

Abstract
BACKGROUND
Depletion of the Ca2+ store by ryanodine receptor (RyR) agonists induces store-operated Ca2+ entry (SOCE). 4-Chloro-3-ethylphenol (4-CEP) and 4-chloro-m-cresol (4-CmC) are RyR agonists commonly used as research tools and diagnostic reagents for malignant hyperthermia. Here, we investigated the effects of 4-CEP and its analogues on SOCE.
EXPERIMENTAL APPROACH
SOCE and ORAI1-3 currents were recorded by Ca2+ imaging and whole-cell patch recordings in rat L6 myoblasts and in HEK293 cells overexpressing STIM1/ORAI1-3.
KEY RESULTS
4-CEP induced a significant release of Ca2+ in rat L6 myoblasts, but inhibited SOCE. The inhibitory effect was concentration-dependent and more potent than its analogues 4-CmC and 4-chlorophenol (4-ClP). In the HEK293 T-REx cells overexpressing STIM1/ORAI1-3, 4-CEP inhibited the ORAI1, ORAI2 and ORAI3 currents evoked by thapsigargin. The 2-APB-induced ORAI3 current was also blocked by 4-CEP. This inhibitory effect was reversible and independent of the Ca2+ release. The two analogues, 4-CmC and 4-ClP, also inhibited the ORAI1-3 channels. Excised patch and intracellular application of 4-CEP demonstrated that the action site was located extracellularly. Moreover, 4-CEP evoked STIM1 translocation and subplasmalemmal clustering through its Ca2+ store-depleting effect via the activation of RyR, but no effect on STIM1 redistribution was observed in cells co-expressing STIM1/ORAI1-3.
CONCLUSION AND IMPLICATIONS
4-CEP not only acts as a RyR agonist to deplete the Ca2+ store and trigger STIM1 subplasmalemmal translocation and clustering, but also directly inhibits ORAI1-3 channels. These findings demonstrate a novel pharmacological property for the chlorophenol derivatives that act as RyR agonists.
Keywords: ryanodine receptors, store-operated Ca2+ channels, STIM1, ORAI, 4-chloro-3-ethylphenol, 4-chloro-m-cresol
Introduction
Ryanodine receptors (RyRs) are important intracellular Ca2+ release channels in the sarcoplasmic/endoplasmic reticulum (SR/ER) (Fill and Copello, 2002). They are ubiquitously expressed in different cell types including both excitable and non-excitable cells (Lanner et al., 2010). RyR1 is the primary isoform in skeletal muscle, while RyR2 is the cardiac isoform, and RyR3, previously referred to as the brain isoform, exists in many tissues (Fill and Copello, 2002; Endo, 2009; Lanner et al., 2010). RyRs are considered particularly important in excitable cells, such as cardiac and skeletal muscles and neurons. Membrane depolarization in cardiomyocytes or neurons activates voltage-gated L-type Ca2+ channels, which results in Ca2+ influx and subsequently triggers the Ca2+-induced Ca2+ release from SR/ER through RyRs, and thus causes muscle contraction or alter the functionality of neurons (Lanner et al., 2010; Rose and Konnerth, 2001). In skeletal muscle, physiological Ca2+ release is induced by t-tubule membrane depolarization, which transmits the signal to RyRs through a mechanism involving a protein-protein interaction (Endo, 2009). Dysfunctional RyRs cause diseases, for example, RyR1 mutations are involved in central core disease and malignant hyperthermia (MH) (Robinson et al., 2006; Hopkins, 2011), and RyR2 mutations are associated with catecholaminergic polymorphic ventricular tachycardia and arrhythmogenic right ventricular cardiomyopathy (Lanner et al., 2010; Priori and Chen, 2011). RyR3 is the least studied isoform, but it has been suggested to play a role in Alzheimer's disease (Supnet et al., 2010).
Caffeine and chlorophenol derivatives, structurally different RyR agonists, are commonly used as pharmacological research tools and diagnostic drugs for MH (Figure 1A) (DiJulio et al., 1997; Westerblad et al., 1998; Baur et al., 2000; Varadi and Rutter, 2002; Gerbershagen et al., 2005). These two classes of structure may have different binding sites to RyRs (Tian et al., 2013). The chlorophenol derivatives 4-chloro-3-ethylphenol (4-CEP) and 4-chloro-m-cresol (4-CmC) can induce a robust release of intracellular Ca2+ by stimulating RyRs at concentrations about 10-fold lower than caffeine (Islam et al., 1998; Jacobson et al., 2006), and have advantages over caffeine and ryanodine as diagnostic drugs for MH (Gerbershagen et al., 2005). It has been demonstrated that activation of RyRs by ryanodine or repetitive depolarization can induce store-operated Ca2+ entry (SOCE) and a Ca2+ release-activated current in smooth muscle and skeletal muscle cells (Fellner and Arendshorst, 2000; Yarotskyy and Dirksen, 2012). ORAI and STIM proteins are recently identified molecules that act as store-operated channels (Zhang et al., 2005; Feske et al., 2006; McNally et al., 2012). The nomenclature and pharmacological characteristics of ORAI store-operated channels are still not known (Vig et al., 2008; Alexander et al., 2013), although they have been shown to be blocked by certain compounds, such as 2-aminoethoxydiphenyl borate (2-APB) (Peinelt et al., 2008), lanthanides and BTP2 (Zitt et al., 2004; Shuttleworth, 2012). Recently, using HEK293 cells transfected with STIM1 tagged with enhance yellow fluorescent protein (EYFP), we showed that the depletion of Ca2+ stores by the RyR agonists caffeine and 4-CEP evoked the release of Ca2+ accompanied by STIM1 translocation and subplasmalemmal clustering (Zeng et al., 2012), and the RyR antagonist tetracaine prevented this intracellular movement of STIM1 (Zeng et al., 2012). This provides direct evidence that RyRs induce the activation of store-operated channels (SOCs) (Zhang et al., 2005). Interestingly, upon re-introduction of external Ca2+ into the bath solution, caffeine induced the release of Ca2+ followed by a significant SOCE, but 4-CEP only induced a robust release of Ca2+ with little Ca2+ influx (Zeng et al., 2012). Furthermore, caffeine evokes intracellular Ca2+ oscillations in bullfrog sympathetic ganglion cells, but the 4-CEP analogue cresol does not (Higure et al., 2006). These observations strongly suggest the existence of functional coupling between RyRs and SOCs via intracellular STIM1 movement, and that caffeine and 4-CEP probably have different effects on the dynamics of intracellular Ca2+. Therefore, we hypothesized that the unexpected reduction in Ca2+ influx induced by 4-CEP could be due to the blockage of store-operated ORAI channels.
Figure 1.
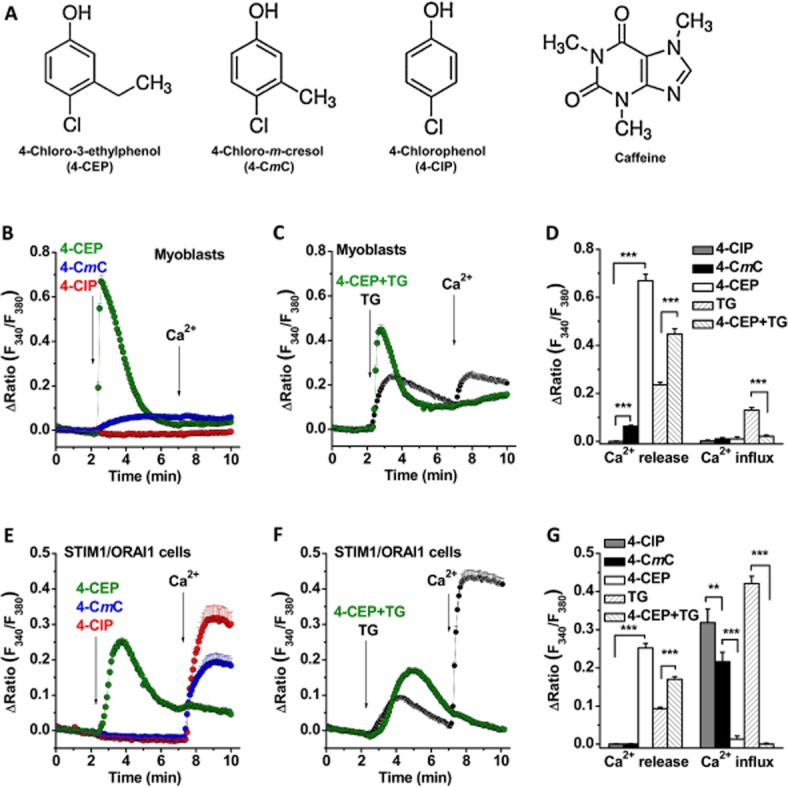
4-CEP induced Ca2+ release but abolished store-operated Ca2+ entry in rat L6 myoblasts and HEK293 cells co-expressing STIM1/ORAI1. (A) Structures for 4-CEP, 4-CmC, 4-ClP and caffeine. (B) The effects of 4-CEP, 4-CmC and 4-ClP on the cytosolic Ca2+ level in rat L6 myoblasts. The cells were maintained in Ca2+ free solution first and then perfused with different compounds (500 μM for each compound) in Ca2+ free and 1.5 mM Ca2+ solution as indicated by the arrows. (C) 4-CEP (500 μM) nearly abolished the TG (1 μM)-induced Ca2+ influx in the myoblasts. (D) Mean ± SEM data for the comparison of Ca2+ release and influx induced by 4-ClP, 4-CmC, 4-CEP and TG in the myoblasts (n = 19–40 cells in each group). (E) The effects of 4-CEP, 4-CmC and 4-ClP (500 μM) in STIM1/ORAI1 cells. (F) 4-CEP (500 μM) abolished the TG (1 μM)-induced Ca2+ influx in STIM1/ORAI1 cells. (G) Mean ± SEM data for Ca2+ release and influx induced by 4-ClP, 4-CmC, 4-CEP and TG in the STIM1/ORAI1 cells (n = 18–26 cells in each group). Ca2+ release was measured as the peak of Ca2+ release signal; and Ca2+ influx was measured at 1 min after Ca2+ re-addition. One-way ANOVA with Bonferroni test was used for (D) and (G). ** P < 0.01, *** P < 0.001.
To test this hypothesis, we investigated the effects of 4-CEP on the SOCE and ORAI1-3 channels using rat L6 myoblasts derived from skeletal muscle and HEK293 cells transfected with ORAI1-3 and STIM1 genes. We also compared the effect of 4-CEP with those of 4-CmC and 4-chlorophenol (4-ClP) to determine any potential structure-activity relationship for their antagonism of ORAI channels.
Methods
Chemicals and reagents
Generally used salts and other chemicals including thapsigargin (TG), 2-APB, caffeine, 4-CEP, 4-CmC, 4-ClP and Fura-PE3/AM were purchased from Sigma-Aldrich (Poole, UK).
Cell culture and transfection
The HEK293 T-REx cells were cultured in DMEM/F-12 medium (Invitrogen, Paisley, UK) supplemented with 10% fetal calf serum, 100 units·mL−1 penicillin and 100 mg·mL−1 streptomycin, and maintained at 37°C under 95% air and 5% CO2. Rat L6 myoblasts were purchased from ATCC (Manassas, VA, USA) and cultured in the same conditions. Plasmids encoding mCherry-ORAI1, mCherry-ORAI2 and monomeric cyan fluorescent protein (mCFP)-ORAI1, mCFP-ORAI2 and mCFP-ORAI3 in the pcDNA4/TO vector were transfected into stable STIM1-EYFP cells and the expression of these genes was induced with 1 μg·mL−1 tetracycline in the culture medium. Details of the procedures for gene cloning and plasmid constructions are as described previously (Zeng et al., 2012). The stable cells were maintained in the cell culture medium containing G418 (200 μg·mL−1).
Electrophysiology
Whole-cell patch-clamp recording was performed at room temperature (23–26°C). Briefly, the signal was amplified with an Axopatch 200B amplifier and controlled by the software pClamp 10. A 1-s ramp voltage protocol from −100 mV to +100 mV was applied at a frequency of 0.2 Hz from a holding potential of 0 mV. Signals were sampled at 10 kHz and filtered at 1 kHz. Glass microelectrodes with resistances of 3–5 MΩ were used. The pipette solution contained (in mM) 145 Cs-methanesulfonate, 10 BAPTA, 8 MgCl2 and 10 HEPES (pH 7.2 adjusted with CsOH). The standard bath solution contained (mM) 130 NaCl, 5 KCl, 8 d-glucose, 10 HEPES, 1.2 MgCl2 and 1.5 CaCl2. The pH was adjusted to 7.4 with NaOH.
Live cell imaging and Ca2+ measurement
The stably transfected STIM1/ORAI1-3 cells were seeded on 13-mm glass cover slips and cultured for 24–48 h. Live cell images for EYFP/mCherry/mCFP fluorescence were captured using a microscope equipped with a Nikon Plan Fluor 100 × /1.30 oil objective by using similar procedures to those reported previously (Zeng et al., 2012). The images were analysed using NIS-Elements software (Version 3.2, Nikon, Tokyo, Japan).
For intracellular Ca2+ measurements, cells were loaded with 2 μM Fura-PE3/AM in standard bath solution for 30 min at 37°C, followed by 5 min wash in standard bath solution at room temperature. Cells were excited alternately by 340 and 380 nm light and emission was collected via a 510-nm filter. Images were sampled every 5 s in pairs for the two excitation wavelengths by a CCD camera (ORCA-R2, Hamamatsu, Japan). The ratio of 340/380 nm fluorescence represented the intracellular Ca2+ level. The standard bath solution was the same as that used in the patch-clamp experiment. The Ca2+-free solution contained (mM): NaCl 130, KCl 5, MgCl2 1.2, HEPES 10, Glucose 8, and EGTA 0.4. The pH was adjusted to 7.4 with NaOH. All experiments were performed at room temperature. The n values given are the numbers of cells from at least three independent Ca2+ imaging experiments.
Statistics
All values are expressed as mean ± SEM. Student's t-test was used to assess the statistical difference between two groups. One-way ANOVA with Bonferroni post hoc test was used for data sets with more than two groups. A P value less than 0.05 was considered significant.
Results
Effects of 4-CEP, 4-CmC and 4-ClP on Ca2+ release and SOCE
The effects of 4-CEP and its analogues on Ca2+ release and SOCE were observed. In rat L6 myoblasts, 4-CEP significantly evoked a Ca2+ release signal in Ca2+ free solution, but the same concentrations of 4-CmC showed only a small increase and 4-ClP showed no effect (Figure 1B). Re-introduction of 1.5 mM Ca2+ did not cause significant cytosolic Ca2+ increase in cells in the absence of TG treatment; however, store-operated Ca2+ entry (SOCE) was evident after store depletion with 1 μM TG. 4-CEP significantly inhibited the influx of store-operated Ca2+ (Figure 1C–D). To further confirm the effects on SOCE, the HEK293 T-REx cells overexpressing STIM1-EYFP and mCFP-ORAI1 were used. The SOCE in STIM1/ORAI1 cells was significantly higher than that in non-transfected cells (Supporting Information Figure S1). Perfusion with 4-CEP (500 μM) induced a robust Ca2+ release signal, as seen in the myoblasts, but 4-CmC (500 μM) and 4-ClP (500 μM) did not cause Ca2+ release (Figure 1E). The different effects on Ca2+ release for the three compounds in the two cell types could be due to the differential expression of RyR isoforms in these cells and the different affinities of the compounds for RyRs; indeed, RyR1 is a dominant isoform in the skeletal muscle derived myoblasts, while RyR3 is dominant in HEK293 cells (Van Petegem, 2012) and RyR3 is sensitive to 4-CEP, but almost insensitive to 4-CmC (Fessenden et al., 2003; Hopkins, 2011). In addition, the Ca2+ influx in the 4-CEP-treated cells was nearly abolished after the addition of 1.5 mM Ca2+ in the bath solution (Figure 1E), which accords with our previous observations in the HEK293 cells transfected with STIM1-EYFP alone (Zeng et al., 2012). The Ca2+ influx in the 4-CmC-treated cells was much higher than that in the 4-CEP group, but lower than that in the cells treated with 4-ClP (Figure 1E). The inhibition of the store-operated Ca2+ influx induced by 4-CEP was also significant in the TG-treated, store-depleted cells (Figure 1F). The IC50 values for 4-CEP, 4-CmC and 4-ClP on the TG-induced SOCE in STIM1/ORAI1 cells were 203.6, 830.9 and 1437.1 μM respectively (Supporting Information Figure S2). These results suggest that 4-CEP can evoke Ca2+ release via activation of RyRs, but significantly inhibits SOCs.
STIM1 translocation evoked by 4-CEP through RyR-mediated Ca2+ store depletion
To elucidate the mechanism of action of 4-CEP on SOCs, we examined the effects of 4-CEP on STIM1 translocation and subplasmalemmal clustering, which are critical steps for store-operated channel activation (Zhang et al., 2005). We recently demonstrated, in HEK293 T-REx cells overexpressing STIM1-EYFP, that 4-CEP can induce STIM1 translocation to the plasma membrane followed by the gradual formation of cytosolic STIM1 clusters (Zeng et al., 2012). However, such cytosolic STIM1 clustering did not occur in our cells co-expressing STIM1-EYFP and ORAIs, although the STIM1 puncta formation at the plasma membrane was reproducible in the STIM1-EYFP/mCFP-ORAI1 cells after the exposure to 500 μM 4-CEP in the Ca2+-containing solution (Figure 2). Importantly, the puncta near the plasma membrane did not disappear or redistribute in the STIM1/ORAI1 cells and almost no STIM1-EYFP clusters were formed in the cytosol after addition of 4-CEP for 5–10 min (Figure 2A, Supporting Information Videos S1 and S2). A similar phenomenon was observed in the cells co-expressing STIM1-EYFP/mCFP-ORAI2 or STIM1-EYFP/mCFP-ORAI3 (Figure 2A). These results suggest that the STIM1/ORAIs complexes are more stable than STIM1 oligomers, and may not disassociate easily and redistribute into the cytosol. This finding is consistent with our previous observation using 2-APB; 2-APB disassembled the subplasmalemmal STIM1 clusters after store depletion in the STIM1–EYFP cells, but lost the ability to disassemble the subplasmalemmal STIM1 clusters in the STIM1–EYFP cells coexpressing mCherry-ORAI1, mCherry-ORAI2 or mCFP-ORAI3 (Zeng et al., 2012). We also observed the effects of 4-CmC and 4-ClP, both compounds at 500 μM failed to induce STIM1 translocation in all of the three STIM1/ORAI co-expressing cell lines, whereas the subsequent perfusion with 500 μM 4-CEP clearly potentiated the formation of STIM1 puncta at the plasma membrane (Figure 2B). These results could be explained by differences in their ability to induce Ca2+ release through RyRs (Figure 1E), suggesting that the Ca2+ store depletion, such as that induced by the RyR agonist 4-CEP, is necessary for triggering STIM1 translocation and subplasmalemmal clustering.
Figure 2.
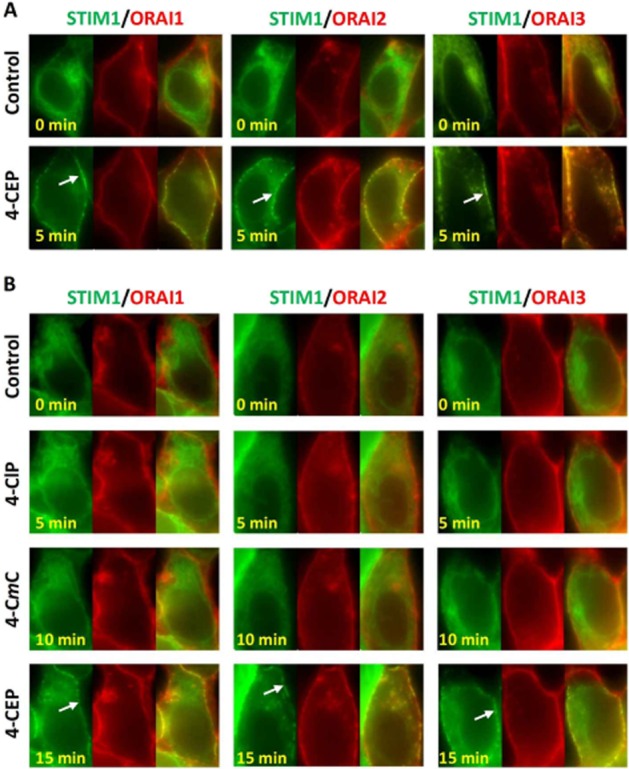
STIM1 translocation and clustering induced by 4-CEP, but not by 4-CmC and 4-ClP in the STIM1/ORAI1-3 cells. (A) Application of 500 μM 4-CEP for 5 min induced persistant STIM1-EYFP puncta (green, indicated by arrow) at the plasma membrane in STIM1-EYFP/mCFP-ORAI1-3 cells, while cytosolic clustering of STIM1 was not evident. The mCFP-ORAI fluorescence was converted into red pseudocolour in the pictures. The merged images are shown on the right of each group. The example cell for each group is a representative of 15 to 26 cells. (B) The cells were successively challenged with 500 μM 4-ClP, 500 μM 4-CmC and 500 μM 4-CEP in 1.5 mM Ca2+ solution with 5 min interval for each drug. STIM1 puncta at the plasma membrane (arrow) seen in 4-CEP-treated cells, but no STIM1 clustering was induced by 4-CmC and 4-CIP.
Inhibition of ORAI channels by 4-CEP, 4-CmC and 4-ClP
The subplasmalemmal translocation of STIM1 induced by 4-CEP is apparently not the mechanism of SOC inhibition, because the persistence of STIM1/ORAI complexes at the plasma membrane induced by 4-CEP implies that STIM1 proteins may keep the active conformation to maintain the ORAI channel opening, therefore, the inhibitory effect of 4-CEP on SOCE or store-operated current seen in the STIM1/ORAI co-expressing cells and myoblasts could be a direct effect on ORAI channels per se. To test this, we determined the effects of 4-CEP on ORAI1-3 store-operated channels in the HEK293 T-REx cells co-expressing STIM1/ORAIs. The ORAI store-operated currents evoked by TG in the ORAI1/STIM1, ORAI2/STIM1 and ORAI3/STIM1 cells were abolished by 500 μM 4-CEP (Figure 3A–C). The ORAI currents were also partially inhibited by 500 μM 4-CmC and 4-ClP (Figure 3D–I), suggesting that 4-CEP inhibits all three store-operated ORAI currents. However, the TG-induced endogenous store-operated current in the non-transfected HEK293 T-REx cells was not significant (Figure 3J). We further tested whether the 2-APB-induced ORAI3 current could be inhibited by the three drugs in the cells overexpressing ORAI3 alone. As shown in Figure 3K, 2-APB at 50 μM potentiated a large outward current and a relatively small inward current in ORAI3 cells, which is independent of STIM1 and Ca2+ store depletion (Shuttleworth, 2012). The ORAI3 current was significantly inhibited by 500 μM of 4-ClP, 4-CmC and 4-CEP successively with potencies in the order of 4-CEP > 4-CmC > 4-ClP (Figure 3K–L, also see Supporting Information Figure S3). These results suggest that both the TG-induced store-operated currents and 2-APB-activated ORAI3 currents are inhibited by the three drugs.
Figure 3.
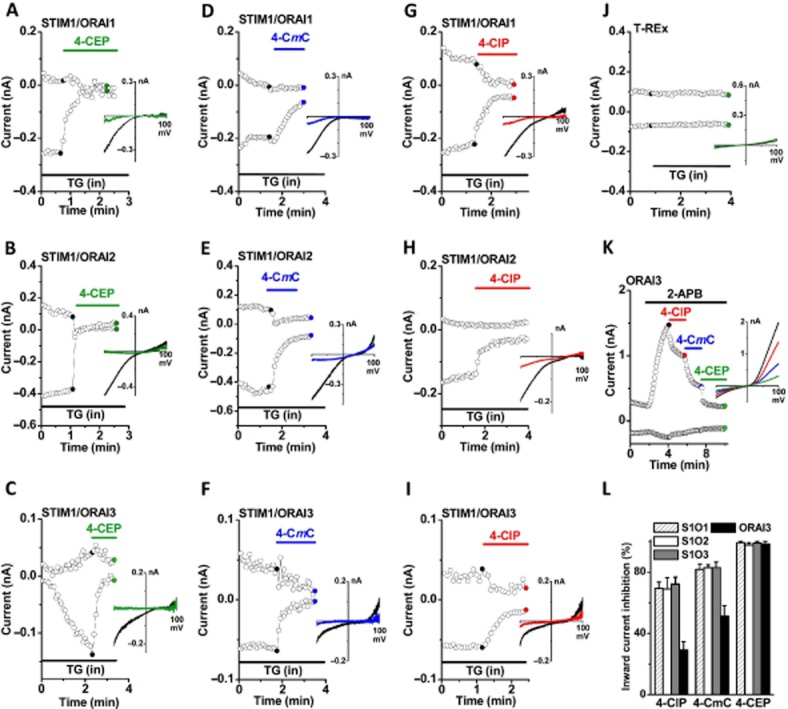
ORAI1-3 currents inhibited by 4-CEP, 4-CmC and 4-ClP in STIM1/ORAIs cells. Whole-cell currents were recorded in the HEK293 T-REx cells co-expressing STIM1-EYFP and ORAI1-3, or expressing ORAI3 alone. The store-operated currents in STIM1/ ORAI1-3 cells were evoked by TG (1 μM) in the pipette solution, and the current in ORAI3 cells was induced by 2-APB (50 μM). The time course for the amplitude of whole-cell currents was given at ±80 mV with corresponding leak-subtracted I/V relationships before and after treatment (inset). (A–C) Effect of 4-CEP (500 μM) on ORAI1-3 currents evoked by TG (1 μM). (D–F) Effect of 4-CmC (500 μM) on ORAI1-3 currents. (G–I) Effect of 4-ClP (500 μM). (J) Effect of 4-CEP on the non-transfected HEK293 T-REx cells. (K) ORAI3 current induced by 2-APB and the effects of 4-ClP, 4-CmC and 4-CEP (500 μM). (L) % inhibition by 4-ClP, 4-CmC and 4-CEP of the whole-cell currents measured at −80 mV in the STIM1/ORAI1 (S1O1), STIM1/ORAI2 (S1O2), and STIM1/ORAI3 (S1O3) (with TG); and ORAI3 (with 2-APB) cells (n = 5–10 in each group).
Extracellular inhibition of ORAI3 by 4-CEP
To further characterize the pharmacological actions of 4-CEP, two strategies were used to examine the action site of 4-CEP on ORAI3. Similar to our previous findings, the current-voltage curve for the 2-APB-induced ORAI3 current displayed double (inward and outward) rectifications (DeHaven et al., 2008; Peinelt et al., 2008). Firstly, the outside-out patch-clamp recordings were used and the characteristic ORAI3 current evoked by 50 μM 2-APB was inhibited by 500 μM 4-CEP in the external solution (Figure 4A–C). Secondly, the intracellular application of 500 μM 4-CEP in the pipette solution failed to block the activation of ORAI3 by 50 μM 2-APB in the whole-cell configuration, whereas the subsequent addition of 500 μM 4-CEP into the extracellular solution rapidly abolished the currents (Figure 4D–E). These results demonstrate that 4-CEP inhibited ORAI3 activities from the extracellular side of the channels.
Figure 4.
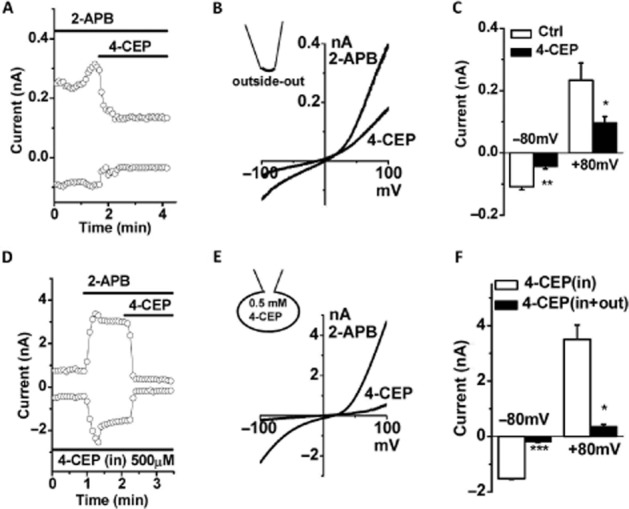
Extracellular effect of 4-CEP on ORAI3 channels. (A) Example of ORAI3 currents inhibited by 4-CEP (500 μM) in the STIM1/ORAI3 cells using outside-out patch configuration. (B) The I/V curves for (A). (C) Mean ± SEM for the current amplitude of outside-out patches (n = 3). (D) Intracellular application of 500 μM 4-CEP in pipette solution did not block the activation of ORAI3 by 2-APB, but extracellular application of 4-CEP (500 μM) significantly inhibited the ORAI3 current. (E) The I/V curves for (D). (F) Mean ± SEM for the amplitude of whole-cell current recorded with intracellular 4-CEP (500 μM) (in) and outside bath perfusion with 4-CEP (out) (n = 3). * P < 0.05, ** P < 0.01 and *** P < 0.001.
Concentration-dependence and reversibility of effects of 4-CEP
The concentration-dependence of the blocking effect of 4-CEP was examined using ORAI3 cells by patch clamp. The whole-cell current of ORAI3 was induced by 50 μM 2-APB. The ORAI3 current was significantly inhibited by 4-CEP and the channel blocking effect was reversible after washout (Figure 5A–B). The inhibition of the ORAI3 channels induced by 4-CEP was concentration-dependent with an IC50 of 71.0 ± 4.2 μM and a slope factor of 0.99 ± 0.04 (Figure 5C).
Figure 5.
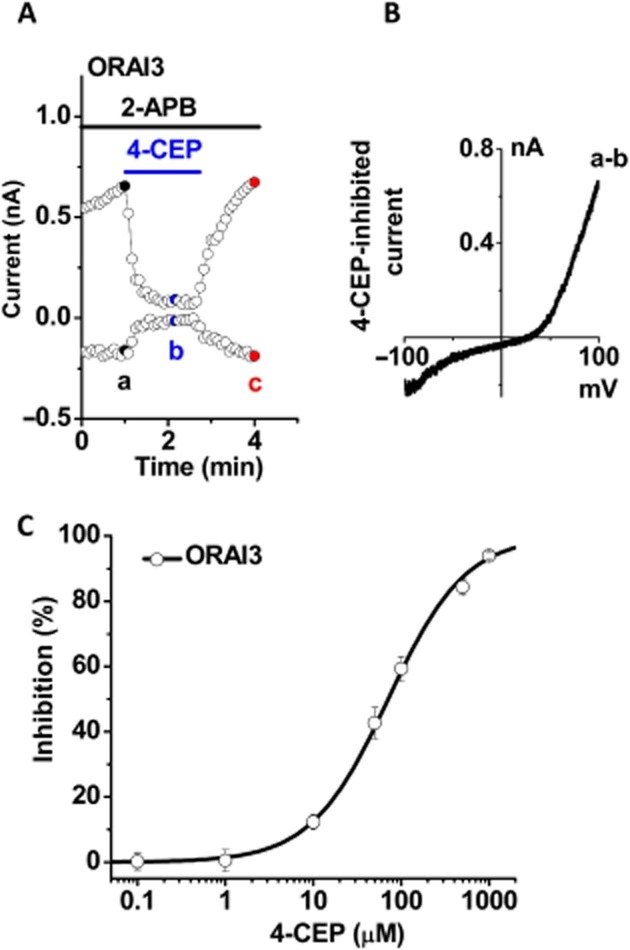
Concentration-dependent inhibition of ORAI3 channels by 4-CEP. The concentration-dependent inhibition by 4-CEP was examined in the HEK293 T-REx cells overexpressing mCFP-ORAI3 using whole-cell patch. (A) Example of ORAI3 current induced by 2-APB (50 μM) and the effect of 4-CEP (500 μM). The inhibition was reversible after washout. (B) The 4-CEP-inhibited I/V curve for (A). (C) Effect of 4-CEP at concentrations ranging from 0.1 to 1000 μM; % inhibition of whole-cell current measured at +80 mV was plotted and the curve was fitted using a logistic model with an IC50 for ORAI3 of 71.0 μM (n = 4–10 recordings for each point).
Discussion
In this study, we have shown that the RyR agonist 4-CEP inhibits the SOCE via extracellular sites of ORAI channels. The inhibitory effect was concentration-dependent, reversible and more potent than its analogues 4-CmC and 4-ClP. The inhibition was also independent of the Ca2+ release induced by activation of RyRs, because the two analogues, 4-CmC and 4-ClP, also inhibit ORAI channels without causing Ca2+ release in the HEK293 cells. In addition, 4-CEP evoked STIM1 translocation and subplasmalemmal clustering through the activation of RyRs and Ca2+ store depletion. However, the STIM1 redistribution in the cytosol was not evident in the STIM1/ORAI co-expressing cells, which could be explained by the stability of STIM1/ORAIs complexes in the plasma membrane. These findings are novel and important for understanding the pharmacology of RyR agonists.
The inhibition of ORAI channels induced by the 4-CEP analogues was structure-dependent. The potency of channel inhibition of the three compounds was positively correlated with the length of the 3-alkyl group, that is 4-CEP (with an ethyl group) was more effective than 4-CmC (with a methyl group) and 4-ClP (no side chain at 3′ position). Elongation of the 3-alkyl chain of 4-CEP may produce analogues with higher potency; however, this has not been investigated in this study due to the availability of the compounds. However, the length of alkyl side chain of chlorophenol derivatives is also associated with their affinity for RyR1 (Beeler and Gable, 1993; Jacobson et al., 2006). Therefore, new compounds with longer alkyl chains may not only be more potent at inhibiting the store-operated channel, but will also strongly interfere with the ER/SR Ca2+ stores via RyR activation. Nonetheless, these structures with direct ORAI channel-blocking activity are important, which provides a new clue for future structure modification to search for more potent and selective compounds, although the potency of 4-CEP (IC50 71 μM for ORAI3) is lower than that of other non-selective store-operated channel blockers, such as 2-APB (Xu et al., 2005), SKF-96365 (Wang et al., 2004), fenamate analogues (Jiang et al., 2012) and BTP2 (Zitt et al., 2004).
Intracellular Ca2+ release induced by 4-CEP and 4-CmC was mediated by the activation of intracellular RyRs, which suggests that the two compounds should be membrane-permeable. Therefore, they may exert their channel-blocking effects from the intracellular side. To determine their action site, we included 4-CEP in the pipette solution using whole-cell patch configuration and found that intracellular application of 4-CEP did not inhibit the ORAI3 channels, but extracellular perfusion with 4-CEP inhibited the current, suggesting that the site of action of 4-CEP is located extracellularly. The experiment using outside-out excised patches further confirmed the extracellular effect of 4-CEP. Furthermore, the blocking effect of 4-CEP on ORAI3 currents could be quickly washed out, which may indirectly indicate that the binding between 4-CEP and the channel proteins could be through non-covalent bonding at the cell surface and thus easily reversed like that of the extracellular blocker 2-APB (Xu et al., 2005).
We also examined STIM1 translocation as an alternative mechanism for SOCE inhibition. 4-CEP may cause cytosolic clustering of STIM1 and this would then lead to an impairment of SOCE, which we demonstrated previously in cells expressing STIM1 alone (Zeng et al., 2012). However, the STIM1/ORAI co-expressing cells had no such cytosolic STIM1 puncta formation, instead they had stable subplasmalemmal cluster accumulation, suggesting that the ratio of STIM1 to ORAI expression in a cell is important and may affect the action of a drug. Nevertheless, the STIM1 clustering effect seems to be unique for 4-CEP, since 4-CmC and 4-ClP do not cause Ca2+ release and STIM1 translocation in the HEK293 cells.
4-CEP and 4-CmC are two commonly used RyR agonists for diagnosis of MH. The RyR agonist-induced skeletal muscle contraction is much stronger in MH susceptible patients with RyR1 mutations than that in the non-susceptible patients (Gerbershagen et al., 2005). Recent findings have shown that SOCE exists in both normal and malignant hyperthermia-susceptible skeletal muscles and the coupling of SR Ca2+ release to SOCE contributes to the sustainability of hypermetabolic reactions (Kurebayashi and Ogawa, 2001; Duke et al., 2010; Launikonis et al., 2010; Stiber and Rosenberg, 2011). The expression of store-operated channel molecules STIM1 and ORAI1 is evident in the skeletal muscles (Lyfenko and Dirksen, 2008; Stiber et al., 2008; Vig et al., 2008). Therefore, the inhibition of Ca2+ influx or SOCE should be considered when the two compounds are used in the in vitro contracture test for the diagnosis of MH, particularly when the Krebs-Ringer solution containing 2.5 mM Ca2+ is recommended in the standard testing procedures (Ording et al., 1997). In order to differentiate the contribution of Ca2+ release and the inhibition of Ca2+ influx by the two agonists, the use of Ca2+-free or low Ca2+ solutions would be a better choice in such muscle contracture test. 4-CEP at the concentrations from 25 to 100 μM can distinguish between the MH susceptible and non-susceptible skeletal muscle samples in the muscle contracture tests, but higher concentrations of 4-CEP, such as 200 μM or higher, are indistinguishable for the diagnosis of MH (Gerbershagen et al., 2005), which could be due to their direct blocking effects on SOCE and lead to changes in intracellular Ca2+ homeostasis.
We conclude that the RyR agonists 4-CEP, 4-CmC and 4-ClP are direct channel inhibitors of ORAI1–3. The use of 4-CEP and 4-CmC in the diagnostic test for MH should be used with caution, as at higher concentrations the two compounds significantly block SOCE, particularly in the presence of extracellular [Ca2+], which may mask their effects on Ca2+ release via RyR activation.
Acknowledgments
This work was supported by British Heart Foundation (PG/08/071/25473) (to S. Z. X.). B. Z. was sponsored by China Scholarship Council. N. D. received 80th anniversary PhD studentship from the University of Hull. We thank Professor AV Tepikin for providing us the STIM1-EYFP construct.
Abbreviations
- 2-APB
2-aminoethoxydiphenyl borate
- 4-CEP
4-chloro-3-ethylphenol
- 4-ClP
4-chlorophenol
- 4-CmC
4-chloro-m-cresol
- EYFP
enhanced yellow fluorescent protein
- mCFP
monomeric cyan fluorescent protein
- MH
malignant hyperthermia
- RyRs
ryanodine receptors
- SOCE
store-operated Ca2+ entry
- SR/ER
sarco/endoplasmic reticulum
- TG
thapsigargin
Conflicts of interest
None.
Supporting information
Additional Supporting Information may be found in the online version of this article at the publisher's web-site: http://dx.doi.org/10.1111/bph.12528
Figure S1 Ca2+ influx in HEK293 T-REx cells and STIM1/ ORAI1 cells. (A) The non-transfected T-REx cells were perfused with free Ca2+ and 1.5 mM Ca2+ solution as indicated by the arrow (n=43 cells). (B) STIM1/ORAI1 cells (n=64 cells). (C) Ca2+ influx in T-REx cells after store depletion with 1 μM TG (n=27 cells). (D) Ca2+ influx in STIM1/ORAI1 cells after store depletion with 1 μM TG (n=50 cells).
Figure S2 Dose-response curves for 4-CEP, 4-CmC and 4-ClP on the inhibition of store-operated Ca2+ influx. The STIM1/ ORAI1 cells were pretreated with 1 μM TG for 30 min and the cytosolic Ca2+ were measured using Ca2+ imaging (n=17–29 cells for each point). The curves were fitted by logistic equation.
Figure S3 Effects of 4-CEP, 4-CmC and 4-ClP on ORAI3 channels activated by 2-APB. The ORAI3 current was induced by 50 μM 2-APB in the T-REx cells expressing mCFP-ORAI3. (A) Example for the effect of 4-CEP (500 μM). (B) IV curves for (A). (C) 4-CmC (500 μM). (D). IV curves for (C). (E) 4-ClP (500 μM). (F) IV curves for (E).
STIM1 cells 4-CEP induced STIM1-EYFP movement in the STIM1 cells. 4-CEP (500 μM) induced STIM1 puncta in the plasma membrane first, and then the accumulation of cytosolic STIM1-EYFP puncta. 4-CEP was added at the time scale 00:01.
STIM1/ORAI1 coexpressed cells 4-CEP induced STIM1-EYFP translocation to the plasma membrane in the cells coexpressing STIM1/ORAI1. ORAI1 protein (red). STIM1 puncta in the plasma membrane was observed after addition of 4-CEP (500 μM), but no cytosolic STIM1-EYFP puncta were formed after perfusion with 4-CEP. 4-CEP was added at the time scale 00:03.
References
- Alexander SPH, Benson HE, Faccenda E, Pawson AJ, Sharman JL, Catterall WA, Spedding M, Peters JA, Harmar AJ CGTP Collaborators. The Concise Guide to PHARMACOLOGY 2013/14: Overview. Br J Pharmacol. 2013;170:1449–1867. doi: 10.1111/bph.12447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baur CP, Bellon L, Felleiter P, Fiege M, Fricker R, Glahn K, et al. A multicenter study of 4-chloro-m-cresol for diagnosing malignant hyperthermia susceptibility. Anesth Analg. 2000;90:200–205. doi: 10.1097/00000539-200001000-00040. [DOI] [PubMed] [Google Scholar]
- Beeler TJ, Gable KS. Activation of Ca2+ release from sarcoplasmic reticulum vesicles by 4-alkylphenols. Arch Biochem Biophys. 1993;301:216–220. doi: 10.1006/abbi.1993.1136. [DOI] [PubMed] [Google Scholar]
- DeHaven WI, Smyth JT, Boyles RR, Bird GS, Putney JW., Jr Complex actions of 2-aminoethyldiphenyl borate on store-operated calcium entry. J Biol Chem. 2008;283:19265–19273. doi: 10.1074/jbc.M801535200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DiJulio DH, Watson EL, Pessah IN, Jacobson KL, Ott SM, Buck ED, et al. Ryanodine receptor type III (Ry3R) identification in mouse parotid acini. Properties and modulation of [3H]ryanodine-binding sites. J Biol Chem. 1997;272:15687–15696. doi: 10.1074/jbc.272.25.15687. [DOI] [PubMed] [Google Scholar]
- Duke AM, Hopkins PM, Calaghan SC, Halsall JP, Steele DS. Store-operated Ca2+ entry in malignant hyperthermia-susceptible human skeletal muscle. J Biol Chem. 2010;285:25645–25653. doi: 10.1074/jbc.M110.104976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Endo M. Calcium-induced calcium release in skeletal muscle. Physiol Rev. 2009;89:1153–1176. doi: 10.1152/physrev.00040.2008. [DOI] [PubMed] [Google Scholar]
- Fellner SK, Arendshorst WJ. Ryanodine receptor and capacitative Ca2+ entry in fresh preglomerular vascular smooth muscle cells. Kidney Int. 2000;58:1686–1694. doi: 10.1046/j.1523-1755.2000.00329.x. [DOI] [PubMed] [Google Scholar]
- Feske S, Gwack Y, Prakriya M, Srikanth S, Puppel SH, Tanasa B, et al. A mutation in Orai1 causes immune deficiency by abrogating CRAC channel function. Nature. 2006;441:179–185. doi: 10.1038/nature04702. [DOI] [PubMed] [Google Scholar]
- Fessenden JD, Perez CF, Goth S, Pessah IN, Allen PD. Identification of a key determinant of ryanodine receptor type 1 required for activation by 4-chloro-m-cresol. J Biol Chem. 2003;278:28727–28735. doi: 10.1074/jbc.M303821200. [DOI] [PubMed] [Google Scholar]
- Fill M, Copello JA. Ryanodine receptor calcium release channels. Physiol Rev. 2002;82:893–922. doi: 10.1152/physrev.00013.2002. [DOI] [PubMed] [Google Scholar]
- Gerbershagen MU, Fiege M, Weisshorn R, Kolodzie K, Schulte am Esch J, Wappler F. Cumulative and bolus in vitro contracture testing with 4-chloro-3-ethylphenol in malignant hyperthermia positive and negative human skeletal muscles. Anesth Analg. 2005;101:710–714. doi: 10.1213/01.ANE.0000160534.23519.1B. [DOI] [PubMed] [Google Scholar]
- Higure Y, Shimazaki Y, Nohmi M. Can 4-chloro-m-cresol be substituted for caffeine as an activator of calcium oscillation in bullfrog sympathetic ganglion cells? Cell Calcium. 2006;39:467–470. doi: 10.1016/j.ceca.2006.01.007. [DOI] [PubMed] [Google Scholar]
- Hopkins PM. Malignant hyperthermia: pharmacology of triggering. Br J Anaesth. 2011;107:48–56. doi: 10.1093/bja/aer132. [DOI] [PubMed] [Google Scholar]
- Islam MS, Leibiger I, Leibiger B, Rossi D, Sorrentino V, Ekstrom TJ, et al. In situ activation of the type 2 ryanodine receptor in pancreatic beta cells requires cAMP-dependent phosphorylation. Proc Natl Acad Sci U S A. 1998;95:6145–6150. doi: 10.1073/pnas.95.11.6145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jacobson AR, Moe ST, Allen PD, Fessenden JD. Structural determinants of 4-chloro-m-cresol required for activation of ryanodine receptor type 1. Mol Pharmacol. 2006;70:259–266. doi: 10.1124/mol.106.022491. [DOI] [PubMed] [Google Scholar]
- Jiang H, Zeng B, Chen GL, Bot D, Eastmond S, Elsenussi SE, et al. Effect of non-steroidal anti-inflammatory drugs and new fenamate analogues on TRPC4 and TRPC5 channels. Biochem Pharmacol. 2012;83:923–931. doi: 10.1016/j.bcp.2012.01.014. [DOI] [PubMed] [Google Scholar]
- Kurebayashi N, Ogawa Y. Depletion of Ca2+ in the sarcoplasmic reticulum stimulates Ca2+ entry into mouse skeletal muscle fibres. J Physiol. 2001;533:185–199. doi: 10.1111/j.1469-7793.2001.0185b.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lanner JT, Georgiou DK, Joshi AD, Hamilton SL. Ryanodine receptors: structure, expression, molecular details, and function in calcium release. Cold Spring Harb Perspect Biol. 2010;2:a003996. doi: 10.1101/cshperspect.a003996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Launikonis BS, Murphy RM, Edwards JN. Toward the roles of store-operated Ca2+ entry in skeletal muscle. Pflugers Arch. 2010;460:813–823. doi: 10.1007/s00424-010-0856-7. [DOI] [PubMed] [Google Scholar]
- Lyfenko AD, Dirksen RT. Differential dependence of store-operated and excitation-coupled Ca2+ entry in skeletal muscle on STIM1 and Orai1. J Physiol. 2008;586:4815–4824. doi: 10.1113/jphysiol.2008.160481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McNally BA, Somasundaram A, Yamashita M, Prakriya M. Gated regulation of CRAC channel ion selectivity by STIM1. Nature. 2012;482:241–245. doi: 10.1038/nature10752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ording H, Brancadoro V, Cozzolino S, Ellis FR, Glauber V, Gonano EF, et al. In vitro contracture test for diagnosis of malignant hyperthermia following the protocol of the European MH Group: results of testing patients surviving fulminant MH and unrelated low-risk subjects. The European Malignant Hyperthermia Group. Acta Anaesthesiol Scand. 1997;41:955–966. doi: 10.1111/j.1399-6576.1997.tb04820.x. [DOI] [PubMed] [Google Scholar]
- Peinelt C, Lis A, Beck A, Fleig A, Penner R. 2-Aminoethoxydiphenyl borate directly facilitates and indirectly inhibits STIM1-dependent gating of CRAC channels. J Physiol. 2008;586:3061–3073. doi: 10.1113/jphysiol.2008.151365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Priori SG, Chen SR. Inherited dysfunction of sarcoplasmic reticulum Ca2+ handling and arrhythmogenesis. Circ Res. 2011;108:871–883. doi: 10.1161/CIRCRESAHA.110.226845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson R, Carpenter D, Shaw MA, Halsall J, Hopkins P. Mutations in RYR1 in malignant hyperthermia and central core disease. Hum Mutat. 2006;27:977–989. doi: 10.1002/humu.20356. [DOI] [PubMed] [Google Scholar]
- Rose CR, Konnerth A. Stores not just for storage. intracellular calcium release and synaptic plasticity. Neuron. 2001;31:519–522. doi: 10.1016/s0896-6273(01)00402-0. [DOI] [PubMed] [Google Scholar]
- Shuttleworth TJ. Orai3 – the ‘exceptional’ Orai? J Physiol. 2012;590:241–257. doi: 10.1113/jphysiol.2011.220574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stiber J, Hawkins A, Zhang ZS, Wang S, Burch J, Graham V, et al. STIM1 signalling controls store-operated calcium entry required for development and contractile function in skeletal muscle. Nat Cell Biol. 2008;10:688–697. doi: 10.1038/ncb1731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stiber JA, Rosenberg PB. The role of store-operated calcium influx in skeletal muscle signaling. Cell Calcium. 2011;49:341–349. doi: 10.1016/j.ceca.2010.11.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Supnet C, Noonan C, Richard K, Bradley J, Mayne M. Up-regulation of the type 3 ryanodine receptor is neuroprotective in the TgCRND8 mouse model of Alzheimer's disease. J Neurochem. 2010;112:356–365. doi: 10.1111/j.1471-4159.2009.06487.x. [DOI] [PubMed] [Google Scholar]
- Tian X, Liu Y, Wang R, Wagenknecht T, Liu Z, Chen SR. Ligand-dependent conformational changes in the clamp region of the cardiac ryanodine receptor. J Biol Chem. 2013;288:4066–4075. doi: 10.1074/jbc.M112.427864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Petegem F. Ryanodine receptors: structure and function. J Biol Chem. 2012;287:31624–31632. doi: 10.1074/jbc.R112.349068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Varadi A, Rutter GA. Dynamic imaging of endoplasmic reticulum Ca2+ concentration in insulin-secreting MIN6 Cells using recombinant targeted cameleons: roles of sarco(endo)plasmic reticulum Ca2+-ATPase (SERCA)-2 and ryanodine receptors. Diabetes. 2002;51(Suppl 1):S190–S201. doi: 10.2337/diabetes.51.2007.s190. [DOI] [PubMed] [Google Scholar]
- Vig M, DeHaven WI, Bird GS, Billingsley JM, Wang H, Rao PE, et al. Defective mast cell effector functions in mice lacking the CRACM1 pore subunit of store-operated calcium release-activated calcium channels. Nat Immunol. 2008;9:89–96. doi: 10.1038/ni1550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang J, Shimoda LA, Sylvester JT. Capacitative calcium entry and TRPC channel proteins are expressed in rat distal pulmonary arterial smooth muscle. Am J Physiol Lung Cell Mol Physiol. 2004;286:L848–L858. doi: 10.1152/ajplung.00319.2003. [DOI] [PubMed] [Google Scholar]
- Westerblad H, Andrade FH, Islam MS. Effects of ryanodine receptor agonist 4-chloro-m-cresol on myoplasmic free Ca2+ concentration and force of contraction in mouse skeletal muscle. Cell Calcium. 1998;24:105–115. doi: 10.1016/s0143-4160(98)90078-1. [DOI] [PubMed] [Google Scholar]
- Xu SZ, Zeng F, Boulay G, Grimm C, Harteneck C, Beech DJ. Block of TRPC5 channels by 2-aminoethoxydiphenyl borate: a differential, extracellular and voltage-dependent effect. Br J Pharmacol. 2005;145:405–414. doi: 10.1038/sj.bjp.0706197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yarotskyy V, Dirksen RT. Temperature and RyR1 regulate the activation rate of store-operated Ca2+ entry current in myotubes. Biophys J. 2012;103:202–211. doi: 10.1016/j.bpj.2012.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zeng B, Chen GL, Xu SZ. Store-independent pathways for cytosolic STIM1 clustering in the regulation of store-operated Ca2+ influx. Biochem Pharmacol. 2012;84:1024–1035. doi: 10.1016/j.bcp.2012.07.013. [DOI] [PubMed] [Google Scholar]
- Zhang SL, Yu Y, Roos J, Kozak JA, Deerinck TJ, Ellisman MH, et al. STIM1 is a Ca2+ sensor that activates CRAC channels and migrates from the Ca2+ store to the plasma membrane. Nature. 2005;437:902–905. doi: 10.1038/nature04147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zitt C, Strauss B, Schwarz EC, Spaeth N, Rast G, Hatzelmann A, et al. Potent inhibition of Ca2+ release-activated Ca2+ channels and T-lymphocyte activation by the pyrazole derivative BTP2. J Biol Chem. 2004;279:12427–12437. doi: 10.1074/jbc.M309297200. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1 Ca2+ influx in HEK293 T-REx cells and STIM1/ ORAI1 cells. (A) The non-transfected T-REx cells were perfused with free Ca2+ and 1.5 mM Ca2+ solution as indicated by the arrow (n=43 cells). (B) STIM1/ORAI1 cells (n=64 cells). (C) Ca2+ influx in T-REx cells after store depletion with 1 μM TG (n=27 cells). (D) Ca2+ influx in STIM1/ORAI1 cells after store depletion with 1 μM TG (n=50 cells).
Figure S2 Dose-response curves for 4-CEP, 4-CmC and 4-ClP on the inhibition of store-operated Ca2+ influx. The STIM1/ ORAI1 cells were pretreated with 1 μM TG for 30 min and the cytosolic Ca2+ were measured using Ca2+ imaging (n=17–29 cells for each point). The curves were fitted by logistic equation.
Figure S3 Effects of 4-CEP, 4-CmC and 4-ClP on ORAI3 channels activated by 2-APB. The ORAI3 current was induced by 50 μM 2-APB in the T-REx cells expressing mCFP-ORAI3. (A) Example for the effect of 4-CEP (500 μM). (B) IV curves for (A). (C) 4-CmC (500 μM). (D). IV curves for (C). (E) 4-ClP (500 μM). (F) IV curves for (E).
STIM1 cells 4-CEP induced STIM1-EYFP movement in the STIM1 cells. 4-CEP (500 μM) induced STIM1 puncta in the plasma membrane first, and then the accumulation of cytosolic STIM1-EYFP puncta. 4-CEP was added at the time scale 00:01.
STIM1/ORAI1 coexpressed cells 4-CEP induced STIM1-EYFP translocation to the plasma membrane in the cells coexpressing STIM1/ORAI1. ORAI1 protein (red). STIM1 puncta in the plasma membrane was observed after addition of 4-CEP (500 μM), but no cytosolic STIM1-EYFP puncta were formed after perfusion with 4-CEP. 4-CEP was added at the time scale 00:03.