

Abstract
Background
One of the most striking features of avian vision is the variation in spectral sensitivity of the short wavelength sensitive (SWS1) opsins, which can be divided into two sub-types: violet- and UV- sensitive (VS & UVS). In birds, UVS has been found in both passerines and parrots, groups that were recently shown to be sister orders. While all parrots are thought to be UVS, recent evidence suggests some passerine lineages may also be VS. The great bowerbird (Chlamydera nuchalis) is a passerine notable for its courtship behaviours in which males build and decorate elaborate bower structures.
Results
The great bowerbird SWS1 sequence possesses an unusual residue combination at known spectral tuning sites that has not been previously investigated in mutagenesis experiments. In this study, the SWS1 opsin of C. nuchalis was expressed along with a series of spectral tuning mutants and ancestral passerine SWS1 pigments, allowing us to investigate spectral tuning mechanisms and explore the evolution of UV/violet sensitivity in early passerines and parrots. The expressed C. nuchalis SWS1 opsin was found to be a VS pigment, with a λmax of 403 nm. Bowerbird SWS1 mutants C86F, S90C, and C86S/S90C all shifted λmax into the UV, whereas C86S had no effect. Experimentally recreated ancestral passerine and parrot/passerine SWS1 pigments were both found to be VS, indicating that UV sensitivity evolved independently in passerines and parrots from a VS ancestor.
Conclusions
Our mutagenesis studies indicate that spectral tuning in C. nuchalis is mediated by mechanisms similar to those of other birds. Interestingly, our ancestral sequence reconstructions of SWS1 in landbird evolution suggest multiple transitions from VS to UVS, but no instances of the reverse. Our results not only provide a more precise prediction of where these spectral sensitivity shifts occurred, but also confirm the hypothesis that birds are an unusual exception among vertebrates where some descendants re-evolved UVS from a violet type ancestor. The re-evolution of UVS from a VS type pigment has not previously been predicted elsewhere in the vertebrate phylogeny.
Keywords: Opsins, Ultraviolet, Bird vision, Visual pigment evolution
Background
Bowerbirds are a remarkable group of passerine birds in which males build elaborate structures of plant material adorned with coloured objects to attract females. These displays are among the most striking examples of sexually selected traits. Consequently, bowerbirds have become a model system in visual ecology and evolutionary biology, particularly with respect to the evolution of visual signals [1-6]. Birds have a visual system well suited for colour detection with four types of cone visual pigments that span a wide range of the electromagnetic spectrum extending into the ultraviolet (UV). UV based signals in particular can play important roles in avian behaviours [7-9], especially in mate choice in passerines [10,11] and parrots [12].
The first step in vision is the absorption of light by visual pigments in the photoreceptor cells of the retina. Visual pigments consist of an opsin protein covalently bound to a light sensitive chromophore via a Schiff base (SB) link. Absorption of a photon of light triggers a cis-trans isomerization in the chromophore that induces subsequent conformational changes in the opsin protein. This change allows the visual pigment to bind and activate the downstream heterotrimeric G-protein, transducin, thus initiating the visual transduction cascade in the photoreceptor cell [13]. The wavelength of maximal absorbance of a visual pigment (λmax) is determined by the interactions between the opsin protein and its chromophore, via a process known as spectral tuning [14].
The short-wavelength-sensitive (SWS1) pigments mediate sensitivity to light in the violet to UV range. This group of pigments exhibits the broadest range in spectral sensitivity across vertebrates, and are generally divided into two groups based on λmax: violet-sensitive (VS: λmax 388–435 nm) and UV-sensitive (UVS: λmax 355–380 nm) [15]. In SWS1 pigments, spectral tuning mechanisms can be quite complicated, and can differ across vertebrate pigments [16-23]. However, among vertebrates, SWS1 spectral tuning mechanisms in birds appear to be fairly unique and unusually straightforward. Mutagenesis studies in a variety of birds indicate the most important site is 90, with mutations at this site responsible for determining whether a pigment absorbs maximally in the violet or UV [17,18,21,24]. Phenylalanine (F) at site 86 appears to be a second mechanism by which birds achieve UVS because it is found in the SWS1 genes of some birds [25-27], and site-directed mutagenesis studies indicate that it can blue shift wavelength sensitivity in some avian VS-type SWS1 pigments [28] as well as in other vertebrates [16,19,29,30], with the exception of some primates [23]. However, the paucity of mutagenesis studies on SWS1 pigments throughout the diverse avian orders somewhat limits our abilities to extrapolate upon the roles of spectral tuning sites across all birds.
Here, we use site-directed mutagenesis and ancestral reconstruction methods in order to characterize the absorption spectra of ancestral passerine/parrot SWS1 pigments, and to investigate SWS1 spectral tuning mechanisms using the great bowerbird pigment as a model system. Until recently, the parrots and passerines were thought to be divergent orders within landbirds, but in fact have been found to be sister groups in a number of recent studies [31-33], though this relationship is not always recovered [34,35]. The relationship between passerines and parrots is relevant to understanding the evolution of UV/violet vision in birds because both groups are thought to contain UVS due to the presence of C90 [17,36-38], raising the question of when UV sensitivity may have arisen in these groups. Recent results indicate some basal songbird lineages may have VS pigments [39,40] and in fact, a variety of other basal passerine lineages including some flycatchers have also been found to possess S90, suggestive of VS pigments [41]. As one of the basal passerine lineages whose ecology and behaviour have been the subject of detailed study, the great bowerbird (Chlamydera nuchalis) provides an ideal system with which to study the function and evolution of avian vision. In this study we not only isolate and characterize the SWS1 pigment from C. nuchalis as a VS-type opsin, we also explore the function and evolution of recreated ancestral SWS1 pigments in passerines and parrots. We present experimental evidence indicating that although passerines and parrots evolved UVS by the same molecular mechanism, the passerine ancestor and parrot/passerine ancestor both had VS-type pigments, indicating UVS evolved independently in these two groups. We also investigate spectral tuning mutants of C. nuchalis SWS1, finding that λmax is affected similarly by the mutations C86S, C86F and S90C as in other avian SWS1 opsins, suggesting spectral tuning in avian SWS1 pigments is unusually consistent compared to other vertebrate groups.
Results
Great bowerbird SWS1 spectral tuning mutants
The sequenced C. nuchalis SWS1 gene was found to contain amino acid residues C86 and S90, a combination found in past sequencing-surveys of avian SWS1 opsins [41,42], but one that has not been investigated in any in vitro expression and mutagenesis experiments. The expressed wild type bowerbird pigment was found to have a VS-type absorption spectrum (λmax = 403 nm, Figure 1). This lies within the range of other expressed VS-type SWS1 avian opsins [17,18,28,43]. Mutating S90C in bowerbird SWS1 resulted in a UVS pigment (363 nm), with a 40 nm blue shift relative to wild type (Figure 2A). A similar effect was found with the C86F mutant, which also resulted in a UVS pigment (365 nm, Figure 2B). However, the mutation C86S had no effect (Figure 2C). The double mutant C86S/S90C had a λmax at 363 nm, identical to the S90C single mutant (Figure 2D, Table 1). Homology modeling studies of bowerbird SWS1 structure confirm that there are only minor differences in side chain orientation for both C86S and S90C; however for C86F there is a large difference in side chain orientation, with F much closer to the protonated Schiff base (Figure 3).
Figure 1.
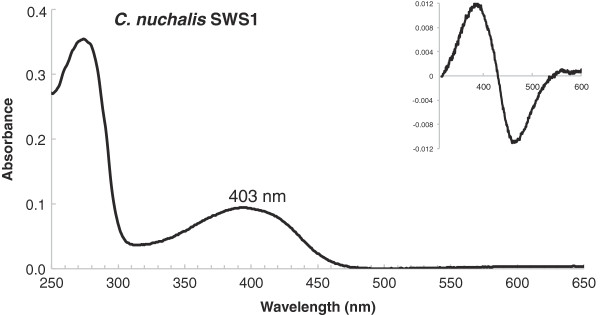
UV-visible dark absorption spectrum of the wild type C. nuchalis SWS1. Estimated absorption maximum values (λmax) noted above the dark spectrum. Inset, Dark-minus-acid bleached difference spectra.
Figure 2.

UV-visible dark absorption spectra of C. nuchalis SWS1 mutants. Dark spectra of (A) S90C, (B) C86F, (C) C86S, and (D) double mutant C86S/S90C, all recorded at pH 6.6. Insets show dark-minus-acid difference spectra. Estimated λmax values indicated for each mutant.
Table 1.
Spectral absorbance characteristics for wild type C. nuchalis SWS1 pigments, site-directed mutants, and ancestral pigments
| Pigment | λ max a (nm) | Shift from C. nuchalis wt pigment b (nm) |
|---|---|---|
| GBS1 wt |
402.97 ± 0.22 |
|
| S90C |
363.05 ± 0.05 |
-40 |
| C86S |
403.05 ± 0.12 |
0 |
| C86Fc |
365.72 ± 1.10 |
-37 |
| C86S/S90C |
362.95 ± 0.26 |
-40 |
| Parrot/Passerine Ancestor |
402.93 ± 0.28 |
0 |
| Passerine Ancestor | 404.28 ± 0.28 | 0 |
a λmax values are given as mean ± standard deviation from at least three different measurements of dark absorbance spectra per expression. b λmax shifts from C. nuchalis wild type (wt) pigment are expressed as negative for blue shifts. c λmax of single mutant C86F calculated from fitting difference spectra of dark and acid denatured species.
Figure 3.

Homology modeling of C. nuchalis SWS1. Models are based on the bovine rhodopsin template, comparing the wild type structure with mutations (A) C86F, (B) C86S, and (C) S90C. Wild type residues are indicated in black, mutant residues in red. The 11-cis retinal chromphore is indicated in purple; with K296 in light blue, the site of chromophore attachment via a protonated Schiff base linkage. Estimated distances to the protonated Schiff base are indicated along the dotted lines.
Due to the short-wavelength λmax of the bowerbird SWS1 and its mutants, a number of assays were performed in order to demonstrate a properly folded protein with bound 11-cis retinal chromophore, and to further characterize its function. In order to demonstrate a covalently bound chromophore, SWS1 pigments regenerated with 11-cis retinal were denatured in HCl, producing absorbance peaks shifted to 440 nm (Figures 1 &2, inset), characteristic of denatured opsin bound to chromophore [44]. All SWS1 pigments with λmax above 400 nm were bleached with light to ~380 nm, characteristic of the biologically active state of visual pigments, metarhodopsin II [45,46]. Finally, the wild type bowerbird SWS1 pigment was found to react in the presence of hydroxylamine (Figure 4), with a t1/2 = ~6 min, typical of cone pigments [47,48].
Figure 4.
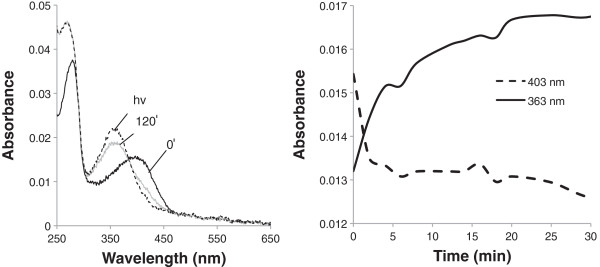
Hydroxylamine reactivity of the C. nuchalis wild type SWS1 pigment. Absorption spectra recorded t = 0 min after hydroxylamine addition (black line), and t = 120 min (grey line), followed by light bleaching (broken line). Right: The absorbance values at 403 nm (broken line) and 363 nm (black line) were plotted as a function of time after addition of hydroxylamine. Half-life for the formation of the retinal oxime in the presence of hydroxylamine was obtained by fitting the plot to a single exponential function.
Some of the pigments were occasionally found to have small secondary absorbance peaks in the longwave arm of the curve, which can have the effect of broadening pigment absorbance curves. These have previously been observed in wild type and mutant SWS1 pigments expressed in solution [17,18,20,24,49-51]. This has also been observed in blue shifted RH1 mutants with mutations at site 90 [51-53]. In this study, experimental attempts to narrow the absorption spectra, including the use of TRIS phosphate buffers, exclusion of glycerol, decreasing purification time and minimizing light and temperature exposure, were unsuccessful, similar to previous experimental studies [18,50,51,54].
In addition to SWS1, four other opsin genes were also isolated from C. nuchalis: SWS2, RH2, and LWS, and rod opsin (RH1) (Additional file 1: Figure S2). All opsin genes were found to contain important structural characteristics typical of functional visual pigments. Phylogenetic analyses show these sequences cluster with expected visual pigment families (Additional file 1: Figure S3).
Reconstructing passerine and parrot/passerine ancestral SWS1 pigments
In order to investigate the evolution of UV sensitivity in passerines and parrots, a combination of Bayesian and maximum likelihood ancestral reconstruction methods were used to infer the sequence of Helix 2 of SWS1 in the ancestors of passerines and parrots (Additional file 1: Table S4). Reconstructed amino acid substitutions at major spectral tuning sites were mapped on a landbird phylogeny (Figure 5). Relative to site 90, less variation was found at sites 86 and 93, with a notable substitution, S86C, occurring at the base of the passerine lineage. Interestingly, substitutions at site 90 were found to occur multiple times throughout the passerine phylogeny, and always involve a change from S to C, suggestive of multiple shifts towards UV sensitivity (but not the reverse). This finding is in disagreement with a previous study proposing that the residue at site 90 has transitioned back and forth between S and C multiple times throughout passerine evolution [41]. Their results would suggest that transitions between UV and violet pigments are quite labile, whereas our results would imply more constrained evolution.
Figure 5.

SWS1 visual pigment evolution, with ancestrally reconstructed substitutions at sites 86, 90 & 93 mapped on a phylogeny of Landbirds [31-33,40,56,80-87]. Experimentally reconstructed ancestral nodes are shown along with measured λmax values. GenBank accession numbers provided in Additional file 1: Table S2.
SWS1 pigments for the ancestors of the passerines and parrots were experimentally recreated in the background of our C. nuchalis pigment. This was done for a number of reasons. First, we were limited by current sequence data, which only exists for Helix 2 for most bird SWS1 genes, as all known spectral tuning sites are thought to be contained in this helix. Second, as a basal passerine, C. nuchalis SWS1 differed from the reconstructed ancestral sequences at specific sites in Helix 2, allowing us to generate the ancestrally reconstructed sequences using site-directed mutagenesis methods. Third, our ability to make direct functional comparisons between the ancestral pigments and that of C. nuchalis allowed us to better interpret the effects of particular amino acid substitutions. The experimentally assayed, recreated ancestral SWS1 pigments were both found to be VS pigments, absorbing maximally in the VS at 403 nm (parrot/passerine ancestor) and 404 nm (passerine ancestor, Figure 6). Both ancestral pigments were found not only to bind retinal, but also to activate in response to light and denature in acid (Figure 6, inset). The reconstructed nodes had high posterior probability values across sites (Additional file 1: Table S4). Reconstructions on an alternate topology favored by previous visual pigment studies [41] did not find any differences with our experimentally recreated sequences (Additional file 1: Figure S4).
Figure 6.

UV-visible dark absorption spectra of the (A) ancestral SWS1 pigment of passerines and parrots and (B) ancestral SWS1 pigment of passerines. Absorption maxima (λmax) noted above the dark spectra. Inset, dark-minus-acid bleached difference spectra.
Discussion
This study extends our understanding of SWS1 opsin function and evolution by investigating evolutionary changes that occurred in avian SWS1 genes. The SWS1 opsin of the great bowerbird C. nuchalis, a basal passerine bird, was expressed along with a series of spectral tuning mutants and ancestral passerine SWS1 pigments allowing us to investigate spectral tuning mechanisms and identify the evolution of UV/violet sensitivity in early passerines and parrots. The C. nuchalis SWS1 opsin was found to be a VS pigment, with a maximal absorbance of 403 nm, which is in agreement with previous MSP studies identifying a λmax of 404 nm [39]. However, our experimentally recreated passerine ancestral SWS1 pigments were also found to be VS, addressing a longstanding issue of ancestral passerine SWS1 spectral tuning in previous studies [25,28,41,55].
Evolution of UV/violet vision in passerines and parrots
Our finding that the passerine ancestor had a violet-type SWS1 reaches slightly different conclusions in comparison with a recent study suggesting that the passerine ancestor was UVS [41], which was the first paper examining avian SWS1 evolution that used a phylogeny in which passerines and parrots were specified sister orders. Not only are the predicted ancestral sequences different, but a VS-type λmax in ancestral pigment was experimentally confirmed in our study. While it is not entirely clear why our study reached such different conclusions, there are a number of important differences. Our analysis included additional outgroup sequences, and used maximum likelihood reconstruction methods (as opposed to parsimony). Furthermore, in our study the ancestral pigments were experimentally recreated and functionally assayed. Finally, our phylogeny is based on the current understanding of phylogenetic relationships among landbirds that includes a recent revision of the relationships among higher lineages [56-58], and therefore is somewhat different from that of Odeen et al. [41]. However, we did not find any differences in our reconstructions of the ancestral passerine SWS1 when we used a tree with the relationships among higher passerines arranged similar to their phylogeny, suggesting that the difference in our findings from previous studies are probably due to methodological differences, such as the use of maximum likelihood reconstruction methods and/or the use of additional outgroup lineages. (Odeen et al. [41] did note that the inclusion of additional outgroup sequences resulted in an ambiguous reconstruction of the passerine ancestor even in their analyses.) Our results support earlier studies that investigated the evolution of UV/violet sensitivity in birds suggesting the passerine ancestor had a VS type SWS1 [25,28,55], but these early studies do not place passerines and parrots as sister orders. Because the parrots are now thought to be closer to the basal passerines than before, our results are more robust than they would be if based upon the older tree.
Our findings, that UVS in passerines and parrots evolved from VS ancestors, and that this occurred independently in at least two lineages, are rather unusual with respect to other vertebrate groups. The ancestral vertebrate state is thought to have been UVS, with VS pigments evolving independently in various lineages within fish, mammals, and amphibians [16,22-24,28,29,50]. Birds are believed to be an interesting exception where a switch to VS is thought to have occurred in the ancestral avian pigment with some descendants subsequently re-evolving UVS [24,50]. Our identification of VS type pigments in both passerine and parrot/passerine ancestors confirm this hypothesis, and our ancestral reconstruction results provide a more precise prediction of where these spectral sensitivity shifts occurred. The re-evolution of UVS from a VS type pigment has not previously been predicted elsewhere in the vertebrate phylogeny. The reasons why bird SWS1 pigments are an exception remain largely unknown, but may be related to their unique spectral tuning mechanisms among vertebrates.
Spectral tuning in C. nuchalis SWS1
The C. nuchalis VS pigment possesses an unusual residue combination at the two spectral tuning sites known to be most important in specifying UVS or VS in vertebrates: C86/S90. This residue combination has been found in a few passerine SWS1 opsins in past sequence-based surveys [41,42], but its spectral relevance has not been examined using mutagenesis experiments, which thus far have only dealt with VS-type pigments with S86/S90, in pigeon and chicken, [18,28] and UVS type with either A86/C90 or C86/C90, in budgerigar and zebra finch, respectively [17,18,59]. Past mutagenesis studies of vertebrate SWS1 pigments have shown the magnitude of λmax shift caused by a given amino acid change can differ significantly among pigments due to synergistic interactions within and between transmembrane regions I-VII [19,50,60,61]. Characterization of C. nuchalis SWS1 mutants was therefore carried out, as it may provide new clarification of the mechanisms contributing to the naturally occurring variation in avian SWS1 pigment spectral sensitivity, particularly among the VS type pigments. These mutants can also help clarify patterns of evolution between VS and UVS visual systems in birds.
Our results showing that S90C shifts the C. nuchalis SWS1 into the UV is consistent with previous studies where similar shifts have been documented in the chicken, pigeon, and the reverse in zebra finch, and budgerigar [17,18,28]. In C. nuchalis, the effect of the double mutant C86S/S90C was identical to that of the single S90C mutant. Thus, in the presence of C90, C86 has no additional effect on sensitivity. In other avian pigments, substitutions at known spectral tuning sites also do not change λmax if expressed with C90 [17,28]. Others have suggested that the effect of C90 is so strong it prevents detection of any subtler effects other residues might have [28]. In birds, all in vitro expressed pigments, whether wild type or mutant, with C90 have λmax ~360 nm. The exception is in chicken where S90C only shifts λmax to 369 nm [18].
The mutation C86F in C. nuchalis also shifts λmax into the UV. Unlike C90, which, as far as we know only has a functional role in avian SWS1 opsins, F86 is an important spectral tuning site across vertebrates where it confers UVS in most pigments in which it occurs [16,19,29,30], the exception being the aye-aye, which is VS despite the presence of F86 [23]. It is, in fact, believed to be the ancestral vertebrate state and substitutions from F86 are responsible for the loss of UVS in many mammalian lineages [16,19,22,23,29,30], and in ancient birds [21]. In C. nuchalis, C86 therefore plays an important role in maintaining sensitivity in the violet range, as the replacement of C86F shifts λmax into the ancestral UV state. F86 is also interesting because it has been suggested to be a second mechanism by which birds achieve UVS: It is found in the SWS1 genes of some birds including the trogon, paleognaths and a few sandgrouses and motmots [25-27], is capable of UV shifting VS pigments of pigeon and chicken [28], and is responsible for UVS in fish and most mammals [19,29,62]. Correspondingly, our mutagenesis results support the hypothesis that extant birds with F86 are UVS, and, therefore, the supposition that there are at least two mechanisms determining UVS in birds [28]. The expression of a wild type pigment with F86 would be needed to confirm this hypothesis.
In contrast to the previous mutants, C86S did not affect λmax in the C. nuchalis SWS1. This mutation was previously suggested as contributing to the broad spectral variation observed among VS type pigments [55,59], which in birds range from 388 nm (pigeon) to 420 nm (chicken) [28]. Site 86 is an important spectral tuning site in other vertebrate SWS1 pigments, and S86C is capable of shifting λmax into the UV in a hypothetical ancestral avian SWS1 [21]. As with C. nuchalis SWS1, S86C barely shifts λmax in the pigeon SWS1 [28], and mutation to serine at site 86 has no effect on the budgerigar SWS1 [17]. Therefore the residues responsible for this large variation in λmax among VS pigments remain unknown. Altogether, these studies indicate that the role of site 86 in avian SWS1 pigments depends not only on the residue at that site, but also on the background in which it is expressed. This is particularly true of mammalian SWS1 pigments where the variation at site 86 is better characterized: in most mammalian pigments the presence of F86 dramatically shifts λmax, into the UV [16,19,29,30], but this is not always the case [23].
Implications for behavioural ecology
While higher passerine lineages with UV type pigments are known to use UV signals in communication [9-11], current evidence indicates no link between colouration and spectral sensitivity in bowerbirds [39]. Here we have shown that despite the fact males display UV reflecting feathers and objects during courtship [3,63,64], C. nuchalis does not possess a UV type SWS1 visual pigment. These findings would seem to contradict evidence demonstrating a strong link between spectral tuning and signal colouration in other vertebrate groups, [65,66], and the belief that UV type pigments offer a dramatic advantage by improving sensitivity in this short wave range [67].
The general correlation between colouration and sensitivity remains because birds with VS pigments can perceive UV; SWS1 visual pigments absorb strongly over most of the UV visible range [6], cone oil droplets are effectively transparent to light in this range [68] and, in most species, avian ocular media transmit most short wavelength light [69]. The difference in UV sensitivity between UVS, VS and the blue shifted bowerbird VS is just a matter of degree. Nevertheless, while UV colouration might be perceived by bowerbirds, its importance in communication is not well understood. In the satin bowerbird (Ptilonorhynchus violaceus) plumage UV reflectance is correlated with factors such as the intensity of infection from blood parasites, feather growth rate, and body size [63], but it is unrelated to mating success [64].
Given that C. nuchalis and other bowerbird ocular media transmit more UV wavelengths than most other species with VS-type visual pigments, they might represent a transitional link in the evolution from a VS to a UVS visual system [39]. This hypothesis is supported by the comparatively blue shifted SWS1 found in bowerbirds, which further augments UV sensitivity. Given the similarly blue shifted λmax of the ancestral SWS1 pigments, this hypothesized transitional state might have originated in the ancestral passerine, and be shared among other basal passerines as well. This could also explain the unusually high number of shifts from VS to UVS in this order. Further investigation into the evolutionary history of ocular transmission would be useful to clarify this possibility.
If an organism with a blue shifted VS pigment, like the great bowerbird, has sufficient UV sensitivity, then the adaptive advantage of a switch to UVS might not be as large as it would be if it could perceive little UV or only had the ancestral VS pigment. Aside from λmax, there are a number of other structural and functional differences between VS and UVS opsins that may be related to a deprotonated Schiff base linkage to the chromophore [48,51,70-74]. These differences may have important consequences for the evolution of UVS in birds and other vertebrates. Therefore, it is possible that the wavelength difference between UVS and VS type pigments might not be the only, or the most important, functional difference between them. Further biochemical and mutagenesis studies would be necessary to refine the functional differences between these two opsin subtypes.
Conclusions
Our in vitro experiments suggest that spectral tuning in C. nuchalis is likely mediated by mechanisms very similar to those of other birds. This is unusual relative to spectral tuning mechanisms within mammals, which vary considerably among and within the major mammalian orders. In addition, despite both parrots and passerines sharing UV sensitivity and the same spectral tuning mechanism the experimentally recreated ancestral passerine and parrot/passerine SWS1 pigments were both found to be maximally sensitive in the violet; this suggests that UV sensitivity may have evolved independently in passerines and parrots from a violet sensitive ancestor. Moreover, our ancestral sequence reconstructions of SWS1 in landbird evolution suggest that transitions from VS to UVS are much more likely than the reverse. Our ancestral reconstruction experiments allow for a more precise prediction of where spectral sensitivity shifts may have occurred, and provide an unusual example where descendants have re-evolved UVS from a violet type ancestor; the reverse being more common in most vertebrates.
Methods
Opsin sequences
Birds were collected using cage traps or mist nets under appropriate Australian (Queensland Parks and Wildlife F1/000331/00/SAA, Australian Bird and Bat Banding Scheme 2434,1310, Commonwealth Scientific, Industrial and Research Organization (CSIRO) Ethics OB15/12, James Cook University Ethics A562, United States Department of Agriculture 47746, Australian Quarantine and Inspection Station 200104468, Environment Australia PWS P20011711, Department of Natural Resources Australia 1576) and US permits and authorizations (UCSB IACUC #10-98-555-1, USDA 47746). Birds were euthanized according to these protocols. Retinas were preserved in RNA Later (Invitrogen), and stored on ice in the field until they could be transferred to -80 for long term storage. RNA was extracted from retinal tissue using TRIzol Reagent (Invitrogen), and a cDNA library was prepared with the SMART cDNA Library Construction Kit (BD Biosciences). Degenerate primers were designed to amplify fragments of the opsin coding regions (Additional file 1: Table S1), with 3′ and 5′ ends of the genes isolated by RACE PCR. Purified PCR products were cloned into pJET1.2 (Fermentas), and sequenced from multiple clones. Site-directed mutagenesis was performed using the QuikChange kit (Stratagene). Blood samples of two individuals (“T + EB” & “BG/Z”) found in the Lavarack Barracks military base in Townsville City Queensland, Australia were preserved in Queen’s lysis buffer (0.01 M Tris, 0.01 M NaCl, 0.01 M sodium EDTA, and 1.0% n-lauroylsarcosine, pH 8.0) [75]. Genomic DNA was extracted from these blood samples using the DNeasy Blood and Tissue Kit (Qiagen). Introns and flanking genomic regions were isolated using PCR with specific primers on a genomic library created with the Genome Walker kit (Clontech).
Expression & purification of wild type and mutant pigments
Full-length coding sequences of C. nuchalis wild type pigments were amplified from cDNA, and cloned into the p1D4-hrGFP II expression vector for transient expression [76]. This vector has a C-terminal 1D4 epitope tag that encodes the last nine amino acids of bovine RH1 [TETSQVAPA], and employs the CMV promoter to drive transgene expression. Cultured HEK293T cells were transiently transfected with the opsin-1D4 construct using the Lipofectamine 2000 reagent (Invitrogen). Typically four 175 cm2 flasks were used per SWS1 expression procedure, with one flask of similarly expressed bovine rhodopsin as a control. Methods for purification of C. nuchalis SWS1 opsins were adapted from those of Starace & Knox [77]. Briefly, cells were harvested, washed with Harvesting Buffer (50 mM HEPES ph 6.6, 140 mM NaCl, 3 mM MgCl2), regenerated with 11-cis retinal chromophore, solubilized (in 1% n-dodecyl-β-D-maltopyranoside detergent (DM) with 20% (w/v) glycerol), and purified by batch immunoaffinity chromatography with the 1D4 monoclonal antibody [78]. The UV-visible absorption spectra of purified visual pigments were recorded at 21°C using a Cary 4000 dual beam spectrophotometer (Agilent). For functional assays, absorbance spectra were also measured after exposure to light (either a 366 nm UV light illuminator for UVS pigments, or a 60-W lamp with 440 nm cutoff filter for VS pigments), to hydrochloric acid (HCl; 100 mM), or to hydroxylamine (NH2OH; 50 mM). To produce difference spectra, either the light or the acid-denatured spectra were subtracted from the dark absorbance spectra. To estimate λmax, the dark absorbance spectra were baseline corrected and fit to a visual pigment template [73]. The F86 mutant λmax was estimated by fitting the dark-acid difference spectrum [29], due to a perturbation in the long wave arm of the dark spectrum. All amino acid numbering in this manuscript is according to the bovine rhodopsin amino acid sequence as a reference.
Ancestral sequence reconstruction
To reconstruct ancestral passerine SWS1 sequences, a dataset of 83 SWS1 genes from passerines, parrots and other related landbirds, as per Hackett et al. [31], was assembled from GenBank for a region of Helix 2 that encompasses all the known SWS1 spectral tuning sites (Additional file 1: Table S2 and Figure S1). For the majority of sequences, this region is the only portion of the SWS1 gene for which sequence data is available. The sequences were aligned with our C. nuchalis sequence using PRANK ([79], Figure S1). For ancestral reconstruction, a topology reflecting current understanding of landbird relationships was used (Figure 5) [31-33,40,56,80-87]. This phylogeny incorporates recent information that places passerines and parrots as derived sister orders relative to other Landbird orders [31-33], and includes a recent revision of the relationships among higher passerine lineages [56-58]. This phylogeny is somewhat different from previous avian SWS1 studies, therefore we also analyzed our data on an alternate phylogeny (Additional file 1: Figure S4) similar to that of Odeen et al. [41], in order to investigate the robustness of our ancestral reconstructions.
For the ancestral sequence reconstruction (ASR), a combination of empirical Bayesian and maximum likelihood (ML) codon-based methods [88] were used (PAML v4.3 [89]). Nested random sites codon models were compared using likelihood ratio tests (LRTs) [90,91], and the best fitting model, M7 [92], was used for the ancestral sequence reconstruction (Additional file 1: Table S3 and S4). Multiple runs were carried out with different starting values to check for convergence in all analyses. In experimentally resurrecting ancestral proteins, focusing solely on the most probable ancestral sequence can introduce biases in amino acid composition, which may in turn alter the functional phenotype of a resurrected protein [93,94]. We addressed this concern using a strategy of weighted random sampling of ancestral sequences from the posterior distribution, in order to avoid this bias [94,95]. For the two ancestral nodes reconstructed, a weighted sampling of 10,000 sequences from the posterior distribution resulted in ancestral sequences that were either identical (parrot/passerine ancestor, 100% identical), or highly similar to (passerine ancestor, 83% or 8343 sequences out of 10,000 identical) the most likely ancestral reconstruction.
Homology modeling
The 3D structure of the C. nuchalis wild-type SWS1 was inferred via homology modeling by Modeller [96], using bovine rhodopsin (PDB code: 1U19, [97]) as template. Fifty models were generated by optimizing the Modeller objective function with the model with the lowest DOPE score [98] selected for further assessment and visualization. Model quality was checked using ProSA-web [99] to ensure the model and template structures have comparable z-scores (an standardized indicator of a structure’s total energy compared to that expected by random chance), and by ProCheck [100], to ensure bond lengths and angles do not have unusual stereochemical conformations. Similar procedures were followed for inferring 3D structures of C86F, C86S and S90C mutants.
Abbreviations
SWS1: Short-wavelength sensitive 1; UV: Ultraviolet; V: Violet; λmax: Wavelength of maximum absorbance.
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
BSWC, JAE and IvH conceived of the study. IvH performed the lab work, compiled the data, performed analyses and drafted the manuscript. AS performed the homology modeling and structural analyses, and LD collected the samples. BSWC guided all aspects of the study, and helped to draft the manuscript. All authors contributed to the final version of the manuscript.
Supplementary Material
Degenerate oligonucleotides for PCR (numbering according to bovine rhodopsin). Table S2. Species names & accession numbers for Landbird SWS1 data set used in ancestral reconstruction analysis. Table S3. Likelihood scores of codon models used for ancestral reconstruction. Table S4. Maximum likelihood ancestral reconstruction of ancestral passerine/parrot, and ancestral passerine SWS1 pigments, with posterior probabilities (numbering according to bovine rhodo). Figure S1. Alignment of SWS1 opsin gene, helix 2 from Landbirds used in ancestral reconstruction, highlighting sites 86, 90 & 93. Figure S2. Alignment of visual pigment sequences in C. nuchalis. Figure S3. Phylogenetic relationships of the C. nuchalis opsin genes with those of other vertebrates. Figure S4. Alternate Landbird topologies used to confirm ancestral sequence reconstruction [101-108].
Contributor Information
Ilke van Hazel, Email: i.vanhazel@utoronto.ca.
Amir Sabouhanian, Email: amir.sabouhanian@utoronto.ca.
Lainy Day, Email: lainyday@olemiss.edu.
John A Endler, Email: john.endler@deakin.edu.au.
Belinda SW Chang, Email: belinda.chang@utoronto.ca.
Acknowledgements
This work was supported by a National Sciences and Engineering Research Council (NSERC) Discovery grant (B.S.W.C.), an NSERC Postgraduate Scholarship (I.v.H.), and a University of Toronto Vision Science Research Program Fellowship (I.v.H.).
References
- Doucet SM, Montgomerie R. Bower location and orientation in satin bowerbirds: optimising the conspicuousness of male display? Emu. 2003;13(2):105–109. doi: 10.1071/MU02024. [DOI] [Google Scholar]
- Endler JA, Day LB. Ornament colour selection, visual contrast and the shape of colour preference functions in great bowerbirds, Chlamydera nuchalis. Anim Behav. 2006;13:1405–1416. doi: 10.1016/j.anbehav.2006.05.005. [DOI] [Google Scholar]
- Endler JA, Westcott DA, Madden JR, Robson T. Animal visual systems and the evolution of color patterns: sensory processing illuminates signal evolution. Evolution. 2005;13(8):1795–1818. doi: 10.1554/04-669.1. [DOI] [PubMed] [Google Scholar]
- Doucet SM, Shawkey MD, Hill GE, Montgomerie R. Iridescent plumage in satin bowerbirds: structure, mechanisms and nanostructural predictors of individual variation in colour. J Exp Biol. 2006;13(Pt 2):380–390. doi: 10.1242/jeb.01988. [DOI] [PubMed] [Google Scholar]
- Endler JA, Endler LC, Doerr NR. Great bowerbirds create theaters with forced perspective when seen by their audience. Curr Biol. 2010;13(18):1679–1684. doi: 10.1016/j.cub.2010.08.033. [DOI] [PubMed] [Google Scholar]
- Endler JA, Mielke PW. Comparing entire colour patterns as birds see them. Biol J Linn Soc. 2005;13(4):405–431. doi: 10.1111/j.1095-8312.2005.00540.x. [DOI] [Google Scholar]
- Viitala J, Korpimäki E, Palokangas P, Koivula M. Attraction of kestrels to vole scent marks visible in ultraviolet-light. Nature. 1995;13(6513):425–427. doi: 10.1038/373425a0. [DOI] [Google Scholar]
- Siitari H, Honkavaara J, Huhta E, Viitala J. Ultraviolet reflection and female mate choice in the pied flycatcher, Ficedula hypoleuca. Anim Behav. 2002;13(1):97–102. doi: 10.1006/anbe.2001.1870. [DOI] [Google Scholar]
- Alonso-Alvarez C, Doutrelant C, Sorci G. Ultraviolet reflectance affects male-male interactions in the blue tit (Parus caeruleus ultramarinus) Behav Ecol. 2004;13(5):805–809. doi: 10.1093/beheco/arh083. [DOI] [Google Scholar]
- Andersson S, Amundsen T. Ultraviolet colour vision and ornamentation in bluethroats. P Roy Soc B-Biol Sci. 1997;13(1388):1587–1591. doi: 10.1098/rspb.1997.0221. [DOI] [Google Scholar]
- Hunt S, Bennett ATD, Cuthill IC, Griffiths R. Blue tits are ultraviolet tits. Proc R Soc Lond B. 1998;13(1395):451–455. doi: 10.1098/rspb.1998.0316. [DOI] [Google Scholar]
- Pearn SM, Bennett ATD, Cuthill IC. Ultraviolet vision, fluorescence and mate choice in a parrot, the budgerigar Melopsittacus undulatus. Proc Biol Sci / The Royal Soc. 2001;13(1482):2273–2279. doi: 10.1098/rspb.2001.1813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Menon ST, Han M, Sakmar TP. Rhodopsin: structural basis of molecular physiology. Physiol Rev. 2001;13(4):1659–1688. doi: 10.1152/physrev.2001.81.4.1659. [DOI] [PubMed] [Google Scholar]
- Kochendoerfer GG, Lin SW, Sakmar TP, Mathies RA. How color visual pigments are tuned. Trends Biochem Sci. 1999;13(8):300–305. doi: 10.1016/S0968-0004(99)01432-2. [DOI] [PubMed] [Google Scholar]
- Bowmaker JK. Evolution of vertebrate visual pigments. Vision research. 2008;13(20):2022–2041. doi: 10.1016/j.visres.2008.03.025. [DOI] [PubMed] [Google Scholar]
- Cowing JA, Poopalasundaram S, Wilkie SE, Robinson PR, Bowmaker JK, Hunt DM. The molecular mechanism for the spectral shifts between vertebrate ultraviolet- and violet-sensitive cone visual pigments. Biochem J. 2002;13(Pt 1):129–135. doi: 10.1042/BJ20020483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilkie SE, Robinson PR, Cronin TW, Poopalasundaram S, Bowmaker JK, Hunt DM. Spectral tuning of avian violet- and ultraviolet-sensitive visual pigments. Biochemistry. 2000;13(27):7895–7901. doi: 10.1021/bi992776m. [DOI] [PubMed] [Google Scholar]
- Yokoyama S, Radlwimmer FB, Blow NS. Ultraviolet pigments in birds evolved from violet pigments by a single amino acid change. Proc Natl Acad Sci. 2000;13(13):7366–7371. doi: 10.1073/pnas.97.13.7366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fasick JI, Applebury ML, Oprian DD. Spectral tuning in the mammalian short-wavelength sensitive cone pigments. Biochemistry. 2002;13(21):6860–6865. doi: 10.1021/bi0200413. [DOI] [PubMed] [Google Scholar]
- Fasick JI, Lee N, Oprian DD. Spectral tuning in the human blue cone pigment. Biochemistry. 1999;13(36):11593–11596. doi: 10.1021/bi991600h. [DOI] [PubMed] [Google Scholar]
- Shi Y, Yokoyama S. Molecular analysis of the evolutionary significance of ultraviolet vision in vertebrates. Proc Natl Acad Sci U S A. 2003;13(14):8308–8313. doi: 10.1073/pnas.1532535100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takahashi Y, Ebrey T. Molecular basis of spectral tuning in the newt short wavelength sensitive visual pigment†. Biochemistry. 2003;13:6025–6034. doi: 10.1021/bi020629+. [DOI] [PubMed] [Google Scholar]
- Carvalho LS, Davies WL, Robinson PR, Hunt DM. Spectral tuning and evolution of primate short-wavelength-sensitive visual pigments. Proc Biol Sci / The Royal Soc. 2012;13(1727):387–393. doi: 10.1098/rspb.2011.0782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yokoyama S, Shi Y. Genetics and evolution of ultraviolet vision in vertebrates. FEBS Lett. 2000;13(2):167–172. doi: 10.1016/S0014-5793(00)02269-9. [DOI] [PubMed] [Google Scholar]
- Ödeen A, Håstad O. Complex distribution of avian color vision systems revealed by sequencing the SWS1 opsin from total DNA. Mol Biol Evol. 2003;13(6):855–861. doi: 10.1093/molbev/msg108. [DOI] [PubMed] [Google Scholar]
- Aidala Z, Huynen L, Brennan PL, Musser J, Fidler A, Chong N, Machovsky Capuska GE, Anderson MG, Talaba A, Lambert D. et al. Ultraviolet visual sensitivity in three avian lineages: paleognaths, parrots, and passerines. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2012;13(7):495–510. doi: 10.1007/s00359-012-0724-3. [DOI] [PubMed] [Google Scholar]
- Ödeen A, Håstad O. The phylogenetic distribution of ultraviolet sensitivity in birds. BMC Evol Biol. 2013;13:36. doi: 10.1186/1471-2148-13-36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carvalho LS, Cowing JA, Wilkie SE, Bowmaker JK, Hunt DM. The molecular evolution of avian ultraviolet- and violet-sensitive visual pigments. Mol Biol Evol. 2007;13(8):1843–1852. doi: 10.1093/molbev/msm109. [DOI] [PubMed] [Google Scholar]
- Parry JWL, Poopalasundaram S, Bowmaker JK, Hunt DM. A novel amino acid substitution is responsible for spectral tuning in a rodent violet-sensitive visual pigment. Biochemistry. 2004;13(25):8014–8020. doi: 10.1021/bi049478w. [DOI] [PubMed] [Google Scholar]
- Yokoyama S, Takenaka N, Agnew DW, Shoshani J. Elephants and human color-blind deuteranopes have identical sets of visual pigments. Genetics. 2005;13(1):335–344. doi: 10.1534/genetics.104.039511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hackett SJ, Kimball RT, Reddy S, Bowie RC, Braun EL, Braun MJ, Chojnowski JL, Cox WA, Han KL, Harshman J. et al. A phylogenomic study of birds reveals their evolutionary history. Science. 2008;13(5884):1763–1768. doi: 10.1126/science.1157704. [DOI] [PubMed] [Google Scholar]
- Suh A, Paus M, Kiefmann M, Churakov G, Franke FA, Brosius J, Kriegs JO, Schmitz J. Mesozoic retroposons reveal parrots as the closest living relatives of passerine birds. Nat Commun. 2011;13:443. doi: 10.1038/ncomms1448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang N, Braun EL, Kimball RT. Testing hypotheses about the sister group of the passeriformes using an independent 30-locus data set. Mol Biol Evol. 2012;13(2):737–750. doi: 10.1093/molbev/msr230. [DOI] [PubMed] [Google Scholar]
- Pratt RC, Gibb GC, Morgan-Richards M, Phillips MJ, Hendy MD, Penny D. Toward resolving deep Neoaves phylogeny: data, signal enhancement and priors. Mol Biol Evol. 2009;13:313–326. doi: 10.1093/molbev/msn248. [DOI] [PubMed] [Google Scholar]
- Pacheco MA, Battistuzzi FU, Lentino M, Aguilar RF, Kumar S, Escalante AA. Evolution of modern birds revealed by mitogenomics: timing the radiation and origin of major orders. Mol Biol Evol. 2011;13:1927–1942. doi: 10.1093/molbev/msr014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilkie SE, Vissers PM, Das D, Degrip WJ, Bowmaker JK, Hunt DM. The molecular basis for UV vision in birds: spectral characteristics, cDNA sequence and retinal localization of the UV-sensitive visual pigment of the budgerigar (Melopsittacus undulatus) Biochem J. 1998;13(Pt 1):541–547. doi: 10.1042/bj3300541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yokoyama S, Blow NS, Radlwimmer FB. Molecular evolution of color vision of zebra finch. Gene. 2000;13(1–2):17–24. doi: 10.1016/s0378-1119(00)00435-2. [DOI] [PubMed] [Google Scholar]
- Carvalho LS, Knott B, Berg ML, Bennett AT, Hunt DM. Ultraviolet-sensitive vision in long-lived birds. Proc Biol Sci / The Royal Soc. 2011;13(1702):107–114. doi: 10.1098/rspb.2010.1100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coyle BJ, Hart NS, Carleton KL, Borgia G. Limited variation in visual sensitivity among bowerbird species suggests that there is no link between spectral tuning and variation in display colouration. J Exp Biol. 2012;13(Pt 7):1090–1105. doi: 10.1242/jeb.062224. [DOI] [PubMed] [Google Scholar]
- Barker FK, Cibois A, Schikler P, Feinstein J, Cracraft J. Phylogeny and diversification of the largest avian radiation. Proc Natl Acad Sci U S A. 2004;13(30):11040–11045. doi: 10.1073/pnas.0401892101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ödeen A, Håstad O, Alström P. Evolution of ultraviolet vision in the largest avian radiation - the passerines. BMC Evol Biol. 2011;13(1):313. doi: 10.1186/1471-2148-11-313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ödeen A, Håstad O. New primers for the avian SWS1 pigment opsin gene reveal new amino acid configurations in spectral sensitivity tuning sites. J Hered. 2009;13(6):784–789. doi: 10.1093/jhered/esp060. [DOI] [PubMed] [Google Scholar]
- Kawamura S, Blow NS, Yokoyama S. Genetic analyses of visual pigments of the pigeon (Columba livia) Genetics. 1999;13(4):1839–1850. doi: 10.1093/genetics/153.4.1839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kito Y, Suzuki T, Azuma M, Sekoguti Y. Absorption spectrum of rhodopsin denatured with acid. Nature. 1968;13(5145):955. doi: 10.1038/218955a0. &. [DOI] [PubMed] [Google Scholar]
- Matthews RG, Hubbard R, Brown PK, Wald G. Tautomeric forms of metarhodopsin. J Gen Physiol. 1963;13:215–240. doi: 10.1085/jgp.47.2.215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koutalos Y, Ebrey TG, Tsuda M, Odashima K, Lien T, Park MH, Shimizu N, Nakanishi K, Gilson HR, Honig B. et al. Regeneration of bovine and octopus opsins in situ with natural and artificial retinals. Biochemistry. 1989;13:2732–2739. doi: 10.1021/bi00432a055. [DOI] [PubMed] [Google Scholar]
- Ma JX, Kono M, Xu L, Das J, Ryan JC, Hazard ES, Oprian DD, Crouch RK. Salamander UV cone pigment: sequence, expression, and spectral properties. Visual Neurosci. 2001;13(3):393–399. doi: 10.1017/S0952523801183057. [DOI] [PubMed] [Google Scholar]
- Das J, Crouch RK, Ma JX, Oprian DD, Kono M. Role of the 9-methyl group of retinal in cone visual pigments. Biochemistry. 2004;13(18):5532–5538. doi: 10.1021/bi036097u. [DOI] [PubMed] [Google Scholar]
- Dukkipati A, Kusnetzow A, Babu KR, Ramos L, Singh D, Knox BE, Birge RR. Phototransduction by vertebrate ultraviolet visual pigments: protonation of the retinylidene Schiff base following photobleaching. Biochemistry. 2002;13(31):9842–9851. doi: 10.1021/bi025883g. [DOI] [PubMed] [Google Scholar]
- Shi Y, Radlwimmer FB, Yokoyama S. Molecular genetics and the evolution of ultraviolet vision in vertebrates. Proc Natl Acad Sci U S A. 2001;13(20):11731–11736. doi: 10.1073/pnas.201257398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsutsui K, Shichida Y. Photosensitivities of rhodopsin mutants with a displaced counterion. Biochemistry. 2010;13(47):10089–10097. doi: 10.1021/bi101020p. [DOI] [PubMed] [Google Scholar]
- Rao VR, Cohen GB, Oprian DD. Rhodopsin mutation G90D and a molecular mechanism for congenital night blindness. Nature. 1994;13(6464):639–642. doi: 10.1038/367639a0. [DOI] [PubMed] [Google Scholar]
- Janz JM, Farrens DL. Engineering a functional blue-wavelength-shifted rhodopsin mutant. Biochemistry. 2001;13(24):7219–7227. doi: 10.1021/bi002937i. [DOI] [PubMed] [Google Scholar]
- Lin SW, Kochendoerfer GG, Carroll KS, Wang D, Mathies RA, Sakmar TP. Mechanisms of spectral tuning in blue cone visual pigments: visible and raman spectroscopy of blue-shifted rhodopsin mutants. J Biol Chem. 1998;13(38):24583–24591. doi: 10.1074/jbc.273.38.24583. [DOI] [PubMed] [Google Scholar]
- Hunt DM, Carvalho LS, Cowing JA, Davies WL. Evolution and spectral tuning of visual pigments in birds and mammals. Philos Trans R Soc Lond B Biol Sci. 2009;13(1531):2941–2955. doi: 10.1098/rstb.2009.0044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Irestedt M, Ohlson JI. The division of the major songbird radiation into passerida and ‘core corvoidea’ (aves: passeriformes) — the species tree vs. gene trees. Zool Scr. 2008;13(3):305–313. doi: 10.1111/j.1463-6409.2007.00321.x. [DOI] [Google Scholar]
- Jønsson KA, Irestedt M, Fuchs J, Ericson PGP, Christidis L, Bowie RCK, Norman JA, Pasquet E, Fjeldså J. Explosive avian radiations and multi-directional dispersal across wallacea: evidence from the campephagidae and other crown corvida (aves) Mol Phylogenet Evol. 2008;13(1):221–236. doi: 10.1016/j.ympev.2008.01.017. [DOI] [PubMed] [Google Scholar]
- Zuccon D, Ericson PGP. Molecular and morphological evidences place the extinct New Zealand endemic Turnagra capensis in the oriolidae. Mol Phylogenet Evol. 2012;13(1):414–426. doi: 10.1016/j.ympev.2011.10.013. [DOI] [PubMed] [Google Scholar]
- Hunt DM, Cowing JA, Wilkie SE, Parry JWL, Poopalasundaram S, Bowmaker JK. Divergent mechanisms for the tuning of shortwave sensitive visual pigments in vertebrates. Photochem Photobiol Sci. 2004;13(6):713. doi: 10.1039/b314693f. [DOI] [PubMed] [Google Scholar]
- Takahashi Y, Yokoyama S. Genetic basis of spectral tuning in the violet-sensitive visual pigment of African clawed frog, Xenopus laevis. Genetics. 2005;13(3):1153–1160. doi: 10.1534/genetics.105.045849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yokoyama S, Starmer WT, Takahashi Y, Tada T. Tertiary structure and spectral tuning of UV and violet pigments in vertebrates. Gene. 2006;13:95–103. doi: 10.1016/j.gene.2005.09.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cowing JA, Poopalasundaram S, Wilkie SE, Bowmaker JK, Hunt DM. Spectral tuning and evolution of short wave-sensitive cone pigments in cottoid fish from Lake Baikal. Biochemistry. 2002;13(19):6019–6025. doi: 10.1021/bi025656e. [DOI] [PubMed] [Google Scholar]
- Doucet SM, Montgomerie R. Multiple sexual ornaments in satin bowerbirds: ultraviolet plumage and bowers signal different aspects of male quality. Behav Ecol. 2003;13(4):503–509. doi: 10.1093/beheco/arg035. [DOI] [Google Scholar]
- Savard J-F, Keagy J, Borgia G. Blue, not UV, plumage color is important in satin bowerbird Ptilonorhynchus violaceus display. J Avian Biol. 2011;13(1):80–84. doi: 10.1111/j.1600-048X.2010.05128.x. [DOI] [Google Scholar]
- Seehausen O, Terai Y, Magalhaes IS, Carleton KL, Mrosso HDJ, Miyagi R, van der Sluijs I, Schneider MV, Maan ME, Tachida H. et al. Speciation through sensory drive in cichlid fish. Nature. 2008;13(7213):620–626. doi: 10.1038/nature07285. [DOI] [PubMed] [Google Scholar]
- Carleton KL, Parry JW, Bowmaker JK, Hunt DM, Seehausen O. Colour vision and speciation in Lake Victoria cichlids of the genus Pundamilia. Mol Ecol. 2005;13(14):4341–4353. doi: 10.1111/j.1365-294X.2005.02735.x. [DOI] [PubMed] [Google Scholar]
- Hausmann F, Arnold KE, Marshall NJ, Owens IPF. Ultraviolet signals in birds are special. Proc R Soc Lond B. 2003;13(1510):61–67. doi: 10.1098/rspb.2002.2200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowmaker JK. Colour-vision in birds and the role of oil droplets. Trends Neurosci. 1980;13(8):196–199. doi: 10.1016/0166-2236(80)90072-7. [DOI] [Google Scholar]
- Hart NS, Vorobyev M. Modelling oil droplet absorption spectra and spectral sensitivities of bird cone photoreceptors. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2005;13(4):381–392. doi: 10.1007/s00359-004-0595-3. [DOI] [PubMed] [Google Scholar]
- Kusnetzow A, Dukkipati A, Babu K, Ramos L, Knox B, Birge R. Vertebrate ultraviolet visual pigments: protonation of the retinylidene schiff base and a counterion switch during photoactivation. Proc Natl Acad Sci U S A. 2004;13(4):941–946. doi: 10.1073/pnas.0305206101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen MH, Kuemmel C, Birge RR, Knox BE. Rapid release of retinal from a cone visual pigment following photoactivation. Biochemistry. 2012;13(20):4117–4125. doi: 10.1021/bi201522h. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo D-G, Yue WWS, Ala-Laurila P, Yau K-W. Activation of visual pigments by light and heat. Science. 2011;13(6035):1307–1312. doi: 10.1126/science.1200172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Govardovskii VI, Fyhrquist N, Reuter T, Kuzmin DG, Donner K. In search of the visual pigment template. Vis Neurosci. 2000;13(4):509–528. doi: 10.1017/S0952523800174036. [DOI] [PubMed] [Google Scholar]
- Tsutsui K, Imai H, Shichida Y. Photoisomerization efficiency in UV-absorbing visual pigments: protein-directed isomerization of an unprotonated retinal schiff base†. Biochemistry. 2007;13(21):6437–6445. doi: 10.1021/bi7003763. [DOI] [PubMed] [Google Scholar]
- Seutin G, White BN, Boag PT. Preservation of avian blood and tissue samples for DNA analyses. Can J Zool. 1991;13(1):82–90. doi: 10.1139/z91-013. [DOI] [Google Scholar]
- Morrow JM, Chang BS. The p1D4-hrGFP II expression vector: a tool for expressing and purifying visual pigments and other G protein-coupled receptors. Plasmid. 2010;13(3):162–169. doi: 10.1016/j.plasmid.2010.07.002. [DOI] [PubMed] [Google Scholar]
- Starace DM, Knox BE. Cloning and expression of a Xenopus short wavelength cone pigment. Exp Eye Res. 1998;13(2):209–220. doi: 10.1006/exer.1998.0507. [DOI] [PubMed] [Google Scholar]
- Molday RS, MacKenzie D. Monoclonal antibodies to rhodopsin: characterization, cross-reactivity, and application as structural probes. Biochemistry. 1983;13(3):653–660. doi: 10.1021/bi00272a020. [DOI] [PubMed] [Google Scholar]
- Loytynoja A, Goldman N. An algorithm for progressive multiple alignment of sequences with insertions. Proc Natl Acad Sci U S A. 2005;13:10557–10562. doi: 10.1073/pnas.0409137102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barker FK, Barrowclough GF, Groth JG. A phylogenetic hypothesis for passerine birds: taxonomic and biogeographic implications of an analysis of nuclear DNA sequence data. Proc Biol Sci / The Royal Soc. 2002;13(1488):295–308. doi: 10.1098/rspb.2001.1883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beresford P, Barker FK, Ryan PG, Crowe TM. African endemics span the tree of songbirds (Passeri): molecular systematics of several evolutionary ‘enigmas’. Proc Biol Sci / The Royal Soc. 2005;13(1565):849–858. doi: 10.1098/rspb.2004.2997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johansson US, Fjeldså J, Bowie RCK. Phylogenetic relationships within passerida (aves: passeriformes): a review and a new molecular phylogeny based on three nuclear intron markers. Mol Phylogenet Evol. 2008;13(3):858–876. doi: 10.1016/j.ympev.2008.05.029. [DOI] [PubMed] [Google Scholar]
- Wright TF, Schirtzinger EE, Matsumoto T, Eberhard JR, Graves GR, Sanchez JJ, Capelli S, Müller H, Scharpegge J, Chambers GK. et al. A multilocus molecular phylogeny of the parrots (psittaciformes): support for a gondwanan origin during the cretaceous. Mol Biol Evol. 2008;13(10):2141–2156. doi: 10.1093/molbev/msn160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gardner JL, Trueman JW, Ebert D, Joseph L, Magrath RD. Phylogeny and evolution of the Meliphagoidea, the largest radiation of Australasian songbirds. Mol Phylogenet Evol. 2010;13(3):1087–1102. doi: 10.1016/j.ympev.2010.02.005. [DOI] [PubMed] [Google Scholar]
- Driskell AC, Norman JA, Pruett-Jones S, Mangall E, Sonsthagen S, Christidis L. A multigene phylogeny examining evolutionary and ecological relationships in the Australo-Papuan wrens of the subfamily malurinae (aves) Mol Phylogenet Evol. 2011;13(3):480–485. doi: 10.1016/j.ympev.2011.03.030. [DOI] [PubMed] [Google Scholar]
- Jønsson KA, Fabre P-H, Ricklefs RE, Fjeldså J. Major global radiation of corvoid birds originated in the proto-Papuan archipelago. Proc Natl Acad Sci U S A. 2011;13(6):2328–2333. doi: 10.1073/pnas.1018956108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee JY, Joseph L, Edwards SV. A species tree for the Australo-Papuan fairy-wrens and allies (Aves: Maluridae) Syst Biol. 2012;13(2):253–271. doi: 10.1093/sysbio/syr101. [DOI] [PubMed] [Google Scholar]
- Yang Z, Kumar S, Nei M. A new method of inference of ancestral nucleotide and amino acid sequences. Genetics. 1995;13(4):1641–1650. doi: 10.1093/genetics/141.4.1641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang Z. PAML 4: phylogenetic analysis by maximum likelihood. Mol Biol Evol. 2007;13(8):1586–1591. doi: 10.1093/molbev/msm088. [DOI] [PubMed] [Google Scholar]
- Felsenstein J. Evolutionary trees from gene-frequencies and quantitative characters - finding maximum-likelihood estimates. Evolution. 1981;13(6):1229–1242. doi: 10.2307/2408134. [DOI] [PubMed] [Google Scholar]
- Yang Z, Kumar S. Approximate methods for estimating the pattern of nucleotide substitution and the variation of substitution rates among sites. Mol Biol Evol. 1996;13(5):650–659. doi: 10.1093/oxfordjournals.molbev.a025625. [DOI] [PubMed] [Google Scholar]
- Yang Z, Nielsen R, Goldman N, Pedersen AK. Codon-substitution models for heterogeneous selection pressure at amino acid sites. Genetics. 2000;13(1):431–449. doi: 10.1093/genetics/155.1.431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams PD, Pollock DD, Blackburne BP, Goldstein RA. Assessing the accuracy of ancestral protein reconstruction methods. PLoS Comput Biol. 2006;13(6):e69. doi: 10.1371/journal.pcbi.0020069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pollock DD, Chang BSW. In: Ancestral sequence reconstruction. Liberles DA, editor. New York: Oxford University Press; 2007. Dealing with uncertainty in ancestral sequence reconstruction: sampling from the posterior distribution; pp. 85–94. [Google Scholar]
- Gaucher EA, Govindarajan S, Ganesh OK. Palaeotemperature trend for precambrian life inferred from resurrected proteins. Nature. 2008;13(7179):704–707. doi: 10.1038/nature06510. [DOI] [PubMed] [Google Scholar]
- Sali A, Blundell TL. Comparative protein modelling by satisfaction of spatial restraints. J Mol Biol. 1993;13(3):779–815. doi: 10.1006/jmbi.1993.1626. [DOI] [PubMed] [Google Scholar]
- Okada T, Sugihara M, Bondar AN, Elstner M, Entel P, Buss V. The retinal conformation and its environment in rhodopsin in light of a new 2.2 A crystal structure. J Mol Biol. 2004;13(2):571–583. doi: 10.1016/j.jmb.2004.07.044. [DOI] [PubMed] [Google Scholar]
- Shen MY, Sali A. Statistical potential for assessment and prediction of protein structures. Protein Sci. 2006;13(11):2507–2524. doi: 10.1110/ps.062416606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wiederstein M, Sippl MJ. ProSA-web: interactive web service for the recognition of errors in three-dimensional structures of proteins. Nucleic Acids Res. 2007;13(Web Server issue):W407–W410. doi: 10.1093/nar/gkm290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laskowski RA, Macarthur MW, Moss DS, Thornton JM. Procheck - a program to check the stereochemical quality of protein structures. J Appl Crystallogr. 1993;13:283–291. doi: 10.1107/S0021889892009944. [DOI] [Google Scholar]
- Das D, Wilkie SE, Hunt DM, Bowmaker JK. Visual pigments and oil droplets in the retina of a passerine bird, the canary Serinus canaria: microspectrophotometry and opsin sequences. Vision Res. 1999;13:2801–2815. doi: 10.1016/S0042-6989(99)00023-1. [DOI] [PubMed] [Google Scholar]
- Felsenstein J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution. 1985;13:783–791. doi: 10.2307/2408678. [DOI] [PubMed] [Google Scholar]
- Guindon S, Gascuel O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst Biol. 2003;13:696–704. doi: 10.1080/10635150390235520. [DOI] [PubMed] [Google Scholar]
- Guindon S, Lethiec F, Duroux P, Gascuel O. PHYML online–a web server for fast maximum likelihood-based phylogenetic inference. Nucleic Acids Res. 2005;13:W557–W559. doi: 10.1093/nar/gki352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nylander J. Program distributed by the author. Uppsala Sweden: Evolutionary Biology Centre, Uppsala University; 2008. MrModeltest v2.3. [Google Scholar]
- Ödeen A, Pruett-Jones S, Driskell AC, Armenta JK, Hastad O. Multiple shifts between violet and ultraviolet vision in a family of passerine birds with associated changes in plumage coloration. Proc Biol Sci. 2012;13:1269–1276. doi: 10.1098/rspb.2011.1777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ronquist F, Huelsenbeck JP. MrBayes 3: bayesian phylogenetic inference under mixed models. Bioinformatics. 2003;13:1572–1574. doi: 10.1093/bioinformatics/btg180. [DOI] [PubMed] [Google Scholar]
- Seddon N, Tobias JA, Eaton M, Odeen A. Human vision can provide a valid proxy for avian perception of sexual dichromatism. Auk. 2010;13:283–292. doi: 10.1525/auk.2009.09070. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Degenerate oligonucleotides for PCR (numbering according to bovine rhodopsin). Table S2. Species names & accession numbers for Landbird SWS1 data set used in ancestral reconstruction analysis. Table S3. Likelihood scores of codon models used for ancestral reconstruction. Table S4. Maximum likelihood ancestral reconstruction of ancestral passerine/parrot, and ancestral passerine SWS1 pigments, with posterior probabilities (numbering according to bovine rhodo). Figure S1. Alignment of SWS1 opsin gene, helix 2 from Landbirds used in ancestral reconstruction, highlighting sites 86, 90 & 93. Figure S2. Alignment of visual pigment sequences in C. nuchalis. Figure S3. Phylogenetic relationships of the C. nuchalis opsin genes with those of other vertebrates. Figure S4. Alternate Landbird topologies used to confirm ancestral sequence reconstruction [101-108].