

Abstract
orf256 is a wheat mitochondrial gene associated with cytoplasmic male sterility (CMS) that has different organization in various species. This study exploited the orf256 gene as a mitochondrial DNA marker to study the genetic fingerprint of Triticum and Aegilops species. PCR followed by sequencing of common parts of the orf256 gene were employed to determine the fingerprint and molecular evolution of Triticum and Aegilops species. Although many primer pairs were used, two pairs of orf256 specific primers (5:-94/C: 482, 5:253/C: 482), amplified DNA fragments of 576 bp and 230 bp respectively in all species were tested. A common 500 bp of nine species of Triticum and Aegilops were aligned and showed consistent results with that obtained from other similar chloroplast or nuclear genes. Base alignment showed that there were various numbers of base substitutions in all species compared to S. cereal (Sc) (the outgroup species). Phylogenetic relationship revealed similar locations and proximity on phylogenetic trees established using plastid and nuclear genes. The results of this study open a good route to use unknown function genes of mitochondria in studying the molecular relationships and evolution of wheat and complex plant genomes.
Keywords: wheat, Triticum, Aegilops, orf256, fingerprinting, phylogenetic, molecular evolution
1. Introduction
Hexaploid bread wheat (Triticum aestivum) is a member of Triticeae tribe, which also includes barley (Hordeum vulgare) and rye (Secale cereale) as well as other diploid and tetraploid wheats. Meiotic studies indicated that the general evolution of the Triticeae tribe has been defined by divergence at the diploid level from a common diploid ancestor and convergence at the polyploid level involving the diverged diploid genomes [1]. Cytological and molecular studies provided information on the identity of donors and the patterns of genome evolution of the Triticum/Aegilops species [2]. The Triticum and Aegilops genera contain 13 diploid and 18 polyploid species [3]. T. monococcum includes the cultivated form T. monococcum ssp. monococcum and the wild form, T. boeoticum. There are two tetraploid wheat species: Triticum temopheevii (AAGG genome) and Triticum turgidum (AABB genome). Finally, there are two hexaploid wheats: Triticum zhukovskyi (AAAAGG) and T. aestivum (AABBDD), including several subspecies [3,4].
T. aestivum is hexaploid with a genome constitution of AABBDD, and was formed about 8,000 years ago from hybridization between T. turgidum (AABB) and A. tauschii (DD) [5,6,7]. The A genome originated with T. urartu (AA), which is closely related to T. monococcum (AA). A. speltoides, A. bicornis, A. searsii, and A. sharonensis appear to have diverged from a common ancestor at about the same time [8]. The grass family (Poaceae) diverged about 50–80 mya into the subfamilies Pooideae (tribe Triticeae containing wheat, barley, rye, Aegilops sp.), Panicoideae (tribe Maydeae containing maize), and Bambusoides (tribe Oryzeae containing rice) [9,10,11]. Maize and sorghum diverged about 16.5 mya [12]; wheat and barley diverged about 10–15 mya [9], with wheat and rye diverging about 7 mya [13]. The cytoplasms of T. aestivum, T. temopheevii, and T. turgidum originate from an ancestor like A. speltoides [14].
It is suggested that the ancestor Aegilops speltoides species (S genome) was the donor of what became the B genome of the bread and durum wheats [15]. It is believed that A. speltoides is the B genome donor [16] and the maternal donor of polyploid wheats [17,18].
Nuclear genes have been used in molecular phylogenetic analysis. Sequence alignment of nuclear genes encoding plastid acetyl-CoA carboxylase (ACCase) and plastid 3-phosphoglycerate kinase (PGK) were used in molecular phylogenetic analysis of the Triticum and Aegilops species. This included A, D, and S diploids and A genome polyploids using a system based on sequences of large fragments [4,19,20]. On the other hand, receptor-like kinase, Lrk, genes were used to study hexaploid wheat evolution from its progenitors, yet the study showed high conservation in gene content and organization [21]. Therefore, molecular evolution studies over a narrow time window with highly conserved genes is not an advantage because changes in DNA sequence and rearrangements are minimal.
The chimeric open reading frame, orf256, is located upstream of coxI in fertile, cytoplasmic male sterile (CMS), and fertility restored (FR) mitochondria from Tt [22,23]. The 5' flanking sequence from −228 to −1 and the first 33 nucleotides of the coding sequence of the orf256 are identical to those of coxI of Ta, but the rest of the orf256 sequence is not related to that of coxI [22]. The orf256 sequence was detected in various species of wheat relatives and progenitors, but was expressed as RNA only in Tt and Aegilops speltoides.
Previous studies on orf256 showed some interesting features including (1) the close evolutionary history of T. aestivum and T. temopheevii, (2) the absence of orf256 in the mitochondrial DNA of T. aestivum, its presence in T. temopheevii, and the presence of a related sequence in rice, (3) the specific transcriptional and translational characteristics of orf256 depending on the source of the nucleus and the relationship to cytoplasmic male sterility, and (4) the lack of a known function for orf256. This gives a good opportunity to follow changes in its sequence, its location, its rearrangement, and its presence or absence in Triticum and Aegilops species. These molecular characteristics of orf256 suggest that this is a rapidly changing gene and make it a suitable molecular handle for evolutionary studies. In this study, orf256 was used as a molecular tool to establish a DNA fingerprint and phylogenetic relationship among Triticum and Aegilops species and their evolutionary changes.
2. Experimental Section
2.1. DNA Primer
Various specific primers (Table 1) were designed on the sequence of orf256 gene to cover different parts of the gene [24].
Table 1.
Nucleotide sequence of primers that were used to detect the orf256 sequence using PCR. Primers with bold face font gave common positive PCR results with all tested species.
| Primer Name | Sequence 5' 3' 3' |
|---|---|
| 1.5: −94 | CCA TAT TCA CGC AAC TGA T |
| 2.5: −215 | CTA CGA GAT CAC CTT CAC G |
| 3.5: −190 | CTG AGC CTT TAC GAG CAG G |
| 4.5: 35 | GCA GGT TTA CTG CTT TC |
| 5.5':253 | CTGAGCCTTTACGAGCAGG |
| 6.C:785 | TCA GAA TTA CTG AGC TAC |
| 7.C:477 | GGA ACG AAG CGC TTC ATC GA |
| 8.C: 219 | GCT TGG GGA TCC TGA ATC |
| 9.C:482 | GAG ATG CTG TTT CCC ACA AC |
| 10.C:980 | ATA GAG AGT CCC AAT ATC C |
| 11.C:1469 | GCT GTC ACT AGA ACG GAC C |
2.2. Growth of Wheat Shoots
Wheat seeds were surface sterilized [13,25]. About 20 g of clean wheat seeds were soaked for 20 hr in 100 mL of 10 ppm ampicillin (Sigma) solution. The antibiotic solution was drained off and 100 mL of 0.1% silver nitrate (w/v) was added. Seeds were shaken vigorously for 10 min, and the silver nitrate solution was replaced with 100 mL of 0.5% of NaCl solution. After 10 min of vigorous shaking, seeds were rinsed three times with sterile, deionized water. Sterilized seeds were spread on 0.1% water agar in a sterilized plastic container and kept in the dark for 7–10 days at room temperature. Shoots were harvested and used directly for mitochondrial isolation or freeze dried for genomic DNA isolation.
2.3. Freeze drying of Wheat Shoots
10-day-old shoots were freeze dried in (Alpha 1–2 LO plus Christ, Vacuubrand, Germany). Dried shoots were ground in a coffee grinder to fine powder and used for DNA isolation.
2.4. Isolation of Mitochondria
Wheat mitochondria were isolated according to Song and Hedgcoth [26]. Mitochondrial pellets were stored at −20 °C.
2.5. Isolation of DNA
DNA was isolated from 10-day-old wheat shoots and or wheat mitochondria of Triticum and Aegilops species (Table 2). DNA was isolated using plant DNA isolation kit (Qiagen, California, USA) following manufacturer instructions. DNA concentration was estimated and used as PCR template. DNA samples were visualized on 1–2% agarose.
Table 2.
Triticum and Aegilops species that were used in this study.
| Species | Description | Ploidy level | Notes |
|---|---|---|---|
| Triticum aestivum (Ta) | Hexaploid wheat | AABBDD | |
| Triticum timopheevii (Tt) | wheat progenitor | GGAA | Possible source of B (G = B) genome |
| Triticum turgidum (Ttu) | wheat progenitor | BBAA | S = B |
| Triticum monococcum monocuccom (Tm) | wheat progenitor | AA | Source of A genome |
| Triticum monocuccum bioeticum (Tb) | wheat progenitor | AA | Source of A genome |
| Aegilops speltoides (Asp) | wheat progenitor | SS | S = B |
| Aegilops bicornis (Ab) | wheat progenitor | SS | |
| Aegilops searsii (Ase) | wheat progenitor | SS | |
| Aegilops taushii (At) | wheat progenitor | DD | Source of D genome |
| Aegilops sharonensis (Ash) | wheat progenitor | SS | |
| Secale cereale (Sc) | wheat relative |
2.6. PCR Amplification
Polymerase Chain Reaction (PCR) was used to amplify diagnostic fragments of orf256 using different combinations of primers. PCR was undertaken in 50 µL total volume containing 5 µL of 10X PCR buffer, 4 µL 25 mM MgCl2, 1 µL (10 ng) of DNA, 1 µL (100 ng, 125 picomole) of each primer (forward and reverse), 1 U of Taq DNA polymerase. PCR amplification conditions were initial denaturation at 95 °C for 5 min, denaturation at 95 °C for 1 min, annealing at 50 °C for 30 sec for 35 cycles, extension at 72 °C 1 min, and final extension at 72 °C for 5 min.
2.7. Sequencing of the PCR Amplified Fragments
The common PCR fragments obtained were amplified in all Triticum and Aegilops species, especially T. turgidum, and were purified and sequenced [24]. Ten samples were sequenced for each species to eliminate the heteroplasmy possibility of mitochondrial genomes.
2.8. Sequence Alignment and Phylogenetic Relationship
The obtained DNA sequences of the orf256 amplified fragments were aligned using CLUSTALW [27]. The phylogenetic relationship among Triticum and Aegilops species was established using PHYLIP program on the Pasteur Institute Server [28].
3. Results and Discussion
3.1. PCR
PCR was used to amplify DNA fragments from Triticum and Aegilops species. Using primer pair 5:-94/C: 482 (Table 1), PCR product of 576 bp was amplified (Figure 1) including Triticum turgidum, whereas using primer pair 5':253 and C: 482 resulted in the amplification of 230 bp fragment in all species tested (Figure 2). Other primer combinations (Table 1) amplified various fragments from different species except Triticum turgidum; therefore, we limited the comparison to these two fragments.
Figure 1.
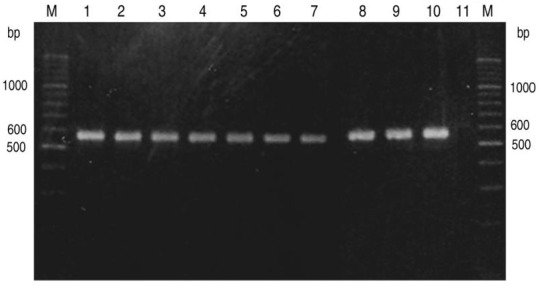
PCR product (576 bp) amplified using primer pair 5': −94 and C: 482. M: 100 bp DNA ladder; 1: Tt, 2: Ttu, 3: Tm, 4: Tb, 5: Asp, 6: Ab, 7:Ase, 8: At, 9: Ash, 10:Sc, 11: Ta. Full scientific names are shown in Table 2.
Figure 2.
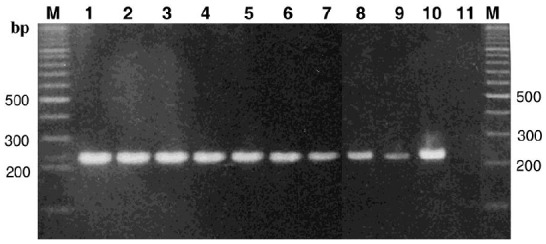
PCR product (230 bp) amplified using primer pair 5':253 and C: 482. 1: Tt, 2: Ttu, 3: Tm, 4: Tb, 5: Asp, 6: Ab, 7: Ase, 8: At, 9: Ash, 10:Sc, 11: Ta. Full scientific names are shown in Table 2.
3.2. Sequencing and Bioinformatic Analysis of PCR Products
The large PCR fragment (576 bp) obtained with primers 5': −94 and C:482 was cleaned and sequenced from the nine Triticum and Aegilops species. The nine DNA sequences obtained were used for multiple alignment using ClustalW2 (Figure 3). Multiple alignments revealed many differences among the nine sequences used in this study. Generally, the 5' third of the aligned sequences showed the most drastic and significant differences, including cluster of deletions or single deletions in some species as well as base substitutions. The middle part of the sequence has fewer changes, whereas the 3' third is more conserved among the species under study. Alignment of 500 bp showed various numbers of base substitutions compared to Secale cereale (out of group species) (Table 3). It showed 49 base substitutions in T. temopheevii and T. turgidum; 50 base substitutions in T. monococcum; 47 base substitutions in T. boeoticum, A. speltoides, A. bicornis; 25 base substitutions in A. searsii; 22 base substitutions in A. tauschii; and 21 base substitutions in A. sharonensis (Table 3).
Figure 3.
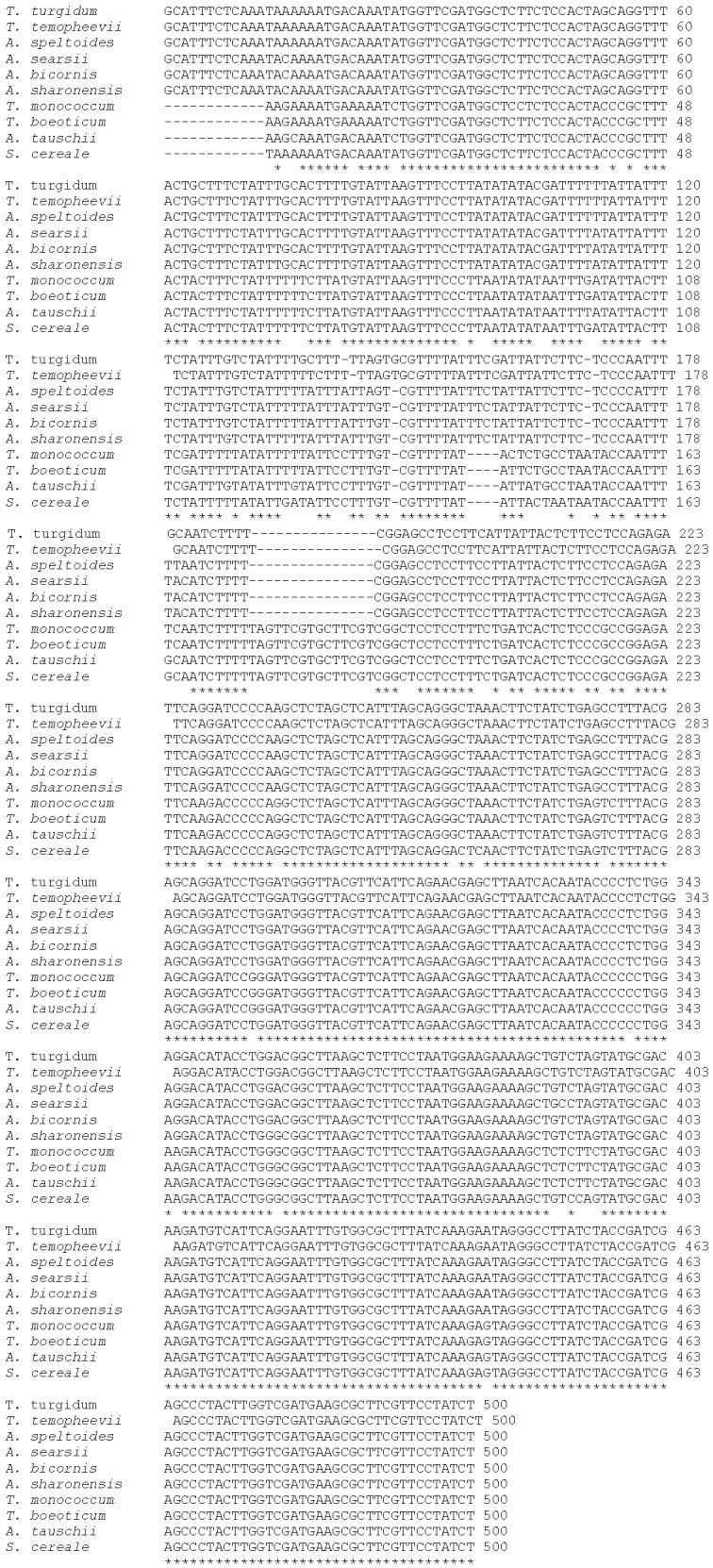
Multiple alignment of 500 bp of orf256 of Triticum and Aegilops species.
Table 3.
Summary of PCR product size obtained and the number of base substitutions in Triticum and Aegilops species used in this study compared to S. cereal sequence.
| Sequence obtained | Species | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Ttu | Tt | Ase | Ab | Ash | Asp | Tm | Tb | At | Sc | ||
| Primers | 5: −94/C:482 | 576 | 576 | 576 | 576 | 576 | 576 | 576 | 576 | 576 | 576 |
| 5:253/C:482 | 230 | 230 | 230 | 230 | 230 | 230 | 230 | 230 | 230 | 230 | |
| Number of base substitutions | 49 | 49 | 50 | 47 | 47 | 47 | 25 | 22 | 21 | - | |
3.3. Phylogenetic Analysis
The longest orf256 sequence obtained from T. turgidum is 576 bp using primer pairs 5: −94/C: 482. Only 500 bp were used (76 bp were eliminated) because of gaps to establish a consensus phylogenetic tree. The tree was established using PHYLIP software on the Pasteur Institute website [28]. The consensus tree was calculated by the UPGMA method. Bootstrap values were calculated as percentages of 1000 trials. Secale cereale was used as outgroup species. Six data sets were included in the calculation of the consensus tree using the nine species. Set one included species A. speltoides, A. sharonesis, A. bicornis. A. searsii, T. temopheevii, T. turgidum. Set two included species A. tauschii, T. boeoticum, and T. monococcum. Set three included species T. temopheevii and T. turgidum. Set four included species T. boeoticum and T. monococcum. Set five included species A. speltoides, A. sharonesis, A. bicornis, A. searsii. Set six included species A. sharonesis, A. bicornis, and A. searsii.
The consensus tree was established by making one thousand trials (Figure 4). The tree has two clades, A and B. Clade A which has the same location (bootstrap) in one thousand trials contains two branches, C and D. Branch C contains one species; A. speltoides and sub-branch E which contains three species A. searsii, A. bicornis, A. sharonesis. Branch D contains two species; T. temopheevii and T. turgidum. Clade B that has the same location (bootstrap) in 975 trials has one species; A. tauschii and one sub-branch F which included two species; T. monococcum, T. boeoticum (Figure 4).
Figure 4.
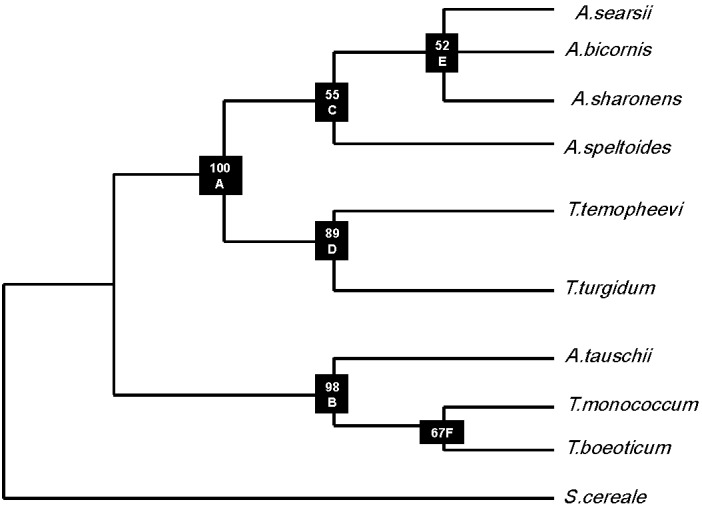
Consensus phylogenetic tree of Triticum and Aegilops species based on the common 500 bp of orf256 sequence and one thousand trials. Bootstraps (the numbers on the branches) indicate the number of times the partition of the species into the two sets, which are separated by that branch, occurred among the trees, out of 999.99 trees.
3.4. Distance Matrix
DNA distances among studied species were calculated using DNAbars software on the Pasteur Institute website [28,Table 4]. T. temopheevii and T. turgidum were the closest species with DNA distance about 0.2. Also, minimum distances occurred between A. serseaii and A. bicornis, A. bicornis and A. sharonesis, T. monococcum and T. boeoticum with distances of 0.2, 0.2, and 0.4 respectively. The highest distance was between T. temopheevii and A. speltoides to A. tauschii with DNA distance of 33.78. S. cereale is the outgroup species. T. temopheevii and A. speltoides were the most separated species of S. cereal with DNA distance of 32.54, although they were located on different sub-branches of the consensus phylogenetic tree, whereas T. boeoticum and A. taushii were the closest species to S. cereale with DNA distance of 4.11, although they were located on different sub-branches of the consensus phylogenetic tree.
Table 4.
DNA distances among Triticum and Aegilops species (1000 trials). Distance Matrix was calculated using the Jukes-Cantor correction method. Base positions 123 in the codon and gap weighting 0.0 were used.
| Species | Ttu | Tt | Ase | Ab | Ash | Asp | Tm | Tb | At | Sc |
|---|---|---|---|---|---|---|---|---|---|---|
| T.turgidum | 0.00 | |||||||||
| T.temopheevii | 0.20 | 0.00 | ||||||||
| A.searsii | 3.06 | 2.85 | 0.00 | |||||||
| A.bicornis | 2.85 | 2.65 | 0.20 | 0.00 | ||||||
| A.sharonensis | 3.06 | 2.85 | 0.40 | 0.20 | 0.00 | |||||
| A.speltoides | 2.44 | 2.23 | 1.41 | 1.21 | 1.41 | 0.00 | ||||
| T.monococcum | 32.85 | 33.16 | 33.16 | 32.85 | 32.54 | 33.16 | 0.00 | |||
| T.boeoticum | 32.85 | 33.16 | 32.85 | 32.54 | 32.23 | 33.16 | 0.40 | 0.00 | ||
| A.tauschii | 33.47 | 33.78 | 33.47 | 33.16 | 32.85 | 33.78 | 1.82 | 1.41 | 0.00 | |
| S.cereale | 32.23 | 32.54 | 32.23 | 31.93 | 31.62 | 32.54 | 4.53 | 4.11 | 4.11 | 0.00 |
T. turgidum gave negative results with other primer pair combinations (Table 1). Results previously obtained from other studies also suggested that this species has a partial orf256 sequence [13]. Common 500 bp were used to study the similarity among the nine different Triticum and Aegilops species. The phylogenetic relationship among studied species, although different, was still consistent with results obtained from previous studies which used other plastid and nuclear genes. In a study using 3 phosphoglycerate kinase (pgk-1) gene of Triticum and Aegilops, results revealed that some species showed similarity of location on the phylogenetic tree. T. temopheevii and T. turgidum along with T. aestivum (not included in this study since it does not have orf256 in its mitochondria DNA) showed closer location on the phylogenetic tree using acetyl-coA carboxylase (ACC-1) and 3-phosphoglycerate kinase (PGK-1) [4,29]. They were mapped on one sub-branch (sub-branch D, Figure 4). A. speltoides showed independent location from other Aegilops or Triticum species using the same genes (PGK-1) [4,29]. The present study showed a similar pattern because it is positioned on a separate branch with bootstrap of 55 (Figure 4). A. searsii, A. bicornis, and A. sharonesis showed a closer location in the present study. They were located on sub-branch E with bootstrap 52 (Figure 4). They showed similar relatedness [4] using 3-phosphoglycerate kinase (PGK-1) while they did not show this close proximity on the phylogenetic tree using acetyl-coA carboxylase (ACC-1) and 3-phosphoglycerate kinase genes (PGK-1) [29]. Triticum monococcum and T. boeoticum also showed close proximity location on phylogenetic trees using 3-phosphoglycerate kinase (PGK-1) [29], yet they did not show this close proximity on phylogenetic trees using the same gene ((PGK-1) [4]. T. taushii is located on an independent branch in this study (Figure 4), but in other studies, it showed close proximity with T. aestivum using ACC-1 and PGK-1 genes [4,29]. A partial sequence of WAG-2 gene was used to study the molecular evolution of wheat and its relatives. Marked variations were reported in single nucleotide polymorphisms (SNIPS) and indel numbers. Similar topology of phylogenetic trees using WGA-2 gene and the orf256 genes were obtained. For example, A. tauchii was located on one separate clade (clade III) using the WGA-2 gene. A similar location on the phylogenetic tree was obtained using orf256 gene (30). A. speltoides and T. turgidum had close phylogenetic topology on trees established using WGA-2 and orf256 genes. This supports the established idea that A. taushii (DD) is the source of D genome of Triticum aestivum (AABBDD).
From the data obtained from this study and similarities of our results with results obtained using other nuclear and plastid genes, it can be concluded that the orf256 represents a suitable molecular tool to study the relationship among Triticum and Aegilops species. Also, this introduces one more mitochondrial gene to study bioinformatic relationships among species with complex genomes which could lead to resolving their evolution at the molecular level. Orf256 and other genes could be used in monitoring gene transfer among cellular organelles, especially the nucleus and mitochondria, and tracking their evolutionary changes.
4. Conclusions
Wheat mitochondrial gene orf256 was used to study the phylogenetic and the evolutionary relationship among Triticum and Aegilops species. The results obtained were consistent with those obtained using plastid and nuclear genes. Also, the phylogenetic tree obtained from this study gave similar locations to many Triticum and Aegilops species which used plastid and nuclear genes. Data conclude that the orf256 gene of wheat mitochondrial DNA is a good molecular tool to study bioinformatic analysis of Triticum and Aegilops genomes.
Acknowledgments
The work was supported by Taif University; grant number 1-432-748.
References
- 1.Cox T.S. Deepening the wheat gene pool. J. Crop Prod. 1998;1 doi: 10.1300/J144v01n01_01. [DOI] [Google Scholar]
- 2.Dvorak J., DiTerlizzi P., Zhang H.B., Resta P. The evolution of polyploid wheats: Identification of the A genome donor species. Genome. 1993;36:21–31. doi: 10.1139/g93-004. [DOI] [PubMed] [Google Scholar]
- 3.Wheat Genetic Resource Center. [(accessed on 19 May 2010)]. Available online: www.ksu.edu/wgrc.
- 4.Huang S., Sirikhachornkit A., Su X., Faris J., Gill B., Haselkorn R., Gornicki P. Phylogenetic analysis of the acetyl-CoA carboxylase and 3-phosphoglycerate kinase loci in wheat and other grasses. Plant Mol. Biol. 2002;48:805–820. doi: 10.1023/a:1014868320552. [DOI] [PubMed] [Google Scholar]
- 5.McFadden E.S., Sears E.R. The origin of Triticum speltoides and its free-threshing hexaploid relatives. J. Hered. 1946;37:81–89. doi: 10.1093/oxfordjournals.jhered.a105590. [DOI] [PubMed] [Google Scholar]
- 6.Kimber G., Sears E.R. Evolution in the Genus Triticum and the Origin of Cultivated Whea. In: Heyne E.G., editor. Wheat and Wheat Improvement. American Society of Agronomy; Madison, WI, USA: 1987. pp. 154–164. [Google Scholar]
- 7.Friebe B., Gill B.S. Methods of Genome Analysis in Plants. CRC Press; Boca Raton, FL, USA: 1996. Chromosome Banding and Genome Analysis in Diploid and Cultivated Polyploid Wheats; pp. 39–60. [Google Scholar]
- 8.Sasanuma T., Miyashita N.T., Tsunewaki K. Wheat phylogeny determined by RFLP analysis of nuclear DNA. 3. Intra- and interspecific variations of five Aegilops Sitopsis species. Theor. Appl. Genet. 1996;92:928–934. doi: 10.1007/BF00224032. [DOI] [PubMed] [Google Scholar]
- 9.Wolfe K.H., Gouy M., Yang Y.W., Sharp P.M., Li W.H. Date of the monocot-dicot divergence estimated from chloroplast DNA sequence data. Proc. Natl. Acad. Sci. USA. 1989;86:6201–6205. doi: 10.1073/pnas.86.16.6201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Doeble J., Durbin M., Golenberg E.M., Clegg M.T., Ma D.P. Evolutionary analysis of the large subunit of carboxylase (rbcL) nucleotide sequence among the grasses (Gramineae) Evolution. 1990;4:1097–1108. doi: 10.1111/j.1558-5646.1990.tb03828.x. [DOI] [PubMed] [Google Scholar]
- 11.Clark L.G., Zhang W., Wendel J.F. A phylogeny of the grass family (Poaceae) based on sequence data. Syst. Bot. 1995;20:436–460. [Google Scholar]
- 12.Gaut B.S., Doeble J. DNA sequence evidence for the segmental allotetraploid origin of maize (duplicated lociychromosomal evolution) Proc. Natl. Acad. Sci. USA. 1997;94:6809–6814. doi: 10.1073/pnas.94.13.6809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hedgcoth C., El-Shehawi A.M., Wei P., Clardson M., Tamalis D. A chimeric open reading frame associated with cytoplasmic male sterility in alloplasmic wheat with Triticum temopheevii mitochondria is present in several Triticum and Aegilops species, barley, and rye. Curr. Genet. 2002;41:357–365. doi: 10.1007/s00294-002-0315-x. [DOI] [PubMed] [Google Scholar]
- 14.Tsunewaki K. Methods of Genome Analysis in Plants. CRC Press; Boca Raton, FL, USA: 1996. Plasmon Analysis as the Counterpart of Genome Analysis; pp. 271–299. [Google Scholar]
- 15.Dvorak J., Zhang H.B. Variation in repeated nucleotide sequences sheds light on the phylogeny of the wheat B and G genomes. Proc. Natl. Acad. Sci. USA. 1990;87:9640–9644. doi: 10.1073/pnas.87.24.9640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Tsunewaki K., Ogihara Y. The Molecular Basis of Genetic Diversity among Cytoplasms of TRITICUM and AEGILOPS Species. II. on the Origin of Polyploid Wheat Cytoplasms as Suggested by Chloroplast DNA Restriction Fragment Patterns. Genetics. 1983;104:155–171. doi: 10.1093/genetics/104.1.155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Wang G.Z., Miyashita N.T., Tsunewaki K. Plasmon analyses of Triticum (wheat) and Aegilops: PCR-single-strand conformational polymorphism (PCR-SSCP) analyses of organellar DNAs. Proc. Natl. Acad. Sci. USA. 1997;94:14570–14577. doi: 10.1073/pnas.94.26.14570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Gaut B.S. Molecular clocks and nucleotide substitution rates in higher plants. Evol. Biol. 1998;30:93–120. [Google Scholar]
- 19.Thompson J.D., Higgins D.G., Gibson T.J. CLUSTALW: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucl. Acids Res. 1994;22:4673–4680. doi: 10.1093/nar/22.22.4673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Faris J., Sirikhachornkit A., Haselkorn R., Gill B., Gornicki P. Chromosome mapping and phylogenetic analysis of the cytosolic acetyl-CoA carboxylase loci in wheat. Mol. Biol. Evol. 2001;18:1720–1733. doi: 10.1093/oxfordjournals.molbev.a003960. [DOI] [PubMed] [Google Scholar]
- 21.Feuillet C., Penger A., Gellner K., Mast A., Keller B. Molecular evolution of receptor-like kinase genes in hexaploid wheat. Independent evolution of orthologs after polyploidization and mechanisms of local rearrangements at paralogous loci. Plant Physiol. 2001;125:1304–1313. doi: 10.1104/pp.125.3.1304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Rathburn H., Hedgcoth C. A chimeric open reading frame in the 5' flanking region of coxI mitochondrial DNA from cytoplasmic male-sterile wheat. Plant Mol. Biol. 1991;16:909–912. doi: 10.1007/BF00015083. [DOI] [PubMed] [Google Scholar]
- 23.Rathburn H., Song J., Hedgcoth C. Cytoplasmic male sterility and fertility restoration in wheat are not associated with rearrangements of mitochondrial DNA in the gene regions for cob, coxII, or coxI. Plant Mol. Biol. 1993;21:195–201. doi: 10.1007/BF00039632. [DOI] [PubMed] [Google Scholar]
- 24.Macrogen Company. [(accessed on 13 June 2010)]. Available online: http://www.macrogen.com.
- 25.Speakman J.B., Krueger W. A comparison of methods to surface sterilize wheat seeds. Trans. Br. Mycol. Soc. 1983;80:374–376. [Google Scholar]
- 26.Song J., Hedgcoth C. A chimeric gene (orf256) is expressed as protein only in cytoplasmic male-sterile lines of wheat. Plant Mol. Biol. 1994;26:535–539. doi: 10.1007/BF00039566. [DOI] [PubMed] [Google Scholar]
- 27.CLUSTALW2 Software. [(accessed on 22 April 2011)]. Available online: http://www.ebi.ac.uk/Tools/msa/clustalw2/
- 28.PHLIP software. [(accessed on 15 June 2011)]. Available online: http://bioweb2.pasteur.fr/phylogeny.
- 29.Huang S., Sirikhachornkit A., Su X., Faris J., Gill B., Haselkorn R., Gornicki P. Genes encoding plastid acetyl-CoA carboxylase and 3-phosphoglycerate kinase of Triticum/Aegilops complex and the evolutionary history of polyploid wheat. Proc. Natl. Aca. Sci. USA. 2002;99:8133–8138. doi: 10.1073/pnas.072223799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wei S., Peng Z., Zhou Y., Yang Z., Wu K., Ouyang Z. Nucleotide diversity and molecular evolution of the WAG-2 gene in common wheat (Triticum aestivum L) and its relatives. Genet. Mol. Biol. 2011;34:606–616. doi: 10.1590/S1415-47572011000400013. [DOI] [PMC free article] [PubMed] [Google Scholar]