

Abstract
Beginning from two plausible starting points—an uninhabited or inhabited Mars—this paper discusses the possible trajectories of martian habitability over time. On an uninhabited Mars, the trajectories follow paths determined by the abundance of uninhabitable environments and uninhabited habitats. On an inhabited Mars, the addition of a third environment type, inhabited habitats, results in other trajectories, including ones where the planet remains inhabited today or others where planetary-scale life extinction occurs. By identifying different trajectories of habitability, corresponding hypotheses can be described that allow for the various trajectories to be disentangled and ultimately a determination of which trajectory Mars has taken and the changing relative abundance of its constituent environments. Key Words: Mars—Habitability—Liquid water—Planetary science. Astrobiology 14, 182–203.
Introduction
Assessing the habitability of Mars has been an objective of scientists for a long time, but it has recently become a sustained focus in light of data being returned from the planet and growing knowledge about life in extreme environments (e.g., McKay and Davis, 1991; Boston et al., 1992; Jakosky and Shock, 1998; Cabrol et al., 1999; Weiss et al., 2000a; Fairén et al., 2005, 2010; Schulze-Makuch et al., 2005; Dartnell et al., 2007; Nisbet et al., 2007; Davila et al., 2008, 2010; Tosca et al., 2008a; Javaux and Dehant, 2010; Stoker et al., 2010; Cockell et al., 2012; Ulrich et al., 2012; Cousins et al., 2013; Grotzinger et al., 2014; Michalski et al., 2013; Westall et al., 2013). This endeavor is important for elucidating the history of Mars and providing a comparative data point for understanding the geological and biological history of Earth (Taylor, 2011). By investigating the habitability trajectories possible for terrestrial-type planets, we gain information with which to assess the habitability of diverse rocky extrasolar planets (Fridlund, 2008).
Assessing the habitability of martian environments depends on the use of an operational definition of habitable. In this paper, it is taken to be an environment that has the necessary conditions for at least one known organism to be active, where active means metabolically active as maintenance, growth, or reproduction. Habitability is necessarily defined by reference to specific organisms. An anaerobic location is not habitable to an iron-oxidizing microorganism that requires oxygen as the terminal electron acceptor; but, for example, it might be habitable to certain anaerobic iron-reducing microorganisms if ferric compounds and accessible organics are present, provided that all other requirements for the organisms exist, such as supplies of the elements C, H, N, O, P, S, liquid water, and appropriate physical and chemical conditions. The more energy sources and essential nutrients an environment contains, the greater the potential diversity of life that the environment is likely to support.
Habitability is therefore a conservative term, circumscribed by the limits of our knowledge of the range of possible life-forms, their environmental and metabolic capabilities, and their energy requirements (e.g., Hoehler, 2007). A habitable environment could exist for a very short period of time as a transient environment. The suitability of environments to host habitable conditions over geological timescales is a more complex calculus, and it is determined by the interplay of factors such as plate tectonics, the hydrological cycle, and planetary temperature regimens (Nisbet et al., 2007).
A common departure point for investigations on the biological potential of Mars is to assume that Mars was habitable and inhabited and then to assess the surface and subsurface of the planet against known metabolic capabilities of microorganisms on Earth (see discussions in Boston et al., 1992; Nisbet et al., 2007; Javaux and Dehant, 2010).
At the current time, there is no unequivocal evidence for life on past or present Mars. Therefore, any assessment as to how the biological potential of that planet has changed over time must start with the formation of early Mars, when it was uninhabitable, and then consider the diversity of trajectories that result on a planet that became inhabited or remained uninhabited.
One way to achieve such a systematic characterization of trajectories is to bin environments into three types (Cockell et al., 2012; Fig. 1). On a Mars that was always uninhabited, there are two potential types of environment: uninhabitable environments and uninhabited habitats. An uninhabitable environment is defined here to be “an environment in which no known organism can be active.” An uninhabited habitat is defined here to be “an environment capable of supporting the activity of at least one known organism but containing no such organism.”
FIG. 1.

Environments on Mars. Types of environments and habitats that may exist and have existed on Mars in two different scenarios of Mars: one in which it has always been uninhabited and one in which it is and/or was inhabited. The arrows and corresponding text show how locations, even at microscopic scales, could transition between these states as environmental conditions alter (adapted from Cockell et al., 2012).
Uninhabited habitats are rare on Earth (Cockell, 2011). A natural example would be a newly formed lava flow transiently uncolonized by metabolically active cells. An artificial example would be a Petri dish containing uninoculated nutrient agar. Indeed, all microbiological media are recipes for making uninhabited habitats. Cockell et al. (2012) predicted that, if habitable conditions exist on Mars, uninhabited habitats could exist, and they could be detected by showing that all the requirements for habitability are met in a candidate environment but that there are no organics associated with life. The report of a habitable environment in Gale Crater, Mars (Grotzinger et al., 2014), but no definitive detection of biologically associated organics (Ming et al., 2014), would be consistent with an uninhabited habitat, although destruction of organics over billions of years could confound determining whether the environment ever contained life.
On an inhabited Mars, there is an additional environment type to consider: inhabited habitats. An inhabited habitat is defined here to be “an environment containing life whose activity can be supported by that environment.”
A categorization of the possible trajectories of martian habitability through time can therefore be addressed by separating out trajectories based on the existence of combinations of these three environments and then considering, within each class of trajectory, the relative abundance and characteristics of these environments at different scales (Fig. 2).
FIG. 2.

Trajectories of martian habitability. Different trajectories of the habitability of Mars through time, beginning with the branch point of an uninhabited and inhabited Mars. Experimental investigations of Mars will allow for the determination of which trajectory applied to Mars and the relative abundance of the different environments. Trajectory 4 is the trajectory that Earth has taken.
In this synthesis, six major potential trajectories of habitability on Mars are identified. Experimental requirements and their associated hypotheses are discussed that can be used to determine which of these trajectories Mars has taken.
Requirements for Microbial Activity on Mars, Past and Present
To identify trajectories for habitability, three requirements for habitability (liquid water, nutrients, and energy) and four factors that can influence the distribution of habitable conditions on Mars—radiation, pH, the presence of brines, and porosity—are taken into consideration. This allows for (1) the identification of trends, particularly with respect to changing liquid water availability over time, which allows for the selection of the habitability trajectories discussed in the following section, and (2) the identification of physical and chemical factors that might have influenced the changing abundance of uninhabited and inhabited habitats and uninhabitable environments on Mars globally and locally over time.
Liquid water through time
Liquid water is the essential solvent for life, and the history of its abundance on Mars drives the habitability trajectories suggested in this synthesis. Without liquid water, the presence of other requirements for habitability is irrelevant.
Past liquid water
The presence of ancient liquid water on Mars is supported by the observations of phyllosilicates associated with ancient crustal units exposed by craters and erosion (Poulet et al., 2005; Bibring et al., 2006; Ehlmann et al., 2013). For example, the Nili Fossae region of Mars (Fig. 3) exhibits an extraordinary range of minerals and is composed of distinct units. Information on this region has been obtained by the Mars Reconnaissance Orbiter (MRO), Compact Reconnaissance Spectrometer for Mars (CRISM), and the High Resolution Science Experiment (HiRISE) instruments. One unit in the Nili Fossae is a brecciated Fe/Mg-smectite-bearing unit that contains meter- to kilometer-sized blocks of altered and unaltered rock and is inferred to represent the ancient crust of Mars, which was torn up in subsequent impact events (Ehlmann et al., 2009). Fe/Mg-smectite clays are the most abundant clays on Mars, followed by chlorites. A second, olivine-rich unit with evidence for aqueous alteration may be impact melt or early stage lava from the Syrtis Major (Wray and Ehlmann, 2011). The presence of prehnite suggests low-temperature (200–350°C) alteration, and the wide variety of hydration products, including kaolinite, chlorite, mica, opal, zeolites, sulfates, serpentine, smectites, and carbonates, suggests complex water-rock interactions (Ehlmann et al., 2009, 2010, 2011; Wray and Ehlmann, 2011).
FIG. 3.

Geological diversity in the Nili Fossae, Mars. Image of part of a fracture in the Nili Fossae region near 21.9°N, 78.2°E; Compact Reconnaissance Imaging Spectrometer for Mars (CRISM). The top left shows the Isidis basin image (small square) superposed on a Mars Orbiter Laser Altimeter (red, higher elevations; blue, lower). The fracture shown, which is 11 km at its narrowest point (top right), is overlain on a Viking digital image (lower left) to show topography. Top right shows results in infrared channels, false-colored. Bright green is phyllosilicates, yellow-brown is olivines, purple is pyroxenes. The CRISM data are superposed on High-Resolution Imaging Science Experiment (HiRISE) (lower right), showing that phyllosilicates are in small eroded outcrops of rock and olivines in sand dunes. (Image credit: NASA/JPL/JHUAPL/Brown University.)
Asteroid and comet impact craters, which can be regarded as “nature's drill,” provide insights into the composition of the martian subsurface and its geochemical characteristics in the ancient past. Central peaks of craters from other regions of Mars show indications of hydrated phases and hydrothermal alteration products (Ehlmann et al., 2011; Rogers, 2011; Quantin et al., 2012; Osinski et al., 2013) (Fig. 4). The central uplift of equatorial Leighton Crater has excavated material from 6 km depth in the martian subsurface. CRISM data show the presence of carbonates, kaolinite-group elements, and Fe/Mg-bearing silicates consistent with serpentine, chlorite, vermiculite, and pumpellyite (Mustard et al., 2009). Impact central uplifts show that the greatest alteration in the Noachian subsurface occurred at depths greater than ∼5 km (Ehlmann et al., 2011) with material at shallower depths (∼2 km) less altered, which suggests an unsaturated subsurface zone (Rogers, 2011).
FIG. 4.

Access into the subsurface of Mars with crater central uplifts. False-color image of the interior (approximately 460 m across) of an unnamed crater on the central uplift of the 467 km diameter Huygens Crater imaged by HiRISE on the Mars Reconnaissance Orbiter. The light-toned rocks contain carbonates and have been excavated in the central uplift from ∼5 km depth. (Image credit: NASA/JPL-Caltech/University of Arizona.)
The presence of deformation bands (Okubo et al., 2009) is suggested to show that water flow in the past subsurface of Mars might have been influenced, and channeled, by these features. Discoloration along the boundaries of the bands is interpreted to show aqueous alteration of primary minerals (Okubo et al., 2009). Variations in discoloration along the bands are taken to suggest heterogeneity in past martian subsurface water flow and spatial differences in subsurface water geochemistry.
Many of these subsurface processes may have occurred in closed systems, which could potentially have limited habitat connectivity. The lack of chloride and sulfates associated with these deposits, the latter generally found in Hesperian and Amazonian terrains exposed to surface conditions (although there are examples of interbedded clays and sulfates), is further evidence that sedimentary mineral formation did not occur and that many of these systems were closed, that is, subsurface and isolated from the surface atmospheric conditions (Poulet et al., 2005; Bibring et al., 2006; Ehlmann et al., 2013). These observations are important because, if any of these closed systems were habitable, they were possibly localized, isolated uninhabited habitats. For trajectories that assume an inhabited Mars, a lack of connectivity between these closed systems and inhabited regions could have increased the abundance of uninhabited habitats.
A non-exhaustive list of other evidence for ancient water could include the presence of valley networks (Sharp and Malin, 1975; Carr, 1986), a possible northern ocean (Head et al., 1999; Clifford and Parker, 2001), lakes (Cabrol and Grin, 2001, 2002), and evidence of seas (Clifford and Parker, 2001; Schwenzer and Kring, 2009). Evidence for ancient water was comprehensively reviewed by Lasue et al. (2013). Direct evidence for liquid water in the late Noachian/early Hesperian is shown by the presence of sedimentation and clay formation in Gale Crater (Grotzinger et al., 2014; Vaniman et al., 2014).
As the Noachian transitioned into the Hesperian, surface water bodies became less abundant; nevertheless, there is evidence for groundwater activity. For example, layered terrains in the Burns Formation, Meridiani Planum, Mars, are interpreted to be sandstones formed in shallow fluvial or eolian systems and have been the subject of intense geochemical and geological discussion (e.g., Grotzinger et al., 2005; McLennan and Grotzinger, 2008). The widespread presence of sulfates and observations of hematite concretions (Arvidson et al., 2006) support a model of groundwater activity. During the Hesperian, the hydrological cycle was generally characterized by low water-rock ratio interactions in the near-surface environment, which would have led to localized acidity (discussed under the section on pH) associated with ferric sulfates (Hurowitz and McLennan, 2007).
Catastrophic outflow channels provide particularly compelling evidence for the occurrence of subsurface and surface water since the Noachian (Fig. 5). Geomorphologically, these latter features begin from a fracture or region of chaotic terrain and consist of broad depressions tens to thousands of kilometers long with streamlined islands and deposits around craters along their beds (Carr, 1986, 1996; Tanaka 1986; Hartmann and Neukum, 2001). The mechanisms by which they might have been formed include the release of groundwater from the cryosphere by impact events, earthquakes, or magmatic intrusions. Some of these channels date back to greater than 3 billion years ago (Lasue et al., 2013). Some, however, such as those associated with Athabasca Valles, may have had activity just a few million years ago (Burr et al., 2002; Neukum et al., 2010). If water is the mechanism of their formation, then outflow channels would suggest the presence of liquid water in the subsurface from the early Hesperian through to the geologically very recent past in catastrophic episodes.
FIG. 5.

Large bodies of subsurface water on Mars since the Noachian. Aromatum Chaos, Mars (1°7′S, ∼43°8′W) images by the Viking orbiter (Mars Digital Image Model). The outflow channel feeds into Hydroates Chaos, ultimately extending onto the plains of Chryse Planitia. (Image credit: NASA.)
Evidence for short-duration water-rock interactions after the Noachian is also found in martian meteorites, for example, secondary alteration textures in the nakhlite meteorites that record events in the Amazonian ∼600 million years ago (Changela and Bridges, 2010).
However, as with Noachian water sources, the connectivity of these post-Noachian water sources is not known. This has important implications for proposed habitability trajectories, since unconnected habitable conditions would imply the widespread presence of uninhabited habitats (Cockell et al., 2012).
Water on present-day Mars
After the Noachian and throughout the Hesperian, the planet's declining heat flow is hypothesized to have led to freezing of most of the water in the surface environment and at gradually increasing depth (Clifford, 1987; Clifford and Parker, 2001). Mars today hosts a large body of frozen water. The near surface (to a depth of tens of centimeters) of Mars is thought to harbor water ice deposits that vary from 2% wt at the equator to pure ice at the polar regions (Christensen, 2006; Bandfield, 2007; Feldman et al., 2011) mixed with surface volcanic regolith as determined by the Mars Odyssey Gamma-Ray Spectrometer (GRS) and Neutron Spectrometer (MONS). Near-surface ice was directly observed at the Phoenix landing site (Smith et al., 2009).
Estimates have put the cryosphere depth at ∼2.5 km at the equator to ∼6.5 km at the poles (Fanale, 1976; Clifford, 1993). However, downward revisions of the geothermal gradient of Mars suggest that the depth could be up to 2–3 times greater (Clifford et al., 2010). Salt solutions would lessen these depths by depressing the freezing point. For example, perchlorate at high concentrations would depress the freezing point to 203 K, thinning the cryosphere (Clifford et al., 2010). The martian cryosphere is estimated to contain an equivalent global layer of water of ∼35 m (Christensen, 2006).
Direct observations of the present-day martian deep subsurface, up to several kilometers depth, were made with the Mars Advanced Radar for Subsurface and Ionospheric Sounding (MARSIS) aboard the Mars Express spacecraft (Picardi et al., 2005), which has a theoretical penetration depth of ∼5 km. The instrument provided information on the subsurface of the north polar layered deposits on Mars down to ∼1.8 km, confirming that the deposits are mainly composed of pure water ice.
The Shallow Radar (SHARAD) instrument on MRO has similarly provided direct information on conditions in the subsurface and has a penetration depth of ∼1.5 km. The study of subsurface structures in lobate debris aprons, which are broad, lobate features that extend up to 20 km away from steep slopes in equatorial regions of Mars, suggests that there are buried glaciers (Holt et al., 2008). The data from the SHARAD instrument suggest that up to ∼28,000 km3 of water ice might be sequestered in lobate debris aprons in the Hellas Basin region of Mars alone, equivalent to a global water layer ∼20 cm thick. The authors attribute their finding to the large obliquity (axial tilt) changes that Mars experiences. Models predict that, when obliquity was ∼45° and in the southern summer solstice, large amounts of water vapor would be formed. The vapor would be transported northward and deposited as snow by condensation and precipitation. Eventually, it was covered by debris. Today, these buried glaciers are testament to climatic changes on Mars over timescales of several million years (Head et al., 2003).
In general, the water ice on Mars, however, follows its predicted depth of stability under current climatic conditions (Bandfield, 2007), consistent with the idea that the subsurface ice conditions of Mars follow orbitally driven climate cycles, with local heterogeneities reflecting differences in topography and material type and preservation of icy deposits from previous epochs.
The presence of ice in the subsurface of Mars is confirmed by impact cratering. Layered ejecta blankets around craters—indicative of high volatile (water) content—can be used to estimate the depth at which subsurface water ice exists based on known relationships between crater diameters and excavation depth (Urbach and Stepinski, 2009; Barlow, 2010).
Quite apart from massive ice deposits at the poles, with an estimated volume of 1.2–1.7×106 km3 (Zuber et al., 1998), and buried ice just discussed, there is a large literature on other glacial and periglacial features on Mars. Evidence for subsurface ice includes inferred features such as polygonal structures, gullies, deformation features in putative permafrost terrain, ice-sublimation–related features, and parallel sorted stone stripes, among others (Squyres and Carr, 1986; Seibert and Kargel, 2001; Mangold, 2003, 2005; Mangold et al., 2004; Levy et al., 2010). Pingos, which are produced by liquid water injection into the subsurface with subsequent freezing that causes upheaval, have been suggested (Mangold, 2005; Dundas and McEwen, 2010; Soare et al., 2013). They are of particular interest, as their formation mechanism requires bulk liquid water movement. Observations from orbit include recently excavated small craters that reveal water ice. The ice is observed to sublimate away after several months (Bryne et al., 2009).
Liquid water at the surface of Mars today is rendered unstable, partly because much of the surface is at the triple point and partly because the low humidity means that, when liquid water is formed, it will rapidly evaporate, even if it does not boil (Haberle et al., 2001). However, evidence has been presented for near-surface present-day liquid water. Gullies that have characteristic alcoves located on a steep slope with an incised sinuous channel leading down to an apron of deposited material have been proposed as evidence of present-day liquid water (Malin and Edgett, 2000; Heldmann and Mellon, 2004; Goldspiel and Squyres, 2011). The observation of some of these features high up on impact crater walls and hills is difficult to reconcile with plausible subsurface water sources. It is hypothesized that gullies could be formed by two different processes: CO2-induced processes such as the fluidization of the regolith and subsurface water (Dienega et al., 2010; Reiss et al., 2010; Schon and Head, 2011).
Seasonally recurrent dark slope streaks (McEwen et al., 2011) could be present-day surface expressions of near-surface salty water. Droplets of putative salt solutions on the legs of the Phoenix lander provide evidence for physical and thermodynamic stability of brines on Mars (Rennó et al., 2009), and deliquescence provides one mechanism by which these brines might form (Martínez and Rennó, 2013). Perchlorate solutions are likely to be metastable on Mars (Gough et al., 2011).
On present-day Mars, it is reasonable to hypothesize that groundwater could exist deep underground where radiogenic heating and lithostatic pressures would allow liquid water to exist above the freezing point. The presence of salts, which depress the freezing point, would reduce the depth at which these waters were plausible (Clifford et al., 2010). Impact events would be another mechanism by which the present-day cryosphere could be melted and disrupted to create a link between the subsurface and surface (Schwenzer et al., 2012). Radar searches for putative present-day bodies of liquid water have not been successful (Nunes et al., 2010). A number of factors, including a very dry conductive surface, have limited radar measurements in most areas of Mars to less than 100 m. The low surface roughness and well-saturated crustal porosity required to optimize radar penetration depth and signal recovery are met only in a small proportion (<20%) of the planet.
Liquid water could exist today in the form of thin water films on soil grains. Dielectric measurements of soils during the nighttime at the Phoenix landing site suggest the present of liquid water, but the lack of conductivity suggests that the water does not move (Stoker et al., 2010), with important implications for its connectivity to other environments. Substantial water quantities have been observed in the <150 μm fraction martian fines by the Curiosity rover (Leshin et al., 2013: Meslin et al., 2013). Thin films of interfacial liquid water on soil grains of just a few nanometers thickness have been proposed as possible microhabitats (Möhlmann, 2009). Water is kept in the liquid phase on the surface of grains by Van der Waals forces above a threshold temperature. They are thought to be able to exist as no more than two monolayers at temperatures down to 163 K within the top 20 cm in the martian subsurface (Möhlmann, 2009, 2011; Martínez and Rennó, 2013). Whether microorganisms can access thin interfacial layers of water or use it as a solvent, since it is tightly bound to the grains, is not known. However, a serious limitation of this water is that, if it remains static (as is suggested for the Phoenix lander site), the microhabitat it creates will geochemically run down and become depleted in essential nutrients, which makes this water a poor environment for the long-term activity of life. Even if it is transiently habitable and Mars was inhabited, the lack of connectivity to other environments would render such static water an uninhabited habitat.
In the more recent geological history of Mars, bulk liquid water might have become available as a result of warming during higher obliquity. This is possibly the case for ices at the Phoenix landing site (Stoker et al., 2010), where, during the last 5 million years, obliquity increases up to 50° would generate surface temperatures in excess of 273 K up to 100 days a year. Ulrich et al. (2012) investigated features of the Utopia Planitia and suggested that during the last 10 million years, thaw processes would have generated liquid water, which would have contributed to geomorphological features in the region and made liquid water available to any putative life. High-obliquity periods between 7.864 and 7.855 Ma and four periods of duration 100–12,000 years between 9.76 and 9.45 Ma could have represented times when liquid water was stable in surface or near-surface environments. Jakosky et al. (2003) examined the conditions required to melt martian polar ice in the past and concluded that, during periods of higher obliquity (>40°), average temperatures could have generated conditions for the formation of interstitial liquid water. The significance of these latter papers is that they suggest the existence of isolated transient liquid water environments that are candidates for uninhabited habitats.
Basic elements
Life requires six basic elements to construct macromolecules (C, H, N, O, P, S). Carbon atoms are likely to have been, and continue to be, present in the surface and subsurface of Mars as a consequence of atmospheric exchange (present day 95.32% CO2; 800 ppm CO) and could be acquired by life through autotrophy. The detection of carbonates (Ehlmann et al., 2008; Michalski and Niles, 2010) suggests that aqueous interactions with these rocks could have generated a source of inorganic autotrophically available carbon throughout martian history. Ancient reservoirs of stored carbon dioxide produced during a time when the martian atmosphere was thicker are an additional plausible source (Kurahashi-Nakamura and Tajika, 2006). The concentration of organic carbon on Mars, a potential source of carbon for heterotrophs, which has been detected in surface missions, is controversial (Leshin et al., 2013; Ming et al., 2014 and citations therein). The infall of carbonaceous chondrites and other organic carbon-bearing material is expected (Benner et al., 2000). Organics are likely to be destroyed by reactive oxygen species, UV radiation, and ionizing radiation in the near-surface environment (e.g., Benner et al., 2000; Kminek and Bada, 2006; Dartnell et al., 2007; Davila et al., 2008; Noblet et al., 2012; Pavlov et al., 2012; Ming et al., 2014), and these factors will greatly influence its preservation and concentrations in different regions and depths on Mars. Reduced magmatic carbon in martian basalts (Steele et al., 2012) might be another source of organic carbon.
Hydrogen atoms are available from water throughout the martian depth profile, which could be split radiolytically in the subsurface (Lin et al., 2005). Hydrogen could also be generated in chemical reactions. The presence of serpentine in impact crater uplifts (Ehlmann et al., 2010, 2011; Quantin et al., 2012) suggests the possibility of hydrogen production through serpentinization reactions, particularly when water flow through ultramafic rocks was more extensive in the Noachian.
Nitrogen gas is present in the modern atmosphere at 2.7%. Fixed nitrogen compounds have been reported in martian meteorites (Wright et al.,
1992; Grady et al.,
1997) and confirmed on the surface of Mars (Ming et al.,
2014). They have been predicted to include nitrate and ammonium based on terrestrial analogues (Mancinelli and Banin, 2003). To be used in biological systems, nitrogen must be in a fixed form. One potential pathway is biological fixation, which was shown to be possible at a pN2 of 5 mbar, but not below 1 mbar, suggesting that this pathway was plausible in a denser early martian atmosphere but unlikely today (Klingler et al.,
1989). Nitrogen fixation on Mars could occur by abiotic processes, including impact events, lightening, and volcanic activity (Segura and Navarro-González, 2005; Summers and Khare, 2007; Manning et al.,
2009), or by processes analogous to reduction by hydrogen in deep subsurface systems on Earth (Brandes et al.,
1998). The concentrations reached and the depths achieved by nitrogen fixed in such processes throughout martian history are unknown. Boxe et al. (2012) used a one-dimensional model to show that fixed nitrogen species, some produced photochemically, for example, 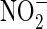 , NO, HNO3, could be generated on the surface of Mars and then transported into near-surface environments in thin water films. Similarly, NO and other abiotically fixed species have been suggested as nitrogen sources and biological electron sinks on early Earth (Ducluzeau et al.,
2008). This transient photochemically produced nitrogen cycle on Mars could provide a source of fixed nitrogen species today, but the depth of its penetration would be low because of lack of surface liquid water. Without a continuous flow of fixed nitrogen into the deep subsurface of Mars, particularly following the cessation of widespread surface hydrological activity on Mars in the Noachian, nitrogen might be, and might have been, one of the limiting factors for life. Despite the detection of fixed nitrogen in meteorites and directly on the surface of Mars, determining the distribution and form of fixed nitrogen in the martian crust, past and present, remains one of the most important challenges in constraining martian habitability (Mancinelli and Banin, 2003).
, NO, HNO3, could be generated on the surface of Mars and then transported into near-surface environments in thin water films. Similarly, NO and other abiotically fixed species have been suggested as nitrogen sources and biological electron sinks on early Earth (Ducluzeau et al.,
2008). This transient photochemically produced nitrogen cycle on Mars could provide a source of fixed nitrogen species today, but the depth of its penetration would be low because of lack of surface liquid water. Without a continuous flow of fixed nitrogen into the deep subsurface of Mars, particularly following the cessation of widespread surface hydrological activity on Mars in the Noachian, nitrogen might be, and might have been, one of the limiting factors for life. Despite the detection of fixed nitrogen in meteorites and directly on the surface of Mars, determining the distribution and form of fixed nitrogen in the martian crust, past and present, remains one of the most important challenges in constraining martian habitability (Mancinelli and Banin, 2003).
Oxygen atoms could be provided by CO2, H2O, sulfates, perchlorates, ferric oxides, and reactive oxygen species. Oxygen atoms are bound to many of the biologically accessible compounds cited and discussed here in association with other elements (C, H, N, P, S).
Phosphate has been reported in martian meteorites (Boctor et al., 1998) and on the surface of Mars in a number of missions. For example, Mössbauer, MiniTES, and APXS spectra from the Mars Exploration Rovers are interpreted to suggest apatite concentrations (wt %) at between 0.1% and 2.4% (McGlynn et al., 2012). Rocks with P2O5 abundances (Wishstone Class) of over 5% were observed in Gusev Crater by the Mars Exploration Rovers in which the primary phosphate-bearing mineral may be merrillite (Usui et al., 2008). Phosphorus was also observed in alkaline basalts studied in Gale Crater at <1% weight abundances (Stolper et al., 2013).
Sulfur has been detected on Mars in meteorites and on the surface in the form of sulfate salts including gypsum, ferric sulfates, jarosite, and other S-bearing species in different oxidation states, including sulfides (Scott, 1999; Gendrin et al., 2005; Langevin et al., 2005; Morris et al., 2006; Bibring et al., 2007; Gaillard et al., 2013; McLennan et al., 2014; Ming et al., 2014). Jarosite-bearing deposits may attest to the oxidation of martian pyrites (Zolotov and Shock, 2005). The extent of these compounds in the subsurface is not known, but the dominance of the sulfur cycle on Mars (reviewed by Gaillard et al., 2013) suggests that sulfur species would have been distributed from the mantle to the surface throughout martian history, potentially including sulfur in microbially accessible gaseous phases such as H2S and SO2.
Other micronutrients and trace elements
The presence of widespread ultramafic and basaltic rocks on Mars and their alteration products shows that a range of major and trace elements required by life are available (Brückner et al., 2003; Gellert et al., 2006), as is the case for igneous rocks on Earth (Taylor and McLennan, 2009). Fe is abundant in the ferrous state in olivines and in the ferric state in a variety of materials from clays to ferric oxides (crystalline and amorphous) and sulfates measured both from orbit and on the surface (Bibring et al., 2007; Bish et al., 2013; Blake et al., 2013; Ehlmann et al., 2013; Vaniman et al., 2014). Mg and Ca are present in materials such as clays and pyroxenes, and K and Na in materials such as muscovite, illite, and plagioclases such as K-feldspar (Bish et al., 2013; Ehlmann et al., 2013; McLennan et al., 2013; Stolper et al., 2013). As would be expected for igneous rocks, other biological trace elements including Mn, Cr, Ni, Zn have been observed (e.g., McLennan et al., 2013; Meslin et al., 2013; Stolper et al., 2013). There is no major or trace element used by biota on Earth and accessible from igneous rocks that is obviously lacking in martian rocks.
There is likely to be a depth and spatial dependence in which the form of these elements is found. In the Noachian deep subsurface (and possibly in the deep subsurface today), in confined closed aqueous systems, water may have been heavily enriched in Ca, Mg, and Fe in ultramafic ultrabasic environments (Michalski et al., 2013). In the near-surface environments, both in the present day and the past, many of these elements would be partitioned in brines, oxides, sulfates, and a variety of minerals (Tosca et al., 2005, 2008b; Bibring et al., 2007; Gaillard et al., 2013), with implications for the coexistence of biologically required suites of elements in any given location at microscopic scales.
Energy and redox couples
A variety of microbial energy sources can be assessed for Mars (Table 1). Photosynthesis is a plausible mode of metabolism when surface water is available (Sagan and Pollack, 1974; Cockell and Raven, 2004). There are depths on the order of millimeters or less in the near surface where the UV biologically effective irradiances are no worse than on Earth today but where photosynthetically active radiation is sufficient for phototrophy (Sagan and Pollack, 1974; Cockell and Raven, 2004), such as anoxygenic photosynthesis that uses ferrous iron or reduced sulfur species. The lack of liquid water on the surface today precludes a productive surface photosynthetic biosphere. As for subsurface life on Earth, photosynthesis is eliminated at depth.
Table 1.
Examples of Potential Redox Couples for Life on Past and Present-Day Mars
| Electron donor | Electron acceptor | Name | Comment |
|---|---|---|---|
| Photosynthesis | |||
| Fe2+ | Photoferrotrophy | Depends on clement surface conditions. Unlikely since the Noachian. | |
| S/S2- | Anoxygenic sulfur photosynthesis | Depends on clement surface conditions. Unlikely since the Noachian. Sulfide suggested in Curiosity data (Ming et al.,2014). Sulfur suggested at Gusev Crater (Morris et al.,2007). | |
| H2O | Oxygenic photosynthesis | Cannot be discounted on early Mars, but no atmospheric evidence for this reaction on present-day Mars. | |
| Chemolithotrophy | |||
| Fe2+ | 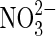 |
Anaerobic iron oxidation | Distribution of 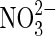 on Mars not known although fixed nitrogen is inferred (Ming et al.,2014). on Mars not known although fixed nitrogen is inferred (Ming et al.,2014). |
| Fe2+ | perchlorates | Anaerobic iron oxidation | Perchlorate can be used to oxidize iron but is not shown to be used for growth in organisms. It is included to highlight the need for investigation of perchlorate-containing redox couples. |
| H2 | CO2 | Methanogenesis, acetogenesis | Hydrogen inferred from presence of olivine and serpentine—substrates and products for H2-evolving water-rock reactions. |
| H2 | Fe3+ | Iron reduction | As above for hydrogen. |
| H2 | 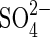 |
Sulfate reduction | As above for hydrogen. |
| H2 | oxidized nitrogen species | Distribution of oxidized nitrogen species on Mars not known. | |
| S | 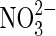 |
Sulfur oxidation | Sulfur suggested at Gusev Crater (Morris et al.,2007). |
| S | Fe3+ | Anaerobic sulfur oxidation | Occurs in acidic conditions. |
| CO | 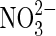 |
Anaerobic carboxydotrophy | Carbon monoxide in atmosphere. |
| Chemoorganotrophy | |||
| organics | Fe3+ | Iron reduction | The accessibility and state of organics on Mars is not known, but they are expected to arrive within carbonaceous chondrites. |
| organics | 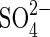 |
Sulfate reduction | As above for organics. |
| organics | 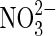 |
Nitrate reduction | As above for organics. |
| organics | perchlorate | Perchlorate reduction | As above for organics. |
| Fermentation (disproportionation) | |||
| organics | organics | Fermentation | As above for organics. |
Red denotes a half-reaction that is known to exist or has existed on Mars, green a half-reaction for which there is strong reason to suspect its presence (meteoritic organics and hydrogen). Reactions are selected from anoxic redox couples. The use of O2 as an electron acceptor for microaerophilic reactions such as hydrogen oxidation on past or present-day Mars is not explicitly ruled out. The table does not include many redox reactions that use different oxidation states of nitrogen (e.g., anaerobic ammonium oxidation with nitrite), since the fixed state of nitrogen in the martian crust is not known. Note that redox couples involving the oxidation and reduction of iron can be performed with other variable valence cations (e.g., Mn, U) that could be present in varying oxidation states in the martian crust.
Chemolithotrophic redox couples are an alternative energy source (Jepsen et al., 2007). Grotzinger et al. (2014) suggested that the Yellowknife Bay site at Gale Crater, Mars, could have supported chemolithoautotrophy. Ferric and sulfate ions as electron acceptors, both of which have been detected on Mars, can be reduced with hydrogen (suggested from the presence of olivine, serpentine, and other substrates or products of hydrogen-evolving mineral weathering). H2 production from serpentinization reactions has been shown to occur in spinel-containing peridotite, olivine, and pyroxene at temperatures of 55°C and 100°C (Mayhew et al., 2013), and it can be produced by anaerobic photochemical oxidation of siderite (Kim et al., 2013), although this latter pathway would be confined to the surface. On Earth, hydrogen can act as the electron donor in the subsurface for microbial redox reactions with sulfate (Matias et al., 2005; Moser et al., 2005; Harris et al., 2007) and ferric iron (Lovley, 1995; Harris et al., 2007). However, as yet the presence of hydrogen on past or present Mars has not been directly demonstrated.
Large resources of ferrous-bearing minerals such as olivines are available for chemolithotrophic iron oxidation. However, Mars probably lacks suitable electron acceptors. Oxygen in the atmosphere (0.14%) is at insufficient concentrations today for aerobic iron oxidation, although localized oxygen concentrations on Mars, produced by abiotic pathways during an early oxidized past (Tuff et al., 2013), are not ruled out. Anaerobic ferrous iron oxidation linked to nitrate (Straub et al., 1996) is difficult to assess, as there is no direct detection of nitrate. Although perchlorates can be used as an electron acceptor for iron oxidation, to date this couple has not been shown to conserve energy for growth (Coates and Achenbach, 2004).
The presence of reduced sulfur species such as sulfides, which have been found in martian meteorites (Scott, 1999) and on the surface of Mars (Leshin et al., 2013; Ming et al., 2014), suggests the possibility of sulfur species oxidation (Grotzinger et al., 2014). However, anaerobic conditions prevent chemolithotrophic sulfur species oxidation with oxygen as the terminal electron acceptor. Sulfur can be oxidized with the use of ferric iron as the electron acceptor (Pronk et al., 1992). This reaction occurs in acidic conditions, and elemental sulfur has been tentatively identified on Mars (Morris et al., 2007). Liquid water on the surface in the Noachian would have allowed for more favorable conditions for sulfur anoxygenic photosynthesis. This would require the co-location of reduced sulfur species, light, and other requirements for habitability at microbial scales.
Other chemolithotrophic redox couples could include methanogenesis and acetogenesis, both with the use of CO2 from the atmosphere or from dissolved carbonates and H2 from serpentinization reactions as observed in the subsurface of Earth (Kotelnikova and Pedersen, 1998; Moser et al., 2005; Harris et al., 2007). Methane itself can be oxidized by microorganisms as a source of energy and can be produced abiotically (Berndt et al., 1996). Serpentinized ultramafic rocks are known to host thriving microbial communities in the subsurface of Earth and in surface discharge (Blank et al., 2009; Okland et al., 2012; Szponar et al., 2013) and could provide analogies to potential water-rock-microbial interactions for the martian subsurface.
Although biogenic methanogenesis cannot be ruled out in the present day, to date a surface detection of methane that would be consistent with such a hypothesis remains elusive (Webster et al., 2013). The presence of CO in the atmosphere, which can be used as an electron donor in anaerobic carboxydotrophy, has also been suggested to indicate the lack of a significant biological sink (Weiss et al., 2000a). Although martian sources and sinks of CH4 and CO are not fully understood, the data do not at the current time provide any evidence for a present-day active chemolithotrophic biosphere on Mars.
Chemoorganotrophy could provide energy for growth, past and present. Iron and sulfate reduction can be accomplished with organic electron donors. Nixon et al. (2012) discussed the available electron acceptors for iron reduction, and by extension sulfate reduction, and conclude that a range of meteoritic organics could be plausible electron donors for iron reduction in the surface and near-surface of Mars (organics could also theoretically provide a substrate for fermentation, a disproportionation reaction), although the biological availability of organics at sufficient concentrations has not been directly demonstrated, and they would be limited by the infall rate.
Finally, there is a wide diversity of other, more unusual redox couples involving alternative anions or cations including arsenite oxidation, uranium reduction, and many others that can be used by microorganisms to conserve energy for growth. The study of more geochemically heterogeneous sites on Mars and a greater investigation of localized distributions of a variety of anions and cations will allow alternative redox couples to be assessed.
The accessibility of theoretically available energy for life would have tracked liquid water availability. As much of the surface water disappeared in the late Noachian, photosynthesis as a theoretical metabolism would have been eliminated. As a greater proportion of the surface and near-surface environment became desiccated and frozen, near-surface exogenous organics from meteoritic material would have become unavailable as a source of energy, until the theoretical energy sources were restricted primarily to chemolithotrophic deep subsurface redox couples and chemoorganotrophy, where organics could be supplied from dead organisms (necromass).
In conclusion, although there are potentially a wide variety of energy sources, a current assessment of energy availability on Mars, past and present, based only on unambiguous detection of half-reactions (Table 1), and not potential electron donors or acceptors, shows that almost all microbial metabolisms lack definitive detection of a complete redox couple, which leads to the conclusion that many martian environments were and are extremely energy-limited. Redox reactions for which available electron donors have been definitely detected tend to lack definitively detected electron acceptors. Those with confirmed electron acceptors lack definitive electron donors. This conclusion of extreme energy limitation can only be removed with the direct detection and determination of the concentration and biological accessibility of a greater variety of electron donors and acceptors in recent and ancient terrains.
Radiation
Ultraviolet radiation is rapidly attenuated in the martian subsurface, so although the surface flux includes wavelength components down to 200 nm (Sagan and Pollack, 1974; Cockell et al., 2000; Rontó et al., 2003; Patel et al., 2004; Schuerger et al., 2006), which generates biologically effective DNA damage about three orders of magnitude higher than on the surface of Earth (Cockell et al., 2000) within a depth of a few tens of microns to millimeters, depending on soil particle size, UV radiation is extinguished (Mancinelli and Klovstad, 2000; Cockell and Raven, 2004; Moores et al., 2007). In present-day desiccated surface environments, it contributes to uninhabitable conditions, but in the presence of liquid water and various protection strategies, including physical and biological (Cockell et al., 2000; Cockell and Raven, 2004), it does not in itself cause uninhabitable conditions.
Ionizing radiations of solar energetic particles and galactic cosmic rays are more penetrating. The total dose of ionizing radiation experienced on the martian surface has been measured as 76 mGy/year (Hassler et al., 2014), which is much lower than the fluxes that can be tolerated by radioresistant organisms such as Deinococcus radiodurans, which can withstand doses in excess of 5 kGy without appreciable loss of viability (Battista, 1997; Dartnell et al., 2007). However, inactivity would result in accumulated damage such that, at 2 m depth in the martian crust, a D. radiodurans population has been estimated to suffer an approximately 6-order-of-magnitude reduction in viability after 450,000 years. For a deep subsurface biota at just a few meters depth or greater on Mars, particularly one that is active and can repair damage in an environment where liquid water is available, radiation would not render the subsurface uninhabitable (Dartnell et al., 2007, 2012; Hassler et al., 2014).
pH
In a variety of martian settings, pH ranges are within the boundaries for life. The pH of the martian near subsurface was measured at the Phoenix lander site. It was found to be slightly alkaline, 7.7–7.9, and carbonate-buffered (Hecht et al., 2009); and the pH at Yellowknife Bay, Gale Crater, was also inferred to be neutral (Grotzinger et al., 2014). This pH range is benign for organisms. Although we have no direct measurements of pH in the deep subsurface, past or present, reactions of fluids with mafic and ultramafic rocks control solution chemistry. It would be expected that, as is the case on Earth, buffered fluids would be anoxic and alkaline or ultrabasic (pH>10) (Okland et al., 2012; Szponar et al., 2013).
Although many environments in the ancient history of Mars may have been neutral to alkaline, the presence of sulfate minerals on the surface of Mars, particularly jarosite and iron sulfates, suggests the potential for locally acidic conditions. Sulfates are found as Hesperian layered sulfates, polar deposits, interior layered deposits, sediments in craters, within the globally ubiquitous martian dust (which may contain 5–10% sulfates), and as sulfate veins within rocks (Clark and Baird, 1979; Clark et al., 2005; Langevin et al., 2005; Bibring et al., 2006; King and McLennan, 2010; Grindrod et al., 2012; Squyres et al., 2012). Mg- and Fe-bearing sulfates are generally more common than Ca-bearing sulfates (Gaillard et al., 2013).
Some of these minerals suggest a period of acidic weathering (pH 2–5) in the Hesperian, during which exhalations of SO2 from martian volcanic activity would have produced acidic conditions, which then weathered martian basalts to produce secondary sulfate minerals in low water-rock ratio interactions (Hurowitz and McLennan, 2007). These sulfate salts are also testament to the fact that the surface and subsurface geochemical cycles of Mars have been strongly influenced by the sulfur cycle, as compared to circumstances on Earth, where the carbon cycle generally dominates (Gaillard et al., 2013). Acidic environments on Earth are not uninhabitable, but they do restrict the range of organisms capable of active growth (Baker-Austin and Dopson, 2007). The restriction of water availability and the presence of acidic conditions during the Hesperian would attest to more widespread inclement environments in the surface and possibility in the subsurface. However, the lowest pHs predicted for Mars would not in themselves make environments uninhabitable. Local differences in pH would merely change the suitability of putative habitats for particular organisms.
Brines
Brines can constrain the boundaries of active life by influencing water activity and other parameters such as chaotropicity (the degree of disorder induced in macromolecules). Extremely low water activities and high chaotropicities can be generated by brines such as chlorides (CaCl2) and mixed sulfate brines (Tosca et al., 2005, 2008a, 2008b; Hallsworth et al., 2007).
The lack of chlorides or sulfates associated with Noachian clays (Ehlmann et al., 2011) suggests that, although deep subsurface ancient brines cannot be excluded, the water was likely not composed of extremely concentrated brines that would have produced deleterious water activities. However, on present-day Mars, seasonally recurrent dark slope streaks (McEwen et al., 2011) could be formed from concentrated briny solutions. Some martian brines are calculated to have water activities below those required for life and would not be habitable environments (Tosca et al., 2008a). Thus, as the hydrological environment of Mars transitioned from the Noachian into the Hesperian and salt-saturated solutions became prevalent in groundwater environments, some of these briny solutions could have rendered localized environments uninhabitable.
Porosity and physical conditions in the subsurface
The porosity of the subsurface of Mars may be greater than that of Earth at comparable depths and lithologies because of the lower gravity (0.38 Earth gravity) on the planet (Clifford et al., 2010). As on Earth, porosity in any location will be controlled by factors including secondary mineral infilling, sediment deposition, local rock pressure environments, and other geological processes, but fundamentally there is no reason why the martian subsurface rock environment should not be accessible to life (McMahon et al., 2013). The permeability of subsurface environments will affect the connectivity of environments with implications for the abundance of uninhabited and inhabited habitats in habitability trajectories that assume an inhabited Mars.
We do not have direct measurements of temperature and pressure profiles into the deep subsurface of Mars. As for the Earth, deep habitability is likely to be constrained when the geothermal gradient temperature exceeds the upper temperature limit for life (122°C; Takai et al., 2008). Geothermal gradients of between ∼10°C/km and ∼20°C/km (Solomon and Head, 1990; Michalski et al., 2013) imply habitable temperature ranges on the order of ∼6–15 km depth. Lithostatic pressures are unlikely to render environments uninhabitable. As the pressure is proportional to the gravity, pressures in the subsurface zone where temperatures are below the upper temperature limit for life will be no higher than those experienced in terrestrial deep subsurface settings.
Trajectories of Martian Habitability
Supported by this previous synthesis of environmental conditions that would have influenced conditions available to support the activity of organisms, it is possible to construct a series of habitability trajectories that are consistent with these data (Fig. 2).
All trajectories of martian habitability begin with the formation of Mars. From early planetesimals (Debaille et al., 2007), an uninhabitable planet formed. As water condensed and the environment cooled, the planet was at a branch point in its long-term trajectory of biological conditions. In one set of trajectories, the planet is defined by its condition as uninhabited (neither an origin of life occurs nor does life transfer to the planet from Earth in meteoritic matter). In the second set of trajectories, the planet is defined by the establishment of life, an event that changes the use of habitable conditions and through feedback effects would itself change the habitability of environments (Nisbet et al., 2007).
Quantifying the relative abundance of environments that make up the trajectories described here will be technically enormously challenging. A comprehensive analysis and characterization of martian habitability will require deep drilling in many locations to examine the environments of the martian deep subsurface (e.g., Smith and McKay, 2005; Zacny et al., 2008; McKay et al., 2013) as well as extensive studies of many surface and near-surface environments. This will allow for one of these trajectories to be identified and the relative abundance of its component environments to be determined.
Trajectories of habitability on an uninhabited Mars
It is not known exactly the conditions that are required to transform prebiotic chemicals into self-replicating life-forms. It is possible to consider various environments and conditions in which an origin of life might have occurred and how prevalent these might have been on early Mars (Stephenson et al., 2013; Westall et al., 2013). However, at the current time it is not possible to quantify the probability of life's origin on Mars, and we do not know whether it did originate.
Although the survival of microorganisms in impact shock (Burchell et al., 2001, 2004; Horneck et al., 2001), in space (Horneck et al., 1994), and potentially in atmospheric transit (Fajardo-Cavazos et al., 2005) might suggest the possibility of the transfer of life from Earth to Mars in its early history (Mileikowsky et al., 2000) in material known to have been transferred (Gladman et al., 1996; Weiss et al., 2000b), there is no empirical evidence to show that this process has occurred.
These factors, taken together, mean that an uninhabited Mars remains a plausible condition for the planet throughout its history and must be taken as one early branch point in systematically identifying habitability trajectories. Three trajectories of martian habitability are envisaged for an uninhabited Mars.
Trajectory 1. Mars is and was always uninhabitable.
This trajectory posits a Mars where the only environment type that has existed (Fig. 1) is uninhabitable environments at both macroscopic and microscopic scales. There are several ways in which this condition might be realized.
One scenario is a lack of spatially contemporaneous requirements for life in any given location. If all the requirements for life are met (Grotzinger et al., 2014) but never all together in one place at the scale of microorganisms (micron to submicron scales), then environments would be uninhabitable. This seems less plausible for early Mars compared to the present day, since water flow might be expected to solubilize many elements and nutrients and generate environments in which a diversity of chemical species coexist at small spatial scales, including the vital elements required for life. As conditions deteriorated during the Hesperian and sustained hydrological processes were terminated, a greater number of conditions that lead to uninhabitable environments, for example, lack of liquid water, low water activity in extreme brines, acidity, would have been realized and co-localized in a greater number of environments, particularly on the surface. In combination, they could have ensured that many environments remained uninhabitable.
Mars might have always lacked a fundamental requirement for life at sufficient concentrations. Although fixed nitrogen has been detected in martian meteorites (Wright et al., 1992; Grady et al., 1997) and on the surface of Mars (Grotzinger et al., 2014; Ming et al., 2014), if these species had not been produced at sufficient concentrations over large scales or co-localized at micron scales with other requirements for life, then this element could have been limiting to life (Mancinelli and Banin, 2003).
Yet another scenario by which this trajectory might be realized is if all environments contained physical or chemical conditions that lie outside habitable conditions, for example, brines with a water activity too low for life (Tosca et al., 2008a). Although some brines on Mars might be, and might have been, too extreme for life, these environments are clearly not globally encompassing (Stoker et al., 2010; Grotzinger et al., 2014).
Trajectory 1 can be experimentally tested by demonstrating that in the most biologically promising aqueous environments on past and present Mars, the environments either lacked or lack a requirement for all known organisms or had or have physicochemical conditions that lie outside the physical and chemical tolerances of all known organisms. This experimental test is underpinned by the hypothesis “All environments on Mars were and are uninhabitable.”
Recent studies by the Curiosity rover do not suggest globally uninhabitable conditions. Grotzinger et al. (2014) suggested a clement early Hesperian aqueous environment at Yellowknife Bay, Gale Crater, with the presence of essential elements and diverse redox states of elements that could have supported life. If this conclusion is correct, then it may already be possible to discount Trajectory 1. However, the trajectory is discussed here for completeness.
Trajectory 2. Uninhabited Mars has hosted uninhabited habitats transiently or continuously during its history.
This trajectory encompasses an uninhabited Mars characterized by two environments throughout its history—uninhabited habitats and uninhabitable environments. During the Noachian, when liquid water was more widespread, uninhabited habitats could have been abundant. The discovery of evidence for clement conditions in Gale Crater by way of the Curiosity rover (Grotzinger et al., 2014), but no evidence for an organic signature associated with life (Ming et al., 2014), could constitute evidence for an ancient uninhabited habitat.
This trajectory supposes that the circulation of ancient water on Mars generated environments where dissolved elements and gases provided vital elements (CHNOPS), nutrients, and energetic redox couples co-located at microscopic scales with clement pH conditions and water activities.
As surface conditions deteriorated during the Hesperian, uninhabited habitats would become more confined and localized as an increasing number of surface environments became uninhabitable at all scales on account of desiccation, long-term irradiation, acidity, the presence of low–water activity brines, and other environmentally deleterious conditions.
Up until the present day, uninhabited habitats become confined to subsurface and near-surface environments where liquid water became transiently available, for example, in impact-induced melting of permafrost at macroscopic scales (Cockell et al., 2012; Fig. 6), layers of water within soil grains at microscopic scales, and production of local liquid water during obliquity changes (Ulrich et al., 2012). They could exist if the geothermal gradient allows for deep aquifers or if habitable liquid water is transiently sustained in near-surface environments such as dilute brines (Clifford et al., 2010; McEwen et al., 2011).
FIG. 6.

An uninhabited habitat on Mars. A scenario for the formation of an uninhabited habitat on Mars by an impact into permafrost, which remains hydrologically isolated at macroscopic and microscopic scales even on a planet that has a hypothetically colonized subsurface (from Cockell et al., 2012).
Thus, this trajectory is characterized by a changing relative abundance of uninhabited habitats and uninhabitable environments through time, with uninhabited habitats becoming less abundant. The trajectory assumes that locations on Mars have harbored habitable conditions throughout its history.
This trajectory includes a scenario where Mars has been uninhabitable but punctuated by infrequent and transient uninhabited habitats through time. For example, uninhabited habitats could be transiently generated by impact events into permafrost (Cockell et al., 2012; Schwenzer et al., 2012) or even destroyed by episodic periods of inclement climatic conditions (Head et al., 2003; Schulze-Makuch et al., 2013), causing temporally variable occurrences of these habitats.
Trajectory 2 can be tested by demonstrating that, throughout martian history, uninhabited habitats have existed at different times. These environments must have no evidence for life, although they could possess organics associated with meteoritic infall or indigenous reduced carbon; but they must have or have had a set of elemental availabilities, appropriate physical and chemical conditions, a potential energy supply, and liquid water that could sustain the activity of at least one known organism. For any given martian environment, this latter requirement can be tested by comparing the combined set of physical and chemical data from a set of martian samples for any given location with what we know about the requirements and limits of different microorganisms on Earth. This experimental test is underpinned by the hypothesis “Mars has always been uninhabited but has possessed uninhabited habitats transiently or continuously throughout its history.”
Trajectory 3. Uninhabited Mars was habitable and possessed uninhabited habitats but is now uninhabitable.
Trajectory 3 is a more extreme version of Trajectory 2. In this scenario, uninhabited habitats existed during the early history of Mars, but deteriorating conditions eventually led to their constriction and localization until they completely disappeared, even at microscopic scales, rendering the entire planet uninhabitable, a condition that remains to the present day. A hypothetically plausible scenario would be the desiccation of the surface, the freezing of the subsurface as the cryosphere extended in depth (Clifford et al., 2010), and, even if deep aquifers persisted to the present day, a geochemical rundown of the availability of nutrients caused by insufficient turnover (Nisbet et al., 2007). It would also require that the formation of transient liquid water in recent times occurred in locations where conditions rendered it uninhabitable, for example, where brine concentrations and water activities of aqueous solutions were extreme (Tosca et al., 2008a) or where there was insufficient energy, nutrients, and other basic elements co-localized at the scale of microorganisms.
Thus, this trajectory posits a well-defined biphasic history for Mars where it was once habitable but is now uninhabitable. This distinctive change is the reason for separating it into a separate trajectory.
Trajectory 3 can be tested by demonstrating that during early martian history environments existed that possessed the conditions required for at least one known organism to be active, but there is no evidence that these environments contained life. Furthermore, more recent environments must have conditions that do not meet requirements for the activity of any known microorganism and suggest a sustained period of globally uninhabitable conditions. This experimental test is underpinned by the hypothesis “Mars has always been uninhabited. However, although it once possessed uninhabited habitats, it is now uninhabitable.”
Trajectories for an inhabited Mars
A second set of trajectories is realized if Mars became inhabited, either by an indigenous origin of life or a transfer of life from Earth to Mars. This event would add inhabited habitats as an environment type (Fig. 1). There are a variety of possible evolutionary trajectories for life if Mars was inhabited that are linked to scenarios for changing environmental conditions and known microbial metabolisms (Schulze-Makuch et al., 2005) and potential analogues on Earth (Fairén et al., 2010). The potential types of life found in the surface and subsurface would be strongly influenced by specific environmental conditions such as the presence of salts (Davila et al., 2010) and redox couples (Fisk and Giovannoni, 1999). In turn, the different plausible microbial metabolisms would influence the potential planetary-scale productivity (Jakosky and Shock, 1998). Three plausible trajectories can be identified for an inhabited Mars.
Trajectory 4. Mars is and was inhabited.
Trajectory 4 posits a sequence where all three environments existed and continue to exist on Mars—uninhabitable environments (in places such as extreme desiccated surface environments), uninhabited habitats (newly formed habitats disconnected from inhabited regions, even if only transiently), and inhabited habitats that contain martian life.
Similarly to other trajectories, the geological evidence observed on Mars suggests that the relative abundance of these three environments would have changed over time. As surface conditions deteriorated and liquid water ceased to be as abundant (Lasue et al., 2013), one would expect the diversity and area of inhabited habitats to decrease from the Noachian through to the Amazonian. As surface environments became desiccated and more acidic, and evaporative brines more extreme, the area of surface uninhabitable environments would have increased through time. The diversity and area of uninhabited habitats would also have concomitantly changed. As the hydrological scale reduced in scope, more habitats would have become isolated and separated from inhabited habitats, potentially increasing the relative volume of uninhabited habitats; but counter to this, generally deteriorating conditions across the whole planet might have acted to reduce the number of habitable spaces overall.
Trajectory 4 can be experimentally demonstrated by showing that fossilized life exists in materials from ancient Mars and the planet also harbors extant active life. This experimental test is underpinned by one of astrobiology's most compelling hypotheses “Life existed and exists on Mars.”
Trajectory 4 is the trajectory that Earth has taken. Habitats on Earth are mostly inhabited with rare uninhabited habitats.
Trajectory 5. Mars was inhabited, life became extinct, but uninhabited habitats remain on Mars.
Trajectory 5 posits the existence of life and all three environments (Fig. 1) on early Mars; but as hydrological conditions deteriorated and geochemical turnover became less efficient, life was eventually constrained to such small pockets of existence that it became functionally extinct, and eventually a total extinction occurred. At this point Mars transitioned into a planet harboring only uninhabitable and uninhabited habitats. The extinction event would not have precluded new habitable places becoming available, for example, from obliquity-driven liquid water formation or in impact-induced hydrothermal systems (Cockell et al., 2012; Fig. 6); but a lack of connectivity and sufficient water flow prevented their colonization from the last remaining vestiges of life until eventually, when life became extinct, there was no life to occupy uninhabited habitats that persist, or are transiently produced, to this day.
Trajectory 5 can be experimentally demonstrated by showing that fossilized life exists in materials from ancient Mars but that the planet today hosts uninhabited habitats. This experimental test is underpinned by the hypothesis “Life existed on Mars, and although locations on Mars are habitable today, there is no life to occupy them.”
Trajectory 6. Mars was inhabited, life became extinct, and the planet became uninhabitable.
Trajectory 6 posits the existence of life and all three environments (Fig. 1) on early Mars, as in Trajectory 5; but as hydrological conditions deteriorated and geochemical turnover became less efficient, life was eventually constrained to such small pockets of existence that it became functionally extinct. Eventually, a total extinction occurred, constituting a distinctive branch point in its biological history. At this point, Mars transitioned into a planet harboring only uninhabitable environments, as even newly formed isolated uninhabited habitats were extinguished. This transition could have occurred through a phase where isolated uninhabited habitats existed for a period of time; that is, Mars hosted inhabited and uninhabited habitats until conditions became so extreme that even uninhabited habitats ceased to exist. This trajectory, as in Trajectories 1 and 3, requires that the combined environmental, chemical, and physical conditions on Mars eventually placed all environments outside the capabilities of all known microorganisms to sustain activity.
Trajectory 6 can be experimentally demonstrated by showing that fossilized life exists in materials from ancient Mars, that there is no evidence for extant life on Mars, and that even the most promising present-day aqueous environments on Mars are uninhabitable. This experimental test is underpinned by the hypothesis “Life existed on Mars but no longer exists, and the planet is uninhabitable.”
Other trajectories
There are other trajectories that can be suggested, but the geological evidence on Mars reviewed in the previous sections makes them less likely than those outlined above. Nevertheless, for completeness, two of them are mentioned here.
Trajectory 7 is a scenario in which uninhabitable environments during much of martian early history in the Noachian ultimately transitioned to uninhabited habitats on an uninhabited Mars at some later stage during the Hesperian/Amazonian. This trajectory, which supposes a planetary-scale improvement in the conditions for life, is difficult to reconcile with the known hydrological and geological evidence, since Noachian Mars had more abundant liquid water than Hesperian and Amazonian Mars (Lasue et al., 2013).
Trajectory 8 is a scenario where Mars, either uninhabitable or possessing uninhabited habitats throughout its early history, transitioned at some later point to an inhabited world in a distinctive biphasic history. This seems unlikely for the same reasons given for Trajectory 7. Such a trajectory could occur if Mars was covered in uninhabited habitats for a significant period of time and was then inoculated by the interplanetary transfer of life from Earth or if it was host to an indigenous origin of life at a late stage of its history. Such a scenario would require an extremely low probability of the transfer of life between planets or an origin of life, allowing for Mars to become inhabited at a late stage.
There are other trajectories of greater complexity that can be envisaged. Examples include an inhabited Mars on which life becomes extinct and then reoriginates (or is transferred from Earth) at some later time. Combinations of the major trajectories discussed here are possible. However, from a position of parsimony, this synthesis is focused on the six major trajectories that can be envisaged based on what we currently know about the history of the martian environment.
Conclusions
The habitability of Mars and its biological condition have been of long-term interest to scientists. A three-environment system can be used to identify six major plausible trajectories for the habitability of Mars through time. The clarification of these trajectories allows for a systematic identification of experimental requirements for demonstrating each trajectory and its corresponding testable hypothesis. Such trajectories have implications for the interpretation of data returned from missions to Mars that investigate habitability on the planet. With sufficient research and missions, it is possible to determine which of these proposed trajectories the planet Mars took and quantify the relative abundance of its component environments. This approach to identifying habitability trajectories can be applied to other planetary bodies.
Acknowledgments
This work was made possible with support from the UK Science and Technology Facilities Council (STFC), Grant No. ST/1001964/1. I thank two anonymous reviewers for their helpful comments.
Abbreviations
CRISM, Compact Reconnaissance Spectrometer for Mars; HiRISE, High Resolution Science Experiment; MRO, Mars Reconnaissance Orbiter; SHARAD, Shallow Radar.
References
- Arvidson R.E., Poulet F., Morris R.V., Bibring J.-P., Bell J.F., Squyres S.W., Christensen P.R., Bellucci G., Gondet B., Ehlmann B.L., Farrand W.H., Fergason R.L., Golombek M., Griffes J.L., Grotzinger M., Guiness E.A., Herkenhoff K.E., Johnson J.R., Klingelhöfer G., Langevin Y., Ming D., Seelos K., Sullivan R.J., Ward J.G., Wiseman S.M., and Wolff M. (2006) Nature and origin of the hematite-bearing plains of Terra Meridiani based on analyses of orbital and Mars Exploration rover data sets. J Geophys Res 111, 10.1029/2006JE002728 [DOI] [Google Scholar]
- Baker-Austin C. and Dopson M. (2007) Life in acid: pH homeostasis in acidophiles. Trends Microbiol 15:165–171 [DOI] [PubMed] [Google Scholar]
- Bandfield J.L. (2007) High-resolution subsurface water-ice distributions on Mars. Nature 447:64–68 [DOI] [PubMed] [Google Scholar]
- Barlow N.G. (2010) What we know about Mars from its impact craters. GSA Bulletin 122:644–657 [Google Scholar]
- Battista J.R. (1997) Against all odds: the survival strategies of Deinococcus radiodurans. Annu Rev Microbiol 51:203–224 [DOI] [PubMed] [Google Scholar]
- Benner S.A., Devine K.G., Matveeva L.N., and Powell D.H. (2000) The missing organic molecules on Mars. Proc Natl Acad Sci USA 97:2425–2430 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berndt M.E., Allen D.E., and Seyfried W.E. (1996) Reduction of CO2 during serpentinization of olivine at 300°C and 500 bar. Geology 24:351–354 [Google Scholar]
- Bibring J.-P., Langevin Y., Mustard J.F., Poulet F., Arvidson R., Gendrin A., Gondet B., Mangold N., Pinet P., Forget F., and the OMEGA Team (2006) Global mineralogical and aqueous Mars history derived from OMEGA/Mars Express data. Science 312:400–404 [DOI] [PubMed] [Google Scholar]
- Bibring J.-P., Arvidson R.E., Gendrin A., Gondet B., Langevin Y., Le Mouelic S., Mangold N., Morris R.V., Mustard J.F., Poulet F., Quantin C., and Sotin C. (2007) Coupled ferric oxides and sulfates on the martian surface. Science 317:1206–1210 [DOI] [PubMed] [Google Scholar]
- Bish D.L., Blake D.F., Vaniman D.T., Chipera S.J., Morris R.V., Ming D.W., Treiman A.H., Sarrazin P., Morrison S.M., Downs R.T., Achilles C.N., Yen A.S., Bristow T.F., Crisp J.A., Morookian J.M., Farmer J.D., Rampe E.B., Stolper E.M., Spanovich N., and the MSL Science Team (2013) X-ray diffraction results from Mars Science Laboratory: mineralogy of Rocknest at Gale Crater. Science 341, 10.1126/science.1238932 [DOI] [PubMed] [Google Scholar]
- Blake D.F., Morris R.V., Kocurek G., Morrison S.M., Downs R.T., Bish D., Ming D.W., Edgett K.S., Rubin D., Goetz W., Madsen M.B., Sullivan R., Gellert R., Campbell I., Treiman A.H., McLennan S.M., Yen A.S., Grotzinger J., Vaniman D.T., Chipera S.J., Achilles C.N., Rampe E.B., Sumner D., Meslin P.-Y., Maurice S., Forni O., Gasnault O., Fisk M., Schmidt M., Mahaffy P., Leshin L.A., Glavin D., Steele A., Freissinet C., Navarro-González R., Yingst R.A., Kah L.C., Bridges N., Lewis K.W., Bristow T.F., Farmer J.D., Crisp J.A., Stolper E.M., Des Marais D.J., Sarrazin P., and the MSL Science Team (2013) Curiosity at Gale Crater, Mars: characterization and analysis of the Rocknest Sand Shadow. Science 341, 10.1126/science.1239505 [DOI] [PubMed] [Google Scholar]
- Blank J.G., Green S., Blake D., Valley J.W., Kita N.T., Treiman A., and Dobson P.F. (2009) An alkaline spring system within the Del Puerto Ophiolite (California, USA): a Mars analog site. Planet Space Sci 57:533–540 [Google Scholar]
- Boctor N.Z., Wang J., Alexander C.M., Hauri E., Bertka C.M., and Fei Y. (1998) Hydrogen isotopic studies of carbonate and phosphate in martian meteorite ALH 84001. Meteorit Planet Sci 32:A18 [Google Scholar]
- Boston P.J., Ivanov M.V., and McKay C.P. (1992) On the possibility of chemosynthetic ecosystems in subsurface habitats on Mars. Icarus 95:300–308 [DOI] [PubMed] [Google Scholar]
- Boxe C.S., Hand K.P., Nealson K.H., Ying Y.L., and Saiz-Lopez A. (2012) An active nitrogen cycle on Mars sufficient to support a subsurface biosphere. International Journal of Astrobiology 11:109–115 [Google Scholar]
- Brandes J.A., Boctor N.Z., Cody G.D., Cooper B.A., Hazen R.M., and Yoder H.S. (1998) Abiotic nitrogen reduction on the early Earth. Nature 395:365–367 [DOI] [PubMed] [Google Scholar]
- Brückner J., Dreibus G., Rieder R., and Wänke H. (2003) Refined data of Alpha Proton X-ray Spectrometer analyses of soils and rocks at the Mars Pathfinder site: implications for surface chemistry. J Geophys Res 108, 10.1029/2003JE002060 [DOI] [Google Scholar]
- Bryne S., Dundas C.M., Kennedy M.R., Mellon M.T., McEwen A.S., Cull S.C., Daubar I.J., Shean D.E., Seelos K.D., Murchie S.L., Cantor B.A., Arvidson R.E., Edgett K.S., Reufer A., Thomas N., Harrison T.N., Posiolova L.V., and Seelos F.P. (2009) Distribution of mid-latitude ground ice on Mars from new impact craters. Science 325:1674–1676 [DOI] [PubMed] [Google Scholar]
- Burchell M.J., Mann J.R., Bunch A.W., and Brando P.F.B. (2001) Survivability of bacteria in hypervelocity impact. Icarus 154:545–547 [Google Scholar]
- Burchell M.J., Mann J.R., and Bunch A.W. (2004) Survival of bacteria and spores under extreme shock pressures. Mon Not R Astron Soc 352:1273–1278 [Google Scholar]
- Burr D.M., Grier J.A., McEwen A.S., and Keszthelyi L.P. (2002) Repeated aqueous flooding from the Cerberus Fossae: evidence for very recently extant, deep groundwater on Mars. Icarus 159:53–73 [Google Scholar]
- Cabrol N.A. and Grin E.A. (2001) The evolution of lacustrine environments on Mars: is Mars only hydrologically dormant? Icarus 149:291–328 [Google Scholar]
- Cabrol N.A. and Grin E.A. (2002) Overview on the formation of paleolakes and ponds in impact craters on Mars. Glob Planet Change 35:199–219 [Google Scholar]
- Cabrol N.A., Grin E.A., Newsom H.E., Landheim R., and McKay C.P. (1999) Hydrogeologic evolution of Gale Crater and its relevance to the exobiological exploration of Mars. Icarus 139:235–245 [Google Scholar]
- Carr M.H. (1986) Mars—a water-rich planet? Icarus 68:187–216 [Google Scholar]
- Carr M.H. (1996) Water on Mars, Oxford University Press, Oxford, UK [Google Scholar]
- Changela H. and Bridges J.C. (2010) Alteration assemblages in the nakhlites: variation with depth on Mars. Meteorit Planet Sci 45:1847–1867 [Google Scholar]
- Christensen P.R. (2006) Water at the poles and in permafrost regions of Mars. Elements 2:151–155 [Google Scholar]
- Clark B.C. and Baird A.K. (1979) Is the martian lithosphere sulphur rich? J Geophys Res 84:8395–8403 [Google Scholar]
- Clark B.C., Morris R.V., McLennan S.M., Gellert R., Jolliff B., Knoll A.H., Squyres S.W., Lowenstein T.K., Ming D.W., Tosca N.J., Yen A., Christensen P.R., Gorevan S., Brückner J., Calvin W., Dreibus G., Farrand W., Klingelhoefer G., Waenke H., Zipfel J., Bell J.F., Grotzinger J., McSween H.Y., and Rieder R. (2005) Chemistry and mineralogy of outcrops at Meridiani Planum. Earth Planet Sci Lett 240:73–94 [Google Scholar]
- Clifford S.M. (1987) Polar basal melting on Mars. J Geophys Res 92:9135–9152 [Google Scholar]
- Clifford S.M. (1993) A model for the hydrologic and climatic behaviour of water on Mars. J Geophys Res 98:10973–11016 [Google Scholar]
- Clifford S.M. and Parker T.J. (2001) The evolution of the martian hydrosphere: implications for the fate of a primordial ocean and the current state of the Northern Plains. Icarus 154:40–79 [Google Scholar]
- Clifford S.M., Lasue J., Heggy E., Boisson J., McGovern P., and Max M.D. (2010) Depth of the martian cryosphere: revised estimates and implications for the existence and detection of subpermafrost water. J Geophys Res 115, 10.1029/2009JE003462 [DOI] [Google Scholar]
- Coates J.D. and Achenbach L.A. (2004) Microbial perchlorate reduction: rocket-fuelled metabolism. Nature 2:569–580 [DOI] [PubMed] [Google Scholar]
- Cockell C.S. (2011) Vacant habitats in the Universe. Trends Ecol Evol 26:73–80 [DOI] [PubMed] [Google Scholar]
- Cockell C.S. and Raven J.A. (2004) Zones of photosynthetic potential on Mars and the early Earth. Icarus 169:300–310 [Google Scholar]
- Cockell C.S., Catling D., Davis W.L., Kepner R.N., Lee P.C., Snook K., and McKay C.P. (2000) The ultraviolet environment of Mars: biological implications past, present and future. Icarus 146:343–359 [DOI] [PubMed] [Google Scholar]
- Cockell C.S., Balme M., Bridges J.C., Davila A., and Schwenzer S.P. (2012) Uninhabited habitats on Mars. Icarus 217:184–193 [Google Scholar]
- Cousins C.R., Crawford I.A., Carrivick J.L., Gunn M., Harris J., Kee T.P., Karlsson M., Carmody L., Cockell C.S., Herschy B., and Joy K.H. (2013) Glaciovolcanic hydrothermal environments in Iceland and implications for their detection on Mars. Journal of Volcanology and Geothermal Research 256:61–77 [Google Scholar]
- Dartnell L.R., Desorgher L., Ward J., and Coates A. (2007) Modelling the surface and subsurface martian radiation environment: implications for astrobiology. Geophys Res Lett 34:L02207 [Google Scholar]
- Dartnell L.R., Page K., Jorge-Villar S.E., Wright G., Munshi T., Scowen I.J., Ward J.M., and Edwards H.G.M. (2012) Destruction of Raman biosignatures by ionising radiation and the implications for life detection on Mars. Anal Bioanal Chem 403:131–144 [DOI] [PubMed] [Google Scholar]
- Davila A.F., Fairén A.G., Gago-Duport L., Stoker C., Amils R., Bonaccorsi R., Zavaleta J., Lim D., Schulze-Makuch D., and McKay C.P. (2008) Subsurface formation of oxidants on Mars and implications for the preservation of organic biosignatures. Earth Planet Sci Lett 272:456–463 [Google Scholar]
- Davila A.F., Duport L.G., Melchiorri R., Jänchen J., Valea S., de los Rios A., Fairén A.G., Möhlmann D., McKay C.P., Ascaso C., and Wierzchos J. (2010) Hygroscopic salts and the potential for life on Mars. Astrobiology 10:617–628 [DOI] [PubMed] [Google Scholar]
- Debaille V., Brandon A.D., Yin Q.Z., and Jacobsen B. (2007) Coupled 142Nd–143Nd evidence for a protracted magma ocean in Mars. Nature 450:525–528 [DOI] [PubMed] [Google Scholar]
- Dienega S., Bryne S., Bridges N.T., Dundas C.M., and McEwen A.S. (2010) Seasonality of present-day martian dune-gully activity. Geology 38:1047–1050 [Google Scholar]
- Ducluzeau A.-L., van Lis R., Duval S., Schoepp-Cothenet B., Russell M.J., and Nitschke W. (2008) Was nitric oxide the first deep electron sink? Trends Biochem Sci 34:9–15 [DOI] [PubMed] [Google Scholar]
- Dundas C.M. and McEwen A.S. (2010) An assessment of evidence for pingos on Mars using HiRISE. Icarus 205:244–258 [Google Scholar]
- Ehlmann B.L., Mustard J.F., Murchie S.L., Poulet F., Bishop J.L., Brown A.J., Calvin W.M., Clark R.N., Des Marais D.J., Milliken R.E., Roach L.H., Roush T.L., Swayze G.A., and Wray J.J. (2008) Orbital detection of carbonate-bearing rocks on Mars. Science 322:1828–1832 [DOI] [PubMed] [Google Scholar]
- Ehlmann B.L., Mustard J.F., Swayze G.A., Clark R.N., Bishop J.L., Poulet F., Des Marais D.J., Roach L.H., Milliken R.E., Wray J.J., Barnouin-Jha O., and Murchie S.L. (2009) Identification of hydrated silicate minerals on Mars using MRO-CRISM: geologic context near Nili Fossae and implications for aqueous alteration. J Geophys Res 144, 10.1029/2009JE003339 [DOI] [Google Scholar]
- Ehlmann B.L., Mustard J.F., and Murchie S.L. (2010) Geologic setting of serpentine deposits on Mars. Geophys Res Lett 37:L06201 [Google Scholar]
- Ehlmann B.L., Mustard J.F., Murchie S.L., Bibring J.-P., Meunier A., Fraeman A.A., and Langevin Y. (2011) Subsurface water and clay mineral formation during the early history of Mars. Nature 479:53–60 [DOI] [PubMed] [Google Scholar]
- Ehlmann B.L., Berger G., Mangold N., Michalski J.R., Catling D.C., Ruff S.W., Chassefière E., Niles P.B., Chevrier V., and Poulet F. (2013) Geochemical consequences of widespread clay mineral formation in Mars' ancient crust. Space Sci Rev 174:329–364 [Google Scholar]
- Fairén A.G., Dohm J.M., Uceda E.R., Rodriguez A.P., Baker V.R., Fernandez-Remolar D., Schulze-Makuch D., and Amils R. (2005) Prime candidate sites for astrobiological exploration through the hydrogeological history of Mars. Planet Space Sci 53:1355–1375 [Google Scholar]
- Fairén A.G., Davila A.F., Lim D., Bramall N., Bonaccorsi R., Zavaleta J., Uceda E.R., Stoker C., Wierzchos J., Dohm J.M., Amils R., Andersen D., and McKay C.P. (2010) Astrobiology through the ages of Mars: the study of terrestrial analogues to understand the habitability of Mars. Astrobiology 10:821–843 [DOI] [PubMed] [Google Scholar]
- Fajardo-Cavazos P., Link L., Melosh H.J., and Nicholson W.L. (2005) Bacillus subtilis spores on artificial meteorites survive hypervelocity atmospheric entry: implications for lithopanspermia. Astrobiology 5:726–736 [DOI] [PubMed] [Google Scholar]
- Fanale F.P. (1976) Martian volatiles: their degassing history and geochemical fate. Icarus 28:179–202 [Google Scholar]
- Feldman W.C., Pathare S., Maurice T.H., Prettyman T.H., Lawrence D.J., Milliken R., and Travis B.J. (2011) Mars Odyssey Neutron data: 2. Search for buried excess water ice deposits at non-polar latitudes on Mars. J Geophys Res 116, 10.1029/2011JE003806 [DOI] [Google Scholar]
- Fisk M.R. and Giovannoni S.J. (1999) Sources of nutrients and energy for a deep biosphere on Mars. J Geophys Res 104:11805–11815 [Google Scholar]
- Fridlund M. (2008) Future space missions to search for terrestrial planets. Space Sci Rev 135:355–369 [Google Scholar]
- Gaillard F., Michalski J., Berger G., McLennan S.M., and Scaillet B. (2013) Geochemical reservoirs and timing of sulphur cycling on Mars. Space Sci Rev 174:251–300 [Google Scholar]
- Gellert R., Rieder R., Brückner J., Clark B.C., Dreibus G., and Klingelhöfer G. (2006) Alpha Particle X-ray Spectrometer (APXS) results from Gusev Crater and calibration report. J Geophys Res 111, 10.1029/2005JE002555 [DOI] [Google Scholar]
- Gendrin A., Mangold N., Bibring J.-P., Langevin Y., Gondet B., Poulet F., Bonello G., Quantin C., Mustard J., Arvidson R., and LeMouélic S. (2005) Sulfates in martian layered terrains: the OMEGA/Mars Express view. Science 307:1587–1591 [DOI] [PubMed] [Google Scholar]
- Gladman B.J., Burns J.A., Duncan M., Lee P., and Levison H.F. (1996) The exchange of impact ejecta between the terrestrial planets. Science 271:1387–1392 [Google Scholar]
- Goldspiel J.M. and Squyres S.W. (2011) Groundwater discharge and gully formation on martian slopes. Icarus 211:238–258 [Google Scholar]
- Gough R.V., Chevrier V.F., Baustian K.J., Wise M.E., and Tolbert M.A. (2011) Laboratory studies of perchlorate phase transitions: support for metastable aqueous perchlorate solutions on Mars. Earth Planet Sci Lett 312:371–377 [Google Scholar]
- Grady M.M., Wright I.P., and Pillinger C.T. (1997) A carbon and nitrogen study of Zagami. J Geophys Res 102:9165–9173 [Google Scholar]
- Grindrod P.M., West M., Warner N.H., and Gupta S. (2012) Formation of an Hesperian-aged sedimentary basin containing phyllosilicates and sulfates in Coprates Catena, Mars. Icarus 218:178–195 [Google Scholar]
- Grotzinger J.P., Arvidson R.E., Calvin W., Clark B.C., Fike D.A., Golombek M., Greeley R., Haldemann A., Herkenhoff E., Joliff B.L., Knoll A.H., Malin M., McLennan S.M., Parker T., Soderblom L., Sohl-Dickstein J.N., Squyres S.W., Tosca N.J., and Watters W. (2005) Stratigraphy and sedimentology of a dry to wet eolian depositional system, Burns Formation, Meridiani Planum, Mars. Earth Planet Sci Lett 240:11–72 [Google Scholar]
- Grotzinger J.P., Sumner D.Y., Kah L.C., Stack K., Gupta S., Edgar L., Rubin D., Lewis K., Schieber J., Mangold N., Milliken R., Conrad P.G., Des Marais D., Farmer J., Siebach K., Calef F., Hurowitz J., McLennan S.M., Ming D., Vaniman D., Crisp J., Vasavada A., Edgett K.S., Malin M., Blake D., Geliert R., Mahaffy P., Wiens R.C., Maurice S., Grant J.A., Wilson S., Anderson R.C., Beegle L., Arvidson R., Hallet B., Sletten R.S., Rice M., Bell J., Griffes J., Ehlmann B., Anderson R.B., Bristow T.F., Dietrich W.E., Dromart G., Eigenbrode J., Fraeman A., Hardgrove C., Herkenhoff K., Jandura L., Kocurek G., Lee S., Leshin L.A., Leveille R., Limonadi D., Maki J., McCloskey S., Meyer M., Minitti M., Newsom H., Oehler D., Okon A., Palucis M., Parker T., Rowland S., Schmidt M., Squyres S., Steele A., Stolper E., Summons R., Treiman A., Williams R., Yingst A., and the MSL Science Team (2014) A habitable fluvio-lacustrine environment at Yellowknife Bay, Gale Crater, Mars. Science 343 10.1126/science.1242777 [DOI] [PubMed] [Google Scholar]
- Haberle R.M., McKay C.P., Schaeffe J., Cabrol N.A., Grin E.A., Zent A.P., and Quinn R. (2001) On the possibility of liquid water on present-day Mars. J Geophys Res 106, 10.1029/2000JE001360 [DOI] [Google Scholar]
- Hallsworth J.E., Yakimov M.M., Golyshin P.N., Gillion J.L.M., D'Auria G., De Lima Alves F., La Cono V., Genovese M., McKew B.A., Hayes S.L., Harris G., Giulioano L., Timmis K.N., and McGenity T. (2007) Limits of life in MgCl2-containing environments: chaotropicity defines the window. Environ Microbiol 9:801–813 [DOI] [PubMed] [Google Scholar]
- Harris S.H., Smith R.L., and Suflita J.M. (2007) In situ hydrogen consumption kinetics as an indicator of subsurface microbial activity. FEMS Microbiol Ecol 60:220–228 [DOI] [PubMed] [Google Scholar]
- Hartmann W. and Neukum G. (2001) Cratering chronology and the evolution of Mars. Space Sci Rev 96:165–194 [Google Scholar]
- Hassler D.M., Zeitlin C., Wimmer-Schweingruber R.F., Ehresmann B., Rafkin S., Eigenbrode J.L., Brinza D.E., Weigle G., Böttcher S., Böhm E., Burmeister S., Guo J., Köhler J., Martin C., Reitz G., Cucinotta A., Myung-Hee K., Grinspoon D., Bullock M.A., Posner A., Gómez-Elvira J., Vasavada A., Grotzinger J.P., and the MSL Science Team (2014) Mars' surface radiation environment measured with the Mars Science Laboratory's Curiosity Rover. Science 343 10.1126/science.1244797 [DOI] [PubMed] [Google Scholar]
- Head J.W., Hiesinger H., Ivanov M.A., Kreslavsky M.A., Pratt S., and Thomson B.J. (1999) Possible ancient oceans on Mars: evidence from Mars Orbiter Laser Altimeter data. Science 296:2134–2137 [DOI] [PubMed] [Google Scholar]
- Head J.W., Mustard J.F., Kreslavsky M.A., Milliken R.E., and Marchant D.R. (2003) Recent ice ages on Mars. Nature 426:797–802 [DOI] [PubMed] [Google Scholar]
- Hecht M.H., Kounaves S.P., Quinn R.C., West S.J., Young S.M., Ming D.W., Catling D.C., Clark B.C., Boynton W.V., Hoffman J., Deflores L.P., Gospodinova K., Kapit J., and Smith P.H. (2009) Detection of perchlorate and the soluble chemistry of martian soil at the Phoenix lander site. Science 324:64–67 [DOI] [PubMed] [Google Scholar]
- Heldmann J.L. and Mellon M.T. (2004) Observations on martian gullies and constraints on potential formation mechanisms. Icarus 168:285–304 [Google Scholar]
- Hoehler T.M. (2007) An energy balance concept for habitability. Astrobiology 7:824–838 [DOI] [PubMed] [Google Scholar]
- Holt J.W., Safaeinili A., Plaut J.J., Head J.W., Phillips R.J., Seu R., Kempf S.D., Choudhary P., Young D.A., Putzig N.E., Biccari D., and Gim Y. (2008) Radar sounding evidence for buried glaciers in the southern mid-latitudes of Mars. Science 322:1235–1238 [DOI] [PubMed] [Google Scholar]
- Horneck G., Bücker H., and Reitz G. (1994) Long-term survival of bacterial spores in space. Adv Space Res 14:41–45 [DOI] [PubMed] [Google Scholar]
- Horneck G., Rettberg P., Reitz G., Wehner J., Eschweiler U., Strauch K., Panitz C., Starke V., and Baumstark-Khan C. (2001) Protection of bacterial spores in space, a contribution to the discussion on panspermia. Orig Life Evol Biosph 31:527–547 [DOI] [PubMed] [Google Scholar]
- Hurowitz J.A. and McLennan S.M. (2007) A ∼3.5 Ga record of water-limited, acidic weathering conditions on Mars. Earth Planet Sci Lett 260:432–443 [Google Scholar]
- Jakosky B.M. and Shock E.L. (1998) The biological potential of Mars, the early Earth and Europa. J Geophys Res 103:19359–19364 [DOI] [PubMed] [Google Scholar]
- Jakosky B.M., Nealson K.H., Bakermans C., Ley R.E., and Mellon M.T. (2003) Subfreezing activity of microorganisms and the potential habitability of Mars' polar regions. Astrobiology 3:343–350 [DOI] [PubMed] [Google Scholar]
- Javaux E.J. and Dehant V. (2010) Habitability: from stars to cells. Astronomy and Astrophysics Review 18:383–416 [Google Scholar]
- Jepsen S.M., Priscu J.C., Grimm R.E., and Bullock M.A. (2007) The potential for lithautotrophic life on Mars: application to shallow interfacial water environments. Astrobiology 7:342–354 [DOI] [PubMed] [Google Scholar]
- Kim J.D., Yee N., Nanda V., and Falkowski P.G. (2013) Anoxic photochemical oxidation of siderite generates molecular hydrogen and iron oxides. Proc Natl Acad Sci USA 110:10073–10077 [DOI] [PMC free article] [PubMed] [Google Scholar]
- King P.L. and McLennan S.M. (2010) Sulfur on Mars. Elements 6:107–112 [Google Scholar]
- Klingler J.M., Mancinelli R.L., and White M.R. (1989) Biological nitrogen fixation under primordial martian partial pressures of dinitrogen. Adv Space Res 9:173–176 [DOI] [PubMed] [Google Scholar]
- Kminek G. and Bada J.L. (2006) The effect of ionizing radiation on the preservation of amino acids on Mars. Earth Planet Sci Lett 245:1–5 [Google Scholar]
- Kotelnikova S. and Pedersen K. (1998) Distribution and activity of methanogens and homoacetogens in deep granitic aquifers at Aspö Hard Rock Laboratory, Sweden. FEMS Microbiol Ecol 26:121–134 [Google Scholar]
- Kurahashi-Nakamura T. and Tajika E. (2006) Atmospheric collapse and transport of carbon dioxide into the subsurface on early Mars. Geophys Res Lett 33, 10.1029/2006GL027170 [DOI] [Google Scholar]
- Langevin Y., Poulet F., Bibring J.-P., and Gondet B. (2005) Sulfates in the north polar region of Mars detected by OMEGA/Mars Express. Science 307:1584–1586 [DOI] [PubMed] [Google Scholar]
- Lasue J., Mangold N., Hauber E., Clifford S.M., Feldman W., Gasnault O., Grima C., Maurice S. and Mousis O. (2013) Quantitative assessments of the martian hydrosphere. Space Sci Rev 174:155–212 [Google Scholar]
- Leshin L.A., Mahaffy P.R., Webster C.R., Cabane M., Coll P., Conrad P.G., Archer P.D., Jr., Atreya S.K., Brunner A.E., Buch A., Eigenbrode J.L., Flesch G.J., Franz H.B., Freissinet C., Glavin D.P., McAdam A.C., Miller K.E., Ming D.W., Morris R.V., Navarro-González R., Niles P.B., Owen T., Pepin R.O., Squyres S., Steele A., Stern J.C., Summons R.E., Sumner D.Y., Sutter B., Szopa C., Teinturier S., Trainer M.G, Wray J.J., Grotzinger J.P., and the MSL Science Team (2013) Volatile, isotope, and organic analysis of martian fines with the Mars Curiosity rover. Science 341, 10.1126/science.1238937 [DOI] [PubMed] [Google Scholar]
- Levy J.S., Head J.W., and Marchant D.R. (2010) Concentric crater fill in the northern mid-latitudes of Mars: formation processes and relationships to similar landforms of glacial origin. Icarus 209:390–404 [Google Scholar]
- Lin L.H., Hall J., Lippmann-Pipke J., Ward J.A., Sherwood-Lollar B., De Flaun M., Rothmel R., Moser D., Gihring T.M., Mislowack B., and Onstott T.C. (2005) Radiolytic H2 in continental crust: nuclear power for deep subsurface microbial communities. Geochemistry, Geophysics, Geosystems 6, 10.1029/2004GC000907 [DOI] [Google Scholar]
- Lovley D.R. (1995) Dissimilatory Fe(III) and Mn(IV) reduction. Microbiol Mol Biol Rev 55:259–287 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malin M.C. and Edgett K.S. (2000) Evidence for recent groundwater seepage and run-off on Mars. Science 288:2330–2335 [DOI] [PubMed] [Google Scholar]
- Mancinelli R.L. and Banin A. (2003) Where is the nitrogen on Mars? International Journal of Astrobiology 2:217–225 [Google Scholar]
- Mancinelli R.L. and Klovstad M. (2000) Martian soil and UV radiation: microbial viability assessment on spacecraft surfaces. Planet Space Sci 48:1093–1097 [Google Scholar]
- Mangold N. (2003) Geomorphic analysis of lobate debris aprons on Mars at MOC scale: evidence for ice sublimation initiated by fractures. J Geophys Res 108, 10.1029/2002JE01885 [DOI] [Google Scholar]
- Mangold N. (2005) High latitude patterned ground on Mars: classification, distribution and climatic control. Icarus 174:336–359 [Google Scholar]
- Mangold N., Maurice S., Feldman W.C., Costard F., and Forget F. (2004) Spatial relationships between patterned ground and ground ice detected by the Neutron Spectrometer on Mars. J Geophys Res 109, 10.1029/2004JE002235 [DOI] [Google Scholar]
- Manning C.V., Zahnle K.J., and McKay C.P. (2009) Impact processing of nitrogen on early Mars. Icarus 199:273–285 [Google Scholar]
- Martínez G.M. and Rennó N.O. (2013) Water and brines on Mars: current evidence and implications for MSL. Space Sci Rev 175:29–51 [Google Scholar]
- Matias P.M., Pereira I.A.C., Soares C.M., and Carrondo M.A. (2005) Sulfate respiration from hydrogen in Desulfovibrio bacteria: a structural biology overview. Prog Biophys Mol Biol 89:292–32915950057 [Google Scholar]
- Mayhew L.E., Ellison E.T., McCollom T.M., Trainor T.P., and Templeton A.S. (2013) Hydrogen generation from low-temperature water-rock interactions. Nat Geosci 6:478–484 [Google Scholar]
- McEwen A.S., Ojha L., Dundas C.M., Mattson S.S., Bryne S., Wray J.J., Cull S.C., Murchie S.L., Thomas N., and Gulick V.C. (2011) Seasonal flows on warm martian slopes. Science 333:740–743 [DOI] [PubMed] [Google Scholar]
- McGlynn I.O., Fedo C.M., and McSween H.Y. (2012) Soil mineralogy at the Mars Exploration Rover landing sites: an assessment of the competing roles of physical sorting and chemical weathering. J Geophys Res 117, 10.1029/2011JE003861 [DOI] [Google Scholar]
- McKay C.P. and Davis W.L. (1991) Duration of liquid water habitats on early Mars. Icarus 90:214–221 [DOI] [PubMed] [Google Scholar]
- McKay C.P., Stoker C.R. and Glass B.J. (2013) The Icebreaker life mission to Mars: a search for biomolecular evidence of life. Astrobiology 13:334–353 [DOI] [PubMed] [Google Scholar]
- McLennan S.M. and Grotzinger J.P. (2008) The sedimentary rock cycle of Mars. In The Martian Surface: Composition, Mineralogy, and Physical Properties, edited by Bell J.F., III, Cambridge University Press, Cambridge, UK, pp 541–577 [Google Scholar]
- McLennan S.M., Anderson R.B., Bell J.F., Bridges J.C., Calef F., Campbell J.L., Clark B.C., Clegg S., Conrad P., Cousin A., Des Marais D.J., Dromart G., Dyar M.D., Edgar L.A., Ehlmann B.L., Fabre C., Forni O., Gasnault O., Gellert R., Gordon S., Grant J.A., Grotzinger J.P., Gupta S., Herkenhoff K.E., Hurowitz J.A., King P.L., Le Mouélic S., Ming D.W., Morris R.V., Nachon M., Newsom H.E., Ollila A.M., Perrett G.M., Rice M.S., Schmidt M.E., Schwenzer S.P., Stack K., Stolper E.M., Sumner D.Y., Treiman A.H., VanBommel S., Vaniman D.T., Vasavada A., Wiens R.C., Yingst R.A., and the MSL Science Team (2014) Elemental geochemistry of sedimentary rocks at Yellowknife Bay, Gale Crater, Mars. Science 343 10.1126/science.1244734 [DOI] [PubMed] [Google Scholar]
- McMahon S., Parnell J., Ponicka J., Hole M., and Boyce A. (2013) The habitability of vesicles in martian basalt. Astronomy & Geophysics 54:1.17–1.21 [Google Scholar]
- Meslin P.-Y., Gasnault O., Forni O., Schröder S., Cousin A., Berger G., Clegg S.M., Lasue J., Maurice S., Sautter V., Le Mouélic S., Wiens R.C., Fabre C., Goetz W., Bish D., Mangold N., Ehlmann B., Lanza N., Harri A.-M., Anderson R., Rampe E., McConnochie T.H., Pinet P., Blaney D., Léveillé R., Archer D., Barraclough B., Bender S., Blake D., Blank J.G., Bridges N., Clark B.C., DeFlores L., Delapp D., Dromart G., Dyar M.D., Fisk M., Gondet B., Grotzinger J., Herkenhoff K., Johnson J., Lacour J.-L., Langevin Y., Leshin L., Lewin E., Madsen M.B., Melikechi N., Mezzacappa A., Mischna M.A., Moores J.A., Newsom H., Ollila A., Perez R., Renno N., Sirven J.-B., Tokar R., de la Torre M., d'Uston L., Vaniman D., Yingst A., and the MSL Science Team (2013) Soil diversity and hydration as observed by ChemCam at Gale Crater, Mars. Science 341, 10.1126/science.1238670 [DOI] [PubMed] [Google Scholar]
- Michalski J.R. and Niles P.B. (2010) Deep crustal carbonate rocks exposed by meteor impact on Mars. Nat Geosci 3:751–755 [Google Scholar]
- Michalski J.R., Cuadros J., Niles P.B., Parnell J., Rogers A.D., and Wright S.P. (2013) Ground water activity on Mars and implications for a deep biosphere. Nat Geosci 6:133–138 [Google Scholar]
- Mileikowsky C., Cucinotta F., Wilson J.W., Gladman B., Horneck G., Lindgren L., Melosh H.J,. Rickman H., Valtonen M.J., and Zheng J.Q. (2000) Natural transfer of viable microbes in space part I: from Mars to Earth and Earth to Mars. Icarus 145:391–427 [DOI] [PubMed] [Google Scholar]
- Ming D.W., Archer P.D., Glavin D.P., Eigenbrode J.L., Franz H.B., Sutter B., Brunner A.E., Stern J.C., Freissinet C., McAdam A.C., Mahaffy P.R., Cabane M., Coll P., Campbell J.L., Atreya S.K., Niles P.B., Bell J.F., Bish D.L., Brinckerhoff W.B., Buch A., Conrad P.G., Des Marais D.J., Ehlmann B.L., Fairén A.G., Farley K., Flesch G.J., Fracois P., Gellert R., Grant J.A., Grotzinger J.P., Gupta S., Herkenhoff K.E., Hurowitz J.A., Leshin L.A., Lewis K.W., McLennan S.M., Miller K.E., Moersch J., Morris R.V., Navarro-González R., Pavlov A.A., Perrett G.M., Pradler I., Squyres S.W., Summons R.E., Steele A., Stolper E.M., Sumner D.Y., Szopa C., Vasavada A.R., Webster C.R., Wray J.J., Yingst R.A., and the MSL Science Team (2014) Volatile and organic compositions of sedimentary rocks in Yellowknife, Gale Crater, Mars. Science 343 10.1126/science.1245267 [DOI] [PubMed] [Google Scholar]
- Möhlmann D. (2009) Are nanometric films of liquid supercooled interfacial water bio-relevant? Cryobiology 58:256–261 [DOI] [PubMed] [Google Scholar]
- Möhlmann D. (2011) Three types of liquid water in icy surfaces of celestial bodies. Planet Space Sci 59:1082–1086 [Google Scholar]
- Moores J.E., Smith P.H., Tanner R., Schuerger A.C., and Venkateswaran K.J. (2007) The shielding effect of small-scale martian surface geometry on ultraviolet flux. Icarus 192:417–433 [Google Scholar]
- Morris R.V., Kilingelhofer C., Schroder C., Rodionov D.S., Ten A., Ming D., de Souza P.A., Wdowiak T., Fleischer I., Gellert R., Evlanov E.N., Hoh J., Gütlich P., Kankeleit E., McCoy T., Mittlefehldt D.W., Renz F., Schmidt M.E., Zubkov B., Squyres A.W., and Arvidson R.E. (2006) Mössbauer mineralogy of rock, soil, and dust at Meridiani Planum, Mars: Opportunity's journey across sulfate-rich outcrop, basaltic sand and dust, and hematite lag deposits. J Geophys Res 114, 10.1029/2006JE002791 [DOI] [Google Scholar]
- Morris R.V., Ming D.W., Yen A., Arvidson R.E., Gruener J., Humm D., Klingelhöfer G., Murchie S., Schröder C., Seelos F., Squyres S., Wisema S., Wolff M., and the MER and CRISM Science Teams (2007) Possible evidence for iron sulfates, iron sulfides, and elemental sulfur at Gusev Crater, Mars, from MER, CRISM, and analog data [abstract 3393]. In Seventh International Conference on Mars, Lunar and Planetary Institute, Houston [Google Scholar]
- Moser D.P., Gihring T.M., Brockman F.J., Fredrickson J.K., Balkwill D.L., Dollhopf M.E., Sherwood-Lollar B., Pratt L.M., Boice E., Southam G., Wanger G., Baker B.J., Pfiffner S.M., Lin L.-H., and Onstott T.C. (2005) Desulfotomaculum and Methanobacterium spp. dominate a 4- to 50-kilometer-deep fault. Appl Environ Microbiol 71:8773–8783 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mustard J.F., Ehlmann B.L., Murchie S.L., Poulet F., Mangold N., Head J.W., Bibring J.-P., and Roach L.H. (2009) Composition, morphology and stratigraphy of Noachian crust around the Isidis Basin. J Geophys Res 114, 10.1029/2009JE003349 [DOI] [Google Scholar]
- Neukum G., Basilevsky A.T., Kneissl T., Chapman M.G., van Gasselt S., Michael G., Jaumann R., Hoffmann H., and Lanz J.K. (2010) The geologic evolution of Mars: episodicity of resurfacing events and ages from cratering analysis of image data and correlation with radiometric ages of martian meteorites. Earth Planet Sci Lett 294:204–222 [Google Scholar]
- Nisbet E., Zahnle K., Gerasimov M.V., Helbert J., Jaumann R., Hofmann B.A., Benzerera K., and Westall F. (2007) Creating habitable zones, at all scales, from planets to mud-micro-habitats, on Earth and on Mars. Space Sci Rev 129:79–121 [Google Scholar]
- Nixon S.L., Cockell C.S., and Tranter M. (2012) Limitations to a microbial iron cycle on Mars. Planet Space Sci 72:116–128 [Google Scholar]
- Noblet A., Stalport F., Guan Y.Y., Poch O., Coll P., Szopa C., Cloix M., Macari F., Raulin F., Chaput D., and Cottin H. (2012) The PROCESS experiment: amino and carboxylic acids under Mars-like surface UV radiation conditions in low-Earth orbit. Astrobiology 12:436–444 [DOI] [PubMed] [Google Scholar]
- Nunes D.C., Smrekar S.E., Safaeinili A., Holt J.W., Phillips R.J., Seu R., and Campbell B.A. (2010) Examination of gully sites on Mars with the shallow radar. J Geophys Res 115, 10.1029/2009JE003509 [DOI] [Google Scholar]
- Okland I., Huang S., Dahle H., Thorseth L.H., and Pedersen R.B. (2012) Low temperature alteration of serpentinized ultramafic rock and implications for microbial life. Chem Geol 318:75–87 [Google Scholar]
- Okubo C.H., Schultz R.A., Chan M.A., Komatsu G., and the HiRISE Team (2009) Deformation band clusters on Mars and implications for subsurface fluid flow. Geol Soc Am Bull 121:474–482 [Google Scholar]
- Osinski G.R., Tornabene L.L., Banerjee N.J., Cockell C.S., Flemming R., Izawa M.R.W., McCutcheon J., Parnell J., Preston L.J., Pickersgill A.E., Pontefract A., Sapers H.M., and Southam G. (2013) Impact-generated hydrothermal systems on Earth and Mars. Icarus 224:347–363 [Google Scholar]
- Patel M.R., Bérces A., Kerékgyárto T., Rontó G., Lammer H., and Zarnecki J.C. (2004) Annual solar UV exposure and biological effective dose rates on the martian surface. Adv Space Res 33:1247–1252 [DOI] [PubMed] [Google Scholar]
- Pavlov A.A., Vasilyev G., Ostryakov V.M., Pavlov A.K., and Mahaffy P. (2012) Degradation of the organic molecules in the shallow subsurface of Mars due to irradiation by cosmic rays. Geophys Res Lett 39:L13202 [Google Scholar]
- Picardi G., Plaut J.J., Bicarri D., Bombaci O., Calabrese D., Cartacci M., Cicchetti A., Clifford S.M., Edenhofer P., Farrell W.M., Federico C., Frigeri A., Gurnett D.A., Hagfors T., Heggy E., Herique A., Huff R.L., Ivanov A.B., Johnson W.T.K., Jordan R.L., Kirchner D.L., Kofman W., Leuschen C.J., Nielsen E., Orosei R., Pettinelli E., Phillips R.J., Plettemeier D., Safaeinili A., Seu R., Stofan E.R., Vannaroni G., Watters T.R., and Zampolini E. (2005) Radar soundings of the subsurface of Mars. Science 310:1925–1928 [DOI] [PubMed] [Google Scholar]
- Poulet F., Bibring J.-P., Mustard J.F., Gendrin A., Mangold N., Langevin Y., Arvidson R.E., Gondet B., Gomez C., Berthé M., Erard S., Forni O., Manaud N., Poulleau G., Soufflot A., Combes M., Drossart P., Encrenaz T., Fouchet T., Melchiorri R., Belluci G., Altieri F., Formisano V., Fonti S., Capaccioni F., Cerroni P., Coradini A., Korablev O., Kottsoy V., Ignatiey N., Titov D., Zasova L., Pinet P., Schmitt V.B., Sotin C., Hauber E., Hoffmann H., Jaumann R., Keller U., and Forget F. (2005) Phyllosilicates on Mars and implications for early martian climate. Nature 438:623–627 [DOI] [PubMed] [Google Scholar]
- Pronk J.T., de Bruyn J.C., Bos P., and Kuenen J.G. (1992) Anaerobic growth of Thiobacillus ferroooxidans. Appl Environ Microbiol 58:2227–2230 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quantin C., Flahut J., Clenet H., Allemand P., and Thomas P. (2012) Composition and structures of the subsurface in the vicinity of Valles Marineris as revealed by central uplifts of impact craters. Icarus 221:436–452 [Google Scholar]
- Reiss D., Erkeling G., Bauch K.E., and Hiesinger H. (2010) Evidence for present-day gully activity on the Russell Crater dune field. Geophys Res Lett 37, 10.1029/2009GL042192 [DOI] [Google Scholar]
- Rennó N.O., Bos B.J., Catling D.C., Clark B.C., Drube L., Fisher D., Goetz W., Hviid S.F., Keller H.U., Kok J.F., Kouvanes S.P., Leer K., Lemmon M., Madsen M.B., Markiewicz W.J., Marshall J., McKay C.P., Mehta M., Smith M., Zorzano M.P., Smith P.H., Stoker C., and Young S.M.M. (2009) Possible physical and thermodynamical evidence for liquid water at the Pheonix landing site. J Geophys Res 114, 10.1029/2009JE003362 [DOI] [Google Scholar]
- Rogers A.D. (2011) Crustal compositions exposed by impact craters in the Tyrrhena Terra region of Mars: considerations for Noachian environments. Earth Planet Sci Lett 301:353–364 [Google Scholar]
- Rontó G., Bérces A., Lammer L., Cockell C.S., Molina-Cuberos G.J., Patel M.R., and Selsis F. (2003) Solar UV irradiation conditions on the surface of Mars. Photochem Photobiol 77:34–40 [DOI] [PubMed] [Google Scholar]
- Sagan C. and Pollack J.B. (1974) Differential transmission of sunlight on Mars: biological implications. Icarus 21:490–495 [Google Scholar]
- Schon S.C. and Head J.W. (2011) Keys to gully formation processes on Mars: relation to climate cycles and sources of meltwater. Icarus 213:428–432 [Google Scholar]
- Schuerger A.C., Richards J.T., Newcombe D.A., and Venkateswaran K. (2006) Rapid inactivation of seven Bacillus spp. under simulated Mars UV irradiation. Icarus 181:52–62 [Google Scholar]
- Schulze-Makuch D., Irwin L.N., Lipps J.H., LeMone D., Dohm J.M., and Fairén A.G. (2005) Scenarios for the evolution of life on Mars. J Geophys Res 110, 10.1029/2005JE002430 [DOI] [Google Scholar]
- Schulze-Makuch D., Irwin L.N., and Fairén A.G. (2013) Drastic environmental change and its effects on a planetary biosphere. Icarus 225:775–780 [Google Scholar]
- Schwenzer S.P. and Kring D.A. (2009) Impact-generated hydrothermal systems: capable of forming phyllosilicates on Noachian Mars. Geology 37:1091–1094 [Google Scholar]
- Schwenzer S.P., Abramov O., Allen C.C., Clifford S.M., Cockell C.S., Filiberto J., Kring D.A., Lasue J., McGovern P.J., Newsom H.E., Treiman A.H., Vaniman D.T., and Wiens R.C. (2012) Puncturing Mars: how impact craters interact with the martian cryosphere. Earth Planet Sci Lett 335:9–17 [Google Scholar]
- Scott E.R. (1999) Origin of carbonate-magnetite-sulfide assemblages in martian meteorite ALH84001. J Geophys Res 104:3803–3813 [DOI] [PubMed] [Google Scholar]
- Segura A. and Navarro-González R. (2005) Nitrogen fixation on early Mars by volcanic lightning and other sources. Geophys Res Lett 32, 10.1029/2004GL021910 [DOI] [Google Scholar]
- Seibert N.M. and Kargel J.S. (2001) Small-scale martian polygonal terrain: implications for liquid surface water. Geophys Res Lett 28, 10.1029/2000GL012093 [DOI] [Google Scholar]
- Sharp R.P. and Malin M.C. (1975) Channels on Mars. Geol Soc Am Bull 86:593–609 [Google Scholar]
- Smith H.D. and McKay C.P. (2005) Drilling in ancient permafrost on Mars for evidence of a second genesis of life. Planet Space Sci 53:1302–1308 [Google Scholar]
- Smith P., Tamppari L.K., Arvidosn R.E., Bass D., Blaney D., Boynton W.V., Carswell A., Catling D.C., Clark B.C., Duck T., Dejong E., Fisher D., Goetz W., Gunnlaugsson H.P., Hecht M.H., Hipkin V., Hoffman J., Hviid S.F., Keller H.U., Kounaves S.P., Lange C.F., Lemmon M.T., Madsen M.B., Markiewicz W.J., Marshall J., McKay C.P., Mellon M.T., Ming D.W., Morris R.V., Pike W.T., Renno N., Staufer U., Stoker C., Taylor P., Whiteway J.A., and Zent A.P. (2009) H2O at the Phoenix landing site. Science 325:58–61 [DOI] [PubMed] [Google Scholar]
- Soare R.J., Conway S.J., Pearce G.D., Dohm J.M, and Grindrod P.M. (2013) Possible crater-based pingos, paleolakes and periglacial landscapes in the high latitudes of Utopia Planitia, Mars. Icarus 225:971–981 [Google Scholar]
- Solomon S.C. and Head J.W. (1990) Heterogeneities in the thickness of the elastic lithosphere of Mars: constraints on heat flow and internal dynamics. J Geophys Res 95:11073–11083 [Google Scholar]
- Squyres S.W. and Carr M.H. (1986) Geomorphic evidence for the distribution of ground ice on Mars. Science 231:249–252 [DOI] [PubMed] [Google Scholar]
- Squyres S.W., Arvidson R.E., Bell J.F., Calef F., Clark B.C., Cohen B.A., Crumpler L.A., de Souza P.A., Farrard W.H., Gellert R., Grant J., Herkenhoff K.E., Hurowitz J.A., Johnson J.R., Jolliff B.L., Knoll A.H., Li R., McLennan S.M., Ming D.W., Mittlefehldt D.W., Parker T.J., Paulsen G., Rice M.S., Ruff S.W., Chroder C., Yen A.S., and Zacny K. (2012) Ancient impact and aqueous processes at Endeavor Crater, Mars. Science 336:570–576 [DOI] [PubMed] [Google Scholar]
- Steele A., McCubbin F.M., Fries M., Kater L., Boctor N.Z., Fogel M.L., Conrad P.G., Glamoclija M., Spencer M., Morrow A.L., Hammond M.R., Zare R.N., Vicenzi E.P., Siljeström S., Bowden R., Herd C.D.K., Mysen B.O., Shirey S.B., Amundsen H.E.F., Treiman A.H., Bullock E.S., and Jull A.J.T. (2012) A reduced organic carbon component in martian basalts. Science 337:212–215 [DOI] [PubMed] [Google Scholar]
- Stephenson J.D., Hallis L.J., Nagashima K., and Freeland S.J. (2013) Boron enrichment in martian clay. PLoS One 8:e64624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stoker C.R., Zent A., Catling D.C., Douglas S., Marshall J.R., Archer D., Clark B., Kouvanes S.P., Lemmon M.T., Quinn R., Renno N., Smith P.H., and Young S.M.M. (2010) Habitability of the Phoenix landing site. J Geophys Res 115, 10.1029/2009JE003421 [DOI] [Google Scholar]
- Stolper E.M., Baker M.B., Newcombe M.E., Schmidt M.E., Treiman A.H., Cousin A., Dyar M.D., Fisk M.R., Gellert R., King P.L., Leshin L., Maurice S., McLennan S.M., Minitti M.E., Perrett G., Rowland S., Sautter V., Wiens R.C., and the MSL Science Team (2013) The petrochemistry of Jake_M: a martian mugearite. Science 341, 10.1126/science.1239463 [DOI] [PubMed] [Google Scholar]
- Straub K.L., Benz M., Schink B., and Widdel F. (1996) Anaerobic, nitrate-dependent microbial oxidation of ferrous iron. Appl Environ Microbiol 62:1458–1460 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Summers D.P. and Khare B. (2007) Nitrogen fixation on early Mars and other terrestrial planets: experimental demonstration of abiotic fixation reactions to nitrite and nitrate. Astrobiology 7:333–341 [DOI] [PubMed] [Google Scholar]
- Szponar N., Brazelton W.L., Screnk M.O., Bower D.M., Steele A., and Morrill P.L. (2013) Geochemistry of a continental site of serpentinization, the Tablelands Ophiolite, Gros Morne National Park: a Mars analogue. Icarus 224:286–296 [Google Scholar]
- Takai K., Nakamura K., Toki T., Tsunogai U., Miyazaki M., Miyazaki J., Hirayama H., Nakagawa S., Nunoura T., and Horikoshi K. (2008) Cell proliferation at 122°C and isotopically heavy CH4 production by a hyperthermophilic methanogen under high pressure cultivation. Proc Natl Acad Sci USA 105:10949–10954 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanaka K.L. (1986) The stratigraphy of Mars. J Geophys Res 91, 10.1029/JB091iB13p0E139 [DOI] [Google Scholar]
- Taylor F.W. (2011) Comparative planetology, climatology and biology of Venus, Earth and Mars. Planet Space Sci 59:889–899 [Google Scholar]
- Taylor R.R. and McLennan S.M. (2009) Planetary Crusts: Their Composition, Origin, and Evolution, Cambridge University Press, Cambridge, UK [Google Scholar]
- Tosca J.N., McLennan S.M., Clark B., Grotzinger J.P., Hurowitz J.A., Knoll A.H., Schröder C., and Squyres S.W. (2005) Geochemical modelling of evaporation processes on Mars: insight from the sedimentary record at Meridiani Planum. Earth Planet Sci Lett 240:122–148 [Google Scholar]
- Tosca N.J., Knoll A.H., and McLennan S.M. (2008a) Water activity and the challenge for life on early Mars. Science 320:1204–1207 [DOI] [PubMed] [Google Scholar]
- Tosca N.J., McLennan S.M., Dyar M.D., Sklute E.C., and Michel F.M. (2008b) Fe oxidation processes at Meridiani Planum and implications for secondary Fe mineralogy on Mars. J Geophys Res 113, 10.1029/2007JE003019 [DOI] [Google Scholar]
- Tuff J., Wade J., and Wood J. (2013) Volcanism on Mars controlled by early oxidation of the upper mantle. Nature 498:342–345 [DOI] [PubMed] [Google Scholar]
- Ulrich M., Wagner D., Hauber E., de Vera J.-P., and Schirrmeister L. (2012) Habitable periglacial landscapes in martian mid-latitudes. Icarus 219:345–357 [Google Scholar]
- Urbach E.R. and Stepinski T.F. (2009) Automatic detection of sub-kilometer craters in high resolution planetary images. Planet Space Sci 57:880–887 [Google Scholar]
- Usui T., McSween H.Y., Jr., and Clark B.C. (2008) Petrogenesis of high-phosphorus Wishstone Class rocks in Gusev Crater, Mars. J Geophys Res 113, 10.1029/2008JE003225 [DOI] [Google Scholar]
- Vaniman D.T., Bish D.L., Ming D.W., Bristow T.F., Morris R.V., Blake D.F., Chipera S.J., Morrison S.M., Treiman A.H., Rampe E.B., Rice M., Achilles C.N., Grotzinger J., McLennan S.M., Williams J., Bell J., Newsom H., Downs R.T., Maurice S., Sarrazin P., Yen A.S., Morookian J.M., Farmer J.D., Stack K., Milliken R.E., Ehlmann B., Sumner D.Y., Berger G., Crisp J.A., Hurowitz J.A., Anderson R., Des Marais D., Stolper E.M., Edgett K.S., Gupta D., Spanovich N., and the MSL Science Team (2014) Mineralogy of a mudstone at Yellowknife Bay, Gale Crater, Mars. Science 343 10.1126/science.1243480 [DOI] [PubMed] [Google Scholar]
- Webster C.R., Mahaffy P.R., Atreya S.K., Flesch G.J., Farley K.A., and the MSL Science Team (2013) Low upper limit to methane abundance on Mars. Science 342:355–357 [DOI] [PubMed] [Google Scholar]
- Weiss B.P., Yung Y.L., and Nealson K.H. (2000a) Atmospheric energy for subsurface life on Mars? Proc Natl Acad Sci USA 97:1395–1399 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weiss B.P., Kirschvink J.L., Baudenbacher F.J., Vali H., Peters N.T., Macdonald F.A., and Wikswo J.P. (2000b) A low temperature transfer of ALH84001 from Mars to Earth. Science 290:791–795 [DOI] [PubMed] [Google Scholar]
- Westall F., Loizeau D., Foucher F., Bost N., Betrand M., Vago J., and Kminek G. (2013) Habitability on Mars from a microbial point of view. Astrobiology 13:887–897 [DOI] [PubMed] [Google Scholar]
- Wray J.J. and Ehlmann B.L. (2011) Geology of possible martian methane source regions. Planet Space Sci 59:196–202 [Google Scholar]
- Wright I.P., Grady M.M., and Pillinger C.T. (1992) Chassigny and the nakhlites: carbon-bearing components and their relationship to martian environmental conditions. Geochim Cosmochim Acta 56:817–826 [Google Scholar]
- Zacny K., Bar-Cohen Y., Brennan M., Briggs G., Cooper G., Davis K., Dolgin B., Glaser D., Glass B., Gorevan S., Guerrero J., McKay C., Paulsen G., Stanley S. and Stoker C. (2008) Drilling systems for extraterrestrial subsurface exploration. Astrobiology 8:665–706 [DOI] [PubMed] [Google Scholar]
- Zolotov M.Y. and Shock E.L. (2005) Formation of jarosite-bearing deposits through aqueous oxidation of pyrite at Meridiani Planum, Mars. Geophys Res Lett 32:L21203 [Google Scholar]
- Zuber M.T., Smith D.E., Solomon S.C., Abshire J.B., Afzal R.S., Aharonson O., Fishbaugh K., Ford P.G., Frey H.V., Garvin J.B., Head J.W., Ivanov A.B., Johnson C.L., Muhleman D.O., Newmann G.A., Pettengill G.H., Pillips R.J., Sun X., Zwally J., Banerdt W.B., and Duxbury T.C. (1998) Observations of the north polar regions of Mars from the Mars Orbiter Laser Altimeter (MOLA). Science 282:2053–2060 [DOI] [PubMed] [Google Scholar]