

MLLT10 (also known as AF10), at chromosome 10 band p12, is emerging as a promiscuous gene. Six partners have been reported to date: PICALM(CALM)/11q14, MLL/11q23, NAP1L1/12q21, HNRNPH1/5q35, DDX3X/Xp11.3 and NUP98/11p15.1,2 All fusions retain the MLLT10 octapeptide motif-leucine-zipper (OM-LZ) domain which induces acute myeloid leukemia in mouse models, suggesting it is critical for leukemogenesis. In pediatric T-cell acute lymphoblastic leukemia (T-ALL) PICALM-MLLT10, HNRNPH1-MLLT10 and DDX3X-MLLT10 fusions shared a specific gene expression profile signature which included NK-like homeobox overexpression, differentiating it from all other genomic rearrangements in the HOXA category.1
Focusing on MLLT10 involvement in adult T-ALL, we used fluorescence in situ hybridization (FISH) to investigate 99 patients (31 females, 68 males; age range 14–69 years, median age 34) who were enrolled in 2 consecutive multicenter GIMEMA (Gruppo Italiano Malattie Ematologiche dell’Adulto) studies (protocols 0904 and 0496). All patients provided informed consent for sample collection. Biological analyses were carried out in accordance with the Declaration of Helsinki. Screening studies were approved by the Institutional Review Board of “Sapienza” University, Rome, Italy.
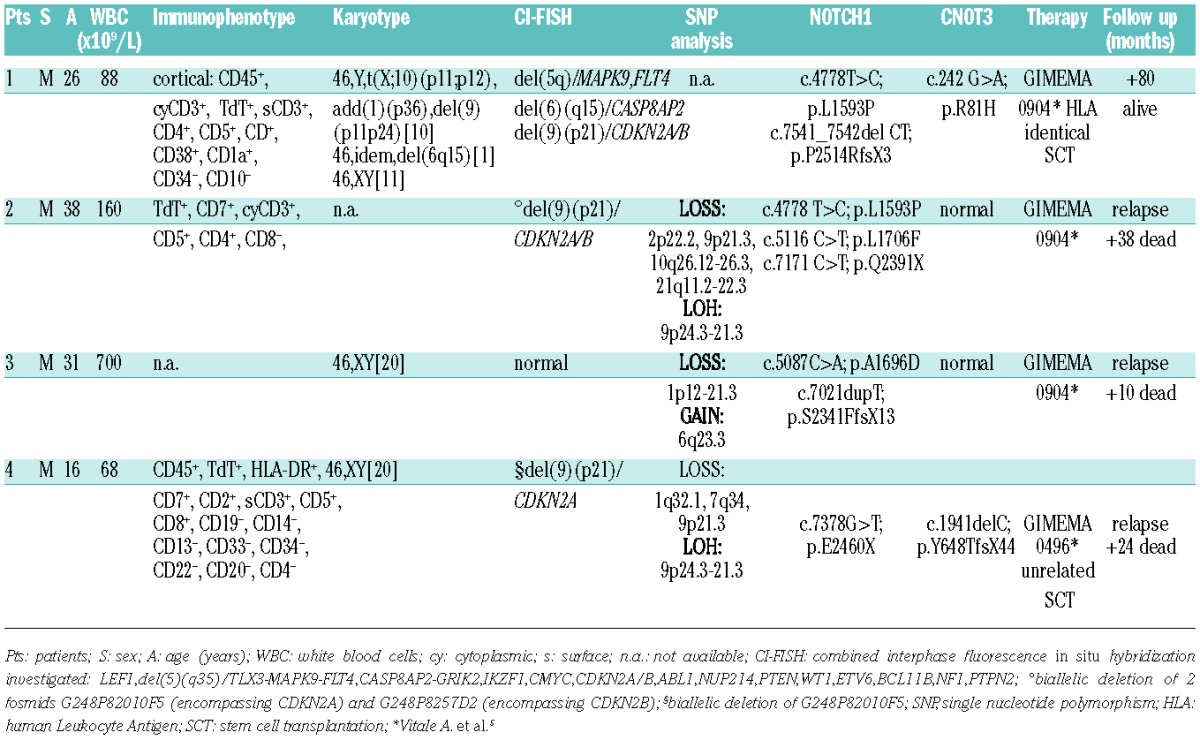
In these 99 patients, the overall incidence of MLLT10 translocations was approximately 10% (10 of 99 patients). FISH investigated for known MLLT10 partners,1,3 finding 7 patients with PICALM-MLLT10 and 3 with DDX3X-MLLT10. A biological sample from a 4th patient was obtained when a parallel study using whole transcriptome sequencing on a Illumina HiSeq2000 (Illumina, San Diego, CA, USA) instrument identified an additional case harboring the DDX3X-MLLT10 fusion transcript in a different cohort of 20 adult T-ALL patients.4 Table 1 reports clinical, hematologic and cytogenetic findings in all 4 DDX3X-MLLT10 positive T-ALL patients. All were males presenting with high white blood cells count. In case 1, T-ALL was arrested at cortical stage as previously described in a child.1 In the other 3 patients, immunophenotype was incomplete or not available. All achieved hematologic remission but 3 relapsed and died. One patient (n. 1) is alive and well at 80 months post-HLA identical stem cell transplantation (HLA-SCT).
Table 1.
Clinical, hematologic and cytogenetic findings in all 4 DDX3X-MLLT10 positive T-ALL patients.
A DDX3X-MLLT10 double color double fusion FISH assay detected balanced translocations in 3 cases and unbalanced in 1 (Figure 1A–C). Reverse-transcription polymerase chain reaction (Thermoscript RT-PCR System, Invitrogen, Carlsbad, CA, USA) and sequencing (AB3500 Genetic analyzer, Applied Biosystem, Foster City, CA, USA) confirmed in-frame DDX3X-MLLT10 transcripts in all patients but breakpoints differed from our previously described pediatric case. DDX3X exon 6 fused to MLLT10 exon 6 in Patient 1 and to exon 4 in Patient 4; DDX3X exon 3 fused to MLLT10 exon 17 in Patient 2 and DDX3X exon 4 fused to MLLT10 exon 7 in Patient 3 (Figure 1D). An additional splicing variant joining DDX3X intron 6–7 to MLLT10 intron 5–6 was identified by RNA-seq in Patient 1. A reciprocal DDX3X-MLLT10 in-frame fusion was detected only in Patients 1 and 2, supporting the hypothesis that DDX3X-MLLT10 is critical in the pathogenesis of this T-ALL subgroup.1
Figure 1.
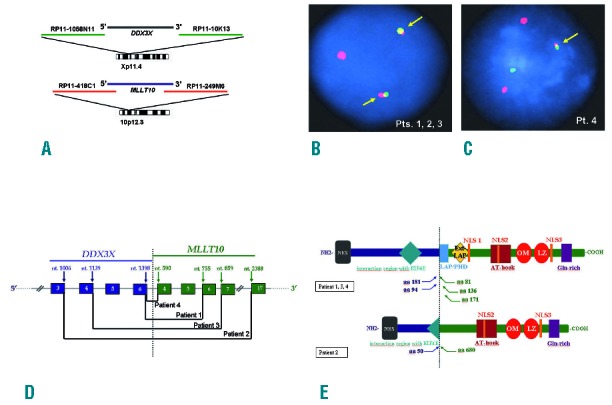
Cytogenetic and molecular characterization of DDX3X-MLLT10 fusions. (A) Double color double fusion FISH assay for DDX3X and MLLT10. (B) FISH showed 2 fused signals in Patients 1, 2 and 3 indicating a balanced translocation (arrows). (C) FISH showed one fused signal in patient 4 indicating an unbalanced translocation (arrow). (D) Schematic representation of DDX3X and MLLT10 breakpoints in the 4 DDX3X-MLLT10 positive cases (arrows). Nucleotide numbers refer to GenBank accession: NM_001356.3 for DDX3X and NM_004641.3 for MLLT10. (E) Putative fusion protein structure. At N terminal DDX3X retained a NES domain in all. Three patients retained the entire EIF4E interacting motif and 1 only half. At C terminal at least 1 NLS, the AT-hook and the OM-LZ domain were retained in all. Pt: patient; Pts: patients; nt.: nucleotide; aa: amino acid; NES: Nuclear Exporting Signal; NLS: Nuclear Localization Signal; LAP/PHD: Leukemia Associated Protein / Plant Homeo Domain; Ext-LAP: Extended LAP; OM-LZ: Octapeptide Motif-Leucine Zipper; Gln: Glutamine.
The MLLT10 leukemogenic OM-LZ domain and at least one nuclear localization signal were maintained at the C-terminal in all fusions (Figure 1E). At the N-terminal, DDX3X retained a nuclear export signal (NES) domain, which interacts with CRM1, and an EIF4E interacting motif that is required for DDX3X modulation of translation. The contributions of these domains to DDX3X-MLLT10 leukemogenesis remains to be established. The PICALM CRM1-dependent NES was shown to play a major role in the onset of PICALM-MLLT10 positive leukemias. Nuclear export of PICALM-MLLT10 mislocalized a DOT1L H3K79 methyltransferase fraction to outside the nucleus, with loss of H3K79 methylation overall except for critical genes such as HOXA which were hypermethylated at lysine 79 and up-regulated.6 Interestingly, a potent and selective inhibitor of DOT1L is under evaluation to enter human clinical trials as a target therapy for acute leukemias bearing MLL translocations.7
Finding DDX3X in these 4 patients with T-ALL ranked it as the second most frequent MLLT10 partner. DDX3X, an ubiquitously expressed gene, belongs to the adenosine 5′-triphosphate-dependent DEAD box RNA helicases family and has been recurrently involved in solid and hematologic tumors. DDX3X is located at Xp11.3 and it is one of the genes that escapes X-inactivation in females.8 As all patients with DDX3X-MLLT10 positive T-ALL (the 4 adults in this study and 1 child previously described)1 were males, no wild-type DDX3X allele was retained in the leukemic blasts, suggesting that the complete absence of a normally functional DDX3X protein might contribute to leukemogenesis. DDX3X appeared to have oncogenic as well as tumor suppressor functions.9,10 DDX3X somatic mutations have recently been discovered in medulloblastoma,11,12 chronic lymphocytic leukemia,13 and Burkitt lymphoma.14 Recurrent DDX3X homozygous deletions were identified in gingivobuccal oral squamous cell carcinoma.15
To identify concurrent molecular hits in our DDX3X-MLLT10 positive T-ALL, Combined interphase-FISH for recurrent T-ALL associated rearrangements,3 SNP array and Sanger sequencing for NOTCH1 and CNOT3 were performed (Table 1). NOTCH1 mutation was common to all. CDKN2A/B deletions and CNOT3 mutations appeared to be accompanying recurrent events in 3 and 2 cases, respectively. CNOT3, a putative tumor suppressor gene, has been recently reported to be mutated in approximately 8% of adult T-ALL.16 Other genomic imbalances, as well as copy neutral loss of heterozygosity, were identified in individual cases.
In our study, longitudinal molecular studies were conducted only in Patients 3 and 4 because of lack of biological material in the others. Both patients achieved hematologic remission, but the DDX3X-MLLT10 fusion persisted after consolidation in Patient 3, and during maintenance in Patient 4 using a nested PCR.
In conclusion, we report for the first time that DDX3X-MLLT10 occurs in approximately 3% of adult T-ALL and characterizes a subgroup of NOTCH1 positive leukemias. The DDX3X-MLLT10 fusion behaved as a primary abnormality and occurred alternatively to rearrangements of other T-cell oncogenes, such as TAL1, TAL2, LMO1, LMO2, TLX1, TLX3, and NKX2-1. CDKN2A/B deletions and CNOT3 alterations were frequent co-operating hits. DDX3X-MLLT10 appeared to be a stable and reliable molecular marker for monitoring residual disease. Although its prognostic impact can only be assessed in a much larger cohort of patients, it is worth noting the only long-term survivor underwent HLA-SCT. Diagnosis of MLLT10 positive leukemias will be helpful to select candidates to target therapy with DOT1L inhibitor.
Acknowledgments
The authors would like to thank Dr. Geraldine Boyd for assistance in preparing the manuscript; Prof F. Di Raimondo (Department of Biomedical Sciences, Hematology Section, University of Catania, Catania, Italy) for providing clinical data of Patient 1; Dr. Zeynep Kalender Atak and Prof. Stein Aerts (Laboratory of Computational Biology, Center for Human Genetics, KU Leuven, Leuven, Belgium) for the bioinformatic analysis of RNA-seq data in Patient 1.
Footnotes
Funding: the work was partially supported by Associazione Italiana per la Ricerca sul Cancro (AIRC, IG-11512 to CM, IG-10612 to AB), Fondo per gli Investimenti della Ricerca di Base (FIRB 2011 RBAP11TF7Z_005 to CM and AIRC Special Program Molecular Clinical Oncology, 5 × 1000, Milan, Italy to VG), Ministero dell’Università e della Ricerca Scientifica (PRIN 2010–2011), Fondazione Cassa di Risparmio di Perugia (Cod. 2012.0108.021 Ricerca scientifica e tecnologica), Interuniversity Attraction Pole (IAP), Belgium. LB is supported by a grant from Beat Leukemia ONLUS and Società Italiana di Ematologia Sperimentale (SIES).
Information on authorship, contributions, and financial & other disclosures was provided by the authors and is available with the online version of this article at www.haematologica.org.
References
- 1.Brandimarte L, Pierini V, Di Giacomo D, Borga C, Nozza F, Gorello P, et al. New MLLT10 gene recombinations in pediatric T-acute lymphoblastic leukemia. Blood. 2013;121(25):5064–7 [DOI] [PubMed] [Google Scholar]
- 2.Soler G, Kaltenbach S, Dobbelstein S, Broccardo C, Radford I, Mozziconacci MJ, et al. Identification of GSX2 and AF10 as NUP98 partner genes in myeloid malignancies. Blood Cancer J. 2013;3:e124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.La Starza R, Lettieri A, Pierini V, Nofrini V, Gorello P, Songia S, et al. Linking genomic lesions with Minimal Residual Disease improves prognostic stratification in children with T-cell acute lymphoblastic leukaemia. Leukemia Res. 2013;37(8):929–35 [DOI] [PubMed] [Google Scholar]
- 4.Atak ZK, Gianfelici V, Hulselmans G, De Keersmaecker K, Devasia AG, Geerdens E, et al. Comprehensive analysis of transcriptome variation uncovers known and novel driver events in T-cell acute lymphoblastic leukemia. PLoS Genet. 2013;9(12):e1003997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Vitale A, Guarini A, Ariola C, Mancini M, Mecucci C, Cuneo A, et al. Adult T-cell acute lymphoblastic leukemia: biologic profile at presentation and correlation with response to induction treatment in patients enrolled in the GIMEMA LAL 0496 protocol. Blood. 2006; 107(2):473–9 [DOI] [PubMed] [Google Scholar]
- 6.Conway AE, Scotland PB, Lavau CP, Wechsler DS. A CALM-derived nuclear export signal is essential for CALM-AF10-mediated leukemogenesis. Blood. 2013;121(23):4758–68 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Daigle SR, Olhava EJ, Therkelsen CA, Basavapathruni A, Jin L, Boriack-Sjodin PA, et al. Potent inhibition of DOT1L as treatment of MLL-fusion leukemia. Blood. 2013;122(6):1017–25 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Lahn BT, Page DC. Functional coherence of the human Y chromosome. Science. 1997; 278(5338):675–80 [DOI] [PubMed] [Google Scholar]
- 9.Chang PC, Chi CW, Chau GY, Li FY, Tsai YH, Wu JC, et al. DDX3, a DEAD box RNA helicase, is deregulated in hepatitis virus-associated hepatocellular carcinoma and is involved in cell growth control. Oncogene. 2006;25(14):1991–2003 [DOI] [PubMed] [Google Scholar]
- 10.Botlagunta M, Vesuna F, Mironchik Y, Raman A, Lisok A, Winnard P, Jr, et al. Oncogenic role of DDX3 in breast cancer biogenesis. Oncogene. 2008;27(28):3912–22 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Robinson G, Parker M, Kranenburg TA, Lu C, Chen X, Ding L, et al. Novel mutations target distinct subgroups of medulloblastoma. Nature. 2012;488(7409):43–8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Jones DT, Jäger N, Kool M, Zichner T, Hutter B, Sultan M, et al. Dissecting the genomic complexity underlying medulloblastoma. Nature. 2012;488(7409):100–5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Wang L, Lawrence MS, Wan Y, Stojanov P, Sougnez C, Stevenson K, et al. SF3B1 and other novel cancer genes in chronic lymphocytic leukemia. N Engl J Med. 2011;365(26):2497–506 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Schmitz R, Young RM, Ceribelli M, Jhavar S, Xiao W, Zhang M, et al. Burkitt lymphoma pathogenesis and therapeutic targets from structural and functional genomics. Nature. 2012;490(7418):116–20 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.India Project Team of the International Cancer Genome Consortium. Mutational landscape of gingivo-buccal oral squamous cell carcinoma reveals new recurrently-mutated genes and molecular subgroups. Nat Comm. 2013;4:2873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.De Keersmaecker K, Atak ZK, Li N, Vicente C, Patchett S, Girardi T, et al. Exome sequencing identifies mutation in CNOT3 and ribosomal genes RPL5 and RPL10 in T-cell acute lymphoblastic leukemia. Nat Genet. 2013;45(2):186–90 [DOI] [PMC free article] [PubMed] [Google Scholar]