

With diabetes reaching epidemic proportions, there is an urgent need for reliable treatment strategies for microvascular (retinopathy, neuropathy, and nephropathy) and macrovascular (coronary heart disease and stroke) diabetes complications. The role of diabetic hyperglycemia in the development of complications has been extensively studied and a number of biochemical pathways activated by hyperglycemia have been identified. Among these, the polyol pathway is perhaps the most investigated and the most controversial pathway.
The initial discovery by Ruth van Heyningen (1) of high sorbitol levels in diabetic rat lenses over 50 years ago followed by studies by Jin Kinoshita (2) laid the foundation for the polyol hypothesis in the pathogenesis of diabetes complications. This hypothesis asserts that diabetes complications result, in part, from direct or indirect consequences of sorbitol production from excess glucose by aldose reductase (AR). AR catalyzes the reduction of glucose to sorbitol, the accumulation of which has been postulated to alter the metabolism of myo-inositol, a six-carbon cyclic polyol, in nerve, ocular, and renal tissues (3,4). Depletion of myo-inositol reduces its incorporation into cellular phosphoinositide pools, resulting in chronic elevation of diacylglycerol (DAG), the chief physiological activator of protein kinase C (PKC) (5). Increased oxidation of sorbitol to fructose may also alter cellular redox potential, resulting in an increase in the cytosolic ratio of NADH/NAD+ (Fig. 1). A number of well-designed animal model experiments were performed to test the role of AR in disease pathogenesis (6–12). Most of these studies reported accumulation of sorbitol (or galactitol in galactosemia model) in diabetic tissues, with AR inhibitors (ARIs) producing various degrees of improvement of microvascular and, in later studies, macrovascular complications.
Figure 1.
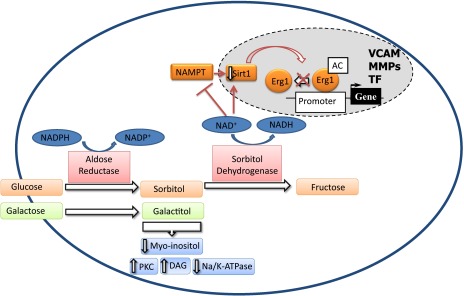
Schematic representation of the role of polyol flux in diabetes complications. AC, acetylation; MMPs, matrix metalloproteinases; Na/K-ATPase, sodium-potassium ATPase; TF, tissue factor; VCAM, vascular cell adhesion molecule.
The polyol pathway, however, turned out to be very evasive. In multiple clinical trials, ARIs showed mixed success or failure in prevention and reversal of long-term diabetes complications (13,14). The short duration of these trials and the low potency (sorbinil) or toxicity (tolrestat) of the test agents used were identified as potential limitations that led to the negative results (15), which led to diminished enthusiasm for the polyol pathway as a therapeutic target. This rather dismal outlook for ARIs was quickly reversed by the outcome of the Aldose Reductase Inhibitor-Diabetes Complications Trial (16) that was performed in Japan. This study showed efficacy for a new ARI agent, epalrestat, in the treatment of diabetic peripheral neuropathy.
Moreover, recent evidence has linked a genetic polymorphism in the AR gene with the susceptibility to diabetes complications. A restriction length polymorphism to an (A-C)n dinucleotide repeat of the microsatellite DNA 5′ of the AR promoter has been identified in a population of Japanese and Chinese subjects with type 2 diabetes and was found to be associated with early-onset retinopathy but not nephropathy (17,18). The C-106T single nucleotide polymorphism in the AR promoter was identified as a susceptibility allele for diabetic retinopathy in Japanese type 2 diabetic patients (19).
The new study by Vedantham et al. (20) in this issue demonstrates a novel role for polyol flux in diabetes. The authors previously showed that overexpression of AR accelerates diabetic atherosclerosis in mice overexpressing human AR (hAR) on an apolipoprotein (apo)E−/− background (21). Using this mouse model and aortic endothelial cell culture studies, the new work demonstrates that a decrease in the cytosolic ratio of NADH/NAD+ due to increased polyol flux inhibits nicotinamide phosphoribosyl transferase (NAMPT), leading to reduced activity of NAD+-dependent deacetylase Sirt-1 and the acetylation and prolonged expression of Egr-1 under high-glucose conditions (Fig. 1). Erg-1 controls gene expression of a number of transcripts that are well established in the pathogeneses of diabetes complications including vascular cell adhesion molecules, matrix metalloproteinases, tumor necrosis factors, and tissue factors (Fig. 1). The authors emphasize that decreases in Sirt-1 activity could result in acetylation of other transcription factors/cofactors, such as forkhead box class O 1, 3 and 4; hypoxia-inducible factor-2α; and nuclear factor-κB, in addition to Erg-1, thereby promoting further tissue pathology.
The results of Vedantham et al. (20) would not explain the deleterious effects of galactosemia in animal models, as galactose is only converted to galactitol, the reaction that utilizes reducing equivalents from NADPH to NADP+ conversion. Importantly, galactitol is not a substrate for sorbitol dehydrogenase thus NAD+ to NADH conversion should not be affected in galactosemic animals (Fig. 1). Interestingly, galactosemic animals have primarily microvascular complications and accumulation of sorbitol was observed in retina, kidney, and peripheral nerves of these animals (3–12). However, no studies have reported the effects of galactosemia on cardiovascular outcomes. Vedantham et al. (20) and previous studies using mice overexpressing hAR in a lipogenic background focused on cardiovascular, but not classic microvascular, diabetes complications (21). In the retina and peripheral nerves, increased polyol flux was associated with PKC-β activation and a decrease in sodium-potassium ATPase levels (3,5). Thus, it would be interesting if AR pathway activation in diabetes results in a different outcome in the microvascular versus macrovascular beds.
Vedantham et al. (20) bring the AR pathway back from exile and give it a new role in the pathogenesis of diabetic macrovascular complications. Since use of ARIs proved to be problematic, new strategies based on this study may include targeting NAD+ levels and/or NAMPT, rather than AR. Furthermore, studies to comprehensively evaluate both microvascular and macrovascular complications are needed to elucidate the involvement of AR in diabetes. However, clearly AR activation as a pathogenic mechanism has made a strong comeback!
Article Information
Funding. Research in J.V.B.’s and M.B.G.’s laboratories is supported by National Institutes of Health (NIH) grant EY-01-6077, Michigan AgBioResearch grant MICL02163 to J.V.B., NIH grants EY-07739 and EY-12601 to M.B.G., and NIH grant DK-09-0730 to M.B.G. and J.V.B.
Duality of Interest. No potential conflicts of interest relevant to this article were reported.
Footnotes
See accompanying original article, p. 761.
References
- 1.van Heyningen R. Formation of polyols by the lens of the rat with ‘sugar' cataract. Nature 1959;184:194–195 [Google Scholar]
- 2.Kinoshita JH. Cataracts in galactosemia. The Jonas S. Friedenwald Memorial Lecture. Invest Ophthalmol 1965;4:786–799 [PubMed] [Google Scholar]
- 3.Greene DA, Lattimer SA, Sima AA. Sorbitol, phosphoinositides, and sodium-potassium-ATPase in the pathogenesis of diabetic complications. N Engl J Med 1987;316:599–606 [DOI] [PubMed] [Google Scholar]
- 4.Haneda M, Kikkawa R, Arimura T, et al. Glucose inhibits myo-inositol uptake and reduces myo-inositol content in cultured rat glomerular mesangial cells. Metabolism 1990;39:40–45 [DOI] [PubMed] [Google Scholar]
- 5.Xia P, Inoguchi T, Kern TS, Engerman RL, Oates PJ, King GL. Characterization of the mechanism for the chronic activation of diacylglycerol-protein kinase C pathway in diabetes and hypergalactosemia. Diabetes 1994;43:1122–1129 [DOI] [PubMed] [Google Scholar]
- 6.Engerman RL, Kern TS, Larson ME. Nerve conduction and aldose reductase inhibition during 5 years of diabetes or galactosaemia in dogs. Diabetologia 1994;37:141–144 [DOI] [PubMed] [Google Scholar]
- 7.Engerman RL, Kern TS, Garment MB. Capillary basement membrane in retina, kidney, and muscle of diabetic dogs and galactosemic dogs and its response to 5 years aldose reductase inhibition. J Diabetes Complications 1993;7:241–245 [PubMed] [Google Scholar]
- 8.Engerman RL, Kern TS. Aldose reductase inhibition fails to prevent retinopathy in diabetic and galactosemic dogs. Diabetes 1993;42:820–825 [DOI] [PubMed] [Google Scholar]
- 9.Kern TS, Engerman RL. Retinal polyol and myo-inositol in galactosemic dogs given an aldose-reductase inhibitor. Invest Ophthalmol Vis Sci 1991;32:3175–3177 [PubMed] [Google Scholar]
- 10.Frank RN, Keirn RJ, Kennedy A, Frank KW. Galactose-induced retinal capillary basement membrane thickening: prevention by sorbinil. Invest Ophthalmol Vis Sci 1983;24:1519–1524 [PubMed] [Google Scholar]
- 11.Robison WG, Jr, Kador PF, Kinoshita JH. Retinal capillaries: basement membrane thickening by galactosemia prevented with aldose reductase inhibitor. Science 1983;221:1177–1179 [DOI] [PubMed] [Google Scholar]
- 12.Kador PF, Akagi Y, Takahashi Y, Ikebe H, Wyman M, Kinoshita JH. Prevention of retinal vessel changes associated with diabetic retinopathy in galactose-fed dogs by aldose reductase inhibitors. Arch Ophthalmol 1990;108:1301–1309 [DOI] [PubMed] [Google Scholar]
- 13.Sorbinil Retinopathy Trial Research Group A randomized trial of sorbinil, an aldose reductase inhibitor, in diabetic retinopathy. Arch Ophthalmol 1990;108:1234–1244 [DOI] [PubMed] [Google Scholar]
- 14.van Gerven JM, Boot JP, Lemkes HH, van Best JA. Effects of aldose reductase inhibition with tolrestat on diabetic retinopathy in a six months double blind trial. Doc Ophthalmol 1994;87:355–365 [DOI] [PubMed] [Google Scholar]
- 15.Pfeifer MA, Schumer MP, Gelber DA. Aldose reductase inhibitors: the end of an era or the need for different trial designs? Diabetes 1997;46(Suppl. 2):S82–S89 [DOI] [PubMed] [Google Scholar]
- 16.Hotta N, Kawamori R, Fukuda M, Shigeta Y; Aldose Reductuase Inhibitor-Diabetes Complications Trial Study Group. Long-term clinical effects of epalrestat, an aldose reductase inhibitor, on progression of diabetic neuropathy and other microvascular complications: multivariate epidemiological analysis based on patient background factors and severity of diabetic neuropathy. Diabet Med 2012;29:1529–1533 [DOI] [PMC free article] [PubMed]
- 17.Ichikawa F, Yamada K, Ishiyama-Shigemoto S, Yuan X, Nonaka K. Association of an (A-C)n dinucleotide repeat polymorphic marker at the 5′-region of the aldose reductase gene with retinopathy but not with nephropathy or neuropathy in Japanese patients with type 2 diabetes mellitus. Diabet Med 1999;16:744–748 [DOI] [PubMed]
- 18.Li Q, Xie P, Huang J, Gu Y, Zeng W, Song H. Polymorphisms and functions of the aldose reductase gene 5′ regulatory region in Chinese patients with type 2 diabetes mellitus. Chin Med J (Engl) 2002;115:209–213 [PubMed] [Google Scholar]
- 19.Watarai A, Nakashima E, Hamada Y, et al. Aldose reductase gene is associated with diabetic macroangiopathy in Japanese type 2 diabetic patients. Diabet Med 2006;23:894–899 [DOI] [PMC free article] [PubMed]
- 20.Vedantham S, Thiagarajan D, Ananthakrishnan R, et al. Aldose reductase drives hyperacetylation of Egr-1 in hyperglycemia and consequent upregulation of proinflammatory and prothrombotic signals. Diabetes 2014;63:761–774 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Vedantham S, Noh H, Ananthakrishnan R, et al. Human aldose reductase expression accelerates atherosclerosis in diabetic apolipoprotein E-/- mice. Arterioscler Thromb Vasc Biol 2011;31:1805–1813 [DOI] [PMC free article] [PubMed] [Google Scholar]