

Abstract
Whether hiding from predators, or avoiding battlefield casualties, camouflage is widely employed to prevent detection. Disruptive coloration is a seemingly well-known camouflage mechanism proposed to function by breaking up an object's salient features (for example their characteristic outline), rendering objects more difficult to recognize. However, while a wide range of animals are thought to evade detection using disruptive patterns, there is no direct experimental evidence that disruptive coloration impairs recognition. Using humans searching for computer-generated moth targets, we demonstrate that the number of edge-intersecting patches on a target reduces the likelihood of it being detected, even at the expense of reduced background matching. Crucially, eye-tracking data show that targets with more edge-intersecting patches were looked at for longer periods prior to attack, and passed-over more frequently during search tasks. We therefore show directly that edge patches enhance survivorship by impairing recognition, confirming that disruptive coloration is a distinct camouflage strategy, not simply an artefact of background matching.
Keywords: crypsis, background matching, disruptive coloration, vision, eye tracking, edge detection
1. Introduction
Camouflage is a ubiquitous phenomenon in nature, with natural selection favouring animals that can avoid being seen and recognized by predators [1–3]. The most intuitive forms of camouflage are based on blending in with the background (background matching) and on mimicking an irrelevant object in the environment (masquerade). However, camouflage is also thought to be achieved in a wide range of species via disruptive coloration. Disruptive coloration functions by obscuring outlines and creating false boundaries, thereby preventing recognition of an animal's salient features [1–4]. While the concept of disruptive coloration has been standard textbook material for over a century [1,2], with applications ranging from military uniforms and equipment to art [5], empirical verification of the functional mechanism underlying disruptive coloration has remained elusive for two main reasons. First, disruptive coloration invariably occurs in conjunction with background matching, so disentangling their individual contributions to camouflage is therefore challenging [6–8]. Second, disruptive coloration is best defined in terms of its function (impairing recognition) rather than its appearance [7,8]. Consequently, despite repeated suggestions that animals from anteaters to zebra are disruptively coloured, one cannot, based on appearance alone, state that an animal's body pattern is disruptive. Cuthill et al. [9] provided the first field demonstration that artificial prey targets with contrastingly patterned edges (‘edge’ targets) had a greater survivorship under bird predation than control targets without edge-intersecting patches. Similar results were subsequently reported with human subjects visually hunting for artificial moth targets displayed on computer screens [10], wild birds foraging in aviaries [11], and wild birds foraging in their natural habitats [12–16]. Unfortunately, however, there is no experimental evidence to confirm that the enhanced survivorship afforded by such markings was attained through impaired object recognition [17]. Put simply, results showing that edge-intersecting patches enhance survivorship are necessary, but not sufficient. For disruptive coloration to work in the manner prescribed, then object recognition must be impaired, otherwise the theory fails [17].
2. Material and methods
Using eye-tracking technology, we test, for the first time to our knowledge, whether edge-intersecting patches on objects impair their recognition and thereby enhance their survivorship. Eye-tracking is widely used in psychophysics [18] and is particularly well suited to distinguish failure of object recognition from a failure to locate a hidden target ([19,20]; see the electronic supplementary material, figure S1). The eye movements of individual subjects were quantified during each search task using a FaceLab eye tracker (Seeing Machine, Canberra, Australia) which recorded foveal (line of sight) eye positions at a frequency of 60 Hz. Raw eye movement data included both small-scale concentrated eye movements (during a bout of visual fixation) and large-scale eye movements (during saccades). As the amount of visual information is limited when the eye moves fast during a saccade, a velocity-based threshold (see the electronic supplementary material, ‘Tracking of eye movements’) was used to identify and exclude saccades data prior to analysis. Analysis of this filtered eye-tracking data allowed us to quantify three inter-related measures of object recognition, namely: (i) inspection time (the total time that each subject's spent foveating within 1.5° of the target), (ii) number of passes (independent bouts during which foveal vision moved from outside to inside the 1.5° zone around the target, equivalent to the number of independent fixations within the target area; see the electronic supplementary material, figures S2 and S3), and (iii) final inspection time (the time spent foveating within 1.5° of the target, in the bout immediately prior to attacking). If disruptive coloration hinders recognition of a target through breaking up a target's outline, then targets with more disruptive colour patterns should be inspected for longer periods and incur more frequent independent passes from the searcher. Moreover, variation in the above measures should explain some of the observed variation in target survival and total search time.
We monitored the eye movements of 48 human subjects hunting for 63 artificial, bi-coloured triangular moth targets with different coloration patterns placed on tree-trunk backgrounds. The trees and moths were displayed on a computer screen using a Visual Basic 8 GUI, which also recorded (i) if targets were discovered, and (ii) total visual search time (in ms) of the subjects for the discovered targets. When a subject recognized the target, he/she immediately rotated the mouse wheel to stop the timer and then moved the cursor over the target and clicked on it to verify that the target had been correctly located. For each of the 63 moth targets, we quantified three of its inter-related coloration properties (see the electronic supplementary material, figure S4 for a gallery illustrating variation), namely, uniformity (the overall heterogeneity of markings, with low values representing highly mottled appearance), the proportion of dark area in the bicoloured targets and the number of edge-intersecting patches (the number of groups of five or more continuous dark pixels that touch the targets outline, used as our measure of putative ‘disruptive’ coloration, as more edge patches contribute to the break-up of target's outline (sensu [21,22]); see the electronic supplementary material, figure S5).
3. Results
Overall, our 63 moth targets differed in their survivorship (i.e. the proportion of each target that went undiscovered over all presentations; see figure 1a for examples of high and low survivorship targets; 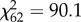 , p < 0.05; range = 0–18%) and mean search time for those found (i.e. total time taken to discover each target type; F62,2544 = 5.05, p < 0.001; means per target 2–10 s). If edge-intersecting patches disrupt object recognition, then we would expect that the survivorship and mean search time of targets should increase with increasing number of edge-intersecting patches, while controlling for other overall aspects of target appearance (in this case, uniformity and proportion of dark area). As predicted, as the number of edge patches increased, so did average survivorship of the target (F1,59 = 4.25, p = 0.044; table 1 and figure 1b), as well as the mean search time for those discovered (F1,59 = 8.36, p = 0.005; table 1 and figure 1c), over and above the effects of target's uniformity and darkness.
, p < 0.05; range = 0–18%) and mean search time for those found (i.e. total time taken to discover each target type; F62,2544 = 5.05, p < 0.001; means per target 2–10 s). If edge-intersecting patches disrupt object recognition, then we would expect that the survivorship and mean search time of targets should increase with increasing number of edge-intersecting patches, while controlling for other overall aspects of target appearance (in this case, uniformity and proportion of dark area). As predicted, as the number of edge patches increased, so did average survivorship of the target (F1,59 = 4.25, p = 0.044; table 1 and figure 1b), as well as the mean search time for those discovered (F1,59 = 8.36, p = 0.005; table 1 and figure 1c), over and above the effects of target's uniformity and darkness.
Figure 1.
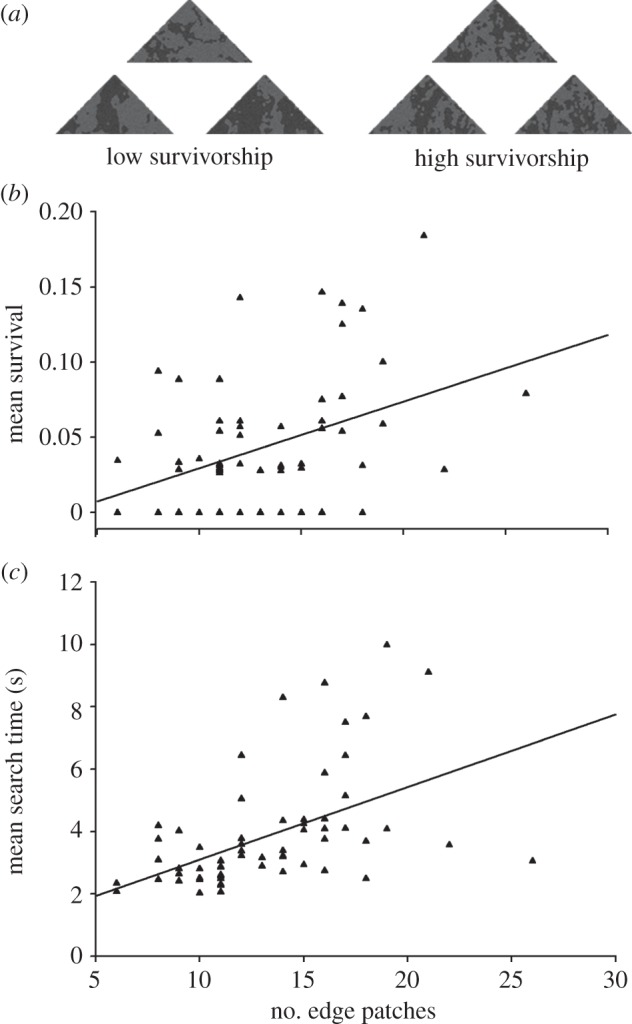
Detection probability of moth targets as a function of edge properties. (a) Examples of moth targets with low and high survival over all trials. Targets with a high number of edge-intersecting patches tended to have (b) higher mean survival, and (c) higher mean search time (of those targets discovered). Each data point represents a target, with individual mean survivorship and search time calculated over 48 independent subject trials. Lines represent fitted least-squares regression, estimated from the fits of general linear models in table 1.
Table 1.
Testing the effects of the moth target's coloration properties on detection and recognition. (Each data column shows the results of the fit of a separate general linear model. All fitted models included three target coloration properties as predictor variables (number of edge-intersecting patches, total proportion of targets dark area and uniformity). These predictors were fitted using type III sums of squares to control for the effect of correlated coloration properties. The geometric means for the detectability and recognizability of each target were calculated across all 63 target presentations. Test statistics reported for the fitted models are F-values (d.f. = 1,59 in all cases), with significance denoted by italics and asterisks. *p < 0.05, **p < 0.01, ***p < 0.001.
| dependent variables |
||||||
|---|---|---|---|---|---|---|
| predictor variables |
detectability |
recognizability |
||||
| type of coloration | target coloration variable | survival (log transformed) | search time (log transformed) | inspection time | final inspection time | no. passes |
| edge | edge-intersecting patches | 4.26, p = 0.044* | 8.36, p = 0.005** | 18.83, p < 0.001*** | 9.34, p = 0.003** | 7.03, p = 0.010** |
| overall | total prop. of dark area | 0.12, p = 0.729 | 0.87, p = 0.360 | 0.08, p = 0.782 | 0.135, p = 0.714 | 0.15, p = 0.700 |
| overall | uniformity | 3.99, p = 0.050* | 7.20, p = 0.009** | 1.401, p = 0.241 | 0.06, p = 0.9390 | 4.48, p = 0.039* |
Targets with more edge-intersecting patches were more difficult to recognize by human subjects. All three recognition metrics, namely total inspection time (F1,59 = 18.83, p < 0.001; figure 2a), final inspection time (F1,59 = 9.34, p = 0.003; figure 2b) and the number of passes (F1,59 = 7.03, p = 0.010; figure 2c) increased as the number of edge-intersecting patches increased, while controlling for other aspects of the targets’ appearance. Targets with more edge-intersecting patches were harder to recognize, despite those targets with a high number of edge patches being less representative of their background (see the electronic supplementary material, figures S6–S8). The number of edge-intersecting patches was not the only target characteristic to affect recognition. Of our background matching metrics, uniformity also affected the number of passes, while target darkness had no effect on any recognition metric (table 1). Lastly, we note that inspection time itself is a significant contributor to total search time (F1,57 = 4.52, p = 0.022; see the electronic supplementary material, figure S9 and table S4), while the number of edge patches interact with inspection time to affect overall search time (F1,57 = 10.41, p < 0.001; see the electronic supplementary material, table S4).
Figure 2.
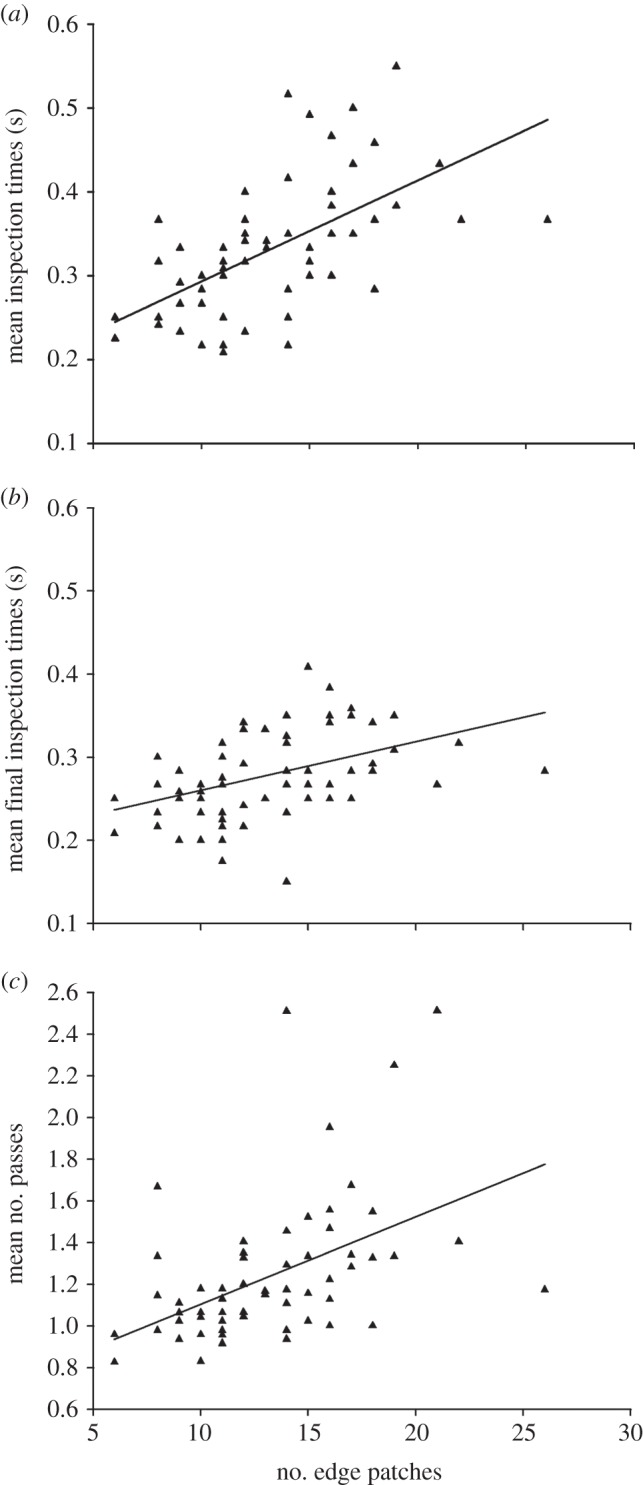
As the number of edge-intersecting patches increase, the recognition of moth targets becomes harder. When visually hunted by human subjects, moth targets with more edge-intersecting patches tend to have (a) a higher mean inspection time, (b) a higher mean final inspection time, and (c) a higher mean number of passes. Lines represent fitted least-squares regression, estimated from the fits of general linear models in table 1.
4. Discussion
Target types with a high number of edge patches survived better and took longer to be discovered, even when they were less representative of the backgrounds ([23]; see the electronic supplementary material, figure S7); therefore, the observed effect of number of edge-intersecting patches could not have arisen as a consequence of targets with a higher number of edge patches being better background matchers. Of the two measures of overall target coloration pattern, lower uniformity (a more mottled appearance) was associated with significantly increased survival and search time, whereas proportion of dark area was not (table 1). Our central finding, that edge-intersecting patches increase survival and search time, remained valid when a more detailed model was fitted to encompass individual trials, with human subject as a random factor (see the electronic supplementary material, tables S2 and S3). Our endpoints are precisely what one would expect if disruption mediates the contribution of object recognition to overall detectability. Intriguingly, inspection time represents a relatively small fraction of total search time, and yet small changes to inspection time have a large effect on total search time (see the electronic supplementary material, figure S9). This outcome may have arisen as a consequence of subjects being reluctant to return to an area that they had previously searched [24]. Alternatively or in addition, the low proportion of inspection time to total search time could miss out other processes that allow object recognition, as our measures exclude the role of subjects’ peripheral vision (see the electronic supplementary material).
Collectively, our results provide, to our knowledge, the first direct evidence that a target's edge patterning hinders object recognition, supporting the view that disruptive coloration can be regarded as a form of camouflage that is functionally distinct [7,8] from background matching. While previous studies have demonstrated that targets with edge markings have higher survivorship [9–16], these unmanipulated disruptive treatments could conceivably have been more representative of the backgrounds on which they are presented. We have overcome this limitation not only by statistically controlling for obvious forms of background matching but also by showing that objects with a high number of edge patches, less representative of their backgrounds, tend to survive at higher rates because they are more difficult to recognize. Only through accounting for how disruption functions can we understand precisely why so many animal species have evolved high-contrast body markings [25] and evaluate what has been dubbed ‘the most important set of principles relating to concealment’ [2].
Acknowledgements
We thank our referees for insightful comments that have helped improve our paper considerably. Francina Jackson, members of the Sherratt Lab and Graeme Ruxton, provided helpful comments on the manuscript.
Our eye-tracking experiment was approved by the Carleton University Psychology Research Ethics Board.
Data accessibility
Data available from the Dryad Digital Repository: http://dx.doi.org/10.5061/dryad.fc109.
Funding statement
The work was supported by NSERC research grants awarded to T.N.S., J-G.J.G. and C.M.H.
References
- 1.Thayer AH. 1909. Concealing coloration in the animal kingdom: an exposition of the laws of disguise through color and pattern. New York, NY: Macmillan [Google Scholar]
- 2.Cott HB. 1940. Adaptive coloration in animals. York, UK: Methuen [Google Scholar]
- 3.Ruxton GD, Sherratt TN, Speed M. 2004. Avoiding attack: the evolutionary ecology of crypsis, warning signals and mimicry. Oxford, UK: Oxford University Press [Google Scholar]
- 4.Troscianko T, Benton CP, Lovell PG, Tolhurst DJ, Pizlo Z. 2009. Camouflage and visual perception. Phil. Trans. R. Soc. B 364, 449–461 (doi:10.1098/rstb.2008.0218) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Behrens RR. 2002. False colors: art, design and modern camouflage. Cedar Falls, IA: Bobolink Books [Google Scholar]
- 6.Silberglied RE, Aniello A, Windsor DM. 1980. Disruptive coloration in butterflies: lack of support in Anartia fatima. Science 209, 617–619 (doi:10.1126/science.209.4456.617) [DOI] [PubMed] [Google Scholar]
- 7.Stevens M. 2007. Predator perception and the interrelation between different forms of protective coloration. Proc. R. Soc. B 274, 1457–1464 (doi:10.1098/rspb.2007.0220) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Stevens M, Merilaita S. 2009. Defining disruptive coloration and distinguishing its functions. Phil. Trans. R. Soc. B 364, 481–488 (doi:10.1098/rstb.2008.0216) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Cuthill IC, Stevens M, Sheppard J, Maddocks T, Párraga CA, Troscianko TS. 2005. Disruptive coloration and background pattern matching. Nature 434, 72–74 (doi:10.1038/nature03312) [DOI] [PubMed] [Google Scholar]
- 10.Fraser S, Callahan A, Klassen D, Sherratt TN. 2007. Empirical tests of the role of disruptive coloration in reducing detectability Proc. R. Soc. B 274, 1325–1331 (doi:10.1098/rspb.2007.0153) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Merilaita S, Lind J. 2005. Background-matching and disruptive coloration, and the evolution of cryptic coloration. Proc. R. Soc. B 272, 665–670 (doi:10.1098/rspb.2004.3000) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Stevens M, Cuthill IC, Windsor AMM, Walker HJ. 2006. Disruptive contrast in animal camouflage. Proc. R. Soc. B 273, 2433–2438 (doi:10.1098/rspb.2006.3614) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Schaefer HM, Stobbe N. 2006. Disruptive coloration provides camouflage independent of background matching. Proc. R. Soc. B 273, 2427–2432 (doi:10.1098/rspb.2006.3615) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Cuthill IC, Stevens M, Windsor AMM, Walker HJ. 2006. The effects of pattern symmetry on detection of disruptive and background-matching coloration. Behav. Ecol. 17, 828–832 (doi:10.1093/beheco/arl015) [Google Scholar]
- 15.Cuthill IC, Szekely A. 2009. Coincident disruptive coloration. Phil. Trans. R. Soc. B 364, 489–496 (doi:10.1098/rstb.2008.0266) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Stevens M, Winney IS, Cantor A, Graham J. 2009. Outline and surface disruption in animal camouflage. Proc. R. Soc. B 276, 781–786 (doi:10.1098/rspb.2008.1450) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Cuthill IC, Troscianko TS. 2011. Animal camouflage: biology meets psychology, computer science and art. In Colour in art, design and nature (eds Brebbia CA, Greated C, Collins MW.), pp. 5–24 Southampton, UK: WIT Press [Google Scholar]
- 18.Duchowski AT. 2002. A breadth-first survey of eye-tracking applications. Behav. Res. Methods Instrum. Comput. 34, 455–470 (doi:10.3758/BF03195475) [DOI] [PubMed] [Google Scholar]
- 19.Credidio HF, Teixeira EN, Reis SDS, Moreira AA, Andrade JS. 2012. Statistical patterns of visual search for hidden objects. Sci. Rep. 2, 1–16 (doi:10.1038/srep00920) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Johnson SP, Amso D, Slemmer JA. 2003. Development of object concepts in infancy: evidence for early learning in an eye-tracking paradigm. Proc. Natl Acad. Sci. USA 100, 10 568–10 573 (doi:10.1073/pnas.1630655100) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Stevens M, Cuthill IC. 2006. Disruptive coloration, crypsis and edge detection in early visual processing. Proc. R. Soc. B 273, 2141–2147 (doi:10.1098/rspb.2006.3556) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Chen HeX, Yung NHC. 2008. Corner detector based on global and local curvature properties. Opt. Eng. 47, 057008 (doi:10.1117/1.2931681) [Google Scholar]
- 23.Merilaita S. 1998. Crypsis through disruptive coloration in an isopod. Proc. R. Soc. Lond. B 265, 1059–1064 (doi:10.1098/rspb.1998.0399) [Google Scholar]
- 24.Klein RM. 2000. Inhibition of return. Trends Cogn. Sci. 4, 138–147 (doi:10.1016/S1364-6613(00)01452-2) [DOI] [PubMed] [Google Scholar]
- 25.Caro T. 2009. Contrasting coloration in terrestrial mammals. Phil. Trans. R. Soc. B 364, 537–548 (doi:10.1098/rstb.2008.0221) [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Data available from the Dryad Digital Repository: http://dx.doi.org/10.5061/dryad.fc109.