

Significance
Recent studies have identified the genetic basis of numerous traits that differentiate modern domestic species from their wild counterparts. In both plants and animals, traits (and the genes underlying them) found ubiquitously in modern breeds are often presumed to have been selected early during the domestication process. Here, by determining genetic variability in ancient European chickens over the past 2,000 years, we show that a mutation thought to be crucial during chicken domestication was not subjected to strong human-mediated selection until much later in time. This result demonstrates that the ubiquity of mutations, which differentiate modern wild and domestic taxa, does not necessarily imply ancient origins.
Keywords: selective sweep, breed formation, animal domestication, Gallus gallus, cultural history
Abstract
Modern domestic plants and animals are subject to human-driven selection for desired phenotypic traits and behavior. Large-scale genetic studies of modern domestic populations and their wild relatives have revealed not only the genetic mechanisms underlying specific phenotypic traits, but also allowed for the identification of candidate domestication genes. Our understanding of the importance of these genes during the initial stages of the domestication process traditionally rests on the assumption that robust inferences about the past can be made on the basis of modern genetic datasets. A growing body of evidence from ancient DNA studies, however, has revealed that ancient and even historic populations often bear little resemblance to their modern counterparts. Here, we test the temporal context of selection on specific genetic loci known to differentiate modern domestic chickens from their extant wild ancestors. We extracted DNA from 80 ancient chickens excavated from 12 European archaeological sites, dated from ∼280 B.C. to the 18th century A.D. We targeted three unlinked genetic loci: the mitochondrial control region, a gene associated with yellow skin color (β-carotene dioxygenase 2), and a putative domestication gene thought to be linked to photoperiod and reproduction (thyroid-stimulating hormone receptor, TSHR). Our results reveal significant variability in both nuclear genes, suggesting that the commonality of yellow skin in Western breeds and the near fixation of TSHR in all modern chickens took place only in the past 500 y. In addition, mitochondrial variation has increased as a result of recent admixture with exotic breeds. We conclude by emphasizing the perils of inferring the past from modern genetic data alone.
The resolution afforded by multiple genetic loci and—more recently—complete genomes has led to an increased understanding of the pattern and process of plant and animal domestication (1, 2). More specifically, genetic analyses have uncovered selective sweeps, quantitative trait loci, and even causative mutations underlying a wide range of behavioral and morphological traits, some of which define specific breeds, and others that differentiate domestic plants and animals from their wild ancestors (1, 3, 4).
Because many of these traits are present in either single or relatively few closely related modern breeds, the earliest occurrences of specific phenotypes (and the underlying causative mutations) are presumed to have occurred well after the initial domestication process. These phenotypes are referred to (at least in the plant genetic literature) as “improvement genes” (2). In animals, these traits include hairlessness in Mexican and Peruvian dogs (5), dorsal hair ridges in Vietnamese, Thai, and Rhodesian Ridgebacks (6), excessive skin folds in western Shar-Peis (7), double muscling in two cattle breeds (8), and a curly coat mutation found in Selkirk Rex cats (9), none of which are thought to have been present during early domestication.
Some causative mutations, however, underlie traits found in numerous, distantly related breeds. Alleles that are fixed in domestic variants—and often presumed to have been under selection at the outset of domestication—are referred to in both the plant (2) and animal (3) domestication literature as “domestication loci” (or domestication genes). In some cases, including gray coloring (10) and altered gaits in horse breeds (11), brachycephaly in dogs (12), and muscle growth in pigs (13), no hypotheses have been proposed for the time-frame of first appearance of these traits. In others, however, the commonality of both small size (14, 15) and chondrodysplasia (16) across modern dog breeds and the widespread occurrence of pea-combs in chickens (17), led the authors of these studies to suggest that the genetic mutations underlying these characteristics were selected for during the early stages of the domestication process. More recently, a whole-genome resequencing study that compared variation in 14 unrelated dog breeds and wolves identified 36 regions potentially targeted during early domestication and included 10 genes that allowed dogs to better digest starches (18). Because increased amylase activity was ubiquitous in dogs but absent in wolves, the authors concluded that this change must have occurred when early dogs began adapting to a starch-rich diet provided by early farmers.
Recent genetic and archaeological research has also shed light on domestic chickens and their primary ancestor, the Red Junglefowl (Gallus gallus) (19). Based on archaeological bones identified from Neolithic sites in the Yellow River basin, chickens were thought to have been domesticated as early as 6000 B.C. (20). This conclusion has recently been questioned, however, because bones presumed to originate from chickens in the original faunal analysis (21, 22) have since been shown to be pheasants (23, 24). As a result, a reevaluation of all of the early finds is necessary to establish the true chronology and geography of chicken domestication.
Genes that differentiate modern domestic chickens from Red Junglefowl include those that underlie the yellow skin phenotype present in the vast majority of Western, commercial chicken breeds, as well as numerous geographically restricted and fancy breeds. Yellow skin is caused by a recessive allele of the BCDO2 (β-carotene dioxygenase 2) gene (25). BCDO2 encodes the β-carotene dioxygenase 2 enzyme that cleaves colorful carotenoids into colorless apocarotenoids (26). Although the expression of the dominant allele in skin tissue results in white skin color, the recessive allele possesses one or more cis-acting and tissue-specific regulatory mutations that inhibit expression of BCDO2 in skin tissue. Provided that sufficient carotenoids are available in the diet, the recessive allele reduces carotenoid cleavage and allows them to be deposited in skin tissue, leading to yellow skin (25). This recessive BCDO2 allele is thought to have been acquired through hybridization with the Gray Junglefowl (Gallus sonneratii) in South Asia (25). Red and Gray Junglefowl are known to hybridize in contact zones in the Indian subcontinent (27, 28), and it is possible that domestic poultry engaged in the same behavior after they were introduced from Southeast Asia. Given the ubiquity and genomic signatures of strong human-driven selection of the yellow skin trait in modern, Western commercial chickens (29), Eriksson et al. (25) suggested that this trait was favored by humans after chickens acquired the trait in South Asia, but before the first wave of domesticated chickens arrived in Europe between 900 and 700 B.C. (30, 31).
In addition, a recent analysis of pooled wild and domestic chicken samples revealed strong selection signatures across a number of loci, as well as a missense mutation in the thyroid-stimulating hormone receptor (TSHR), a locus possibly linked to shifts in seasonal mating (29). Given its ubiquity in domestic breeds (264 of 271 birds representing 36 global populations were homozygous for the sweep allele; the remaining 7 were heterozygous) and the general absence of the derived allele in Red Junglefowl, the authors of that study concluded that the TSHR locus may have played a crucial role during chicken domestication (29).
Here, we investigate whether the TSHR gene was selected for during the early stages of chicken domestication (29), and if early poultry keepers favored the BCDO2 gene that underlies yellow skin in chickens soon after it was acquired from the Gray Junglefowl (25, 29). To do so, we genotyped SNPs linked with the sweep alleles in both TSHR and BCDO2 in 80 ancient European chickens dating from ∼280 B.C. to the 18th century A.D. (Table S1 and SI Materials and Methods). If TSHR played a critical role during the domestication process, all of the samples analyzed here should have been fixed for the derived TSHR allele, as has been demonstrated in worldwide modern chicken populations (29). Similarly, if BCDO2 and the yellow skin phenotype was favored and maintained soon after its introgression from Gray Junglefowl, a significant proportion of the ancient European individuals should also possess this phenotype. Finally, we assess the hypothesis that the presence of mitochondrial DNA (mtDNA) control region (CR) haplogroups A–D has resulted from the recent introduction of East Asian chickens into the European gene pool, and that haplogroup E is historically associated with European chickens (32).
Results
For each ancient individual, we attempted to amplify a 58-bp fragment surrounding the candidate missense (Gly > Arg) SNP in the TSHR gene (29), a 51-bp fragment surrounding a SNP in the BCDO2 gene associated with the yellow skin allele (SNP B in table 1 of ref. 25), and a 201-bp fragment of the mtDNA CR (33). Overall, 55 of 80 (69%) ancient chicken remains provided reproducible results for at least one of three loci (Fig. 1, Fig. S1, and Table S1). We observed allelic drop out in a number of heterozygous specimens for both TSHR and BCDO2. However, we estimated the probability of falsely assigning a true heterozygous individual as a homozygote to <0.01. In addition, external replication on 12 samples yielded identical mtDNA CR and TSHR sequences (SI Materials and Methods).
Fig. 1.

(A) A map showing the locations and chronology of archaeological sites (black dots) from three different European regions where ancient chicken remains analyzed in this study were excavated (see also SI Materials and Methods and Fig. S1). The pairs of colored boxes adjacent to each region show which of two alleles of two nuclear genes (TSHR and BCDO2) were present in each genotyped individual (Table S1). (B) Pie charts showing the differing allele frequencies of all (pooled) ancient samples (from 290 B.C. to the 18th century A.D.) (Table S2). Allele frequencies found in modern chicken populations at the TSHR locus were derived from ref. 29 and at the BCDO2 locus from ref. 25.
Among the 44 specimens from whom the TSHR locus was successfully genotyped, 8 individuals were homozygous for the derived (domestic) sweep allele, 14 were homozygous for the wild-type allele, and 22 specimens were heterozygous (Fig. 1 and Table S1). The results of a Fisher’s exact test revealed that the sweep allele was significantly less frequent in the ancient sample than in modern chickens (P < 0.0001). A binomial probability test demonstrated that observing these frequencies among the ancient samples, assuming the frequencies of the modern samples (Fig. S2 and Table S2) (29), is very unlikely (P < 0.0001). In addition, a χ2 test on TSHR genotype frequencies derived from the second to third century A.D. Quintana/Künzing Roman population (Fig. 1 and Tables S1 and S3) revealed that the observed frequencies are consistent with Hardy–Weinberg equilibrium (P > 0.95, χ2 = 0.004, df = 1), although one group had fewer than the recommended minimum number of expected genotypes/individuals.
Of the 25 ancient specimens successfully genotyped for the BCDO2 locus, 20 were homozygous for the white skin allele (found in Red Junglefowl) and five individuals were heterozygous (Fig. 1 and Table S2). Because only one functional copy of the BCDO2 gene is necessary to effectively cleave carotenoids, the yellow skin phenotype can only be expressed in chickens that are homozygous for the yellow skin allele and consume sufficient carotenoids in their diet. Of the 25 successfully genotyped chicken samples, none could express the yellow skin phenotype. Because genotype/phenotype frequencies reported previously were selected on the basis of their phenotypes (nonrandom sampling) (25), we did not carry out statistical comparisons of allele frequencies between ancient and modern populations.
The targeted mtDNA CR fragment was successfully sequenced in 38 individuals (Table S1). The topology of a maximum-likelihood tree constructed from an alignment of 201-bp haplotypes matched the neighbor-joining tree generated by Liu et al. (19), confirming previous observations that this specific 201-bp fragment is sufficient for recovering the major clades present in the chicken mitochondrial tree (Fig. 2 and Fig. S3) (33, 34). We identified a total of three haplotypes among the ancient specimens, all of which clustered within the E clade on the chicken mitochondrial tree (19, 35) (Fig. 2 and Table S4). The E3 (n = 1) and E6 (n = 2) haplotypes (19) were present only in Medieval and post-Medieval chickens from England (Tables S1 and S4), whereas the remaining 35 individuals possessed a 201-bp haplotype corresponding to haplotypes E1, E5, E12, E15, or E16 described using a 519-bp fragment (Table S4) (19). We find a significant difference in haplogroup frequencies (pooled into two groups of chickens: those belonging to haplogroup E and those belonging to haplogroup A–D) between the ancient and modern datasets (Fisher's exact test, P < 0.002) (Table S5). Assuming the frequency reported for modern European chickens (Table S5) (i.e., ∼15% of modern European chickens possess haplotypes from clades A–D), a binomial test revealed that the probability of observing only the E haplogroup in 43 ancient specimens (the unique 38 sequences combined with previously published data) (Table S5) is <0.001.
Fig. 2.
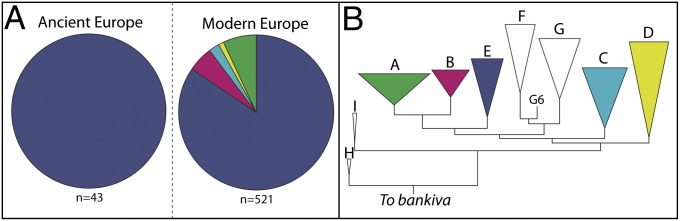
(A) Two pie charts showing differences in mtDNA haplogroup frequencies between ancient and modern European chickens. The ancient sample consists of specimens sequenced in this study (n = 38) and from a previous publication (n = 5) (34) and the modern data was compiled from (19, 32, 39) (Table S5). The colors in A correspond to haplogroups depicted in a phylogenetic tree in B. (B) A maximum-likelihood phylogenetic tree constructed using 519-bp haplotypes defined in ref. 19. White (or colorless) clades were not identified in either modern or ancient European chickens. A more detailed maximum-likelihood tree, including detailed description of methods and results (including nodal support values), is described in the SI Materials and Methods (see also Fig. S3).
Discussion
TSHR Domestication Locus.
The locus encoding TSHR on chromosome 5 in domestic chickens has recently been shown to have undergone a massive selective sweep (29). A nonconservative amino acid substitution (a missense mutation Gly558Arg) was identified as a potential causal and target mutation for the selective sweep. Although the function (and corresponding phenotype) associated with the derived allele remains unknown, it is possible that this gene variant affects photoperiod control and the absence of strict seasonal reproduction, a trait commonly found in domestic animals but rare or absent among their wild relatives (29). Because 264 of 271 modern birds representing 36 globally distributed populations were homozygous for the derived sweep allele (the seven remaining were heterozygous), and because the Red Junglefowl individuals that also possessed the mutation were thought have acquired it from domestic chickens, Rubin et al. (29) suggested that TSHR was a domestication locus. If this selective sweep occurred during the early phase of domestication, all ancient chickens that postdate this event (and are located outside the natural distribution range of Red Junglefowl, thus eliminating the potential for backcrossing with wild birds) should also possess the derived allele.
The results presented here, however, demonstrate that although the derived allele was present in European chickens dated to ∼280 B.C. to the 16th–18th century A.D., it was only found on 43% of the typed chromosomes and the wild-type allele persisted at intermediate frequencies until at least the 16th–18th century AD (Fig. 1 and Tables S1 and S2). These results suggest that the TSHR mutation was neither a prerequisite, nor critical in the immediate aftermath of chicken domestication. Although the strength of the selection pressure that drove the sweep haplotype in modern populations is not in doubt (29), the data presented here suggest that the fixation of the derived allele in European chickens was likely much closer to the present-day and certainly within the last 500 y, possibly commensurate with the improvement of farmyard animals that began during the industrial revolution (27).
In one possible scenario, the sweep allele was fixed in Western Europe and spread across the world with newly synthesized commercial breeds that emerged during the second half of the 19th century (27). This hypothesis seems overly simplistic, however, given that the TSHR allele was found to be fixed (or nearly so) in modern Egyptian Fayoumi chickens and Silkie, Cochin, and Hua-Tung chickens from China (29) that are unlikely to have been strongly influenced by European commercial breeds. It is therefore possible that the TSHR sweep allele became fixed in populations originating outside Europe well before the creation of modern breeds, and that these birds in turn replaced the initial populations introduced into Europe. This narrative is supported by the observation that the Classical Greek chickens (Kassope) were the only population analyzed in this study that was fixed for the sweep allele (Fig. 1 and Table S1). Although the earliest Central European chickens that arrived north of the Alps ∼600–400 B.C. likely descended from founder populations present in eighth to fifth century B.C. Greek settlements on the Mediterranean coast, the first century B.C. chicken population from Kassope could have arrived as part of a secondary introduction of chickens. During the Achaemenid (550–330 B.C.) and Hellenistic (323–31 B.C.) periods, the Aegean region was a commercial hub that drew trade from most of the Near and Middle East, including the Indus Valley (36). Such trade networks could have been linked to the introduction of chicken populations that had undergone a selective sweep at the TSHR locus. Crucially, however, none of the Greek individuals possessed the introgressed Gray Junglefowl BCDO2 allele (Fig. 1), suggesting these two loci were selected for and fixed at different times and places.
The BCDO2 Yellow Skin Locus.
The study that discovered the TSHR sweep (29) also confirmed a selective sweep encompassing the BCDO2 locus. Unlike TSHR, however, BCDO2 is only fixed in a limited number of (often commercial and geographically widespread) breeds (25). The yellow skin phenotype was previously shown to result from the presence of a BCDO2 allele that domestic chickens acquired not from their primary ancestor—the Red Junglefowl—but from introgression between domestic chickens and Gray Junglefowl indigenous to South Asia (25).
The data presented here reveal that although the yellow skin allele was present on ∼10% of chromosomes in ancient European chickens, not a single ancient bird was homozygous for the Gray Junglefowl allele (and therefore capable of expressing the yellow skin phenotype) (Tables S1 and S2). This finding is consistent with 17th–19th century records suggesting that a number of widespread, prolific, and economically important breeds raised in western and southern Europe, including the Dorking, Houdan, Sultan, Spanish, or B/W Bantams, clearly had white legs, but the Hamburgh, Polish, Turkish, and Crève Cœur breeds had leg colors ranging between slate blue and dark leaden-blue (27, 37). However, some 17th century European breeds, including the Padua, did possess yellow legs (37) and the trait was explicitly mentioned in relation to heavy, fast-growing types, including Cochin, Brahma, and Malay breeds imported by sea (and therefore named Captain’s birds) from the Far East into Europe and the United States during the 1820s–1850s (27). It is therefore possible that the rise in frequency of the yellow skin phenotype occurred only after recent introductions of foreign birds to Europe and the breed formation process that consequently led to the creation of modern, widespread, commercial broiler, and egg-laying breeds. Tegetmeier (27), for example, noted the remarkable rapid growth and great size attained by crossbred birds produced by mating Cochins with the large traditional French Crève Cœur, La Flèche, and Houdan breeds. The ubiquity of the yellow skin phenotype in commercial, modern, and some rare, geographically restricted breeds can, therefore, be explained by rapid worldwide spread of newly synthesized commercial chickens. This suggestion is further supported by the fact that the yellow skin phenotype appears infrequently or is completely absent in rare breeds, such as the Friesian Fowl, Houdan, and Westfälischer Totleger (25).
The Mitochondrial Control Region.
The mtDNA control region is a widely used locus in chicken genetic studies and has frequently been used to investigate domestication, admixture, and migration (19, 34, 35, 38). Although previous studies have concluded that modern European chickens (and Western commercial breeds) primarily possess mitochondrial haplotypes belonging to the E clade (19, 35), additional studies (32, 39) have demonstrated that haplogroups A–D are also present in a wide variety of breeds, most likely as a result of the intentional importation and hybridization of East Asian breeds into Europe during breed formation and breed improvement over the past 500 y (32).
Although a recent study of complete mtDNA chicken genomes revealed the existence of 14 mitochondrial clades found worldwide (35), all 38 ancient chickens sequenced in this study, and five archaeological Spanish chickens typed in a previous study (34), possessed exclusively E-clade haplotypes (which represent 3 out of the 14 major clades in ref. 35) (Tables S1 and S5). These results first demonstrate that chickens initially introduced to Europe possessed a small fraction of the variability present in Southeast Asia, where chickens were originally domesticated. Second, these results demonstrate that all haplotypes belonging to other haplogroups therefore represent recent introductions. Interestingly, because all but three ancient European chickens possessed a single haplotype (probably corresponding to the common E1 haplotype) (19) (Table S4), the presence of haplotypes E3 and E6 in Medieval and post-Medieval contexts from the United Kingdom may imply secondary introductions.
Finally, although the nuclear loci typed in this study show a dramatic reduction in variability between ancient and modern populations, the mitochondrial signatures reveal an initial uniformity followed by an increase in haplogroup diversity in modern birds. These seemingly divergent genetic patterns reflect different human goals over the last two centuries, including not only intensive selection for traits associated with behavior, production (TSHR), and skin color (BCDO2), but also for phenotypic variability in fancy breeds achieved through the importation of East Asian varieties (32).
Conclusions
A variety of genetic techniques and analytical approaches have led to the identification of selective sweeps and causative mutations that differentiate populations of domestic plants and animals from their modern wild counterparts. The ubiquity of sweep alleles across numerous breeds has often been used as an argument to infer their ancient origins, and as an argument that the SNPs and associated traits were selected for during the early phases of domestication (4, 14, 17, 18, 40). The patterns of allelic diversity presented here, however, challenge the hypothesis that modern variation can be directly mapped onto the past. Instead, the allelic variability within TSHR in ancient chickens demonstrates that this locus was not crucial for the early development of domestic chickens, and that the fixation of the derived allele (at least in European chickens) took place only in the past few hundred years. Similarly, although the yellow skin allele was present in ancient European chickens, the phenotype was very rare in the past. In addition, the fixation of the BCDO2 allele in numerous modern chicken breeds was likely also a recent occurrence, possibly coincident with recent breed formation instigated during the industrial revolution (27), and the subsequent development of modern commercial chicken breeds. This pattern demonstrates the ease with which we can underestimate the potential of alleles to become ubiquitous through rapid strong selection leading to fixation, followed by geographic proliferation through human-assisted migration.
This result is consistent with most ancient DNA studies that have revealed the hazards of assuming that modern genetic data can be used to draw accurate conclusions about past population variability. For example, a mutation in the NAM-B1 gene, associated with increased grain size in wheat, was thought (on the basis of fixation in modern cultivars) to have been selected for during the early phases of domestication (40). A genetic survey of 19th century historical seeds, however, revealed that fixation of the modern, sweep allele occurred only recently during crop improvement (41). A similar study of ancient maize concluded that, although two genes (tb1 and pbf) had been fixed for the domestic variant by ca. 4,000 y ago, one gene (su1) still possessed significant variability as long ago as ca. 2,000 y ago (42). Similarly, a number of studies of ancient mtDNA have shown that lineage replacement (often rapid and geographically widespread) was common among both domestic (43, 44) and wild (45, 46) populations.
The processes of recent breed formation, coupled with strong directed selection and admixture with exotic breeds, have rapidly and radically shaped the gene pools of modern domestic plants and animals, often distorting their deeper genetic history (47). As a result, hypotheses regarding past population dynamics drawn solely from modern datasets require verification through direct observation. This need is especially true where zooarcheological or historical records suggest recent origins for some traits. As mentioned at the outset, conclusions drawn from the genetic variability in modern samples suggest that small size (14, 15) and chondrodysplasia (16) in dogs, and pea-combs in chickens (17), occurred early during domestication. However, small dogs (<30-cm shoulder height) do not appear in the Eurasian archaeological record until after the first millennium B.C. (30), the earliest reported evidence for chondrodysplasia is found in Egyptian tomb art dating to the late third millennium B.C. (48), and the peacomb trait in chickens was still rare in mid-19th century English fowl (27).
These lines of evidence alone do not undermine claims for much earlier appearances of these and other domestication-related traits, but they do suggest that modern genetic data should be considered cautiously and within a broader context that includes an appreciation for the potential of rapid and widespread demographic shifts. Future studies that investigate ancient genetic variability in loci known to underlie behavioral, dietary, and phenotypic differences between wild and domestic plants and animals will help to reveal the first appearance and timing of selection pressures. These data will lead to the establishment of a significantly more robust geographic and temporal interpretative framework to more fully understand the early patterns and processes of domestication.
Materials and Methods
Ancient chicken specimens were selected to represent three geographical locations and two major time bins (first, Late Iron Age, Late Hellenistic and Roman contexts, dating to ∼280 B.C. to the fifth century A.D., and second, Medieval and post-Medieval contexts, dating to approximately the 10th to 18th century A.D.) (Table S1). The majority of chicken bones come from closed contexts that have been dated either through stratigraphic (cultural) association or direct radiocarbon dating (SI Materials and Methods).
DNA extraction was performed in dedicated ancient DNA laboratories at Durham University and Uppsala University following stringent laboratory procedures according to commonly applied guidelines (49, 50). The work was carried out by laboratory personal wearing protective laboratory coats and overshoes, or coveralls and dedicated laboratory clogs, and double pairs of gloves (with the outer pairs of gloves changed in between every step of the preparation/extraction procedure). All equipment and work surfaces are routinely cleaned before and after each use with a dilute solution of bleach [5–10% (wt/vol) active sodium hypochlorite] followed by rinsing with ddH2O and ethanol [70–99% (vol/vol)]. A strict one-way system for entering the laboratories is in use to avoid carry-over of post-PCR contaminants.
DNA extractions were performed as previously described (44) (see SI Materials and Methods for a detailed account of experimental procedures). To authenticate the results, 12 chickens were independently replicated for the CR and TSHR SNP at Uppsala University (Table S1). CR sequences were amplified using previously published PCR primers GG144F-GG387R (33), and PCR and sequencing primers for TSHR and BCDO2 were designed in PSQ Assay Design (Qiagen) (Table S6). CR sequences were Sanger-sequenced in both directions at the DNA sequencing facility at Durham University. TSHR and BCDO2 PCR amplicons were genotyped on the Q24 (Qiagen) pyrosequencer in the Department of Archaeology at Durham University, United Kingdom, or on a PSQ 96MA (Biotage) pyrosequencer at Uppsala University. Full PCR cycling conditions and sequencing methods are presented in detail in the SI Materials and Methods.
MtDNA CR sequence data were analyzed and assembled in Geneious v.5.4 (51) and manually edited in Se-Al (http://tree.bio.ed.ac.uk/software/seal). Reference sequences were compiled from previous publications (19, 32) and aligned with the unique sequences deposited into GenBank (KF753251- KF753289).
Supplementary Material
Acknowledgments
We thank Anders Götherström for assistance with data replication; and the London Archaeological Archive and Research Centre, Museum of London, Alex Croom (Arbeia Roman Fort and Museum), and Deborah Jaques for sample material and discussion. Sample material from Kassope and Central European sites is courtesy of Bavarian State Collection of Anthropology and Palaeoanatomy, Munich; material from Beverley is courtesy of Humber Field Archaeology, Hull; and material from York is courtesy of Deborah Jaques. This work was supported by a Research Councils UK Academic Fellowship (to G.L.); Grant NE/F003382/1 from the Natural Environment Research Council; and Grant F/00 128/AX from the Leverhulme Trust. This manuscript resulted from a catalysis meeting entitled “Domestication as an Evolutionary Phenomenon: Expanding the Synthesis” that was awarded and hosted by the National Evolutionary Synthesis Centre, National Science Foundation #EF-0905606 in 2011.
Footnotes
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
2Centre for Genome-Enabled Biology and Medicine, University of Aberdeen, Aberdeen AB24 3RY, Scotland.
Data deposition: The sequences reported in this paper has been deposited in the GenBank database (accession nos. KF753251–KF753289).
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1308939110/-/DCSupplemental.
References
- 1.Andersson L. Molecular consequences of animal breeding. Curr Opin Genet Dev. 2013;23(3):295–301. doi: 10.1016/j.gde.2013.02.014. [DOI] [PubMed] [Google Scholar]
- 2.Olsen K-M, Wendel J-F. A bountiful harvest: Genomic insights into crop domestication phenotypes. Annu Rev Plant Biol. 2013;64:47–70. doi: 10.1146/annurev-arplant-050312-120048. [DOI] [PubMed] [Google Scholar]
- 3.Andersson L. How selective sweeps in domestic animals provide new insight into biological mechanisms. J Intern Med. 2012;271(1):1–14. doi: 10.1111/j.1365-2796.2011.02450.x. [DOI] [PubMed] [Google Scholar]
- 4.Wayne RK, vonHoldt BM. Evolutionary genomics of dog domestication. Mamm Genome. 2012;23(1–2):3–18. doi: 10.1007/s00335-011-9386-7. [DOI] [PubMed] [Google Scholar]
- 5.Drögemüller C, et al. A mutation in hairless dogs implicates FOXI3 in ectodermal development. Science. 2008;321(5895):1462. doi: 10.1126/science.1162525. [DOI] [PubMed] [Google Scholar]
- 6.Salmon Hillbertz NHC, et al. Duplication of FGF3, FGF4, FGF19 and ORAOV1 causes hair ridge and predisposition to dermoid sinus in Ridgeback dogs. Nat Genet. 2007;39(11):1318–1320. doi: 10.1038/ng.2007.4. [DOI] [PubMed] [Google Scholar]
- 7.Olsson M, et al. A novel unstable duplication upstream of HAS2 predisposes to a breed-defining skin phenotype and a periodic fever syndrome in Chinese Shar-Pei dogs. PLoS Genet. 2011;7(3):e1001332. doi: 10.1371/journal.pgen.1001332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.McPherron AC, Lee S-J. Double muscling in cattle due to mutations in the myostatin gene. Proc Natl Acad Sci USA. 1997;94(23):12457–12461. doi: 10.1073/pnas.94.23.12457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Filler S, et al. Selkirk Rex: Morphological and genetic characterization of a new cat breed. J Hered. 2012;103(5):727–733. doi: 10.1093/jhered/ess039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Rosengren Pielberg G, et al. A cis-acting regulatory mutation causes premature hair graying and susceptibility to melanoma in the horse. Nat Genet. 2008;40(8):1004–1009. doi: 10.1038/ng.185. [DOI] [PubMed] [Google Scholar]
- 11.Andersson LS, et al. Mutations in DMRT3 affect locomotion in horses and spinal circuit function in mice. Nature. 2012;488(7413):642–646. doi: 10.1038/nature11399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Bannasch D, et al. Localization of canine brachycephaly using an across breed mapping approach. PLoS ONE. 2010;5(3):e9632. doi: 10.1371/journal.pone.0009632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Van Laere A-S, et al. A regulatory mutation in IGF2 causes a major QTL effect on muscle growth in the pig. Nature. 2003;425(6960):832–836. doi: 10.1038/nature02064. [DOI] [PubMed] [Google Scholar]
- 14.Gray MM, Sutter NB, Ostrander EA, Wayne RK. The IGF1 small dog haplotype is derived from Middle Eastern grey wolves. BMC Biol. 2010;8(1):16. doi: 10.1186/1741-7007-8-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Sutter NB, et al. A single IGF1 allele is a major determinant of small size in dogs. Science. 2007;316(5821):112–115. doi: 10.1126/science.1137045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Parker HG, et al. An expressed fgf4 retrogene is associated with breed-defining chondrodysplasia in domestic dogs. Science. 2009;325(5943):995–998. doi: 10.1126/science.1173275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Wright D, et al. Copy number variation in intron 1 of SOX5 causes the Pea-comb phenotype in chickens. PLoS Genet. 2009;5(6):e1000512. doi: 10.1371/journal.pgen.1000512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Axelsson E, et al. The genomic signature of dog domestication reveals adaptation to a starch-rich diet. Nature. 2013;495(7441):360–364. doi: 10.1038/nature11837. [DOI] [PubMed] [Google Scholar]
- 19.Liu Y-P, et al. Multiple maternal origins of chickens: Out of the Asian jungles. Mol Phylogenet Evol. 2006;38(1):12–19. doi: 10.1016/j.ympev.2005.09.014. [DOI] [PubMed] [Google Scholar]
- 20.West B, Zhou B-X. Did chickens go north? New evidence for domestication. J Archaeol Sci. 1988;15(5):515–533. [Google Scholar]
- 21.Chow BS. The animal remains discovered at Cishan village, Wu’an, Hebei Province. Kaogu Xuebao. 1981;62(3):339–348. Chinese. [Google Scholar]
- 22.Chow BS. The animal remains discovered at Baiyingsite, Tangyin, Henan Province. Papers on Chinese Archaeology. 1983;3:48–50. Chinese. [Google Scholar]
- 23.Deng H, Yuan J, Song G-D, Wang C-S, Masaki E. The re-examination of the Domestic Chicken in Ancient China. Archaeology. 2013;6(6):83–96. Chinese. [Google Scholar]
- 24.Peters J. Römische Tierhaltung und Tierzucht. Eine Synthese aus archäozoologischer Untersuchung und schriftlich-bildlicher Überlieferung. Passauer Universitätsschriften zur Archäologie 5. Rahden, Westfalen, Germany: Marie Leidorf; 1998. [Google Scholar]
- 25.Eriksson J, et al. Identification of the yellow skin gene reveals a hybrid origin of the domestic chicken. PLoS Genet. 2008;4(2):e1000010. doi: 10.1371/journal.pgen.1000010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kiefer C, et al. Identification and characterization of a mammalian enzyme catalyzing the asymmetric oxidative cleavage of provitamin A. J Biol Chem. 2001;276(17):14110–14116. doi: 10.1074/jbc.M011510200. [DOI] [PubMed] [Google Scholar]
- 27.Tegetmeier WB. The Poultry Book: Comprising the Breeding and Management of Profitable and Ornamental Poultry, Their Qualities and Characteristics; To Which is added “The Standard of Excellence in Exhibition Birds,” Authorized by the Poultry. London: G. Routledge and Sons; 1867. [Google Scholar]
- 28.McGowan PJK. Phasianidae (Pheasants and Partridges) In: del Hoyo J, Elliott A, Sargatal J, editors. Handbook of the Birds of the World. Barcelona, Spain: Lynx Edicions; 1994. pp. 434–552. [Google Scholar]
- 29.Rubin C-J, et al. Whole-genome resequencing reveals loci under selection during chicken domestication. Nature. 2010;464(7288):587–591. doi: 10.1038/nature08832. [DOI] [PubMed] [Google Scholar]
- 30.Benecke N. Archäozoologische Studien zur Entwicklung der Haustierhaltung in Mitteleuropa und Südskandinavien von den Anfängen bis zum ausgehenden Mittelalter. Berlin: Akademie; 1994. [Google Scholar]
- 31.Carrasquila Hernández F. Some comments on the introduction of domestic fowl in Iberia. Archaeofauna. 1992;1:45–93. [Google Scholar]
- 32.Dana N, et al. East Asian contributions to Dutch traditional and western commercial chickens inferred from mtDNA analysis. Anim Genet. 2010;42(2):125–133. doi: 10.1111/j.1365-2052.2010.02134.x. [DOI] [PubMed] [Google Scholar]
- 33.Storey AA, et al. Radiocarbon and DNA evidence for a pre-Columbian introduction of Polynesian chickens to Chile. Proc Natl Acad Sci USA. 2007;104(25):10335–10339. doi: 10.1073/pnas.0703993104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Storey AA, et al. Investigating the global dispersal of chickens in prehistory using ancient mitochondrial DNA signatures. PLoS ONE. 2012;7(7):e39171. doi: 10.1371/journal.pone.0039171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Miao YW, et al. Chicken domestication: an updated perspective based on mitochondrial genomes. Heredity (Edinb) 2013;110(3):277–282. doi: 10.1038/hdy.2012.83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Roaf M. Cultural Atlas of Mesopotamia and the Ancient Near East. New York: Equinox, Facts on File; 1990. [Google Scholar]
- 37.Aldrovandi U. Ornithologiae Tomus Alter cum Indice Copiosissimo Variarum Linguarum. Bologna: Giovanni Battista Bellagamba; 1600. [Google Scholar]
- 38.Gongora J, et al. Indo-European and Asian origins for Chilean and Pacific chickens revealed by mtDNA. Proc Natl Acad Sci USA. 2008;105(30):10308–10313. doi: 10.1073/pnas.0801991105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Revay T, Bodzsar N, Mobegi VE, Hanotte O, Hidas A. Origin of Hungarian indigenous chicken breeds inferred from mitochondrial DNA D-loop sequences. Anim Genet. 2010;41(5):548–550. doi: 10.1111/j.1365-2052.2010.02041.x. [DOI] [PubMed] [Google Scholar]
- 40.Uauy C, Distelfeld A, Fahima T, Blechl A, Dubcovsky J. A NAC Gene regulating senescence improves grain protein, zinc, and iron content in wheat. Science. 2006;314(5803):1298–1301. doi: 10.1126/science.1133649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Asplund L, Hagenblad J, Leino MW. Re-evaluating the history of the wheat domestication gene NAM-B1 using historical plant material. J Archaeol Sci. 2010;37(9):2303–2307. [Google Scholar]
- 42.Jaenicke-Després V, et al. Early allelic selection in maize as revealed by ancient DNA. Science. 2003;302(5648):1206–1208. doi: 10.1126/science.1089056. [DOI] [PubMed] [Google Scholar]
- 43.Malmström H, et al. Barking up the wrong tree: Modern northern European dogs fail to explain their origin. BMC Evol Biol. 2008;8:71. doi: 10.1186/1471-2148-8-71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Ottoni C, et al. Pig domestication and human-mediated dispersal in western Eurasia revealed through ancient DNA and geometric morphometrics. Mol Biol Evol. 2013;30(4):824–832. doi: 10.1093/molbev/mss261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Barnes I, Matheus P, Shapiro B, Jensen D, Cooper A. Dynamics of Pleistocene population extinctions in Beringian brown bears. Science. 2002;295(5563):2267–2270. doi: 10.1126/science.1067814. [DOI] [PubMed] [Google Scholar]
- 46.Shapiro B, et al. Rise and fall of the Beringian steppe bison. Science. 2004;306(5701):1561–1565. doi: 10.1126/science.1101074. [DOI] [PubMed] [Google Scholar]
- 47.Larson G, et al. Rethinking dog domestication by integrating genetics, archeology, and biogeography. Proc Natl Acad Sci USA. 2012;109(23):8878–8883. doi: 10.1073/pnas.1203005109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Boessneck J. Die Tierwelt des Alten Ägypten. Munich: C. H. Beck; 1988. [Google Scholar]
- 49.Cooper A, Poinar HN. Ancient DNA: Do it right or not at all. Science. 2000;289(5482):1139. doi: 10.1126/science.289.5482.1139b. [DOI] [PubMed] [Google Scholar]
- 50.Gilbert MT, Bandelt HJ, Hofreiter M, Barnes I. Assessing ancient DNA studies. Trends Ecol Evol. 2005;20(10):541–544. doi: 10.1016/j.tree.2005.07.005. [DOI] [PubMed] [Google Scholar]
- 51.Drummond AJ, et al. 2011. Geneious v5.4, Available from www.geneious.com/. Accessed July 1, 2011.
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.