

Abstract
The concentration of CXCL12/SDF-1 in the bloodstream is tightly regulated, given its central role in leucocyte and stem/progenitor cell egress from bone marrow and recruitment to sites of inflammation or injury. The mechanism responsible for this regulation is unknown. Here we show that both genetic deletion and pharmacological inhibition of CXCR7, a high-affinity CXCL12 receptor, caused pronounced increases in plasma CXCL12 levels. The rise in plasma CXCL12 levels was associated with an impairment in the ability of leucocytes to migrate to a local source of CXCL12. Using a set of complementary and highly sensitive techniques, we found that CXCR7 protein is expressed at low levels in multiple organs in both humans and mice. In humans, CXCR7 was detected primarily on venule endothelium and arteriole smooth muscle cells. CXCR7 expression on venule endothelium was also documented in immunodeficient mice and CXCR7+/lacZ mice. The vascular expression of CXCR7 therefore gives it immediate access to circulating CXCL12. These studies suggest that endothelial CXCR7 regulates circulating CXCL12 levels and that CXCR7 inhibitors might be used to block CXCL12-mediated cell migration for therapeutic purposes.
Keywords: chemotaxis, CXCL12, CXCR4, CXCR7, endothelial
Introduction
The chemokine CXCL12/SDF-1 has been implicated in many biological processes, including organ development, angiogenesis and vasculogenesis, stem cell biology, leucocyte trafficking, cancer, neurodegenerative disorders, inflammatory disorders, atherosclerosis and HIV pathology.1–9 CXCL12-mediated recruitment of leucocytes and cancer cells from the bloodstream into tissues is a fundamental process in inflammation and tumour metastasis. The concentration of CXCL12 in the bloodstream is tightly controlled, and elevations in plasma CXCL12 levels as little as twofold have been reported to initiate mobilization of pro-angiogenic progenitor cells from the bone marrow to tumours and injured tissues.10,11
CXCL12 binds to the receptor CXCR7 with even higher affinity than the chemokine's first-described receptor CXCR4.12 Unlike CXCR4, which triggers calcium mobilization and chemotaxis in response to CXCL12, CXCR7 does not effectively mediate these processes.12,13 In fact, the CXCR7-mediated cellular responses to CXC12 are largely unknown, although CXCL12 binding does trigger CXCR7 association with β-arrestin 2 and CXCL12/CXCR7 internalization in vitro.13–16 As such, CXCR7 has been described as an atypical chemokine receptor.16–19 CXCR7 has also been shown to associate with CXCR420,21 and regulate CXCR4/CXCL12-mediated processes such as transendothelial migration of human renal progenitors and tumour cells13,22,23 and primordial germ cell migration during zebrafish development,24,25 reflecting a complex interplay between CXCL12, CXCR4 and CXCR7.
Interestingly, treatment of mice with CXCR7-specific compounds that inhibit CXCL12 binding in vitro results in increased CXCL12 levels in the bloodstream.19,26 Wang et al.19 observed a greater than twofold increase in plasma CXCL12 after 2 days of treatment with the CXCR7 inhibitor CCX754; the authors suggested that CXCR7-expressing cells might act as a sink for CXCL12 in spleen. Luker et al.26 observed an approximately 70% increase in plasma CXCL12 levels after 1 week of treatment with the CXCR7 inhibitor CCX771, an effect ascribed by the authors to scavenging of plasma CXCL12 by CXCR7-expressing cells. These results are similar to those obtained in CXCR7−/− zebrafish, which have elevated levels of CXCL12.27,28
Despite over 20 years of research, the expression profile of CXCR7 protein in healthy tissues is still unclear. Initially identified in 1989, mRNA for CXCR7 (RDC1) has been detected by Northern blotting in multiple tissues in both dog and mouse.12,29,30 In these species, CXCR7 mRNA is most abundant in heart and kidney, and is also expressed in spleen, lung, brain, testis and ovary, but not in liver or thymus. (CXCR7 mRNA expression patterns in healthy human organs have not been reported.) However, in contrast to the mRNA expression patterns, CXCR7 protein was not detected in healthy mouse organs, raising the possibility that CXCR7 is subject to post-transcriptional regulation.12 By immunohistochemistry (IHC), CXCR7 protein has been detected in human tonsil,31 brain,32 placenta,33 kidney34 and umbilical cord.18 However, the CXCR7 antibodies used in some of these studies are not CXCR7-specific,35 and accordingly the IHC staining may be inauthentic. We previously reported that CXCR7 protein was expressed on transformed cells and tumour-associated vasculature,12 as well as in various pathological settings (e.g. rejecting renal transplant34 and inflamed joints36). We also reported, using the best validated reagents available, the absence of CXCR7 protein expression in mouse and human leucocytes.37
To study the process by which CXCR7 inhibitors increase plasma CXCL12 levels in mice, and to identify which CXCR7+ cells are responsible for this effect, we set out to expand our previous analysis of CXCR7 protein expression in mouse and human tissues. We used a set of complementary techniques of enhanced sensitivity, which included tyramide-based IHC, [125I]CXCL12 binding and β-galactosidase expression in CXCR7+/lacZ mice. This new and comprehensive analysis confirms our previous observations that solid tumours and tumour-associated vasculature express much higher levels of CXCR7 than healthy tissues, and therefore is easier to detect with less sensitive IHC techniques. However, lower level CXCR7 protein expression was seen in a large number of tissues, although it was restricted to certain cell types, mostly venule endothelial cells, vascular smooth muscle cells and selected epithelial cells.
The results of this study indicate that CXCL12 levels in the bloodstream may be regulated by CXCR7 expressed on the vascular endothelium and that systemic pharmacological inhibition of CXCR7 may be an effective way to modulate CXCL12 and the biological processes it controls.
Materials and methods
Antibodies and reagents
CXCR7 monoclonal antibody (mAb) 11G8, isotype control mAb IgG1 and recombinant CXCL11 and CXCL12 were purchased from R&D Systems (Minneapolis, MN). AMD3100 was purchased from Sigma (St Louis, MO). [125I]CXCL12 was purchased from PerkinElmer (Waltham, MA). Mouse CXCL12 ELISA kits were purchased from RayBiotech (Norcross, GA). The CXCR7-specific compound CCX771 and the CXCR7-inactive control compound CCX704 were synthesized by the ChemoCentryx Medicinal Chemistry Department and have been described previously.13 The compounds were maintained as 10-mm stock solutions in DMSO for in vitro experiments and were formulated in 10% captisol for in vivo experiments (subcutaneous dosing). CXCR7−/− mice were generated at ChemoCentryx and have been described previously.37 All animal procedures were approved by the ChemoCentryx Institutional Animal Care and Use Committee.
CXCL12 uptake assay
CXCL12 (final concentration 1 nm) was added to a flask of human umbilical cord endothelial cells (HUVEC passage 3; Lonza, Inc., Allendale, NJ), after which CCX771 or CCX704 (final concentration 1 μm) was added to the cells. At multiple time-points, an aliquot of the medium was removed from the flask and frozen. After all of the aliquots were collected, they were thawed and analysed for CXCL12 levels using a CXCL12 ELISA kit (R&D Systems).
In vivo migration assay
Air pouches were created on the backs of BALB/c mice by injection of 3 ml air on day 0 and 1 ml air on days 2, 4 and 5. On day 6, 30 mg/kg CCX771 or vehicle was injected subcutaneously in a distal site and, 1 hr later, 1 ml PBS containing 30 µg CXCL12 was injected into the air pouch. Twenty-four hours later, cells in the air pouch were collected by rinsing the pouch with 4 ml PBS. Cells were counted on a haematocytometer and analysed for myeloid cells by flow cytometry with phycoerythrin-conjugated anti-mouse CD11b (BD Biosciences, San Jose, CA) using standard techniques.
Cell isolation and organ homogenization
Cells were isolated from mouse organs by mechanical dispersion through a 70-μm nylon strainer. Organ homogenates were prepared by mincing the organs and homogenizing the pieces in PBS containing 10 mm EDTA and complete protease inhibitors (Roche Applied Science, Indianapolis, IN) using a Polytron PT3100 (Kinematica, Bohemia, NY). Total protein content in the homogenates was determined with the Coomassie Plus Protein Assay Reagent (Thermo Fisher, Rockford, IL) according to the manufacturer's instructions.
[125I]CXCL12 binding assay
Cells (106 per well) or organ homogenates (200 μg per well) were diluted in Hanks' balanced salt solution containing 0·1% BSA and transferred to 96-well plates (100 μl per well) containing 5 μl per well recombinant CXCL11 or CXCL12 (final concentration 100 nm) or compounds CCX771 or AMD3100 (final concentration 10 μm). Then, 100 μl per well of cold buffer containing 0·025 μCi (1 nm) [125I]CXCL12 (PerkinElmer) was added and the plates were shaken at 4° for 3 hr. Cells were transferred onto polyethyleneimine-treated GF/B glass fibre filters (PerkinElmer) with a cell harvester (Tomtec, Hamden, CT) and washed with washing buffer (25 mm HEPES, 500 mm NaCl, 1 mm CaCl2, 5 mm MgCl2, pH 7·1). Fifty microlitres per well of MicroScint-20 (PerkinElmer) was added to the filters and counts/min were measured on a Packard TopCount scintillation counter (PerkinElmer). This assay is optimized for CXCR7 detection, using very low levels of [125I]CXCL12. Due to the 10× to 20× lower affinity of CXCL12 for CXCR4 than for CXCR7,12 the binding assay does not yield an appreciable CXCR4 signal.
Northern blotting
Multi-tissue human mRNA blots were purchased from Ambion (Austin, TX). A CXCR7 riboprobe containing the full human CXCR7 cDNA12 was generated with the Strip-EZ RNA kit (Ambion), using [32P]UTP (PerkinElmer). After pre-hybridization, blots were incubated overnight at 68° in Ultrahyb (Ambion) containing 28 million counts/min CXCR7 riboprobe, then rinsed sequentially with low- and high-stringency buffers (Ambion) at 68°. Blots were exposed to Hyperfilm MP (GE Healthcare, Piscataway, NJ) at −80° for 22 hr.
In situ detection of β-galactosidase
To detect β-galactosidase, organs from CXCR7+/lacZ mice37 were excised and immersed for 1 hr in PBS containing 3% paraformaldehyde (Sigma). Organs were rinsed in PBS for 10 min and then immersed in β-galactosidase detection reagent (Millipore, Billerica, MA). The following day, the organs were rinsed in water for 10 min and then fixed overnight in 10% neutral-buffered formalin (Sigma). The organs were dehydrated and embedded in paraffin, after which 5-μm sections were cut, deparaffinized and hydrated by conventional methods. Some sections were counterstained with Neutral Red (Sigma). Sections were rinsed with water, dried and coverslipped with Permount (Sigma).
Immunohistochemistry
Mouse organs were fixed in 10% neutral-buffered formalin (Sigma), dehydrated and embedded in paraffin by conventional methods, and then 5-μm sections were cut and transferred to glass slides. Formalin-fixed, paraffin-embedded sections of human tissues were purchased from Biochain (Newark, CA). Sections were deparaffinized with xylenes and hydrated through graded alcohols by conventional methods. For standard IHC, sections were rinsed in water and immersed in Tris-buffered saline (TBS) containing 1% BSA (Sigma) for 10 min. Sections were incubated for 1 hr in antibody diluent (Dako, Carpinteria, CA) containing 10 μg/ml CXCR7 mAb 11G8 or mouse IgG1 isotype control antibody. Sections were rinsed in TBS-Tween and incubated for 30 min in TBS containing biotinylated anti-mouse IgG (Jackson Immunoresearch, West Grove, PA). Sections were rinsed in TBST and incubated for 20 min in TBS containing streptavidin-conjugated alkaline phosphatase (Dako). Sections were rinsed in TBS-Tween and stained with Fuchsin+ reagent (Dako). Sections were rinsed with water, counterstained with Mayer's haematoxylin (Sigma), rinsed with water and coverslipped with Faramount (Dako). In the sensitive IHC protocol, tissue sections were rinsed in water, incubated for 20 min in 10 mm citric acid, 0·05% Tween-20 pH 6 at 98° and slow-cooled to room temperature for 30 min. Sections were rinsed in TBST and stained with 11G8 or isotype control antibody at 2 μg/ml using the Catalyzed Signal Amplification system (Dako), which employs a tyramide-based amplification step. Sections were rinsed, counterstained and coverslipped as described above. Histological evaluation of stained images was performed by Lifespan Biosciences (Seattle, WA). For fluorescence IHC, CB17 SCID mouse spleen was immersed in Cryomatrix (Thermo Fisher) and frozen by immersion in partially frozen isopentane (Sigma). Five-micrometer sections were cut, transferred to glass slides, air-dried, fixed for 10 min in acetone (Sigma), air-dried and immersed in TBS for 5 min. Sections were immersed in TBS containing 1% BSA, 0·1% Triton X-100 and 0·05% Tween-20 (all from Sigma) for 30 min. 11G8 or isotype control antibody (10 μg/ml) was mixed with mouse sinusoidal endothelial cell clone IBL-9/2 (a gift of Peter Balogh, University of Pécs, Hungary) and the mixtures were added to the sections and incubated for 1 hr at room temperature. Sections were rinsed in TBST and incubated for 30 min in TBS containing a mixture of Alexa Fluor 488-conjugated anti-rat IgG and Alexa Fluor 555-conjugated anti-mouse IgG (Invitrogen, Carlsbad, CA; each diluted 1 : 500). Sections were rinsed in PBS and incubated with DAPI (Invitrogen) for 15 min. Sections were rinsed in PBS, transferred to water and coverslipped with GelMount (VWR, West Chester, PA).
Results
Regulation of CXCL12 levels by CXCR7 in vitro and in vivo
Human umbilical cord venous endothelial cells express CXCR7, as demonstrated by their [125I]CXCL12 binding pattern, in which the tracer can only be displaced by excess CXCL11, CXCL12 or the CXCR7-specific molecule CCX771, but not by the CXCR7-inactive chemical analogue CCX704 or the CXCR4 antagonist AMD310013 (Fig. 1a). Addition of CXCL12 to a flask of human umbilical cord venous endothelial cells resulted in a rapid removal of the chemokine from the medium, consistent with previous reports that CXCR7 binds to and internalizes CXCL12 (Fig. 1b). CCX771, but not CCX704, inhibited the removal of CXCL12 (Fig. 1b). To analyse this effect in vivo, the regulation of plasma CXCL12 by CXCR7 was evaluated in two ways. First, plasma CXCL12 levels were compared between CXCR7+/+, CXCR7+/− and CXCR7−/− mice. Mean plasma CXCL12 levels were 421 ± 42, 796 ± 43, and 2393 ± 226 pg/ml, respectively (Fig. 1c). Secondly, CXCR7+/+ mice were treated for 1 hr with CCX771 (Fig. 1d). Mean plasma CXCL12 levels were 422 ± 19 pg/ml in vehicle-treated mice (n = 3) and 1674 ± 102 pg/ml CXCL12 in CCX771-treated mice (n = 9). Hence, genetic deletion or pharmacological inhibition of CXCR7 resulted in fourfold to fivefold increases in plasma CXCL12 levels. Similar effects with several selective CXCR7 inhibitors, including CCX771, have been seen in mice, rats and dogs (data not shown). Hence, CXCR7 regulates plasma CXCL12 levels.
Figure 1.
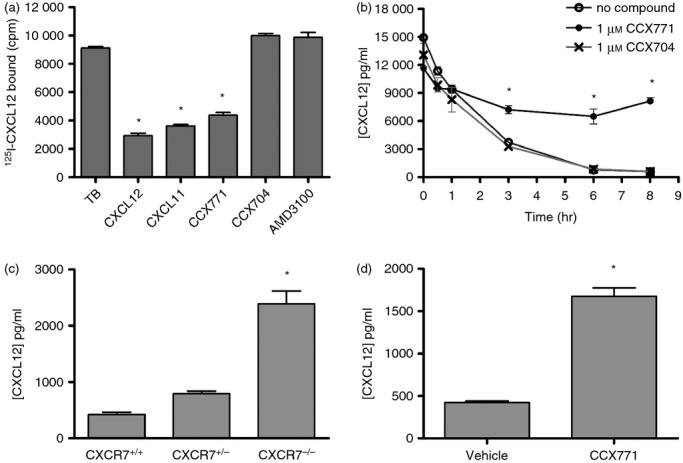
Regulation of CXCL12 levels by CXCR7 in vitro and in vivo. (a) Binding of [125I]CXCL12 to human umbilical cord venous endothelial cells (HUVECs) in the absence (TB = total bound) or presence of various inhibitors. Binding was inhibited by CXCL12, CXCL11 and the CXCR7 inhibitor CCX771, but not by the structurally related but inactive compound CCX704 or the CXCR4 antagonist AMD3100 (*P < 0·05 versus TB; one-way analysis of variance (anova) with Tukey's post tests), indicating that HUVECs express CXCR7. The experiment was repeated twice. (b) Amount of CXCL12 in the culture medium of HUVECs at multiple time-points after addition of CXCL12 and either CCX771, CCX704 or a vehicle control. Only CCX771 blocked the removal of CXCL12 by the HUVEC cells (*P < 0·05 versus CCX704; two way anova with repeated measures and Tukey's post tests). The experiment was repeated twice. (c) CXCL12 levels in plasma of CXCR7+/+, CXCR7+/− and CXCR7−/− mice (10 age-matched mice each, comprising five males and five females). The mice were littermates, born from CXCR7+/− parents and sampled at a variety of ages (12–35 weeks). CXCL12 levels were elevated in CXCR7−/− mice (*P < 0·05 versus CXCR7+/+ mice; two-way anova with Tukey's post tests). (d) CXCL12 levels in plasma of 10-week-old female C3H/HeJ mice treated with 30 mg/kg of CCX771 (nine mice) or vehicle control (three mice), sampled 1-hr post-dosing. CCX771 increased plasma CXCL12 levels (*P < 0·05 versus vehicle; Student's t-test). The experiment was repeated twice. Error bars indicate SEM.
Regulation of leucocyte migration by CXCR7 in vivo
We next investigated whether the increase in plasma CXCL12 levels induced by CCX771 correlated with altered leucocyte migration in vivo. Towards this end, we employed an air pouch model in which CXCL12 was injected into the air pouches of CCX771-treated or vehicle-treated mice. Twenty-four hours after CXCL12 injection, cells that had migrated into the air pouch were counted and analysed by flow cytometry (Fig. 2). CCX771 caused a decrease in the number of CD11b+ myeloid cells, a cell type previously shown to migrate to air pouches containing CXCL12.38 Hence, pharmacological inhibition of CXCR7 can regulate CXCL12-mediated migration of leucocytes in vivo.
Figure 2.
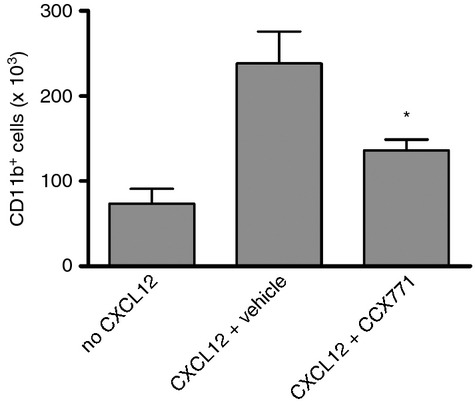
Regulation of leucocyte migration by CXCR7 in vivo. Number of CD11b+ myeloid cells recruited to CXCL12-containing air pouches of 10-week-old male BALB/c mice injected subcutaneously 1 hr earlier with 30 mg/kg of the CXCR7 inhibitor CCX771 or a vehicle control. CCX771 inhibited the migration of CD11b+ cells (*P < 0·05 versus vehicle; one-way analysis of variance with Tukey's post tests) into the air pouch. The number of cells in air pouches lacking CXCL12 indicates non-CXCL12-mediated cell recruitment. n = 7 (no CXCL12), n = 10 (CXCL12 + vehicle) and n = 9 (CXCL12 + CCX771) mice. The experiment was repeated three times. Error bars indicate SEM.
Evaluation of CXCR7 expression in mice
Competitive [125I]CXCL12 binding assay
To investigate the mechanism by which CXCR7 regulates plasma CXCL12 levels, we sought to determine where CXCR7 protein is expressed. Towards this end, we first analysed a subset of organs in healthy mice using a competitive [125I]CXCL12 binding assay. In this assay, CXCR7 is identified by the ability of CXCR7-specific compounds (such as CCX771) and the other CXCR7 chemokine ligand CXCL11, but not the CXCR4-specific antagonist AMD3100, to inhibit the binding of [125I]CXCL12 to cells or cell membranes. In one of our previous studies, we performed the competitive [125I]CXCL12 binding assay on cells isolated from mouse heart, liver and lung by mechanical dispersion and observed no CXCR7-specific [125I]CXCL12 binding to these cells.12 To look for CXCR7 protein inside these cells, as well as cells (e.g. endothelial cells) that may have been excluded using the dispersion method, we homogenized whole organs from multiple strains of mice (C57BL/6, BALB/c, C3H/Hej and 129S) as well as CXCR7−/− mice. Homogenates of wild-type mouse heart, kidney, lung and spleen, but not liver, displayed the CXCR7 profile in the competitive [125I]CXCL12 binding assay (Fig. 3). In contrast, homogenates of the CXCR7−/− mouse organs did not display the CXCR7 binding profile, confirming the specificity of the assay (Fig. 3). Hence, CXCR7 protein is indeed expressed in these organs in healthy mice.
Figure 3.
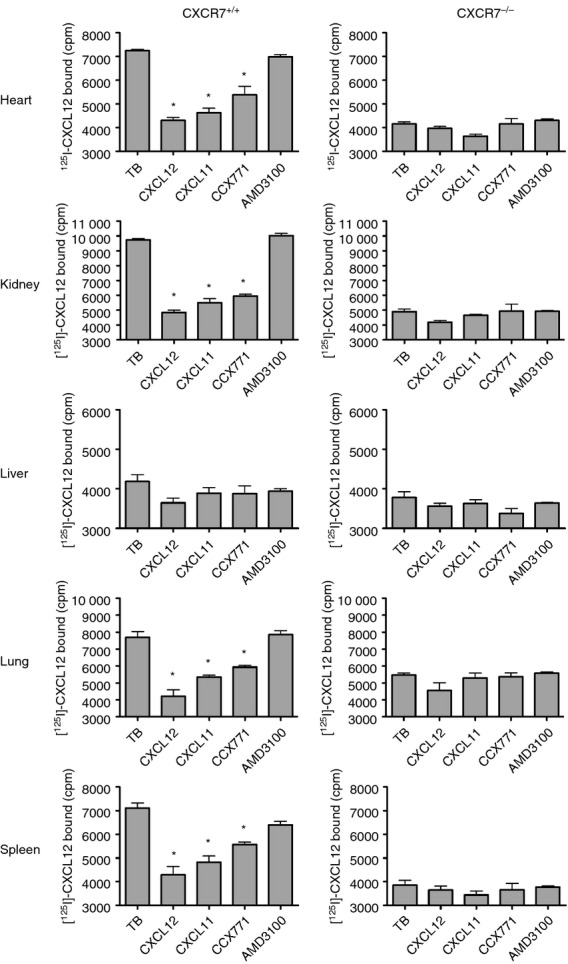
Comparison of CXCR7+/+ and CXCR7−/− mice in the competitive [125I]CXCL12 binding assay. Homogenates from wild-type and CXCR7−/− organs were tested in a competitive [125I]CXCL12 binding assay. Wild-type but not CXCR7−/− mouse heart, kidney, lung and spleen displayed the CXCR7 profile, in that [125I]CXCL12 binding to these organs was inhibited by CXCL12, CXCL11 and the CXCR7 inhibitor CCX771, but not by the CXCR4 antagonist AMD3100 (*P < 0·05 versus TB; one-way analysis of variance with Tukey's post tests). Homogenates from wild-type and CXCR7−/− mouse liver failed to display the CXCR7 profile. The experiment was repeated twice. Error bars indicate SEM.
Immunohistochemistry
To determine which cells express CXCR7 protein in healthy mouse organs, we performed IHC initially using conventional methods. We used CXCR7 mAb 11G8 because other CXCR7 antibodies either do not work or are non-specific in IHC.35 Since 11G8 is a mouse mAb and therefore requires detection with an anti-mouse IgG secondary antibody, we analysed immunodeficient mice in order to avoid detection of endogenous IgG. Of all tissues analysed, only spleen was consistently stained by 11G8 (Fig. 4a). The splenic cells were identified as venous sinusoidal endothelial cells by virtue of co-staining with IBL-9/2, an antibody specific for mouse splenic sinusoidal endothelial cells (Fig. 4b).39
Figure 4.
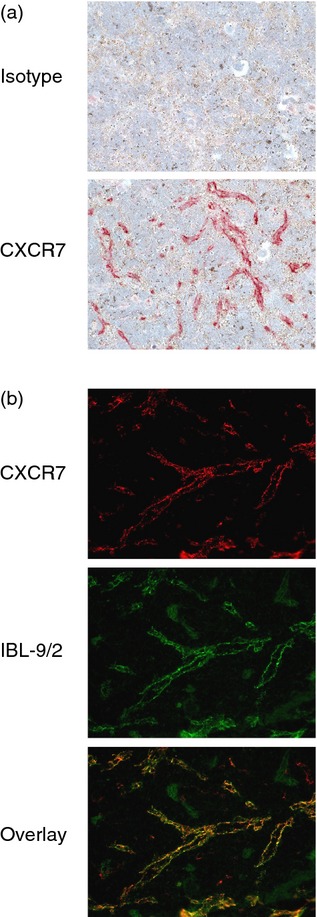
Analysis of CXCR7 protein in mouse spleen by conventional immunohistochemistry (IHC). (a) SCID mouse spleen sections were stained by conventional IHC methods with the CXCR7 monoclonal antibody (mAb) 11G8 or an isotype control mAb. 11G8 stained vessels in the red pulp. Red colour: 11G8 staining. Blue colour: nucleus counterstaining. Original magnification 200×. (b) SCID mouse spleen sections were co-stained with 11G8 and IBL-9/2, an antibody specific for splenic sinusoidal endothelial cells. An overlay of each antibody's staining pattern confirms that 11G8 stained sinusoidal endothelial cells. Original magnification 400×.
Given the expression of CXCR7 mRNA in multiple organs12,29 and CXCR7 protein in mouse heart, kidney and lung (Fig. 3), the lack of CXCR7 staining by conventional IHC was notable. To determine whether healthy tissues express levels of CXCR7 protein too low to be detected by conventional IHC methods, we performed a sensitive IHC procedure including antigen retrieval and tyramide-based signal amplification. Direct comparison of the two methods confirmed the superior sensitivity of the sensitive method in normal mouse tissues (Fig. 5a). Some neoplastic tissues, which are readily stained by 11G8 by conventional IHC methods,40 presumably due to relatively high levels of CXCR7 protein, do not require the sensitive IHC method (Fig. 5a). Using the sensitive IHC method, 11G8 staining was detected in healthy (but immunodeficient) mouse organs previously not stained using the conventional IHC method, including heart, kidney and lung (Fig. 5b). In heart, 11G8 stained endothelial cells lining the atria, valves and some coronary vessels. In lung, 11G8 stained endothelial cells lining a subset of venules. In kidney, 11G8 stained medullary and papillary tubules and a subset of cortical tubules. In kidney glomeruli, 11G8 stained parietal epithelium and, variably, podocytes and/or endothelial cells. 11G8 did not stain mouse liver, consistent with the competitive binding assay (Fig. 3) and Northern blotting,12,29 even when tested against multiple mouse strains (data not shown). 11G8 did not stain lymphatic endothelium (see Supplementary material, Fig. S1).
Figure 5.
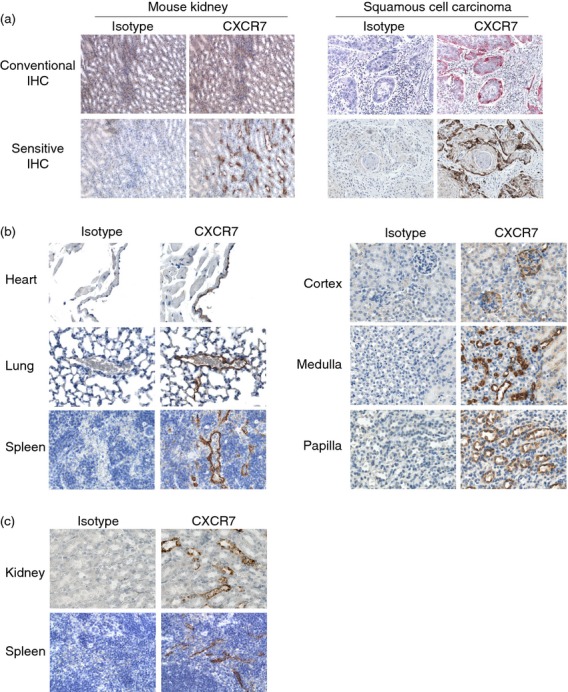
Analysis of CXCR7 protein in mouse organs by the sensitive immunohistochemistry (IHC) method. (a) Sections from SCID mouse organs and human carcinomas were evaluated for 11G8 staining by conventional and sensitive IHC methods. By conventional IHC, 11G8 stained tumours but not normal tissues (red colour: 11G8 staining; blue colour: nucleus counterstaining). With the sensitive IHC method, 11G8 stained both normal tissues and tumours (brown colour: 11G8 staining; blue colour: nucleus counterstaining). Original magnification 200×. (b) Mouse SCID tissue sections were stained with 11G8 by the sensitive IHC method. Left images: 11G8, but not the isotype control mAb, stained endothelial cells of heart cardiac vessels, lung venules and spleen sinusoids. Right images: 11G8, but not the isotype control mAb, stained epithelial cells in a subset of tubules in kidney cortex, medulla and papilla. 11G8 also stained parietal epithelial cells and podocytes and/or endothelial cells in kidney glomeruli (top right image). Brown colour: 11G8 staining; blue colour: nucleus counterstaining. Original magnification 400×. (c) Wild-type c57BL/6 mouse tissue sections were stained with biotin-conjugated 11G8 by the sensitive IHC method. Biotinylated 11G8, but not the biotinylated isotype control mAb, stained epithelial cells in a subset of kidney tubules and splenic sinusoidal endothelial cells. Original magnification 400×.
To perform IHC for CXCR7 on immunocompetent mice, we used a biotinylated conjugate of 11G8 and eliminated the biotinylated anti-mouse IgG secondary antibody. Skipping this step eliminates the ‘mouse-on-mouse’ problem, but also decreases the sensitivity of the IHC procedure because the secondary antibody is polyclonal. When we made this change, only the relatively high CXCR7-expressing cells (splenic sinusoidal endothelium and kidney tubules) were stained (Fig. 5c).
Corroboration of CXCR7 expression
To look for CXCR7 expression in situ without the use of antibodies, we analysed tissue sections from CXCR7+/lacZ mice for β-galactosidase enzymatic activity. β-galactosidase was detected in multiple organs, including heart, kidney, lung and spleen, but not in the liver (Fig. 6). The β-galactosidase patterns were nearly identical to the mouse 11G8 staining patterns (Fig. 5). In the heart, β-galactosidase was expressed by endothelial cells lining coronary and cardiac vessels. In the kidney, β-galactosidase was expressed by epithelial cells in a subset of tubules and podocytes and/or endothelial cells in glomeruli. In the lung, β-galactosidase was expressed by endothelial cells lining a subset of venules. In the spleen, β-galactosidase was expressed by venous sinusoidal endothelial cells. Leucocytes, including splenic marginal zone B cells, did not express β-galactosidase.
Figure 6.
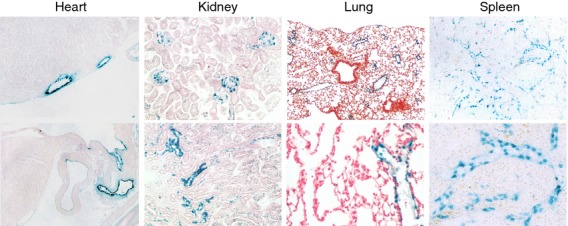
Analysis of β-galactosidase expression in CXCR7+/lacZ mice. Tissue sections from CXCR7+/lacZ mice were evaluated for lacZ expression with an in situ enzymatic assay. β-galactosidase activity was detected in endothelial cells of coronary and cardiac vessels, lung venules and spleen venous sinusoids, as well as kidney glomeruli and tubules. Blue colour: β-galactosidase. Red colour (kidney and lung): nucleus counterstaining. Original magnification 100–400×.
Evaluation of CXCR7 expression in humans
Northern blotting
To assess CXCR7 in healthy human organs, we first analysed CXCR7 mRNA levels by Northern blotting. Northern blots have been published for mouse and dog but not human CXCR7. In the human Northern blot, CXCR7 mRNA was expressed in many human organs (Fig. 7), in a pattern similar to the published mouse CXCR7 Northern blots.12,29 Unlike the mouse Northern blots, however, the human Northern blots contained abundant CXCR7 mRNA in liver but low levels in kidney (Fig. 7).
Figure 7.
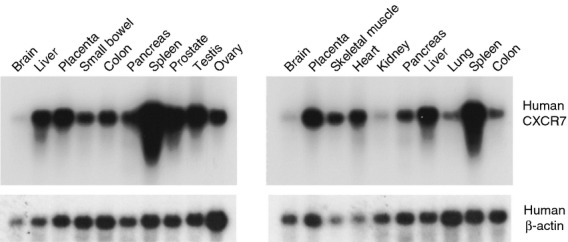
Analysis of CXCR7 mRNA in human tissues by Northern blotting. Two multi-organ mRNA blots were probed for CXCR7 mRNA. CXCR7 mRNA was detected in many human tissues. The blots were then stripped and probed for β-actin mRNA as a loading control.
Immunohistochemistry
Using conventional IHC methods on multiple healthy human tissues, mAb 11G8 stained only splenic sinusoidal endothelial cells (data not shown), as observed in mouse. Using the sensitive IHC method, mAb 11G8 stained venule endothelium in many of the organs analysed (Fig. 8; Table S1). In arterioles, 11G8 usually stained the vascular smooth muscle cells but not the endothelial cells (see Supplementary material, Fig. S2). 11G8 also stained sinusoidal endothelial cells in spleen and lymph nodes but not in bone marrow or liver, and stained endothelial cells lining the heart and its valves but not large arteries or veins. Capillary endothelium was noted in a minor subset of organs. 11G8 did not stain lymphatic endothelium (see Supplementary material, Fig. S3).
Figure 8.
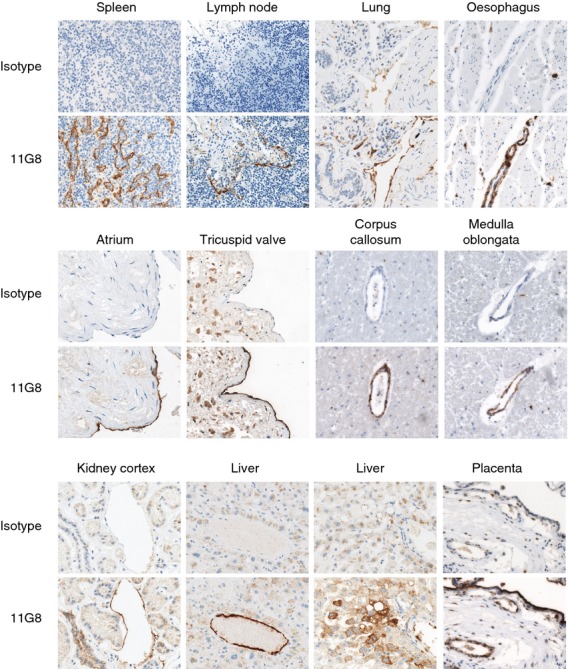
Analysis of CXCR7 protein in human tissues by the sensitive immunohistochemistry (IHC) method. Human tissue arrays were stained with 11G8 by the sensitive IHC method. 11G8, but not the isotype control monoclonal antibody (mAb), stained endothelial cells and/or vascular smooth muscle cells of small vessels in most tissues. 11G8 also stained kidney tubules, liver hepatocytes and placenta trophoblast. In some cases, both 11G8 and the isotype control mAb stained other structures non-specifically (i.e. connective tissue and macrophages within the tricuspid valve leaflet). Brown colour: 11G8 staining; blue colour: nucleus counterstaining. Original magnification 200× and 400×.
11G8 also stained other cell types (Fig. 8). In kidney, 11G8 stained medullary tubules but did not stain glomeruli, in contrast to glomerular expression in mice (Fig. 5b). In liver, 11G8 stained patches of hepatocytes close to portal tracts. In placenta, 11G8 strongly stained trophoblast cells. 11G8 also stained a variety of other cell types, but only in a small number of organs (see Supplementary material, Table S1). Of note, 11G8 did not stain lymphocytes in any organ, including splenic marginal zone B cells. Some macrophages in a subset of non-lymphoid tissues were stained by 11G8; however, some of these macrophages were also stained by the isotype control mAb, raising the possibility that 11G8 staining of macrophages was non-specific. Indeed, 11G8 and the isotype control mAb strongly stained mast cells in multiple tissues (data not shown).
Discussion
In this study we addressed the regulation of CXCL12 levels in the bloodstream by CXCR7 and how manipulation of this receptor can impact leucocyte migration in vivo. Previous studies using CXCR7 gene deletion or CXCR7-specific inhibitors have indicated that CXCL12 binding to and internalization by CXCR7 regulates CXCL12 levels in vivo, both in tissues26–28 and the bloodstream.19,26 However, to date no one has provided a suitable hypothesis for how or where this in vivo regulation takes place. Although we and others have detected CXCR7 in neoplastic tissues, there have been no thorough analyses of CXCR7 protein expression in healthy tissues. In addition, several published analyses of CXCR7 protein expression in individual tissues are not particularly informative given the questionable specificity of the antibodies used.35 To address this, we used a combination of tools, including Northern blotting, tyramide-based IHC, a competitive [125I]CXCL12 binding assay, and β-galactosidase detection in CXCR7+/lacZ mice to probe healthy human and mouse tissues for expression of CXCR7.
We previously postulated that, because CXCR7 protein was not detected on cells isolated by mechanical dispersion from healthy mouse organs, the CXCR7 mRNA found in these organs was not being translated. However, endothelial cells and other interstitial cells do not disperse readily, and therefore the previous analyses may not have included all of the cells in the organs. Using the tools described herein, we now show that CXCR7 mRNA is often translated into protein. However, the levels of CXCR7 protein expressed by non-neoplastic cells seem to be very low, as CXCR7 is rarely detected by conventional IHC methods. In fact, the only cell type in normal human and mouse tissues that is consistently stained by the CXCR7-specific mAb 11G8 using conventional IHC is the splenic venous sinusoidal endothelial cell. In contrast, the mAb readily stains tumour cells and tumour-associated vasculature by conventional IHC.40 Using a sensitive IHC method incorporating antigen retrieval and a tyramide-based amplification step, CXCR7 was detected on various cell types in multiple organs.
In mice, CXCR7 protein was detected in heart, kidney, lung and spleen. Homogenates of these organs from wild-type but not CXCR7−/− mice displayed the CXCR7 profile in the competitive [125I]CXCL12 binding assay. The CXCR7 binding profile was not detected on intact cells isolated by mechanical dispersion, in agreement with previous observations,12 indicating that CXCR7 protein may be located intracellularly and/or that the CXCR7+ cells (such as endothelial cells) are excluded during the dispersion procedure. Using the sensitive IHC method on tissues from immunodeficient mice (immunocompetent mice could not be used due to artefactual detection of endogenous IgG), we detected CXCR7 protein in each of these four organs: in heart, endothelial cells lining cardiac and coronary blood vessels; in kidney, epithelial cells in tubules as well as epithelial cells, podocytes and/or endothelial cells in glomeruli; in lung, endothelial cells in a subset of venules; and in spleen, venous sinusoidal endothelial cells. Interestingly, CXCR7 was not detected on lymphatic or capillary endothelium or pericytes in these organs in mice. In genetically modified CXCR7+/lacZ mice, in which the mouse CXCR7 promoter drives lacZ expression, β-galactosidase was expressed by the same cells that were stained by IHC, except that β-galactosidase expression in glomeruli was stronger in endothelial cells/podocytes than in parietal epithelial cells. CXCR7 protein was not detected in mouse liver, which lacks CXCR7 mRNA,12 by any of the three methods.
Since CXCR7 expression can be modulated by Toll-like receptor signalling and inflammatory mediators,41,42 it is possible that the CXCR7 expression patterns we noted in immunodeficient mice might be different in immunocompetent mice. When we performed the sensitive IHC method on tissues from immunocompetent mice using a biotinylated conjugate of CXCR7 mAb 11G8, and omitted the biotinylated anti-mouse IgG secondary antibody, only splenic sinusoids and kidney tubules were stained. The lack of staining of venules was probably the result of the decreased sensitivity of the amended IHC procedure, because the omitted secondary antibody is polyclonal.
In healthy human tissues, CXCR7 protein was detected in the same cells in kidney, heart, lung and spleen as in mice, except that human glomeruli were not stained. Unlike mouse liver, human liver was stained by 11G8, primarily on endothelial cells of central and portal veins, but also on patches of hepatocytes. The expression of CXCR7 in human liver might result from increased pathogen exposure in humans versus mice kept in pathogen-free conditions. In humans, CXCR7 protein was detected on endothelial cells in a subset of venules in many organs, including appendix, brain, colon, oesophagus, gall bladder, lymph node, ovary, placenta, prostate, small intestine, stomach, testis, thymus, tonsil and uterus. CXCR7 protein was not detected on lymphatic endothelium, but was detected on capillary endothelium in a smaller subset of organs. Interestingly, CXCR7 protein was detected in vascular smooth muscle cells lining arterioles in many organs; these cells may be related to CXCR7+ pericytes which surround tumour-associated blood vessels.40 Finally, CXCR7 protein was detected in several other cell types, although non-vascular staining was restricted to relatively few tissues (see Table S1). The expression of CXCR7 in kidney medullary tubules, especially at the cortico-medullary junction, may be because these areas are normally hypoxic,43 as CXCR7 is up-regulated in regions of hypoxia.44–47
In this study we provide data that the chemokine receptor CXCR7 regulates the levels of chemokine CXCL12 in the bloodstream. Compared with wild-type littermates, CXCR7−/− mice exhibited fivefold higher levels of plasma CXCL12, and CXCR7+/− mice exhibited twofold higher levels of plasma CXCL12. Elevation in chemokine levels has been noted in other chemokine receptor knockout mice.48,49 In addition, treatment of wild-type mice with the CXCR7 antagonist CCX771 resulted in a fourfold increase in the plasma levels of CXCL12, in agreement with other studies.19,26 We also observed that CCX771 blocks the uptake of CXCL12 by CXCR7+ endothelial cells in culture, resulting in higher levels of CXCL12 in the culture medium. Finally, we demonstrated that the CCX771-mediated increase in plasma CXCL12 levels in vivo correlated with an inhibition of leucocyte migration to a local site of CXCL12. Although CXCR7 inhibitors can directly regulate chemotaxis of CXCR4+ CXCR7+ tumour cells to CXCL12 in vitro,13 it is unlikely that CCX771 inhibited the in vivo leucocyte migration to CXCL12 directly, because leucocytes do not express detectable CXCR7 protein.37 Instead, the inhibition of leucocyte migration to the local site of CXCL12 may have been caused by the elevation in plasma CXCL12 levels, which has already been shown to affect leucocyte migration in vivo.38
There are a number of ways in which plasma CXCL12 levels might be increased as a result of CXCR7 inhibition/deletion, including direct displacement of CXCL12 bound to vascular CXCR7, inhibition of CXCR7-mediated removal of CXCL12 from the bloodstream, and increased production of CXCL12. Displacement of receptor-bound chemokines has been invoked50 to explain the increased plasma levels of such chemokines following administration of a receptor antagonist. While we have not assessed the displacement mechanism directly, we think it is unlikely to be a major contributor in the case of CXCL12/CXCR7, mainly because the plasma half-life of CXCL12 is approximately 1 hr;38 displacement of CXCR7-bound CXCL12, in the absence of any other effects, should produce a brief spike in CXCL12 levels, not the sustained increase that we and others describe. At this time, we believe that the most likely explanation for these findings is that genetic deletion or pharmacological antagonism of CXCR7 interferes with its ability to bind to and internalize CXCL12. In support of this explanation (and against the direct displacement model), increased CXCL12 plasma levels are seen only when CCX771 plasma levels are high enough to engage ≥ 90% of the CXCR7 molecules – the same concentration of CCX771 required to block CXCR7-mediated CXCL12 uptake in cell culture (data not shown).
DARC and D6 are, like CXCR7, atypical chemokine receptors expressed primarily on endothelium that function to scavenge chemokines from the vessel lumen.51,52 The expression of CXCR7 on blood but not lymphatic vessels suggests that CXCR7 is more like DARC, which is expressed on blood vessels, than D6, which is expressed on lymphatics. However, both CXCR7 and D6 are also expressed on placental trophoblasts. Since trophoblast-bearing D6 facilitates fetal survival by reducing levels of maternal pro-inflammatory chemokines,53,54 it is an intriguing possibility that trophoblast-bearing CXCR7 scavenges maternal CXCL12 during pregnancy.
In summary, we describe here the expression of CXCR7 protein on vascular endothelium in a large number of human and mouse tissues, providing a direct mechanism for this receptor to control the systemic levels of CXCL12. As CXCL12 regulates a number of important biological processes, we anticipate that CCX771 and other CXCR7 inhibitors might have profound biological and therapeutic effects. It is possible that some of the pharmacological effects already described for CCX771 and other CXCR7 ligands in experimental disease models36,55,56 result from systemic regulation of CXCL12/CXCR4-mediated leucocyte homing.
Acknowledgments
Financial support for these experiments was provided by ChemoCentryx, Inc.
Disclosures
All authors are current or former employees of ChemoCentryx, Inc.
Supporting Information
Additional Supporting Information may be found in the online version of this article:
Figure S1. CXCR7 is detected on mouse venule but not capillary or lymphatic endothelium.
Figure S2. CXCR7 is detected on human vascular smooth muscle.
Figure S3. CXCR7 is not detected on human lymphatic endothelium.
Table S1. Immunohistochemical staining of human tissue arrays by 11G8a,b.
References
- 1.Alkhatib G. The biology of CCR5 and CXCR4. Curr Opin HIV AIDS. 2009;4:96–103. doi: 10.1097/COH.0b013e328324bbec. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Braunersreuther V, Mach F, Steffens S. The specific role of chemokines in atherosclerosis. Thromb Haemost. 2007;97:714–21. [PubMed] [Google Scholar]
- 3.Gelmini S, Mangoni M, Serio M, Romagnani P, Lazzeri E. The critical role of SDF-1/CXCR4 axis in cancer and cancer stem cells metastasis. J Endocrinol Invest. 2008;31:809–19. doi: 10.1007/BF03349262. [DOI] [PubMed] [Google Scholar]
- 4.Iwamoto T, Okamoto H, Toyama Y, Momohara S. Molecular aspects of rheumatoid arthritis: chemokines in the joints of patients. FEBS J. 2008;275:4448–55. doi: 10.1111/j.1742-4658.2008.06580.x. [DOI] [PubMed] [Google Scholar]
- 5.Levesque JP, Winkler IG. Mobilization of hematopoietic stem cells: state of the art. Curr Opin Organ Transplant. 2008;13:53–8. doi: 10.1097/MOT.0b013e3282f42473. [DOI] [PubMed] [Google Scholar]
- 6.Miller RJ, Banisadr G, Bhattacharyya BJ. CXCR4 signaling in the regulation of stem cell migration and development. J Neuroimmunol. 2008;198:31–8. doi: 10.1016/j.jneuroim.2008.04.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Mines M, Ding Y, Fan GH. The many roles of chemokine receptors in neurodegenerative disorders: emerging new therapeutical strategies. Curr Med Chem. 2007;14:2456–70. doi: 10.2174/092986707782023686. [DOI] [PubMed] [Google Scholar]
- 8.Schober A. Chemokines in vascular dysfunction and remodeling. Arterioscler Thromb Vasc Biol. 2008;28:1950–9. doi: 10.1161/ATVBAHA.107.161224. [DOI] [PubMed] [Google Scholar]
- 9.Vandercappellen J, Van Damme J, Struyf S. The role of CXC chemokines and their receptors in cancer. Cancer Lett. 2008;267:226–44. doi: 10.1016/j.canlet.2008.04.050. [DOI] [PubMed] [Google Scholar]
- 10.Jin DK, Shido K, Kopp HG, et al. Cytokine-mediated deployment of SDF-1 induces revascularization through recruitment of CXCR4+ hemangiocytes. Nat Med. 2006;12:557–67. doi: 10.1038/nm1400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Orimo A, Gupta PB, Sgroi DC, et al. Stromal fibroblasts present in invasive human breast carcinomas promote tumor growth and angiogenesis through elevated SDF-1/CXCL12 secretion. Cell. 2005;121:335–48. doi: 10.1016/j.cell.2005.02.034. [DOI] [PubMed] [Google Scholar]
- 12.Burns JM, Summers BC, Wang Y, et al. A novel chemokine receptor for SDF-1 and I-TAC involved in cell survival, cell adhesion, and tumor development. J Exp Med. 2006;203:2201–13. doi: 10.1084/jem.20052144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zabel BA, Wang Y, Lewen S, et al. Elucidation of CXCR7-mediated signaling events and inhibition of CXCR4-mediated tumor cell transendothelial migration by CXCR7 ligands. J Immunol. 2009;183:3204–11. doi: 10.4049/jimmunol.0900269. [DOI] [PubMed] [Google Scholar]
- 14.Luker KE, Gupta M, Steele JM, Foerster BR, Luker GD. Imaging ligand-dependent activation of CXCR7. Neoplasia. 2009;11:1022–35. doi: 10.1593/neo.09724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Luker KE, Steele JM, Mihalko LA, Ray P, Luker GD. Constitutive and chemokine-dependent internalization and recycling of CXCR7 in breast cancer cells to degrade chemokine ligands. Oncogene. 2010;29:4599–610. doi: 10.1038/onc.2010.212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Rajagopal S, Kim J, Ahn S, Craig S, Lam CM, Gerard NP, Gerard C, Lefkowitz RJ. Beta-arrestin- but not G protein-mediated signaling by the “decoy” receptor CXCR7. Proc Natl Acad Sci U S A. 2010;107:628–32. doi: 10.1073/pnas.0912852107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Mahabaleshwar H, Boldajipour B, Raz E. Killing the messenger: the role of CXCR7 in regulating primordial germ cell migration. Cell Adh Migr. 2008;2:69–70. doi: 10.4161/cam.2.2.6027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Naumann U, Cameroni E, Pruenster M, Mahabaleshwar H, Raz E, Zerwes HG, Rot A, Thelen M. CXCR7 functions as a scavenger for CXCL12 and CXCL11. PLoS ONE. 2010;5:e9175. doi: 10.1371/journal.pone.0009175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Wang H, Beaty N, Chen S, Qi CF, Masiuk M, Shin DM, Morse HC., 3rd The CXCR7 chemokine receptor promotes B-cell retention in the splenic marginal zone and serves as a sink for CXCL12. Blood. 2012;119:465–8. doi: 10.1182/blood-2011-03-343608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Levoye A, Balabanian K, Baleux F, Bachelerie F, Lagane B. CXCR7 heterodimerizes with CXCR4 and regulates CXCL12-mediated G protein signaling. Blood. 2009;113:6085–93. doi: 10.1182/blood-2008-12-196618. [DOI] [PubMed] [Google Scholar]
- 21.Luker KE, Gupta M, Luker GD. Imaging chemokine receptor dimerization with firefly luciferase complementation. FASEB J. 2009;23:823–34. doi: 10.1096/fj.08-116749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Mazzinghi B, Ronconi E, Lazzeri E, et al. Essential but differential role for CXCR4 and CXCR7 in the therapeutic homing of human renal progenitor cells. J Exp Med. 2008;205:479–90. doi: 10.1084/jem.20071903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zabel BA, Lewen S, Berahovich RD, Jaen JC, Schall TJ. The novel chemokine receptor CXCR7 regulates trans-endothelial migration of cancer cells. Mol Cancer. 2011;10:73. doi: 10.1186/1476-4598-10-73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Boldajipour B, Mahabaleshwar H, Kardash E, et al. Control of chemokine-guided cell migration by ligand sequestration. Cell. 2008;132:463–73. doi: 10.1016/j.cell.2007.12.034. [DOI] [PubMed] [Google Scholar]
- 25.Dambly-Chaudiere C, Cubedo N, Ghysen A. Control of cell migration in the development of the posterior lateral line: antagonistic interactions between the chemokine receptors CXCR4 and CXCR7/RDC1. BMC Dev Biol. 2007;7:23. doi: 10.1186/1471-213X-7-23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Luker KE, Lewin SA, Mihalko LA, Schmidt BT, Winkler JS, Coggins NL, Thomas DG, Luker GD. Scavenging of CXCL12 by CXCR7 promotes tumor growth and metastasis of CXCR4-positive breast cancer cells. Oncogene. 2012;31:4750–8. doi: 10.1038/onc.2011.633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Sanchez-Alcaniz JA, Haege S, Mueller W, et al. Cxcr7 controls neuronal migration by regulating chemokine responsiveness. Neuron. 2011;69:77–90. doi: 10.1016/j.neuron.2010.12.006. [DOI] [PubMed] [Google Scholar]
- 28.Wang Y, Li G, Stanco A, et al. CXCR4 and CXCR7 have distinct functions in regulating interneuron migration. Neuron. 2011;69:61–76. doi: 10.1016/j.neuron.2010.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Heesen M, Berman MA, Charest A, Housman D, Gerard C, Dorf ME. Cloning and chromosomal mapping of an orphan chemokine receptor: mouse RDC1. Immunogenetics. 1998;47:364–70. doi: 10.1007/s002510050371. [DOI] [PubMed] [Google Scholar]
- 30.Libert F, Parmentier M, Lefort A, et al. Selective amplification and cloning of four new members of the G protein-coupled receptor family. Science. 1989;244:569–72. doi: 10.1126/science.2541503. [DOI] [PubMed] [Google Scholar]
- 31.Infantino S, Moepps B, Thelen M. Expression and regulation of the orphan receptor RDC1 and its putative ligand in human dendritic and B cells. J Immunol. 2006;176:2197–207. doi: 10.4049/jimmunol.176.4.2197. [DOI] [PubMed] [Google Scholar]
- 32.Hattermann K, Held-Feindt J, Lucius R, Muerkoster SS, Penfold ME, Schall TJ, Mentlein R. The chemokine receptor CXCR7 is highly expressed in human glioma cells and mediates antiapoptotic effects. Cancer Res. 2010;70:3299–308. doi: 10.1158/0008-5472.CAN-09-3642. [DOI] [PubMed] [Google Scholar]
- 33.Tripathi V, Verma R, Dinda A, Malhotra N, Kaur J, Luthra K. Differential expression of RDC1/CXCR7 in the human placenta. J Clin Immunol. 2009;29:379–86. doi: 10.1007/s10875-008-9258-4. [DOI] [PubMed] [Google Scholar]
- 34.Neusser MA, Kraus AK, Regele H, et al. The chemokine receptor CXCR7 is expressed on lymphatic endothelial cells during renal allograft rejection. Kidney Int. 2010;77:801–8. doi: 10.1038/ki.2010.6. [DOI] [PubMed] [Google Scholar]
- 35.Berahovich R, Penfold ME, Schall T. Nonspecific CXCR7 antibodies. Immunol Lett. 2010;133:112–4. doi: 10.1016/j.imlet.2010.06.010. [DOI] [PubMed] [Google Scholar]
- 36.Watanabe K, Penfold ME, Matsuda A, et al. Pathogenic role of CXCR7 in rheumatoid arthritis. Arthritis Rheum. 2010;62:3211–20. doi: 10.1002/art.27650. [DOI] [PubMed] [Google Scholar]
- 37.Berahovich RD, Wang Y, Jaen JC, Schall TJ. CXCR7 protein is not expressed on human or mouse leukocytes. J Immunol. 2010;185:5130–9. doi: 10.4049/jimmunol.1001660. [DOI] [PubMed] [Google Scholar]
- 38.O'Boyle G, Mellor P, Kirby JA, Ali S. Anti-inflammatory therapy by intravenous delivery of non-heparan sulfate-binding CXCL12. FASEB J. 2009;23:3906–16. doi: 10.1096/fj.09-134643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Balazs M, Horvath G, Grama L, Balogh P. Phenotypic identification and development of distinct microvascular compartments in the postnatal mouse spleen. Cell Immunol. 2001;212:126–37. doi: 10.1006/cimm.2001.1847. [DOI] [PubMed] [Google Scholar]
- 40.Miao Z, Luker KE, Summers BC, et al. CXCR7 (RDC1) promotes breast and lung tumor growth in vivo and is expressed on tumor-associated vasculature. Proc Natl Acad Sci U S A. 2007;104:15735–40. doi: 10.1073/pnas.0610444104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Singh RK, Lokeshwar BL. The IL-8-regulated chemokine receptor CXCR7 stimulates EGFR signaling to promote prostate cancer growth. Cancer Res. 2011;71:3268–77. doi: 10.1158/0008-5472.CAN-10-2769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Xu H, Wu Q, Dang S, et al. Alteration of CXCR7 expression mediated by TLR4 promotes tumor cell proliferation and migration in human colorectal carcinoma. PLoS ONE. 2011;6:e27399. doi: 10.1371/journal.pone.0027399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Brezis M, Rosen S. Hypoxia of the renal medulla – its implications for disease. N Engl J Med. 1995;332:647–55. doi: 10.1056/NEJM199503093321006. [DOI] [PubMed] [Google Scholar]
- 44.Costello CM, Howell K, Cahill E, et al. Lung-selective gene responses to alveolar hypoxia: potential role for the bone morphogenetic antagonist gremlin in pulmonary hypertension. Am J Physiol Lung Cell Mol Physiol. 2008;295:L272–84. doi: 10.1152/ajplung.00358.2007. [DOI] [PubMed] [Google Scholar]
- 45.Costello CM, McCullagh B, Howell K, Sands M, Belperio JA, Keane MP, Gaine S, McLoughlin P. A role for the CXCL12 receptor, CXCR7, in the pathogenesis of human pulmonary vascular disease. Eur Respir J. 2012;39:1415–24. doi: 10.1183/09031936.00044911. [DOI] [PubMed] [Google Scholar]
- 46.Gambaryan N, Perros F, Montani D, et al. Targeting of c-kit+ haematopoietic progenitor cells prevents hypoxic pulmonary hypertension. Eur Respir J. 2011;37:1392–9. doi: 10.1183/09031936.00045710. [DOI] [PubMed] [Google Scholar]
- 47.Zhou D, Wang J, Zapala MA, Xue J, Schork NJ, Haddad GG. Gene expression in mouse brain following chronic hypoxia: role of sarcospan in glial cell death. Physiol Genomics. 2008;32:370–9. doi: 10.1152/physiolgenomics.00147.2007. [DOI] [PubMed] [Google Scholar]
- 48.Ariel A, Fredman G, Sun YP, Kantarci A, Van Dyke TE, Luster AD, Serhan CN. Apoptotic neutrophils and T cells sequester chemokines during immune response resolution through modulation of CCR5 expression. Nat Immunol. 2006;7:1209–16. doi: 10.1038/ni1392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Cardona AE, Sasse ME, Liu L, Cardona SM, Mizutani M, Savarin C, Hu T, Ransohoff RM. Scavenging roles of chemokine receptors: chemokine receptor deficiency is associated with increased levels of ligand in circulation and tissues. Blood. 2008;112:256–63. doi: 10.1182/blood-2007-10-118497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Charo IF, Ransohoff RM. The many roles of chemokines and chemokine receptors in inflammation. N Engl J Med. 2006;354:610–21. doi: 10.1056/NEJMra052723. [DOI] [PubMed] [Google Scholar]
- 51.Hadley TJ, Lu ZH, Wasniowska K, Martin AW, Peiper SC, Hesselgesser J, Horuk R. Postcapillary venule endothelial cells in kidney express a multispecific chemokine receptor that is structurally and functionally identical to the erythroid isoform, which is the Duffy blood group antigen. J Clin Investig. 1994;94:985–91. doi: 10.1172/JCI117465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Nibbs RJ, Kriehuber E, Ponath PD, et al. The β-chemokine receptor D6 is expressed by lymphatic endothelium and a subset of vascular tumors. Am J Pathol. 2001;158:867–77. doi: 10.1016/s0002-9440(10)64035-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Madigan J, Freeman DJ, Menzies F, et al. Chemokine scavenger D6 is expressed by trophoblasts and aids the survival of mouse embryos transferred into allogeneic recipients. J Immunol. 2010;184:3202–12. doi: 10.4049/jimmunol.0902118. [DOI] [PubMed] [Google Scholar]
- 54.Martinez de la Torre Y, Buracchi C, Borroni EM, et al. Protection against inflammation- and autoantibody-caused fetal loss by the chemokine decoy receptor D6. Proc Natl Acad Sci USA. 2007;104:2319–24. doi: 10.1073/pnas.0607514104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Cruz-Orengo L, Holman DW, Dorsey D, et al. CXCR7 influences leukocyte entry into the CNS parenchyma by controlling abluminal CXCL12 abundance during autoimmunity. J Exp Med. 2011;208:327–39. doi: 10.1084/jem.20102010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Sartina E, Suguihara C, Ramchandran S, et al. Antagonism of CXCR7 attenuates chronic hypoxia-induced pulmonary hypertension. Pediatr Res. 2012;71:682–8. doi: 10.1038/pr.2012.30. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.