7) and lipids (e.g., gangliosides) contribute to the organization of neuronal signaling components that regulate synpatic function and plasticity[154, 155]. As noted above, Flot is a cytofacial leaflet scaffolding and cholesterol binding protein that is up-regulated in regenerating optic nerve axons[154]. Flot clusters prion proteins (PrP) and amyloid precursor proteins (APP) at the membrane prior to clathrin-dependent endocytosis[171]. Flot contributes to the clustering of the tyrosine kinases Src, Fyn, and Lck in addition to Rho-GTPases (e.g., Rac, Rho, and Cdc42) into active signaling platforms, potentially facilitating axonal regeneration[172–175]. Flots also localize proteins, such as adhesion molecules (e.g., cadherins and integrins) and FAK, to the leading edge of neurites; such proteins help alter the cytoskeletal organization required for neuronal growth. More recent work has demonstrated a physical association of MLR with neuronal membrane postsynaptic densities, which included cytoskeletal (actin) and cytoskeletal-binding components such as drebrin, fodrin, shank, and homer[14]. Along with Cav-1 and Flot, TSPN-enriched MLR are also critical for neuronal growth[155]. TSPN7 promotes filopodial and dendritic spine formation in hippocampal neurons through interactions with the PDZ domain of PICK1 (protein interacting with C kinase 1), GluR2/3, and AMPAR. TSPN7 regulates PI4K type II activity and its association with actin filaments, facilitates β1-integrin recruitment, and modulates AMPA receptor currents and trafficking, which is crucial for synaptic plasticity, learning and memory[176]. Non-protein components of MLR, such as gangliosides and cholesterol, activate the pro-growth neurotrophin signaling receptor TrkA and promote interaction with microtubules and thereby, axonal growth[177, 178]. Because MLR-associated components (Cav, Flot, gangliosides, cholesterol) are up-regulated and appear to be required for axonal growth and regeneration, interventions that enhance MLR formation in neurons may be a novel therapy following spinal cord or brain injury. In addition, as the cytoskeleton appears to be a key element of maintenance of MLR, cytoskeletal stabilization may offer a second therapeutic approach.
MLR and the Immunologic Synapse
Many factors initiate inflammatory responses to entities that include microorganisms, antigens and foreign bodies. Responses of the immune system are initiated by extracellular cues to membrane-localized signaling complexes. The organization of receptors, channels, and effector enzymes in MLR of immune cells is an important aspect of these responses. Within MLR, cholesterol, sphingolipids, and the underlying cytoskeleton play an essential role in communication of immune cells (Figure 5). What have been termed “immunologic synapses” are formed by the presentation of an antigen from the antigen presenting cell (APC) to the T cell. In this process, peripheral MLR cluster towards a central MLR region to become a supramolecular activation cluster (cSMAC); this clustering and T-cell antigen receptor (TCR) activation (i.e., its dimerization) is cholesterol-, sphingomyelin-, and Cav-1-dependent[179, 180]. MLR-cytoskeletal interactions are essential for T cell activation[181–183]. The lateral membrane mobility and clustering of MLR is thought to enhance signal transduction at immunologic synapses. This clustering depends upon Rho-mediated actin rearrangement; disruption of rafts prevents formation of immunologic synapses[10]. Within MLR, Cav-1 contributes to membrane polarity and TCR-induced actin polymerization of CD8[180]. In addition, CD26 binds Cav-1 in APCs to induce T cell proliferation in a TCR/CD3-dependent manner[184].
Figure 5. Membrane/lipid rafts and the immunologic synapse.
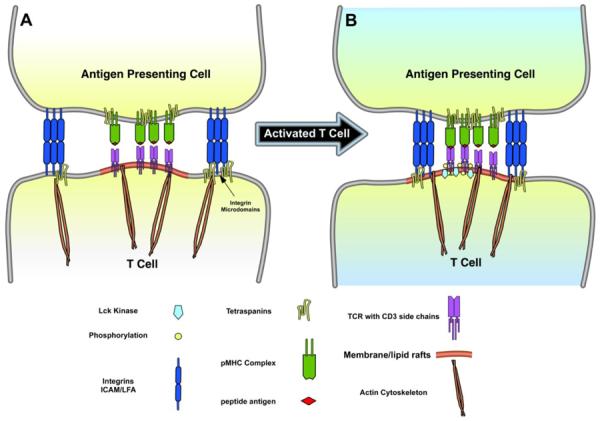
Panel A, or pre-activation state, depicts an antigen presenting cell (APC) that is presenting peptide antigens extracellularly via the major histocompatibility complex (pMHC) to the T cell receptor (TCR) located within MLR located in the central supramolecular activation cluster (cSMAC). APC and T cells connect via cell adhesion molecules (integrins) located within tetraspanin-enriched microdomains (TEMs) located in the peripheral supramolecular activation cluster (pSMAC). Panel B, or activation state, shows that after TCR-pMHC interactions, the pSMAC MLR migrates laterally towards the cSMAC via actin cytoskeletal rearrangement. cSMAC regulates termination of the signaling through subsequent TCR downregulation.
In addition to Cav, the MLR-associated proteins Flots and TSPNs play a role in T cell activation. Flot in polarized regions of T cells form a `cap' where TCR complexes organize following stimulation[154]. This `cap' is necessary for T cell activation in that it localizes the TCR and initiates communication with actin through Fyn, Lck, Src, and Rho GTPases. Similar to subcellular actions exhibited in neurons, Flots interact with the GPI-anchored protein PrP to form PrP clusters and subsequent recruitment of CD3. TSPNs faciliate similar clustering events and regulate the lateral movement of SMAC, leading to the formation of cSMAC in an actin-dependent manner following TCR activation and CD3 chain phosphorylation by Lck[185, 186].
Certain other functions of blood cells are dependent upon MLR/cytoskeletal interaction. For example, MLR are involved in erythoblast enucleation in a Rac-microtubule dependent manner[187]. Platelet interaction with damaged endothelial regions is dependent in part on the anchoring of glyoprotein GPlb-IX-V complex to MLR via interactions with filamin and the actin cytoskeleton[188]. Moreover, platelet activation is dependent upon PrP localization to MLR and interactions with the platelet cytoskeleton[189].
MLR and Disease: Entry of pathogens and toxins, migration of malignant cells and Alzheimer's disease
Pathogen Invasion
Endocytosis plays an important role in the communication between eukaryotic cells and their outside environment by regulating cell dynamics and homeostasis. Cytoskeletal rearrangement allows (and is required for) cellular entry via MLR of many types of cargo, including nutrients, receptor-ligand complexes, lipid, antigens, DNA nanoparticles, and infectious agents that include fungal, bacterial, and viral pathogens[190–195]. Of note, especially with respect to disease, are the roles of MLR and interaction with cytoskeletal components in the endocytosis of microbial-derived toxins and pathogens. One example is the neurotropic fungal pathogen Cryptococcus neoformans, which crosses the blood brain barrier of brain microvascular endothelial cells via a MLR-mediated endocytotic pathway[192]. This occurs in GM1-enriched regions and is regulated by dual specificity tyrosine-phosphorylation-regulated kinase 3 (Dyrk3)[192]. Pore-forming toxins such as equinatoxin II invade cells by binding to MLR, and subsquently re-organize the actin cytoskeleton to facilitate toxin entry that leads to cell death[94]. As part of their invasion of cells, bacterial toxins such as the exotoxin of Vibrio cholerae, CTX and Shigella dysenteriae Shiga (STX) bind to the MLR glycosphingolipids GM1 and globotriosylceramide Gb3, respectively[194]. Such toxins (which also include Shiga-like toxin, Escherichia coli Heat labile enterotoxin 1 and Heat labile toxin IIb) are composed of a catalytic A subunit and a pentameric B subunit; the latter flat-like ring structure binds to the carbohydrate moiety of glycosphingolipids to achieve internalization. Clostridial neurotoxins, such as Tetanus and Botulinium, possess a single binding subunit and invade cells by binding to gangliosides GT1b, GD1b, GQ1b and GD1a, GT1b[196, 197]. Several types of viruses utilize glycosphingolipids (MLR) for eukaryotic cell invasion; examples include Simian virus 40, murine, Merkel cell and BK polyomavirus, and murine norovirus[194].
Cancer Cell Metastasis
Migration of metastic cells and their invasion of tissues depends upon many cellular events, including detachment from adhesion molecules[198]. MLR-resident proteins at focal adhesion sites play a central role in initiating detachment from the ECM[158]. For example, prostate cancer cells appear to require Cav-1, cavin-1, Src, FAK, and the actin cytoskeleton to achieve adherence to or detachment from extracellular components[199–201]. In some cases MLR proteins (e.g.,Cav-1) are released by these cells, an event that alters recruitment of the actin cytoskeleton to the PM[202]. MLR also contribute to the regulation of polarized cell adhesion through growth factor receptor activation in a ligand-independent manner[203], via CD24-integrin-mediated transmission of contractile forces[204], or as a platform for association of the KCl cotransporter with a myosin-actin motor protein[205]. Neuroblastoma/glioma cell migration may occur, at least in part, through the action of MLR-localized P2Y2 receptor-Gq/11-PLC signaling[206].
Neurodegenerative Diseases
MLR play contrasting roles (i.e., preventive or exacerbating) in certain neurodegenerative diseases, such as Alzheimer's disease (AD). Altered MLR homeostasis or “lipid raft aging” has been suggested to be a factor in the processing of amyloid precursor protein (APP) to the toxic amyloid beta (Aβ) peptide[207–210]. Enhanced phospholipid/cholesterol ratios are found in brain membranes from AD mice compared to those of wild-type animals, suggesting that “lipid raft aging” results from alterations in the structure and physicochemical properties of raft microdomains[208, 211, 212]. Within MLR, both Cav-1 and Flot have been implicated in AD, in particular with regards to γ-secretase-mediated APP processing[210] and estrogen receptor-voltage-dependent anion channel (VDAC) neuroprotection[209]. However, it is unclear if these raft proteins enhance or dampen the production of the neurotoxic peptides[210, 213]. Thathiah et al have demonstrated that a GPCR-β-arrestin 2 interaction in MLR enhances γ–secretase activity and subsequent generation of Aβ peptide and that GPCRs enhance Aβ production through their association with β-arrestin 2 in MLR[214]. Aβ production can alter membrane mechanical properties, such as membrane fluidity and molecular order[215], suggesting that alterations in physiochemical properties of the PM may be the result of Aβ aggregates rather than the cause of such alterations.
Alterations in MLR integrity due to oxidative stress[216], changes in lipid bilayer components that lead to altered MLR physiochemcial properties[217] or depletion in membrane cholesterol content[218, 219], not only alter cellular polarity[220], signal transduction[221], and morphology, but also can contribute to aberrant intercellular communication leading to organ pathology, including in the nervous system [166, 222, 223].
A general conclusion of work to date (in the nervous system and others) is that expression of MLR in animal cell membranes depends upon sterols, sphingolipids, and an active, energy-dependent actin/myosin meshwork. This dependency on cellular energy implies a role for mitochondria in close promixity to the MLR/cytoskeletal interaction. Studies are needed to investigate mitochondrial transport[224, 225] and how mitochondria that are closely apposed to rafts in the PM interact with the actin/myosin-containing components[226]. Data from such efforts should further our understanding of normal cell physiology and how disease, injury or oxidative stress affects MLR clustering, signaling, and cell function or fate both within and outside the central nervous system.
Conclusion
MLR are involved in numerous cellular functions, which include signaling, trafficking, adhesion, migration, and growth—all of which may involve an interaction between rafts and the actin/myosin and tubulin cytoskeleton. Certain components that regulate actin polymerization and can stabilize microtubules are associated with membrane rafts (e.g., Src family kinases and small Rho GTPases). Considerable data show that microtubules and the actin cytoskeleton are intimately associated with MLR. This association contributes to cytoskeletal organization and recruitment of `cargo' to the PM, which in turn facilitate cell polarity, adherence and migration, stretch and contraction, and the transduction of extracellular signals to the intracellular milieu.
MLR facilitate the assembly of cell-surface proteins and signaling `hubs' at the PM. These signaling platforms communicate with the cytoskeleton (via small GTPases) so as to facilitate the transport and recruitment of bulk membrane material (i.e., lipids and proteins) from internal vesicular stores to MLR and likely to other portions of the PM. “Clustering” of proteins helps establish polarized platforms necessary for ECM adhesion, cellular migration, membrane growth, expansion or guidance (filopodia, neuritic processes), and in the case of smooth and striated muscle, mechanotransduction (stretch and contraction/relaxation). Within MLR, receptors, channels, and small GTPases can communicate with high fidelity, interactions that occur, at least in part, as a consequence of their close proximity to one another.
Extracellular stimuli that trigger membrane signal transduction, protein recruitment, and alterations to the cytoskeletal architecture, depend upon MLR integrity and function. Signaling and activation of small GTPases to regulate actin dynamics is regulated by the presence of protein scaffolds such as Cavs, reggies/flotillin, GPI-anchored proteins, and tetraspanins. These protein scaffolds not only contribute to the formation of membrane microdomains but also to the lateral clustering of microdomains within the PM (e.g., tetraspinin-enriched integrin domains clustering with TCR-enriched liquid ordered nanodomains) and thereby, help facilitate high fidelity extracellular-intracellular signaling, especially via cytoskeletal components. The clustering of lipids and proteins forms this signaling platform.
Although the precise properties and functions of MLR in the resting state of the PM remain incompletely understood (and controversial), a role for liquid-ordered domains in signal transduction, and actin-containing cytoskeletal components in the formation of larger functional MLR via clustering of nanoscale membrane domains is more widely accepted[30, 227]. Signaling enhances MLR clustering into a centric or focal region of the cellular membrane, thus leading to the question as to whether MLR enhance signaling or if initial signaling events enhance MLR formation and clustering. Irrespective of the answer to this “chicken-egg” question, the interplay between MLR-associated proteins and the cytoskeleton impacts on cellular signal transduction, polarity, and morphological changes. Advances in understanding the biology and chemistry of MLR (for example, the precise identity and functional role of the full complement of lipids and proteins in MLR from different cells) and how they are intimately connected with the ECM and underlying cytoskeleton should provide new insights regarding cellular communication and regulation. Other questions remain as well. For example, do different subcellular MLR species regulate membrane functional properties via differences in assembly and interaction of components within the same cell (e.g., apical versus basal membrane formation in epithelial and endothelial cells or axonal versus dendritic neuritic conversion in neurons)? Is there a common pattern within different cell types with respect to the cytoskeletal components that interact with MLR? Are MLR therapeutic targets? We believe that findings related to MLR may have therapeutic implications, for example by identifying components to which therapies could be developed (for example, cell-type-specific targeted overexpression of proteins, microRNAs (miRs), antagomirs) or ones that are modified to restore, maintain or repair membrane raft integrity and function in diseased or injured cells and tissues.
Highlights
APC presents antigens via pMHC to TCR localized in MLR in the central SMAC.
APC and T cells connect via CAMs (integrins) located within MLR in peripheral SMAC.
Receptor activation causes cytoskeletal rearrangement and MLR lateral migration.
MLR migrates laterally towards the central SMAC is actin dependent.
Central SMAC terminates signaling through TCR downregulation.
ACKNOWLEDGMENTS
Work in the authors' laboratories is supported by Veteran Affairs Merit Award from the Department of Veterans Affairs BX001225 (B. P. Head) and BX001963 (H. H. Patel) and the National Institutes of Health, Bethesda, MD, U.S.A., R01 NS073653 (B. P. Head), RO1 HL091071 (H. H. Patel) and HL107200 (H. H. Patel).
Abbreviations
- AC
adenylyl cyclases
- AD
Alzheimer's disease
- AJ
adherent junctions
- AMPAR
alpha-amino-3-hydroxyl-5-methyl-4-isoxazole-propionate receptor
- APC
antigen presenting cell
- APP
amyloid precursor protein
- Aβ
amyloid beta peptide
- CAM
cellular adhesion molecules
- cAMP
cyclic adenosine 3',5' monophosphate
- Cav
caveolin
- CBD
caveolin binding domain
- CRAC
cholesterol recognition/interaction amino acid consensus
- CSD
caveolin scaffolding domain
- cSMAC
central supramolecular activation cluster
- CTX
cholera toxin
- EC
endothelial cell
- ECM
extracellular matrix
- eNOS
endothelial nitric oxide synthase (NOS3)
- FAK
focal adhesion kinase
- Flot
flotillin
- GD
ganglioside disialic acid
- GJ
gap junctions
- GM
ganglioside monosialic acid
- GPCR
G-protein-coupled receptor
- GPI
glycosylphosphatidylinositol
- ICAM/VCAM
inter/vascular CAM
- JAM
junctional adhesion molecules
- mGluR
metabotropic glutamate receptor
- MLR
membrane/lipid rafts
- MT
microtubules
- NMDAR
N-methyl-D-aspartate receptor
- pMHC
peripheral major histocompatibility complex
- PrP
prion protein
- PTRF
polymerase I and transcript release factor
- RTK
receptor tyrosine kinases
- TCR
T cell receptor
- TEM
tetraspanin-enriched microdomains
- TJ
tight junctions
- Trk
tropomyosin receptor kinase
- TRPC1
transient receptor potential cation channel
- TSPN
tetraspanin
- VGCC
voltage-gated Ca2+channels
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
REFERENCES
- [1].Gorter E, Grendel F. On Bimolecular Layers of Lipoids on the Chromocytes of the Blood. The Journal of experimental medicine. 1925;41:439–443. doi: 10.1084/jem.41.4.439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [2].Simons K, Sampaio JL. Membrane organization and lipid rafts. Cold Spring Harb Perspect Biol. 2011;3:1–17. doi: 10.1101/cshperspect.a004697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Mouritsen OG, Zuckermann MJ. What's so special about cholesterol? Lipids. 2004;39:1101–1113. doi: 10.1007/s11745-004-1336-x. [DOI] [PubMed] [Google Scholar]
- [4].Roduit C, van der Goot FG, De Los Rios P, Yersin A, Steiner P, Dietler G, Catsicas S, Lafont F, Kasas S. Elastic membrane heterogeneity of living cells revealed by stiff nanoscale membrane domains. Biophysical journal. 2008;94:1521–1532. doi: 10.1529/biophysj.107.112862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Lundbaek JA, Andersen OS, Werge T, Nielsen C. Cholesterol-induced protein sorting: an analysis of energetic feasibility. Biophysical journal. 2003;84:2080–2089. doi: 10.1016/S0006-3495(03)75015-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Singer SJ, Nicolson GL. The fluid mosaic model of the structure of cell membranes. Science. 1972;175:720–731. doi: 10.1126/science.175.4023.720. [DOI] [PubMed] [Google Scholar]
- [7].Simons K, van Meer G. Lipid sorting in epithelial cells. Biochemistry. 1988;27:6197–6202. doi: 10.1021/bi00417a001. [DOI] [PubMed] [Google Scholar]
- [8].Simons K, Ikonen E. Functional rafts in cell membranes. Nature. 1997;387:569–572. doi: 10.1038/42408. [DOI] [PubMed] [Google Scholar]
- [9].Pike LJ. Rafts defined: a report on the Keystone Symposium on Lipid Rafts and Cell Function. Journal of lipid research. 2006;47:1597–1598. doi: 10.1194/jlr.E600002-JLR200. [DOI] [PubMed] [Google Scholar]
- [10].Gonnord P, Blouin CM, Lamaze C. Membrane trafficking and signaling: two sides of the same coin. Seminars in cell & developmental biology. 2012;23:154–164. doi: 10.1016/j.semcdb.2011.11.002. [DOI] [PubMed] [Google Scholar]
- [11].Edidin M. Lipids on the frontier: a century of cell-membrane bilayers. Nature reviews. Molecular cell biology. 2003;4:414–418. doi: 10.1038/nrm1102. [DOI] [PubMed] [Google Scholar]
- [12].Head BP, Insel PA. Do caveolins regulate cells by actions outside of caveolae? Trends Cell Biol. 2007;17:51–57. doi: 10.1016/j.tcb.2006.11.008. [DOI] [PubMed] [Google Scholar]
- [13].Head BP, Patel HH, Roth DM, Murray F, Swaney JS, Niesman IR, Farquhar MG, Insel PA. Microtubules and actin microfilaments regulate lipid raft/caveolae localization of adenylyl cyclase signaling components. The Journal of biological chemistry. 2006;281:26391–26399. doi: 10.1074/jbc.M602577200. [DOI] [PubMed] [Google Scholar]
- [14].Suzuki T, Zhang J, Miyazawa S, Liu Q, Farzan MR, Yao WD. Association of membrane rafts and postsynaptic density: proteomics, biochemical, and ultrastructural analyses. Journal of neurochemistry. 2011;119:64–77. doi: 10.1111/j.1471-4159.2011.07404.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Viola A, Gupta N. Tether and trap: regulation of membrane-raft dynamics by actin-binding proteins. Nature reviews. Immunology. 2007;7:889–896. doi: 10.1038/nri2193. [DOI] [PubMed] [Google Scholar]
- [16].Simons K, Gerl MJ. Revitalizing membrane rafts: new tools and insights. Nature reviews. Molecular cell biology. 2010;11:688–699. doi: 10.1038/nrm2977. [DOI] [PubMed] [Google Scholar]
- [17].Whitehead SN, Gangaraju S, Aylsworth A, Hou ST. Membrane raft disruption results in neuritic retraction prior to neuronal death in cortical neurons. Bioscience trends. 2012;6:183–191. doi: 10.5582/bst.2012.v6.4.183. [DOI] [PubMed] [Google Scholar]
- [18].Goudenege S, Dargelos E, Claverol S, Bonneu M, Cottin P, Poussard S. Comparative proteomic analysis of myotube caveolae after milli-calpain deregulation. Proteomics. 2007;7:3289–3298. doi: 10.1002/pmic.200700124. [DOI] [PubMed] [Google Scholar]
- [19].Goswami D, Gowrishankar K, Bilgrami S, Ghosh S, Raghupathy R, Chadda R, Vishwakarma R, Rao M, Mayor S. Nanoclusters of GPI-anchored proteins are formed by cortical actin-driven activity. Cell. 2008;135:1085–1097. doi: 10.1016/j.cell.2008.11.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [20].Ritchie K, Iino R, Fujiwara T, Murase K, Kusumi A. The fence and picket structure of the plasma membrane of live cells as revealed by single molecule techniques (Review) Molecular membrane biology. 2003;20:13–18. doi: 10.1080/0968768021000055698. [DOI] [PubMed] [Google Scholar]
- [21].Kusumi A, Koyama-Honda I, Suzuki K. Molecular dynamics and interactions for creation of stimulation-induced stabilized rafts from small unstable steady-state rafts. Traffic. 2004;5:213–230. doi: 10.1111/j.1600-0854.2004.0178.x. [DOI] [PubMed] [Google Scholar]
- [22].Suzuki KG, Kasai RS, Hirosawa KM, Nemoto YL, Ishibashi M, Miwa Y, Fujiwara TK, Kusumi A. Transient GPI-anchored protein homodimers are units for raft organization and function. Nature chemical biology. 2012;8:774–783. doi: 10.1038/nchembio.1028. [DOI] [PubMed] [Google Scholar]
- [23].Suzuki KG. Lipid rafts generate digital-like signal transduction in cell plasma membranes. Biotechnology journal. 2012;7:753–761. doi: 10.1002/biot.201100360. [DOI] [PubMed] [Google Scholar]
- [24].Gowrishankar K, Ghosh S, Saha S, Mayor RC,S, Rao M. Active remodeling of cortical actin regulates spatiotemporal organization of cell surface molecules. Cell. 2012;149:1353–1367. doi: 10.1016/j.cell.2012.05.008. [DOI] [PubMed] [Google Scholar]
- [25].Yamada E. The fine structure of the gall bladder epithelium of the mouse. The Journal of biophysical and biochemical cytology. 1955;1:445–458. doi: 10.1083/jcb.1.5.445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26].Palade G. Fine structure of blood capilaries. J. Appl. Phys. 1953;24:1419. [Google Scholar]
- [27].Karnovsky MJ, Kleinfeld AM, Hoover RL, Klausner RD. The concept of lipid domains in membranes. The Journal of cell biology. 1982;94:1–6. doi: 10.1083/jcb.94.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Fra AM, Williamson E, Simons K, Parton RG. Detergent-insoluble glycolipid microdomains in lymphocytes in the absence of caveolae. The Journal of biological chemistry. 1994;269:30745–30748. [PubMed] [Google Scholar]
- [29].Field KA, Holowka D, Baird B. Fc epsilon RI-mediated recruitment of p53/56lyn to detergent-resistant membrane domains accompanies cellular signaling. Proceedings of the National Academy of Sciences of the United States of America. 1995;92:9201–9205. doi: 10.1073/pnas.92.20.9201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].Jacobson K, Mouritsen OG, Anderson RG. Lipid rafts: at a crossroad between cell biology and physics. Nature cell biology. 2007;9:7–14. doi: 10.1038/ncb0107-7. [DOI] [PubMed] [Google Scholar]
- [31].Brown DA, Rose JK. Sorting of GPI-anchored proteins to glycolipid-enriched membrane subdomains during transport to the apical cell surface. Cell. 1992;68:533–544. doi: 10.1016/0092-8674(92)90189-j. [DOI] [PubMed] [Google Scholar]
- [32].Schuck S, Honsho M, Ekroos K, Shevchenko A, Simons K. Resistance of cell membranes to different detergents. Proceedings of the National Academy of Sciences of the United States of America. 2003;100:5795–5800. doi: 10.1073/pnas.0631579100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].Minogue S, Waugh MG. Lipid rafts, microdomain heterogeneity and inter-organelle contacts: impacts on membrane preparation for proteomic studies. Biology of the cell / under the auspices of the European Cell Biology Organization. 2012;104:618–627. doi: 10.1111/boc.201200020. [DOI] [PubMed] [Google Scholar]
- [34].Song KS, Li S, Okamoto T, Quilliam LA, Sargiacomo M, Lisanti MP. Co-purification and direct interaction of Ras with caveolin, an integral membrane protein of caveolae microdomains. Detergent-free purification of caveolae microdomains. The Journal of biological chemistry. 1996;271:9690–9697. doi: 10.1074/jbc.271.16.9690. [DOI] [PubMed] [Google Scholar]
- [35].Smart EJ, Ying YS, Mineo C, Anderson RG. A detergent-free method for purifying caveolae membrane from tissue culture cells. Proceedings of the National Academy of Sciences of the United States of America. 1995;92:10104–10108. doi: 10.1073/pnas.92.22.10104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [36].Macdonald JL, Pike LJ. A simplified method for the preparation of detergent-free lipid rafts. Journal of lipid research. 2005;46:1061–1067. doi: 10.1194/jlr.D400041-JLR200. [DOI] [PubMed] [Google Scholar]
- [37].Schnitzer JE, McIntosh DP, Dvorak AM, Liu J, Oh P. Separation of caveolae from associated microdomains of GPI-anchored proteins. Science. 1995;269:1435–1439. doi: 10.1126/science.7660128. [DOI] [PubMed] [Google Scholar]
- [38].Stan RV, Roberts WG, Predescu D, Ihida K, Saucan L, Ghitescu L, Palade GE. Immunoisolation and partial characterization of endothelial plasmalemmal vesicles (caveolae) Mol Biol Cell. 1997;8:595–605. doi: 10.1091/mbc.8.4.595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].Pike LJ. The challenge of lipid rafts. Journal of lipid research. 2009;50(Suppl):S323–328. doi: 10.1194/jlr.R800040-JLR200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [40].Lebiedzinska M, Szabadkai G, Jones AW, Duszynski J, Wieckowski MR. Interactions between the endoplasmic reticulum, mitochondria, plasma membrane and other subcellular organelles. The international journal of biochemistry & cell biology. 2009;41:1805–1816. doi: 10.1016/j.biocel.2009.02.017. [DOI] [PubMed] [Google Scholar]
- [41].Waugh MG, Hsuan JJ. Preparation of membrane rafts. Methods in molecular biology. 2009;462:403–414. doi: 10.1007/978-1-60327-115-8_26. [DOI] [PubMed] [Google Scholar]
- [42].Head BP, Patel HH, Roth DM, Lai NC, Niesman IR, Farquhar MG, Insel PA. G-protein-coupled receptor signaling components localize in both sarcolemmal and intracellular caveolin-3-associated microdomains in adult cardiac myocytes. The Journal of biological chemistry. 2005;280:31036–31044. doi: 10.1074/jbc.M502540200. [DOI] [PubMed] [Google Scholar]
- [43].Owen DM, Magenau A, Williamson D, Gaus K. The lipid raft hypothesis revisited--new insights on raft composition and function from super-resolution fluorescence microscopy. BioEssays : news and reviews in molecular, cellular and developmental biology. 2012;34:739–747. doi: 10.1002/bies.201200044. [DOI] [PubMed] [Google Scholar]
- [44].Leslie M. Mysteries of the cell. How does a cell know its size? Science. 2011;334:1047–1048. doi: 10.1126/science.334.6059.1047. [DOI] [PubMed] [Google Scholar]
- [45].Levental I, Byfield FJ, Chowdhury P, Gai F, Baumgart T, Janmey PA. Cholesterol-dependent phase separation in cell-derived giant plasma-membrane vesicles. The Biochemical journal. 2009;424:163–167. doi: 10.1042/BJ20091283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [46].Meder D, Moreno MJ, Verkade P, Vaz WL, Simons K. Phase coexistence and connectivity in the apical membrane of polarized epithelial cells. Proceedings of the National Academy of Sciences of the United States of America. 2006;103:329–334. doi: 10.1073/pnas.0509885103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [47].Lillemeier BF, Pfeiffer JR, Surviladze Z, Wilson BS, Davis MM. Plasma membrane-associated proteins are clustered into islands attached to the cytoskeleton. Proceedings of the National Academy of Sciences of the United States of America. 2006;103:18992–18997. doi: 10.1073/pnas.0609009103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [48].Zheng W, Kollmeyer J, Symolon H, Momin A, Munter E, Wang E, Kelly S, Allegood JC, Liu Y, Peng Q, Ramaraju H, Sullards MC, Cabot M, Merrill AH., Jr. Ceramides and other bioactive sphingolipid backbones in health and disease: lipidomic analysis, metabolism and roles in membrane structure, dynamics, signaling and autophagy. Biochimica et biophysica acta. 2006;1758:1864–1884. doi: 10.1016/j.bbamem.2006.08.009. [DOI] [PubMed] [Google Scholar]
- [49].Morales-Serna JA, Boutureira O, Diaz Y, Matheu MI, Castillon S. Recent advances in the glycosylation of sphingosines and ceramides. Carbohydrate research. 2007;342:1595–1612. doi: 10.1016/j.carres.2007.03.028. [DOI] [PubMed] [Google Scholar]
- [50].Young MM, Kester M, Wang HG. Sphingolipids: regulators of crosstalk between apoptosis and autophagy. Journal of lipid research. 2013;54:5–19. doi: 10.1194/jlr.R031278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [51].Sonnino S, Mauri L, Chigorno V, Prinetti A. Gangliosides as components of lipid membrane domains. Glycobiology. 2007;17:1R–13R. doi: 10.1093/glycob/cwl052. [DOI] [PubMed] [Google Scholar]
- [52].Merritt EA, Sixma TK, Kalk KH, van Zanten BA, Hol WG. Galactose-binding site in Escherichia coli heat-labile enterotoxin (LT) and cholera toxin (CT) Molecular microbiology. 1994;13:745–753. doi: 10.1111/j.1365-2958.1994.tb00467.x. [DOI] [PubMed] [Google Scholar]
- [53].Blank N, Schiller M, Krienke S, Wabnitz G, Ho AD, Lorenz HM. Cholera toxin binds to lipid rafts but has a limited specificity for ganglioside GM1. Immunology and cell biology. 2007;85:378–382. doi: 10.1038/sj.icb.7100045. [DOI] [PubMed] [Google Scholar]
- [54].Simons K, Vaz WL. Model systems, lipid rafts, and cell membranes. Annual review of biophysics and biomolecular structure. 2004;33:269–295. doi: 10.1146/annurev.biophys.32.110601.141803. [DOI] [PubMed] [Google Scholar]
- [55].Glenney JR, Jr., Soppet D. Sequence and expression of caveolin, a protein component of caveolae plasma membrane domains phosphorylated on tyrosine in Rous sarcoma virus-transformed fibroblasts. Proceedings of the National Academy of Sciences of the United States of America. 1992;89:10517–10521. doi: 10.1073/pnas.89.21.10517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [56].Rothberg KG, Heuser JE, Donzell WC, Ying YS, Glenney JR, Anderson RG. Caveolin, a protein component of caveolae membrane coats. Cell. 1992;68:673–682. doi: 10.1016/0092-8674(92)90143-z. [DOI] [PubMed] [Google Scholar]
- [57].Kurzchalia TV, Dupree P, Parton RG, Kellner R, Virta H, Lehnert M, Simons K. VIP21, a 21-kD membrane protein is an integral component of trans-Golgi-network-derived transport vesicles. The Journal of cell biology. 1992;118:1003–1014. doi: 10.1083/jcb.118.5.1003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [58].Dupree P, Parton RG, Raposo G, Kurzchalia TV, Simons K. Caveolae and sorting in the trans-Golgi network of epithelial cells. The EMBO journal. 1993;12:1597–1605. doi: 10.1002/j.1460-2075.1993.tb05804.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [59].Murata M, Peranen J, Schreiner R, Wieland F, Kurzchalia TV, Simons K. VIP21/caveolin is a cholesterol-binding protein. Proceedings of the National Academy of Sciences of the United States of America. 1995;92:10339–10343. doi: 10.1073/pnas.92.22.10339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [60].Rietveld A, Simons K. The differential miscibility of lipids as the basis for the formation of functional membrane rafts. Biochimica et biophysica acta. 1998;1376:467–479. doi: 10.1016/s0304-4157(98)00019-7. [DOI] [PubMed] [Google Scholar]
- [61].Zocher M, Zhang C, Rasmussen SG, Kobilka BK, Muller DJ. Cholesterol increases kinetic, energetic, and mechanical stability of the human beta2-adrenergic receptor. Proceedings of the National Academy of Sciences of the United States of America. 2012;109:E3463–3472. doi: 10.1073/pnas.1210373109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [62].Glenney JR., Jr. Tyrosine phosphorylation of a 22-kDa protein is correlated with transformation by Rous sarcoma virus. The Journal of biological chemistry. 1989;264:20163–20166. [PubMed] [Google Scholar]
- [63].Salanueva IJ, Cerezo A, Guadamillas MC, del Pozo MA. Integrin regulation of caveolin function. Journal of cellular and molecular medicine. 2007;11:969–980. doi: 10.1111/j.1582-4934.2007.00109.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [64].Schlegel A, Arvan P, Lisanti MP. Caveolin-1 binding to endoplasmic reticulum membranes and entry into the regulated secretory pathway are regulated by serine phosphorylation. Protein sorting at the level of the endoplasmic reticulum. The Journal of biological chemistry. 2001;276:4398–4408. doi: 10.1074/jbc.M005448200. [DOI] [PubMed] [Google Scholar]
- [65].Fielding PE, Chau P, Liu D, Spencer TA, Fielding CJ. Mechanism of platelet-derived growth factor-dependent caveolin-1 phosphorylation: relationship to sterol binding and the role of serine-80. Biochemistry. 2004;43:2578–2586. doi: 10.1021/bi035442c. [DOI] [PubMed] [Google Scholar]
- [66].Mundy DI, Machleidt T, Ying YS, Anderson RG, Bloom GS. Dual control of caveolar membrane traffic by microtubules and the actin cytoskeleton. J Cell Sci. 2002;115:4327–4339. doi: 10.1242/jcs.00117. [DOI] [PubMed] [Google Scholar]
- [67].Epand RM, Sayer BG, Epand RF. Caveolin scaffolding region and cholesterol-rich domains in membranes. Journal of molecular biology. 2005;345:339–350. doi: 10.1016/j.jmb.2004.10.064. [DOI] [PubMed] [Google Scholar]
- [68].Hoop CL, Sivanandam VN, Kodali R, Srnec MN, van der Wel PC. Structural characterization of the caveolin scaffolding domain in association with cholesterol-rich membranes. Biochemistry. 2012;51:90–99. doi: 10.1021/bi201356v. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [69].Couet J, Li S, Okamoto T, Ikezu T, Lisanti MP. Identification of peptide and protein ligands for the caveolin-scaffolding domain. Implications for the interaction of caveolin with caveolae-associated proteins. The Journal of biological chemistry. 1997;272:6525–6533. doi: 10.1074/jbc.272.10.6525. [DOI] [PubMed] [Google Scholar]
- [70].Insel PA, Head BP, Ostrom RS, Patel HH, Swaney JS, Tang CM, Roth DM. Caveolae and lipid rafts: G protein-coupled receptor signaling microdomains in cardiac myocytes. Annals of the New York Academy of Sciences. 2005;1047:166–172. doi: 10.1196/annals.1341.015. [DOI] [PubMed] [Google Scholar]
- [71].Fuhs SR, Insel PA. Caveolin-3 undergoes SUMOylation by the SUMO E3 ligase PIASy: sumoylation affects G-protein-coupled receptor desensitization. The Journal of biological chemistry. 2011;286:14830–14841. doi: 10.1074/jbc.M110.214270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [72].Byrne DP, Dart C, Rigden DJ. Evaluating caveolin interactions: do proteins interact with the caveolin scaffolding domain through a widespread aromatic residue-rich motif? PloS one. 2012;7:e44879. doi: 10.1371/journal.pone.0044879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [73].Pike LJ. Lipid rafts: bringing order to chaos. Journal of lipid research. 2003;44:655–667. doi: 10.1194/jlr.R200021-JLR200. [DOI] [PubMed] [Google Scholar]
- [74].Yamamoto M, Toya Y, Schwencke C, Lisanti MP, Myers MG, Jr., Ishikawa Y. Caveolin is an activator of insulin receptor signaling. The Journal of biological chemistry. 1998;273:26962–26968. doi: 10.1074/jbc.273.41.26962. [DOI] [PubMed] [Google Scholar]
- [75].Wary KK, Mariotti A, Zurzolo C, Giancotti FG. A requirement for caveolin-1 and associated kinase Fyn in integrin signaling and anchorage-dependent cell growth. Cell. 1998;94:625–634. doi: 10.1016/s0092-8674(00)81604-9. [DOI] [PubMed] [Google Scholar]
- [76].Head BP, Hu Y, Finley JC, Saldana MD, Bonds JA, Miyanohara A, Niesman IR, Ali SS, Murray F, Insel PA, Roth DM, Patel HH, Patel PM. Neuron-targeted caveolin-1 protein enhances signaling and promotes arborization of primary neurons. The Journal of biological chemistry. 2011;286:33310–33321. doi: 10.1074/jbc.M111.255976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [77].Pelkmans L, Fava E, Grabner H, Hannus M, Habermann B, Krausz E, Zerial M. Genome-wide analysis of human kinases in clathrin- and caveolae/raft-mediated endocytosis. Nature. 2005;436:78–86. doi: 10.1038/nature03571. [DOI] [PubMed] [Google Scholar]
- [78].Sargiacomo M, Scherer PE, Tang Z, Kübler E, Song KS, Sanders MC, Lisanti MP. Oligomeric structure of caveolin: implications for caveolae membrane organization. Proceedings of the National Academy of Sciences of the United States of America. 1995;92:9407–9411. doi: 10.1073/pnas.92.20.9407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [79].Monier S, Parton RG, Vogel F, Behlke J, Henske A, Kurzchalia TV. VIP21-caveolin, a membrane protein constituent of the caveolar coat, oligomerizes in vivo and in vitro. Mol Biol Cell. 1995;6:911–927. doi: 10.1091/mbc.6.7.911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [80].Fernandez I, Ying Y, Albanesi J, Anderson RG. Mechanism of caveolin filament assembly. Proceedings of the National Academy of Sciences of the United States of America. 2002;99:11193–11198. doi: 10.1073/pnas.172196599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [81].Whiteley G, Collins RF, Kitmitto A. Characterization of the molecular architecture of human caveolin-3 and interaction with the skeletal muscle ryanodine receptor. The Journal of biological chemistry. 2012;287:40302–40316. doi: 10.1074/jbc.M112.377085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [82].Stern CM, Mermelstein PG. Caveolin regulation of neuronal intracellular signaling. Cellular and molecular life sciences : CMLS. 2010;67:3785–3795. doi: 10.1007/s00018-010-0447-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [83].Liu L, Pilch PF. A critical role of cavin (polymerase I and transcript release factor) in caveolae formation and organization. The Journal of biological chemistry. 2008;283:4314–4322. doi: 10.1074/jbc.M707890200. [DOI] [PubMed] [Google Scholar]
- [84].Hansen CG, Nichols BJ. Exploring the caves: cavins, caveolins and caveolae. Trends Cell Biol. 2010;20:177–186. doi: 10.1016/j.tcb.2010.01.005. [DOI] [PubMed] [Google Scholar]
- [85].Aboulaich N, Vainonen JP, Stralfors P, Vener AV. Vectorial proteomics reveal targeting, phosphorylation and specific fragmentation of polymerase I and transcript release factor (PTRF) at the surface of caveolae in human adipocytes. The Biochemical journal. 2004;383:237–248. doi: 10.1042/BJ20040647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [86].Chadda R, Mayor S. PTRF triggers a cave in. Cell. 2008;132:23–24. doi: 10.1016/j.cell.2007.12.021. [DOI] [PubMed] [Google Scholar]
- [87].Schulte T, Paschke KA, Laessing U, Lottspeich F, Stuermer CA. Reggie-1 and reggie-2, two cell surface proteins expressed by retinal ganglion cells during axon regeneration. Development. 1997;124:577–587. doi: 10.1242/dev.124.2.577. [DOI] [PubMed] [Google Scholar]
- [88].Lang DM, Lommel S, Jung M, Ankerhold R, Petrausch B, Laessing U, Wiechers MF, Plattner H, Stuermer CA. Identification of reggie-1 and reggie-2 as plasmamembrane-associated proteins which cocluster with activated GPI-anchored cell adhesion molecules in non-caveolar micropatches in neurons. Journal of neurobiology. 1998;37:502–523. doi: 10.1002/(sici)1097-4695(199812)37:4<502::aid-neu2>3.0.co;2-s. [DOI] [PubMed] [Google Scholar]
- [89].Solis GP, Hoegg M, Munderloh C, Schrock Y, Malaga-Trillo E, Rivera-Milla E, Stuermer CA. Reggie/flotillin proteins are organized into stable tetramers in membrane microdomains. The Biochemical journal. 2007;403:313–322. doi: 10.1042/BJ20061686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [90].Palade GE. Blood capillaries of the heart and other organs. Circulation. 1961;24:368–388. doi: 10.1161/01.cir.24.2.368. [DOI] [PubMed] [Google Scholar]
- [91].Lucero HA, Robbins PW. Lipid rafts-protein association and the regulation of protein activity. Archives of biochemistry and biophysics. 2004;426:208–224. doi: 10.1016/j.abb.2004.03.020. [DOI] [PubMed] [Google Scholar]
- [92].Simons K, Toomre D. Lipid rafts and signal transduction. Nature reviews. Molecular cell biology. 2000;1:31–39. doi: 10.1038/35036052. [DOI] [PubMed] [Google Scholar]
- [93].Sebastiao AM, Colino-Oliveira M, Assaife-Lopes N, Dias RB, Ribeiro JA. Lipid rafts, synaptic transmission and plasticity: impact in age-related neurodegenerative diseases. Neuropharmacology. 2013;64:97–107. doi: 10.1016/j.neuropharm.2012.06.053. [DOI] [PubMed] [Google Scholar]
- [94].Garcia-Saez AJ, Buschhorn SB, Keller H, Anderluh G, Simons K, Schwille P. Oligomerization and pore formation by equinatoxin II inhibit endocytosis and lead to plasma membrane reorganization. The Journal of biological chemistry. 2011;286:37768–37777. doi: 10.1074/jbc.M111.281592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [95].Harder T, Engelhardt KR. Membrane domains in lymphocytes - from lipid rafts to protein scaffolds. Traffic. 2004;5:265–275. doi: 10.1111/j.1600-0854.2003.00163.x. [DOI] [PubMed] [Google Scholar]
- [96].Pelkmans L, Helenius A. Endocytosis via caveolae. Traffic. 2002;3:311–320. doi: 10.1034/j.1600-0854.2002.30501.x. [DOI] [PubMed] [Google Scholar]
- [97].Yang B, Radel C, Hughes D, Kelemen S, Rizzo V. p190 RhoGTPase-activating protein links the beta1 integrin/caveolin-1 mechanosignaling complex to RhoA and actin remodeling. Arteriosclerosis, thrombosis, and vascular biology. 2011;31:376–383. doi: 10.1161/ATVBAHA.110.217794. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [98].Sbalzarini IF. Active flows cluster cell surface proteins. Developmental cell. 2012;22:1121–1122. doi: 10.1016/j.devcel.2012.05.015. [DOI] [PubMed] [Google Scholar]
- [99].Parton RG, Simons K. The multiple faces of caveolae, Nature reviews. Molecular cell biology. 2007;8:185–194. doi: 10.1038/nrm2122. [DOI] [PubMed] [Google Scholar]
- [100].Sowa G. Caveolae, caveolins, cavins, and endothelial cell function: new insights. Frontiers in physiology. 2012;2:120. doi: 10.3389/fphys.2011.00120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [101].Ochoa CD, Stevens T. Studies on the cell biology of interendothelial cell gaps, American journal of physiology. Lung cellular and molecular physiology. 2012;302:L275–286. doi: 10.1152/ajplung.00215.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [102].Pokutta S, Drees F, Yamada S, Nelson WJ, Weis WI. Biochemical and structural analysis of alpha-catenin in cell-cell contacts. Biochemical Society transactions. 2008;36:141–147. doi: 10.1042/BST0360141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [103].Hartsock A, Nelson WJ. Adherens and tight junctions: structure, function and connections to the actin cytoskeleton. Biochimica et biophysica acta. 2008;1778:660–669. doi: 10.1016/j.bbamem.2007.07.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [104].Dodelet-Devillers A, Cayrol R, van Horssen J, Haqqani AS, de Vries HE, Engelhardt B, Greenwood J, Prat A. Functions of lipid raft membrane microdomains at the blood-brain barrier. Journal of molecular medicine. 2009;87:765–774. doi: 10.1007/s00109-009-0488-6. [DOI] [PubMed] [Google Scholar]
- [105].Creighton J, Jian M, Sayner S, Alexeyev M, Insel PA. Adenosine monophosphate-activated kinase alpha1 promotes endothelial barrier repair. FASEB journal : official publication of the Federation of American Societies for Experimental Biology. 2011;25:3356–3365. doi: 10.1096/fj.10-179218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [106].Komarova Y, Malik AB. Regulation of endothelial permeability via paracellular and transcellular transport pathways. Annual review of physiology. 2010;72:463–493. doi: 10.1146/annurev-physiol-021909-135833. [DOI] [PubMed] [Google Scholar]
- [107].Frank PG, Pavlides S, Cheung MW, Daumer K, Lisanti MP. Role of caveolin-1 in the regulation of lipoprotein metabolism. American journal of physiology. Cell physiology. 2008;295:C242–248. doi: 10.1152/ajpcell.00185.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [108].Ge S, Song L, Serwanski DR, Kuziel WA, Pachter JS. Transcellular transport of CCL2 across brain microvascular endothelial cells. Journal of neurochemistry. 2008;104:1219–1232. doi: 10.1111/j.1471-4159.2007.05056.x. [DOI] [PubMed] [Google Scholar]
- [109].Hu G, Vogel SM, Schwartz DE, Malik AB, Minshall RD. Intercellular adhesion molecule-1-dependent neutrophil adhesion to endothelial cells induces caveolae-mediated pulmonary vascular hyperpermeability. Circulation research. 2008;102:e120–131. doi: 10.1161/CIRCRESAHA.107.167486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [110].Chen Z, Bakhshi FR, Shajahan AN, Sharma T, Mao M, Trane A, Bernatchez P, van Nieuw Amerongen GP, Bonini MG, Skidgel RA, Malik AB, Minshall RD. Nitric oxide-dependent Src activation and resultant caveolin-1 phosphorylation promote eNOS/caveolin-1 binding and eNOS inhibition. Mol Biol Cell. 2012;23:1388–1398. doi: 10.1091/mbc.E11-09-0811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [111].Siddiqui MR, Komarova YA, Vogel SM, Gao X, Bonini MG, Rajasingh J, Zhao YY, Brovkovych V, Malik AB. Caveolin-1-eNOS signaling promotes p190RhoGAP-A nitration and endothelial permeability. The Journal of cell biology. 2011;193:841–850. doi: 10.1083/jcb.201012129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [112].Sundivakkam PC, Kwiatek AM, Sharma TT, Minshall RD, Malik AB, Tiruppathi C. Caveolin-1 scaffold domain interacts with TRPC1 and IP3R3 to regulate Ca2+ store release-induced Ca2+ entry in endothelial cells. American journal of physiology. Cell physiology. 2009;296:C403–413. doi: 10.1152/ajpcell.00470.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [113].Mirza MK, Yuan J, Gao XP, Garrean S, Brovkovych V, Malik AB, Tiruppathi C, Zhao YY. Caveolin-1 deficiency dampens Toll-like receptor 4 signaling through eNOS activation. The American journal of pathology. 2010;176:2344–2351. doi: 10.2353/ajpath.2010.091088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [114].Muriel O, Echarri A, Hellriegel C, Pavon DM, Beccari L, Del Pozo MA. Phosphorylated filamin A regulates actin-linked caveolae dynamics. J Cell Sci. 2011;124:2763–2776. doi: 10.1242/jcs.080804. [DOI] [PubMed] [Google Scholar]
- [115].Sverdlov M, Shinin V, Place AT, Castellon M, Minshall RD. Filamin A regulates caveolae internalization and trafficking in endothelial cells. Mol Biol Cell. 2009;20:4531–4540. doi: 10.1091/mbc.E08-10-0997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [116].Klein IK, Predescu DN, Sharma T, Knezevic I, Malik AB, Predescu S. Intersectin-2L regulates caveola endocytosis secondary to Cdc42-mediated actin polymerization. The Journal of biological chemistry. 2009;284:25953–25961. doi: 10.1074/jbc.M109.035071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [117].Berrier AL, Yamada KM. Cell-matrix adhesion. Journal of cellular physiology. 2007;213:565–573. doi: 10.1002/jcp.21237. [DOI] [PubMed] [Google Scholar]
- [118].Lock JG, Wehrle-Haller B, Stromblad S. Cell-matrix adhesion complexes: master control machinery of cell migration. Seminars in cancer biology. 2008;18:65–76. doi: 10.1016/j.semcancer.2007.10.001. [DOI] [PubMed] [Google Scholar]
- [119].Cao X, Surma MA, Simons K. Polarized sorting and trafficking in epithelial cells. Cell research. 2012;22:793–805. doi: 10.1038/cr.2012.64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [120].Corbeil D, Marzesco AM, Wilsch-Brauninger M, Huttner WB. The intriguing links between prominin-1 (CD133), cholesterol-based membrane microdomains, remodeling of apical plasma membrane protrusions, extracellular membrane particles, and (neuro)epithelial cell differentiation. FEBS letters. 2010;584:1659–1664. doi: 10.1016/j.febslet.2010.01.050. [DOI] [PubMed] [Google Scholar]
- [121].Marchiando AM, Shen L, Graham WV, Weber CR, Schwarz BT, Austin JR, 2nd, Raleigh DR, Guan Y, Watson AJ, Montrose MH, Turner JR. Caveolin-1-dependent occludin endocytosis is required for TNF-induced tight junction regulation in vivo. The Journal of cell biology. 2010;189:111–126. doi: 10.1083/jcb.200902153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [122].Itallie CM, Anderson JM. Caveolin binds independently to claudin-2 and occludin. Annals of the New York Academy of Sciences. 2012;1257:103–107. doi: 10.1111/j.1749-6632.2012.06535.x. [DOI] [PubMed] [Google Scholar]
- [123].Naota M, Shimada A, Morita T, Yamamoto Y, Inoue K, Takano H. Caveolae-mediated Endocytosis of Intratracheally Instilled Gold Colloid Nanoparticles at the Air-Blood Barrier in Mice. Toxicologic pathology. 2012 doi: 10.1177/0192623312457271. [DOI] [PubMed] [Google Scholar]
- [124].Lapierre LA, Ducharme NA, Drake KR, Goldenring JR, Kenworthy AK. Coordinated regulation of caveolin-1 and Rab11a in apical recycling compartments of polarized epithelial cells. Experimental cell research. 2012;318:103–113. doi: 10.1016/j.yexcr.2011.10.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [125].Scheiffele P, Verkade P, Fra AM, Virta H, Simons K, Ikonen E. Caveolin-1 and -2 in the exocytic pathway of MDCK cells. The Journal of cell biology. 1998;140:795–806. doi: 10.1083/jcb.140.4.795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [126].Lahtinen U, Honsho M, Parton RG, Simons K, Verkade P. Involvement of caveolin-2 in caveolar biogenesis in MDCK cells. FEBS letters. 2003;538:85–88. doi: 10.1016/s0014-5793(03)00135-2. [DOI] [PubMed] [Google Scholar]
- [127].Fiedler K, Parton RG, Kellner R, Etzold T, Simons K. VIP36, a novel component of glycolipid rafts and exocytic carrier vesicles in epithelial cells. The EMBO journal. 1994;13:1729–1740. doi: 10.1002/j.1460-2075.1994.tb06437.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [128].Mazzone A, Tietz P, Jefferson J, Pagano R, LaRusso NF. Isolation and characterization of lipid microdomains from apical and basolateral plasma membranes of rat hepatocytes. Hepatology. 2006;43:287–296. doi: 10.1002/hep.21039. [DOI] [PubMed] [Google Scholar]
- [129].Pena E, Arderiu G, Badimon L. Subcellular localization of tissue factor and human coronary artery smooth muscle cell migration. Journal of thrombosis and haemostasis : JTH. 2012 doi: 10.1111/j.1538-7836.2012.04910.x. [DOI] [PubMed] [Google Scholar]
- [130].Corriden R, Insel PA. New insights regarding the regulation of chemotaxis by nucleotides, adenosine, and their receptors. Purinergic signalling. 2012;8:587–598. doi: 10.1007/s11302-012-9311-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [131].Jagielska J, Kapopara PR, Salguero G, Scherr M, Schutt H, Grote K, Schieffer B, Bavendiek U. Interleukin-1 assembles a proangiogenic signaling module consisting of caveolin-1, tumor necrosis factor receptor-associated factor 6, p38-mitogen-activated protein kinase (MAPK), and MAPK-activated protein kinase 2 in endothelial cells. Arteriosclerosis, thrombosis, and vascular biology. 2012;32:1280–1288. doi: 10.1161/ATVBAHA.111.243477. [DOI] [PubMed] [Google Scholar]
- [132].Vincent C, Siddiqui TA, Schlichter LC. Podosomes in migrating microglia: components and matrix degradation. Journal of neuroinflammation. 2012;9:190. doi: 10.1186/1742-2094-9-190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [133].Verstraeten SV, Mackenzie GG, Oteiza PI. The plasma membrane plays a central role in cells response to mechanical stress. Biochimica et biophysica acta. 2010;1798:1739–1749. doi: 10.1016/j.bbamem.2010.06.010. [DOI] [PubMed] [Google Scholar]
- [134].Mitchell JS, Brown WS, Woodside DG, Vanderslice P, McIntyre BW. Clustering T-cell GM1 lipid rafts increases cellular resistance to shear on fibronectin through changes in integrin affinity and cytoskeletal dynamics. Immunology and cell biology. 2009;87:324–336. doi: 10.1038/icb.2008.103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [135].Radel C, Carlile-Klusacek M, Rizzo V. Participation of caveolae in beta1 integrin-mediated mechanotransduction. Biochemical and biophysical research communications. 2007;358:626–631. doi: 10.1016/j.bbrc.2007.04.179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [136].Best JM, Kamp TJ. Different subcellular populations of L-type Ca2+ channels exhibit unique regulation and functional roles in cardiomyocytes. Journal of molecular and cellular cardiology. 2012;52:376–387. doi: 10.1016/j.yjmcc.2011.08.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [137].Golini L, Chouabe C, Berthier C, Cusimano V, Fornaro M, Bonvallet R, Formoso L, Giacomello E, Jacquemond V, Sorrentino V. Junctophilin 1 and 2 proteins interact with the L-type Ca2+ channel dihydropyridine receptors (DHPRs) in skeletal muscle. The Journal of biological chemistry. 2011;286:43717–43725. doi: 10.1074/jbc.M111.292755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [138].Sabourin J, Cognard C, Constantin B. Regulation by scaffolding proteins of canonical transient receptor potential channels in striated muscle. Journal of muscle research and cell motility. 2009;30:289–297. doi: 10.1007/s10974-010-9206-9. [DOI] [PubMed] [Google Scholar]
- [139].Maguy A, Hebert TE, Nattel S. Involvement of lipid rafts and caveolae in cardiac ion channel function. Cardiovascular research. 2006;69:798–807. doi: 10.1016/j.cardiores.2005.11.013. [DOI] [PubMed] [Google Scholar]
- [140].Macdougall DA, Agarwal SR, Stopford EA, Chu H, Collins JA, Longster AL, Colyer J, Harvey RD, Calaghan S. Caveolae compartmentalise beta2-adrenoceptor signals by curtailing cAMP production and maintaining phosphatase activity in the sarcoplasmic reticulum of the adult ventricular myocyte. Journal of molecular and cellular cardiology. 2012;52:388–400. doi: 10.1016/j.yjmcc.2011.06.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [141].Asghari P, Scriven DR, Hoskins J, Fameli N, van Breemen C, Moore ED. The structure and functioning of the couplon in the mammalian cardiomyocyte. Protoplasma. 2012;249(Suppl 1):S31–38. doi: 10.1007/s00709-011-0347-5. [DOI] [PubMed] [Google Scholar]
- [142].Pfleger C, Ebeling G, Blasche R, Patton M, Patel HH, Kasper M, Barth K. Detection of caveolin-3/caveolin-1/P2X7R complexes in mice atrial cardiomyocytes in vivo and in vitro. Histochemistry and cell biology. 2012;138:231–241. doi: 10.1007/s00418-012-0961-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [143].Alday A, Urrutia J, Gallego M, Casis O. alpha1-adrenoceptors regulate only the caveolae-located subpopulation of cardiac K(V)4 channels. Channels. 2010;4:168–178. doi: 10.4161/chan.4.3.11479. [DOI] [PubMed] [Google Scholar]
- [144].Patel HH, Murray F, Insel PA. G-protein-coupled receptor-signaling components in membrane raft and caveolae microdomains. Handbook of experimental pharmacology. 2008:167–184. doi: 10.1007/978-3-540-72843-6_7. [DOI] [PubMed] [Google Scholar]
- [145].Insel PA, Patel HH. Membrane rafts and caveolae in cardiovascular signaling. Current opinion in nephrology and hypertension. 2009;18:50–56. doi: 10.1097/MNH.0b013e3283186f82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [146].Liu L, Li Y, Lin J, Liang Q, Sheng X, Wu J, Huang R, Liu S, Li Y. Connexin43 interacts with Caveolin-3 in the heart. Molecular biology reports. 2010;37:1685–1691. doi: 10.1007/s11033-009-9584-5. [DOI] [PubMed] [Google Scholar]
- [147].Vega-Moreno J, Tirado-Cortes A, Alvarez R, Irles C, Mas-Oliva J, Ortega A. Cholesterol depletion uncouples beta-dystroglycans from discrete sarcolemmal domains, reducing the mechanical activity of skeletal muscle. Cellular physiology and biochemistry : international journal of experimental cellular physiology, biochemistry, and pharmacology. 2012;29:905–918. doi: 10.1159/000186933. [DOI] [PubMed] [Google Scholar]
- [148].Formigli L, Sassoli C, Squecco R, Bini F, Martinesi M, Chellini F, Luciani G, Sbrana F, Zecchi-Orlandini S, Francini F, Meacci E. Regulation of transient receptor potential canonical channel 1 (TRPC1) by sphingosine 1-phosphate in C2C12 myoblasts and its relevance for a role of mechanotransduction in skeletal muscle differentiation. J Cell Sci. 2009;122:1322–1333. doi: 10.1242/jcs.035402. [DOI] [PubMed] [Google Scholar]
- [149].Waddell LB, Lemckert FA, Zheng XF, Tran J, Evesson FJ, Hawkes JM, Lek A, Street NE, Lin P, Clarke NF, Landstrom AP, Ackerman MJ, Weisleder N, Ma J, North KN, Cooper ST. Dysferlin, annexin A1, and mitsugumin 53 are upregulated in muscular dystrophy and localize to longitudinal tubules of the T-system with stretch. Journal of neuropathology and experimental neurology. 2011;70:302–313. doi: 10.1097/NEN.0b013e31821350b0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [150].Nuno DW, England SK, Lamping KG. RhoA localization with caveolin-1 regulates vascular contractions to serotonin. American journal of physiology. Regulatory, integrative and comparative physiology. 2012;303:R959–967. doi: 10.1152/ajpregu.00667.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [151].Jessica AF, Xiaoqiang Y, Geraldine R. TRPV4 and the regulation of vascular tone. Journal of cardiovascular pharmacology. 2012 doi: 10.1097/FJC.0b013e318279ba42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [152].Kuszczak I, Samson SE, Pande J, Shen DQ, Grover AK. Sodium-calcium exchanger and lipid rafts in pig coronary artery smooth muscle. Biochimica et biophysica acta. 2011;1808:589–596. doi: 10.1016/j.bbamem.2010.11.029. [DOI] [PubMed] [Google Scholar]
- [153].Sharma P, Ghavami S, Stelmack GL, McNeill KD, Mutawe MM, Klonisch T, Unruh H, Halayko AJ. beta-Dystroglycan binds caveolin-1 in smooth muscle: a functional role in caveolae distribution and Ca2+ release. J Cell Sci. 2010;123:3061–3070. doi: 10.1242/jcs.066712. [DOI] [PubMed] [Google Scholar]
- [154].Stuermer CA. The reggie/flotillin connection to growth. Trends Cell Biol. 2010;20:6–13. doi: 10.1016/j.tcb.2009.10.003. [DOI] [PubMed] [Google Scholar]
- [155].Bassani S, Passafaro M. TSPAN7: A new player in excitatory synapse maturation and function. Bioarchitecture. 2012;2:95–97. doi: 10.4161/bioa.20829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [156].Lingwood D, Ries J, Schwille P, Simons K. Plasma membranes are poised for activation of raft phase coalescence at physiological temperature. Proceedings of the National Academy of Sciences of the United States of America. 2008;105:10005–10010. doi: 10.1073/pnas.0804374105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [157].Sekino Y, Kojima N, Shirao T. Role of actin cytoskeleton in dendritic spine morphogenesis. Neurochem Int. 2007;51:92–104. doi: 10.1016/j.neuint.2007.04.029. [DOI] [PubMed] [Google Scholar]
- [158].Grider MH, Park D, Spencer DM, Shine HD. Lipid raft-targeted Akt promotes axonal branching and growth cone expansion via mTOR and Rac1, respectively. J Neurosci Res. 2009 doi: 10.1002/jnr.22140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [159].Davare MA, Fortin DA, Saneyoshi T, Nygaard S, Kaech S, Banker G, Soderling TR, Wayman GA. Transient receptor potential canonical 5 channels activate Ca2+/calmodulin kinase Igamma to promote axon formation in hippocampal neurons. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2009;29:9794–9808. doi: 10.1523/JNEUROSCI.1544-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [160].Nguyen L, He Q, Meiri KF. Regulation of GAP-43 at serine 41 acts as a switch to modulate both intrinsic and extrinsic behaviors of growing neurons, via altered membrane distribution. Molecular and cellular neurosciences. 2009;41:62–73. doi: 10.1016/j.mcn.2009.01.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [161].Shelly M, Lim BK, Cancedda L, Heilshorn SC, Gao H, Poo MM. Local and long-range reciprocal regulation of cAMP and cGMP in axon/dendrite formation. Science. 2010;327:547–552. doi: 10.1126/science.1179735. [DOI] [PubMed] [Google Scholar]
- [162].Murray AJ, Tucker SJ, Shewan DA. cAMP-dependent axon guidance is distinctly regulated by Epac and protein kinase A. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2009;29:15434–15444. doi: 10.1523/JNEUROSCI.3071-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [163].Ayling LJ, Briddon SJ, Halls ML, Hammond GR, Vaca L, Pacheco J, Hill SJ, Cooper DM. Adenylyl cyclase AC8 directly controls its micro-environment by recruiting the actin cytoskeleton in a cholesterol-rich milieu. J Cell Sci. 2012;125:869–886. doi: 10.1242/jcs.091090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [164].Lai Y, Cheng K, Kisaalita W. Three dimensional neuronal cell cultures more accurately model voltage gated calcium channel functionality in freshly dissected nerve tissue. PloS one. 2012;7:e45074. doi: 10.1371/journal.pone.0045074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [165].Head BP, Patel HH, Tsutsumi YM, Hu Y, Mejia T, Mora RC, Insel PA, Roth DM, Drummond JC, Patel PM. Caveolin-1 expression is essential for N-methyl-D-aspartate receptor-mediated Src and extracellular signal-regulated kinase 1/2 activation and protection of primary neurons from ischemic cell death. FASEB journal : official publication of the Federation of American Societies for Experimental Biology. 2008;22:828–840. doi: 10.1096/fj.07-9299com. [DOI] [PubMed] [Google Scholar]
- [166].Head BP, Peart JN, Panneerselvam M, Yokoyama T, Pearn ML, Niesman IR, Bonds JA, Schilling JM, Miyanohara A, Headrick J, Ali SS, Roth DM, Patel PM, Patel HH. Loss of caveolin-1 accelerates neurodegeneration and aging. PloS one. 2010;5:e15697. doi: 10.1371/journal.pone.0015697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [167].Francesconi A, Kumari R, Zukin RS. Regulation of group I metabotropic glutamate receptor trafficking and signaling by the caveolar/lipid raft pathway. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2009;29:3590–3602. doi: 10.1523/JNEUROSCI.5824-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [168].Boulware MI, Kordasiewicz H, Mermelstein PG. Caveolin proteins are essential for distinct effects of membrane estrogen receptors in neurons. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2007;27:9941–9950. doi: 10.1523/JNEUROSCI.1647-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [169].Hong YH, Kim JY, Lee JH, Chae HG, Jang SS, Jeon JH, Kim CH, Kim J, Kim SJ. Agonist-induced internalization of mGluR1alpha is mediated by caveolin. Journal of neurochemistry. 2009;111:61–71. doi: 10.1111/j.1471-4159.2009.06289.x. [DOI] [PubMed] [Google Scholar]
- [170].Mermelstein PG. Membrane-localised oestrogen receptor alpha and beta influence neuronal activity through activation of metabotropic glutamate receptors. J Neuroendocrinol. 2009;21:257–262. doi: 10.1111/j.1365-2826.2009.01838.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [171].Langhorst MF, Reuter A, Jaeger FA, Wippich FM, Luxenhofer G, Plattner H, Stuermer CA. Trafficking of the microdomain scaffolding protein reggie-1/flotillin-2. European journal of cell biology. 2008;87:211–226. doi: 10.1016/j.ejcb.2007.12.001. [DOI] [PubMed] [Google Scholar]
- [172].Riento K, Frick M, Schafer I, Nichols BJ. Endocytosis of flotillin-1 and flotillin-2 is regulated by Fyn kinase. J Cell Sci. 2009;122:912–918. doi: 10.1242/jcs.039024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [173].Munderloh C, Solis GP, Bodrikov V, Jaeger FA, Wiechers M, Malaga-Trillo E, Stuermer CA. Reggies/flotillins regulate retinal axon regeneration in the zebrafish optic nerve and differentiation of hippocampal and N2a neurons. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2009;29:6607–6615. doi: 10.1523/JNEUROSCI.0870-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [174].Langhorst MF, Jaeger FA, Mueller S, Sven Hartmann L, Luxenhofer G, Stuermer CA. Reggies/flotillins regulate cytoskeletal remodeling during neuronal differentiation via CAP/ponsin and Rho GTPases. European journal of cell biology. 2008;87:921–931. doi: 10.1016/j.ejcb.2008.07.001. [DOI] [PubMed] [Google Scholar]
- [175].Schrock Y, Solis GP, Stuermer CA. Regulation of focal adhesion formation and filopodia extension by the cellular prion protein (PrPC) FEBS letters. 2009;583:389–393. doi: 10.1016/j.febslet.2008.12.038. [DOI] [PubMed] [Google Scholar]
- [176].Keifer J, Zheng Z. AMPA receptor trafficking and learning. The European journal of neuroscience. 2010;32:269–277. doi: 10.1111/j.1460-9568.2010.07339.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [177].Pryor S, McCaffrey G, Young LR, Grimes ML. NGF causes TrkA to specifically attract microtubules to lipid rafts. PloS one. 2012;7:e35163. doi: 10.1371/journal.pone.0035163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [178].Xu X, Warrington AE, Wright BR, Bieber AJ, Van Keulen V, Pease LR, Rodriguez M. A human IgM signals axon outgrowth: coupling lipid raft to microtubules. Journal of neurochemistry. 2011;119:100–112. doi: 10.1111/j.1471-4159.2011.07416.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [179].Molnar E, Swamy M, Holzer M, Beck-Garcia K, Worch R, Thiele C, Guigas G, Boye K, Luescher IF, Schwille P, Schubert R, Schamel WW. Cholesterol and Sphingomyelin Drive Ligand-independent T-cell Antigen Receptor Nanoclustering. The Journal of biological chemistry. 2012;287:42664–42674. doi: 10.1074/jbc.M112.386045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [180].Tomassian T, Humphries LA, Liu SD, Silva O, Brooks DG, Miceli MC. Caveolin-1 orchestrates TCR synaptic polarity, signal specificity, and function in CD8 T cells. Journal of immunology. 2011;187:2993–3002. doi: 10.4049/jimmunol.1101447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [181].Lim TS, Mortellaro A, Lim CT, Hammerling GJ, Ricciardi-Castagnoli P. Mechanical interactions between dendritic cells and T cells correlate with T cell responsiveness. Journal of immunology. 2011;187:258–265. doi: 10.4049/jimmunol.1100267. [DOI] [PubMed] [Google Scholar]
- [182].Chichili GR, Westmuckett AD, Rodgers W. T cell signal regulation by the actin cytoskeleton. The Journal of biological chemistry. 2010;285:14737–14746. doi: 10.1074/jbc.M109.097311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [183].Lin SL, Chien CW, Han CL, Chen ES, Kao SH, Chen YJ, Liao F. Temporal proteomics profiling of lipid rafts in CCR6-activated T cells reveals the integration of actin cytoskeleton dynamics. Journal of proteome research. 2010;9:283–297. doi: 10.1021/pr9006156. [DOI] [PubMed] [Google Scholar]
- [184].Ohnuma K, Uchiyama M, Hatano R, Takasawa W, Endo Y, Dang NH, Morimoto C. Blockade of CD26-mediated T cell costimulation with soluble caveolin-1-Ig fusion protein induces anergy in CD4+T cells. Biochemical and biophysical research communications. 2009;386:327–332. doi: 10.1016/j.bbrc.2009.06.027. [DOI] [PubMed] [Google Scholar]
- [185].Fooksman DR, Vardhana S, Vasiliver-Shamis G, Liese J, Blair DA, Waite J, Sacristan C, Victora GD, Zanin-Zhorov A, Dustin ML. Functional anatomy of T cell activation and synapse formation. Annual review of immunology. 2010;28:79–105. doi: 10.1146/annurev-immunol-030409-101308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [186].Huse M, Klein LO, Girvin AT, Faraj JM, Li QJ, Kuhns MS, Davis MM. Spatial and temporal dynamics of T cell receptor signaling with a photoactivatable agonist. Immunity. 2007;27:76–88. doi: 10.1016/j.immuni.2007.05.017. [DOI] [PubMed] [Google Scholar]
- [187].Konstantinidis DG, Pushkaran S, Johnson JF, Cancelas JA, Manganaris S, Harris CE, Williams DA, Zheng Y, Kalfa TA. Signaling and cytoskeletal requirements in erythroblast enucleation. Blood. 2012;119:6118–6127. doi: 10.1182/blood-2011-09-379263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [188].Munday AD, Gaus K, Lopez JA. The platelet glycoprotein Ib-IX-V complex anchors lipid rafts to the membrane skeleton: implications for activation-dependent cytoskeletal translocation of signaling molecules. Journal of thrombosis and haemostasis : JTH. 2010;8:163–172. doi: 10.1111/j.1538-7836.2009.03656.x. [DOI] [PubMed] [Google Scholar]
- [189].Brouckova A, Holada K. Cellular prion protein in blood platelets associates with both lipid rafts and the cytoskeleton. Thrombosis and haemostasis. 2009;102:966–974. doi: 10.1160/TH09-02-0074. [DOI] [PubMed] [Google Scholar]
- [190].Chen X, Shank S, Davis PB, Ziady AG. Nucleolin-mediated cellular trafficking of DNA nanoparticle is lipid raft and microtubule dependent and can be modulated by glucocorticoid. Molecular therapy : the journal of the American Society of Gene Therapy. 2011;19:93–102. doi: 10.1038/mt.2010.214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [191].Wang H, Wang AX, Barrett EJ. Insulin-induced endothelial cell cortical actin filament remodeling: a requirement for trans-endothelial insulin transport. Molecular endocrinology. 2012;26:1327–1338. doi: 10.1210/me.2012-1003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [192].Huang SH, Long M, Wu CH, Kwon-Chung KJ, Chang YC, Chi F, Lee S, Jong A. Invasion of Cryptococcus neoformans into human brain microvascular endothelial cells is mediated through the lipid rafts-endocytic pathway via the dual specificity tyrosine phosphorylation-regulated kinase 3 (DYRK3) The Journal of biological chemistry. 2011;286:34761–34769. doi: 10.1074/jbc.M111.219378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [193].Thaa B, Herrmann A, Veit M. Intrinsic cytoskeleton-dependent clustering of influenza virus M2 protein with hemagglutinin assessed by FLIM-FRET. Journal of virology. 2010;84:12445–12449. doi: 10.1128/JVI.01322-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [194].Ewers H, Helenius A. Lipid-Mediated Endocytosis. Cold Spring Harb Perspect Biol. 2011;3 doi: 10.1101/cshperspect.a004721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [195].Goldston AM, Powell RR, Temesvari LA. Sink or swim: lipid rafts in parasite pathogenesis. Trends in parasitology. 2012;28:417–426. doi: 10.1016/j.pt.2012.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [196].Binz T, Rummel A. Cell entry strategy of clostridial neurotoxins. Journal of neurochemistry. 2009;109:1584–1595. doi: 10.1111/j.1471-4159.2009.06093.x. [DOI] [PubMed] [Google Scholar]
- [197].Brunger AT, Rummel A. Receptor and substrate interactions of clostridial neurotoxins. Toxicon : official journal of the International Society on Toxinology. 2009;54:550–560. doi: 10.1016/j.toxicon.2008.12.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [198].Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144:646–674. doi: 10.1016/j.cell.2011.02.013. [DOI] [PubMed] [Google Scholar]
- [199].Su B, Gao L, Meng F, Guo LW, Rothschild J, Gelman IH. Adhesion-mediated cytoskeletal remodeling is controlled by the direct scaffolding of Src from FAK complexes to lipid rafts by SSeCKS/AKAP12. Oncogene. 2012 doi: 10.1038/onc.2012.218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [200].Thompson TC, Tahir SA, Li L, Watanabe M, Naruishi K, Yang G, Kadmon D, Logothetis CJ, Troncoso P, Ren C, Goltsov A, Park S. The role of caveolin-1 in prostate cancer: clinical implications. Prostate cancer and prostatic diseases. 2010;13:6–11. doi: 10.1038/pcan.2009.29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [201].Tahir SA, Park S, Thompson TC. Caveolin-1 regulates VEGF-stimulated angiogenic activities in prostate cancer and endothelial cells. Cancer biology & therapy. 2009;8:2286–2296. doi: 10.4161/cbt.8.23.10138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [202].Inder KL, Zheng YZ, Davis MJ, Moon H, Loo D, Nguyen H, Clements JA, Parton RG, Foster LJ, Hill MM. Expression of PTRF in PC-3 Cells modulates cholesterol dynamics and the actin cytoskeleton impacting secretion pathways. Molecular & cellular proteomics : MCP. 2012;11:M111 012245. doi: 10.1074/mcp.M111.012245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [203].Lu YC, Chen HC. Involvement of lipid rafts in adhesion-induced activation of Met and EGFR. Journal of biomedical science. 2011;18:78. doi: 10.1186/1423-0127-18-78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [204].Mierke CT, Bretz N, Altevogt P. Contractile forces contribute to increased glycosylphosphatidylinositol-anchored receptor CD24-facilitated cancer cell invasion. The Journal of biological chemistry. 2011;286:34858–34871. doi: 10.1074/jbc.M111.245183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [205].Fu Y, Bannach O, Chen H, Teune JH, Schmitz A, Steger G, Xiong L, Barbazuk WB. Alternative splicing of anciently exonized 5S rRNA regulates plant transcription factor TFIIIA. Genome Res. 2009;19:913–921. doi: 10.1101/gr.086876.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [206].Ando K, Obara Y, Sugama J, Kotani A, Koike N, Ohkubo S, Nakahata N. P2Y2 receptor-Gq/11 signaling at lipid rafts is required for UTP-induced cell migration in NG 108–15 cells. The Journal of pharmacology and experimental therapeutics. 2010;334:809–819. doi: 10.1124/jpet.110.167528. [DOI] [PubMed] [Google Scholar]
- [207].Evangelisti E, Wright D, Zampagni M, Cascella R, Bagnoli S, Relini A, Nichino D, Scartabelli T, Nacmias B, Sorbi S, Cecchi C. Lipid Rafts Mediate Amyloid-induced Calcium Dyshomeostasis and Oxidative Stress in Alzheimer's Disease. Current Alzheimer research. 2012 doi: 10.2174/1567205011310020004. [DOI] [PubMed] [Google Scholar]
- [208].Fabelo N, Martin V, Marin R, Santpere G, Aso E, Ferrer I, Diaz M. Evidence for premature lipid raft aging in APP/PS1 double-transgenic mice, a model of familial Alzheimer disease. Journal of neuropathology and experimental neurology. 2012;71:868–881. doi: 10.1097/NEN.0b013e31826be03c. [DOI] [PubMed] [Google Scholar]
- [209].Marin R. Signalosomes in the brain: relevance in the development of certain neuropathologies such as Alzheimer's disease. Frontiers in physiology. 2011;2:23. doi: 10.3389/fphys.2011.00023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [210].Kapoor A, Hsu WM, Wang BJ, Wu GH, Lin TY, Lee SJ, Yen CT, Liang SM, Liao YF. Caveolin-1 regulates gamma-secretase-mediated AbetaPP processing by modulating spatial distribution of gamma-secretase in membrane. Journal of Alzheimer's disease : JAD. 2010;22:423–442. doi: 10.3233/JAD-2010-100531. [DOI] [PubMed] [Google Scholar]
- [211].Hicks DA, Nalivaeva NN, Turner AJ. Lipid rafts and Alzheimer's disease: protein-lipid interactions and perturbation of signaling. Frontiers in physiology. 2012;3:189. doi: 10.3389/fphys.2012.00189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [212].Vetrivel KS, Thinakaran G. Membrane rafts in Alzheimer's disease beta-amyloid production. Biochimica et biophysica acta. 2010;1801:860–867. doi: 10.1016/j.bbalip.2010.03.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [213].Schneider A, Rajendran L, Honsho M, Gralle M, Donnert G, Wouters F, Hell SW, Simons M. Flotillin-dependent clustering of the amyloid precursor protein regulates its endocytosis and amyloidogenic processing in neurons. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2008;28:2874–2882. doi: 10.1523/JNEUROSCI.5345-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [214].Thathiah A, Horre K, Snellinx A, Vandewyer E, Huang Y, Ciesielska M, De Kloe G, Munck S, De Strooper B. beta-arrestin 2 regulates Abeta generation and gamma-secretase activity in Alzheimer's disease. Nature medicine. 2013;19:43–49. doi: 10.1038/nm.3023. [DOI] [PubMed] [Google Scholar]
- [215].Yang X, Askarova S, Lee JC. Membrane biophysics and mechanics in Alzheimer's disease. Molecular neurobiology. 2010;41:138–148. doi: 10.1007/s12035-010-8121-9. [DOI] [PubMed] [Google Scholar]
- [216].de la Haba C, Palacio JR, Martinez P, Morros A. Effect of oxidative stress on plasma membrane fluidity of THP-1 induced macrophages. Biochimica et biophysica acta. 2013;1828:357–364. doi: 10.1016/j.bbamem.2012.08.013. [DOI] [PubMed] [Google Scholar]
- [217].Diaz ML, Fabelo N, Marin R. Genotype-induced changes in biophysical properties of frontal cortex lipid raft from APP/PS1 transgenic mice. Frontiers in physiology. 2012;3:454. doi: 10.3389/fphys.2012.00454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [218].Aliche-Djoudi F, Podechard N, Chevanne M, Nourissat P, Catheline D, Legrand P, Dimanche-Boitrel MT, Lagadic-Gossmann D, Sergent O. Physical and chemical modulation of lipid rafts by a dietary n-3 polyunsaturated fatty acid increases ethanol-induced oxidative stress. Free radical biology & medicine. 2011;51:2018–2030. doi: 10.1016/j.freeradbiomed.2011.08.031. [DOI] [PubMed] [Google Scholar]
- [219].Clarion L, Schindler M, de Weille J, Lolmede K, Laroche-Clary A, Uro-Coste E, Robert J, Mersel M, Bakalara N. 7beta-Hydroxycholesterol-induced energy stress leads to sequential opposing signaling responses and to death of C6 glioblastoma cells. Biochemical pharmacology. 2012;83:37–46. doi: 10.1016/j.bcp.2011.09.022. [DOI] [PubMed] [Google Scholar]
- [220].Wani NA, Kaur J. Reduced levels of folate transporters (PCFT and RFC) in membrane lipid rafts result in colonic folate malabsorption in chronic alcoholism. Journal of cellular physiology. 2011;226:579–587. doi: 10.1002/jcp.22525. [DOI] [PubMed] [Google Scholar]
- [221].Tian G, Kong Q, Lai L, Ray-Chaudhury A, Lin CL. Increased expression of cholesterol 24S-hydroxylase results in disruption of glial glutamate transporter EAAT2 association with lipid rafts: a potential role in Alzheimer's disease. Journal of neurochemistry. 2010;113:978–989. doi: 10.1111/j.1471-4159.2010.06661.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [222].Ferrer I. Altered mitochondria, energy metabolism, voltage-dependent anion channel, and lipid rafts converge to exhaust neurons in Alzheimer's disease. J Bioenerg Biomembr. 2009;41:425–431. doi: 10.1007/s10863-009-9243-5. [DOI] [PubMed] [Google Scholar]
- [223].Li X, Becker KA, Zhang Y. Ceramide in redox signaling and cardiovascular diseases. Cellular physiology and biochemistry : international journal of experimental cellular physiology, biochemistry, and pharmacology. 2010;26:41–48. doi: 10.1159/000315104. [DOI] [PubMed] [Google Scholar]
- [224].Tang Y, Scott DA, Das U, Edland SD, Radomski K, Koo EH, Roy S. Early and selective impairments in axonal transport kinetics of synaptic cargoes induced by soluble amyloid beta-protein oligomers. Traffic. 2012;13:681–693. doi: 10.1111/j.1600-0854.2012.01340.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [225].Sorice M, Garofalo T, Misasi R, Manganelli V, Vona R, Malorni W. Ganglioside GD3 as a raft component in cell death regulation. Anti-cancer agents in medicinal chemistry. 2012;12:376–382. doi: 10.2174/187152012800228670. [DOI] [PubMed] [Google Scholar]
- [226].Fridolfsson HN, Kawaraguchi Y, Ali SS, Panneerselvam M, Niesman IR, Finley JC, Kellerhals SE, Migita MY, Okada H, Moreno AL, Jennings M, Kidd MW, Bonds JA, Balijepalli RC, Ross RS, Patel PM, Miyanohara A, Chen Q, Lesnefsky EJ, Head BP, Roth DM, Insel PA, Patel HH. Mitochondria-localized caveolin in adaptation to cellular stress and injury. FASEB journal : official publication of the Federation of American Societies for Experimental Biology. 2012;26:4637–4649. doi: 10.1096/fj.12-215798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [227].Neumann AK, Itano MS, Jacobson K. Understanding lipid rafts and other related membrane domains. F1000 biology reports. 2010;2:31. doi: 10.3410/B2-31. [DOI] [PMC free article] [PubMed] [Google Scholar]