

Abstract
Background
Coronaviruses are the diverse group of RNA virus. From 1960, six strains of human coronaviruses have emerged that includes SARS-CoV and the recent infection by deadly MERS-CoV which is now going to cause another outbreak. Prevention of these viruses is urgent and a universal vaccine for all strain could be a promising solution in this circumstance. In this study we aimed to design an epitope based vaccine against all strain of human coronavirus.
Results
Multiple sequence alignment (MSA) approach was employed among spike (S), membrane (M), enveloped (E) and nucleocapsid (N) protein and replicase polyprotein 1ab to identify which one is highly conserve in all coronaviruses strains. Next, we use various in silico tools to predict consensus immunogenic and conserved peptide. We found that conserved region is present only in the RNA directed RNA polymerase protein. In this protein we identified one epitope WDYPKCDRA is highly immunogenic and 100% conserved among all available human coronavirus strains.
Conclusions
Here we suggest in vivo study of our identified novel peptide antigen in RNA directed RNA polymerase protein for universal vaccine – which may be the way to prevent all human coronavirus disease.
Keywords: Coronavirus, SARS-CoV, MERS-CoV, RNA directed RNA polymerase, Epitope, Universal vaccine
Background
Coronaviruses are the diverse group of virus which infects domestic animals, birds as well as human [1]. Coronaviruses are enveloped viruses which are the members of Coronaviridae family [2]. Coronaviruses have positive strand RNA genome which is approximately 26–32 kb long. The overall structures of all coronaviruses are composed of the spike (S), envelope (E), membrane (M) and nucleocapsid (N) protein. The other non-structural proteins like RNA directed RNA polymerase, helicase, 3CL like proteinases etc are produced by the cleavage of replicase polyprotein 1ab or ORF 1ab polyprotein [3].
The first coronavirus 229E was identified in 1960. Since then different types of coronaviruses have emerged. HCoV-229E, HCoV-OC43, SARS-CoV, HCoV-NL63, HCoV-HKU1 and MERS-CoV are the coronaviruses which infect human [4]. Most of the coronaviruses cause respiratory, enteric, hepatic, or neurological diseases with highly variable severity in their hosts [5]. The first two coronavirus HCoV-229E and HCoV-OC43 infects lower respiratory tract where the later SARS-CoV, HCoV-NL63, HCoV-HKU1 infect both lower and upper respiratory tract [4,6-8]. But the newly emerged MERS-CoV infects both lung and kidney that reflects how these viruses are changing their cell tropism and becoming highly pathogenic [9].
Most infections caused by human coronaviruses were relatively mild. Among all the human coronaviruses, SARS-CoV and MERS-CoV are much more deadly. The severe acute respiratory syndrome (SARS) outbreak caused by SARS-CoV in 2002 to 2003. The SARS-CoV created outbreak resulted in a total of 916 deaths among more than 8000 confirmed cases in over 30 countries [10,11]. The newly emerged MERS-CoV is now posing a great threat for human. According to WHO, 75 people have died among more than 178 confirmed cases caused by MERS-CoV [4]. Though this MERS-CoV virus was first found in Saudi Arabia 2012, now it has been emerged in UK, France, Tunisia, Spain and Italy that indicates it’s going to create another outbreak like SARS-CoV [12-14]. From 1960 to till now there is no recommended drug or vaccine for MERS-CoV infection and treatment relies on exclusively supportive care, which gives the high case-fatality rate, is not highly effective [15].
In 2003 after the discovery of SARS-CoV, there were a significant increase in research on coronavirus, but no definitive antiviral or therapeutic treatment for coronavirus infections came from these researches [16]. From the clinical experience of SARS-CoV found that a number of interventions including ribavirin with and without corticosteroids, ribavirin with protease inhibitors and interferon with corticosteroids may improve outcome. But a definitive treatment was not clearly established and the therapeutic interventions have not been evaluated in vivo[17].
The identification of therapeutics is a high priority and though there is currently no specific therapy or vaccine for human coronaviruses, this disease has been severe with a high case-fatality rate [18]. As these viruses are now becoming pathogenic and causing outbreaks, so steps have to be taken to prevent human death. Vaccination is one of the most efficient ways to prevent infectious disease [19]. Effective vaccines controlling virus spread and disease are available for a number of infections, such as smallpox, poliomyelitis, measles, mumps, rubella, influenza, hepatitis A, and hepatitis B [20]. For coronavirus, this vaccine approach is hindered by the fact that human coronavirus strains are not structurally related and they are changing rapidly by recombination [21]. Therefore, designing a universal vaccine against conserved regions for all human coronaviruses is a major challenge at present.
With the disclosure of huge sequence information, epitope based vaccine design now has become a most promising approach for viral vaccine preparation [22]. In order to prepare vaccines, computational prediction of epitopes and vaccine design can reliably aid this process to reduce time and cost. Although the epitope based vaccine design is now a familiar concept, not much work has been done in case of coronaviruses.
In this study, we design an epitope based universal vaccine which can be use to prevent all kind of human coronaviruses. For this, bioinformatics analyses of viral proteins were done for finding the conserved peptide region and for mapping the evolutionary conserved epitope. The 3D structure of RNA directed RNA polymerase was determined by threading modeling technique and a highly immunogenic, accessible and conserved epitope was identified. This epitope can be used as a universal vaccine against all human coronaviruses.
Results
RNA directed RNA polymerase is highly conserved in all human coronavirus strains
To find a conserved region, MSA by clustalW [23] and protein variability index [24] analyses were performed. No conserved region was found in case of S, E, M and N proteins (Additional file 1: Figure S1, Additional file 2: Figure S2, Additional file 3: Figure S3, Additional file 4: Figure S4 respectively). From the MSA of replicase polyprotein 1ab coronaviruses were found to be conserved in RNA directed RNA polymerase (Additional file 5: Figure S5). MSA of this RNA directed RNA polymerase region (Figure 1) and protein variability index (Figure 2) identified a 385 amino acid long conserved region among all human coronaviruses. The conserve sequence was then used to determine immunogenicity.
Figure 1.

Conserved peptide in RNA directed RNA polymerase. Multiple sequence alignment of the 46 replicase polyprotein 1ab of all coronaviruses revealed that human coronaviruses are conserved in RNA directed RNA polymerase. This alignment was visualized by Jalview 2.8 [25]. Along the alignment this tool provides a graphical (bar chart) conservation summery using 11-base scale for conservancy and BLOSUM 62 for quality. For Conservation yellow color bar and star sign indicates the full conservation. Black bars showed the consensus sequence and yellow color indicates good quality. All the colors changes according to the conservation and alignment quality.
Figure 2.
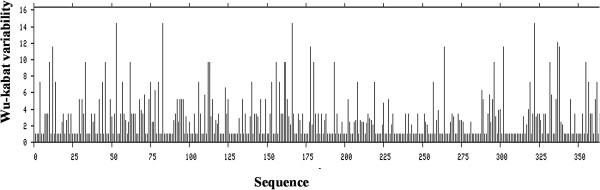
Protein variability index of the conserved peptide. The protein variability index of the conserved peptide was determined by using PVS server [24]. The conservancy threshold was 1.0 in this analysis. X axis indicates the amino acid position in sequences and Y axis indicates the Wu-kabat variability.
Both YPKCDRA and YYVKPG identified as consensus and highly immunogenic epitopes by two different algorithms
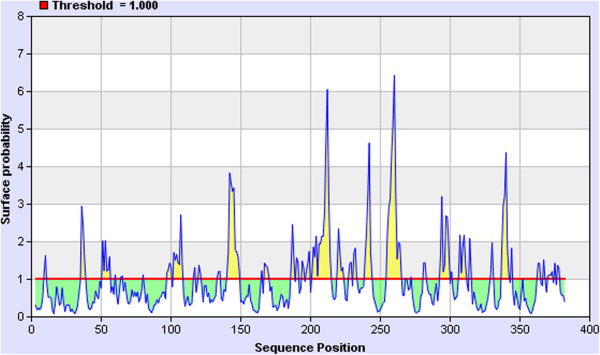
For vaccine design the peptide has to be immunogenic and antigenic [26]. The conserved peptide was found to be highly antigenic (Figure 3) in IEDB epitope prediction analysis [27]. In this analysis 1.000 threshold was used and most of the residues in the peptides were found above the threshold level. B-cell epitopes were predicted using Immune Epitope Database (IEDB) [28] B-cell epitope prediction tool and Bepipred [29] using the conserved protein sequence. Several epitopes were predicted (Table 1) by these algorithms, but only those epitopes sequences that are found full or at least 90% overlap between IEDB B-cell epitope prediction tool [28] and Bepipred prediction [29] are chosen as desired epitopes (Table 2). YPKCDRA and YYVKPG epitopes were found to be consensus among both tools predicted epitopes.
Figure 3.

Antigenicity of the conserved peptide. The conserved peptide was found to be highly antigenic in the IEDB analysis [27]. Most of the residues were found above the threshold 1.00. Residues in the yellow region are antigenic and in the green region are below the threshold (red line).
Table 1.
|
Bepipred analysis |
IEDB analysis |
||
|---|---|---|---|
| Peptide | Length (aa) | Peptide | Length (aa) |
| VRPGNFNQ |
8 |
DYKVQLFEKYFKY |
13 |
| GNAAIT |
6 |
HANCVNCTDDRCVLHCANFNVLFAM |
25 |
| DKSAGHPFN |
9 |
KTCFGPIVRKIFVDGVPFVVSCGYHYKELGLV |
32 |
| SYQEQ |
5 |
VSLHRHRLSLK |
11 |
| DVDNP |
5 |
ADPAMHI |
7 |
| YPKCDRA |
7 |
SSNAFLDLRTSCFSVAALTTG |
21 |
| YYVKPGGTSSGDATTAY |
17 |
YDFVVSKG |
8 |
| |
|
SVTLKHFFFA |
10 |
| |
|
DYNYYSYN |
8 |
| |
|
PTMCDIKQMLFCMEVVNKYFEI |
22 |
| |
|
DGGCLNASEVVVNN |
14 |
| |
|
KARVYYESM |
9 |
| |
|
LKYAIS |
6 |
| |
|
TVAGVSILS |
9 |
| |
|
GATCVIGTT |
9 |
| |
|
LKTLYKDV |
8 |
| |
|
YPKCDRA |
7 |
| |
|
CRIFASLILARKHGTCCTT |
19 |
| |
|
YRLANECAQVLSEYVLCGGGYYVKPG |
26 |
| |
|
ANSVFNILQA |
10 |
| TANVSAL | 7 | ||
Table 2.
| Consensus Epitope | Length (aa) |
|---|---|
| YPKCDRA |
7 |
| YYVKPG | 6 |
Nine surface accessible epitopes were predicted from the conserved peptide
To become a vaccine, an epitope should be accessible to the antibody. If the antibody can bind to the epitope that will be able to induce an immune response [26]. The surface accessibility of the conserved peptide (Figure 4) was determined using 1.000 threshold level and nine accessible epitopes were found to be above the threshold level (Figure 4) (Table 3) using Immune Epitope Database (IEDB) [28] Emini surface accessibility prediction analysis [30]. Among these nine epitopes, WDYPKC epitope overlaps with the Bepipred [29] and IEDB [28] predicted consensus epitope YPKCDRA.
Figure 4.
Conserved peptide’s surface accessibility. The surface accessible residues of the conserved peptide which are above the cut off are located in the yellow region. The red horizontal line indicates surface accessibility cutoff (1.000).
Table 3.
Predicted surface accessible antigenic sites by using Emini surface accessibility prediction analysis [30]
| Peptide | Length (aa) |
|---|---|
| HRHRLS |
6 |
| GNFNQDF |
7 |
| TDYNYYSYNL |
10 |
| ARVYYESMSYQEQD |
14 |
| AKNRAR |
6 |
| MTNRQYHQKML |
11 |
| TLYKDVD |
7 |
| WDYPKC |
6 |
| TRDRFY | 6 |
WDYPKCDRA is fully conserved among all human corona virus isolates
The conservancies of all epitopes were determined by IEDB conservancy analysis tools [31]. From the IEDB predicted epitopes, two epitopes (YPKCDRA, LKYAIS) and from Bepipred predicted epitopes, YPKCDRA epitope were found to be 100% conserved among all human coronavirus isolates (Table 4). WDYPKC epitope from the surface accessible epitope was also found to be 100% conserved (Table 4). Among the two consensus epitopes of Bepipred [29] and IEDB [28] analysis, YPKCDRA epitope was found to be 100% conserved among all human coronavirus isolates (Figure 5). This YPKCDRA and WDYPKC epitopes are in the same region and 100% conserved in all human coronaviruses. Therefore, the whole epitope WDYPKCDRA which is 100% conserved was then selected as the desired universal vaccine candidate.
Table 4.
Predicted conservancy of the antigenic site by using IEDB conservancy analysis [31]
| Sequence | Identity % |
|---|---|
|
Bepipred analysis | |
| VRPGNFNQ |
17.39 |
| GNAAIT |
41.30 |
| DKSAGHPFN |
17.39 |
| SYQEQ |
17.39 |
| DVDNP |
41.30 |
| YPKCDRA |
100.00 |
| YYVKPGGTSSGDATTAY |
17.39 |
|
IEDB analysis | |
| DYKVQLFEKYFKY |
17.39 |
| HANCVNCTDDRCVLHCANFNVLFAM |
17.39 |
| KTCFGPIVRKIFVDGVPFVVSCGYHYKELGLV |
17.39 |
| VSLHRHRLSLK |
17.39 |
| ADPAMHI |
17.39 |
| SSNAFLDLRTSCFSVAALTTG |
17.39 |
| YDFVVSKG |
17.39 |
| SVTLKHFFFA |
17.39 |
| DYNYYSYN |
17.39 |
| PTMCDIKQMLFCMEVVNKYFEI |
17.39 |
| DGGCLNASEVVVNN |
17.39 |
| KARVYYESM |
17.39 |
| LKYAIS |
100.00 |
| TVAGVSILS |
41.30 |
| GATCVIGTT |
17.39 |
| LKTLYKDV |
17.39 |
| YPKCDRA |
100.00 |
| CRIFASLILARKHGTCCTT |
17.39 |
| YRLANECAQVLSEYVLCGGGYYVKPG |
17.39 |
| ANSVFNILQA |
17.39 |
| TANVSAL |
17.39 |
|
Surface accessibility prediction analysis | |
| HRHRLS |
17.39 |
| GNFNQDF |
41.30 |
| TDYNYYSYNL |
17.39 |
| ARVYYESMSYQEQD |
17.39 |
| AKNRAR |
56.52 |
| MTNRQYHQKML |
17.39 |
| TLYKDVD |
17.39 |
| WDYPKC |
100.00 |
| TRDRFY | 17.39 |
Figure 5.

Conservancy of the predicted consensus epitopes. Three of the four epitopes were found to be 100% conserved. Here Y axis indicates the epitopes and X axis indicates the conservancy percentage.
WDYPKCDRA is also an accessible epitope
Hydrophilicity is desired feature of B cell epitope which indicates the accessibility of the epitope. The WDYPKCDRA epitope was found to be hydrophilic (Figure 6) in nature as determined by the IEDB Parker hydrophilicity analysis [32]. A threshold of 3.448 was used which is indicated by the red line and the residues of the epitope which are hydrophilic are in the yellow region. The maximum level was found as 4.5 (Figure 6) in the epitope.
Figure 6.
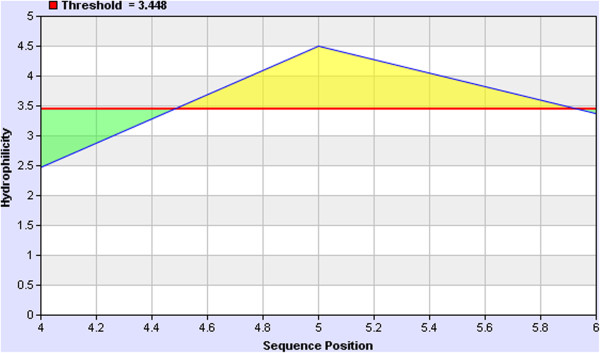
Hydrophilicity of the WDYPKCDRA epitope. Most of the residues of the desired WDYPKCDRA epitope were found to be hydrophilic in nature (in the yellow colored region). The residues which are below the cut off 3.448 (red line) are in the green region.
A tertiary structure of RNA directed RNA polymerase was predicted and validated using in silico approach
As the experimental tertiary structure of the RNA directed RNA polymerase is not available, we modeled a 3D structure by I-TASSER server [33] by multiple threading alignments. I-TASSER analysis deduced 5 different models (data not shown) for this protein. The quality of all the predicted protein models was checked by PROCHECK analysis [34]. From the PROCHECK analysis results, the protein model in which maximum numbers of amino acids residues were in maximum favorable region and G factor was highest was taken as the desired best model. The model in which 89.3% residues are found to be in the most favored region in Ramachandran plot (Additional file 6: Figure S6) and G-factor was -0.31 from the PROCHECK analysis was selected as the desired model. Along with the surface accessibility analysis and hydrophilicity analysis, the targeted WDYPKCDRA epitope was also found to be in the surface and accessible in the RNA directed RNA polymerase 3D structure (marked as green color) (Figure 7).
Figure 7.
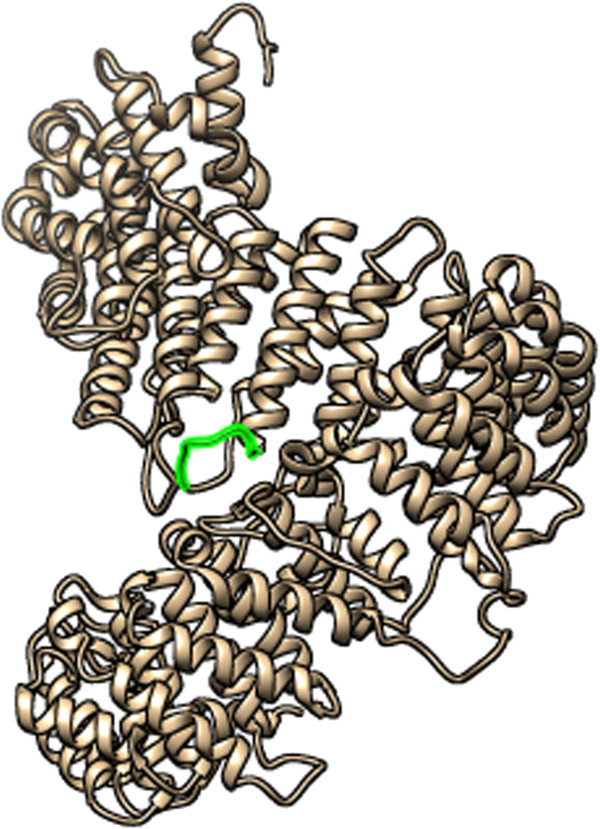
3D structure of RNA directed RNA polymerase protein. Predicted conserved WDYPKCDRA epitope mapped onto protein 3D structure using UCSF Chimera [35] visualization tool. Here green colored region indicates the conserved epitope WDYPKCDRA.
Discussion
Coronaviruses are one of the most diverse groups of virus which are becoming a deadly virus day by day. Though the first two strains were not so much deadly but the other members were pathogenic. After SARS outbreak, a new coronavirus strain called MERS-CoV is now going to cause another outbreak [4]. The cell tropism and cellular receptor of the six types of coronaviruses are not similar (Additional file 7: Table S1). Though at first it was thought that SARS-CoV and MERS-CoV are structurally similar and tried to treat MERS-CoV infected patient with the SASR-CoV treatment. But it was found that they bind to two different receptors, namely ACE2 and DPP4 or CD26 respectively [36]. These viruses are actually zoonotic origin, undergo recombination, and may be in future another strain of this group of virus will come [37]. Therefore, it is important to take preventing measures not only to prevent this new strain of coronavirus, also against all the strain of coronavirus. There is no recommended vaccine for coronaviruses which is necessary to prevent. Most of the cases, vaccines were designed by targeting spike protein. Similarly, researchers also reported to design vaccine against SARS-CoV and MERS-CoV spike protein [38,39]. Fernando et al. also designed lived-attenuated MERS coronavirus by mutating MERS-CoV envelope protein as a vaccine which will be for only MERS-CoV [40]. These vaccines would thus be effective for only those strains not for others. Giving a universal vaccine for all strain of viruses is much more promising solution rather than giving individual vaccine for individual strain.
The concept of prevention of viruses by designing a universal vaccine has also been reported previously, for example against Influenza virus. In case of influenza virus, universal vaccine against matrix 2 protein which was found to be conserved among all Influenza subtypes was reported [41]. An attempt to design universal vaccine against members of coronaviruses, like feline infectious peritonitis (FIPV), canine coronavirus (CCV), gastroenteritis coronavirus (TGEV), bovine coronavirus (BCV) targeting their spike protein in 1993 were observed [42]. But this concept was not applied to human coronaviruses.
Vaccine development has been one of the most important contributions of immunology to public health to date. Traditionally vaccines were based on the intact pathogen, either inactivated or live attenuated. These types of vaccines had some crucial drawbacks like safety consideration and the loss of efficacy due to the genetic variation of the pathogen. But now a day’s these vaccine concepts are greatly replaced by novel vaccine approaches like naked DNA vaccine, epitope based vaccine. The main benefit of immunization with an epitope-based vaccine is the ability to immunize with a minimal structure and it will stimulate an effective specific immune response, while avoiding potential undesirable effects [43].
In this study, we aimed to design an epitope-based universal vaccine for all human coronavirus strain. For this purpose we did multiple sequence alignment of the spike (S), envelope (E), membrane (M), nucleocapsid (N) protein and replicase polyprotein 1ab of all six human coronaviruses. Replicase polyprotein 1ab was taken to check whether there is any conservancy among the non structural protein as this replicase polyprotein cleaved into 15 non-structural proteins. In case of S, E, M and N protein, no putative conserved region was found. But conserved region was found in case of replicae polyprotein 1ab where the conserved region was in the RNA directed RNA polymerase. This indicates that this protein is less mutating than the S, E, M and N protein. This RNA directed RNA polymerase protein was targeted to determine antigenic sites based on immunogenicity and surface accessibility using different bioinformatics analyses. The consensus antigenic sites were the desired one and their conservancy was also determined. From the conservancy analysis it was found that Bepipred [29] and IEDB [28] analysis predicted consensus epitope YPKCDRA and the surface accessibility analysis [30] predicted epitope WDYPKC are 100% conserved. This 100% conserved WDYPKC and YPKCDRA are actually located in the same region of RNA directed RNA polymerase and it was then taken as the targeted epitope. This epitope was found to be accessible and hydrophilic which is one of the crucial requirements for an epitope to be used as a vaccine. This reflects a promising scope to use this conserved epitope as a universal vaccine both as preventive and therapeutic treatment. As this epitope remain long been conserved since 1960, it may be possible to use this vaccine in future for upcoming human coronavirus strains as well. To become an effective vaccine, it needs to be highly immunogenic, stable inside the body. If it is poorly immunogenic or unstable, it needs to be conjugated with adjuvant [44]. Though this epitope based vaccine is designed by in silico analyses, the actual immunogenicity, stability, efficacy and their delivery strategy inside the recipients body can’t be determined by this in silico analysis. To address these questions in vitro and in vivo experiments are essential.
Conclusions
This study shows that though the human coronaviruses are not structurally related but it is possible to design an epitope-based universal vaccine for all human coronavirus strains. Our results are based on sequence analysis and computational predictions show predicted epitope would be a candidate target for the universal vaccine; and to determine the actual effectiveness of the peptide for mounting an immune response both in vitro and in vivo studies can be performed.
Methods
Retrieving coronavirus structural and nonstructural protein sequences
A total available 46 replicase polyprotein 1ab, 17 spike (S) protein, 18 envelope (E) protein, 18 membrane (M) and 18 nucleocapsid (N) protein sequence data were retrieved from NCBI GenBank sequence database [45] (Additional file 7: Table S1).
Identification of conserved region
To find the conserved region, retrieved sequences were aligned using EBI-clustalW program [23]. This multiple sequence alignment (MSA) was done with Gonnet matrix [23]. Protein variability server (PVS) was used to calculate protein variability index using Wu-kabat Variability coefficient [24]. From the multiple sequence alignment where the highest number of identical and similar amino acid and no gap was found, the sequence was selected as a conserved region. That conserved region was then used for antigenic site prediction.
Detection of immunogenicity of conserved peptides
To evaluate the immunogenicity of the conserved peptides, various bioinformatics algorithms and computational tools were used. Bepipred (v1.0) [29] and B cell epitope prediction tools of The Immune Epitope Database (IEDB) [28] were used for this purpose. Bepipred predicts linear B-cell epitopes using hidden Markov model [29]. Default threshold 0.35 was used for Bepipred analysis. Among B cell epitope prediction tools of IEDB, prediction of linear epitopes from protein sequence tool was used. The Immune Epitope Database (IEDB) linear epitope prediction tools [28] made the option of using different prediction methods. Finally Kolaskar and Tongaonkar Antigenicity method [27] was applied in this study using a threshold of 1.000 because it predicts the antigenicity of the provided protein sequence. The epitopes which were found to be fully or at least 90% overlap between IEDB B-cell epitope prediction tool [28] and Bepipred prediction [29] are chosen as desired epitope sequences.
Prediction of surface accessible epitopes
To predict the surface accessible epitope of the conserved peptide, Emini surface accessibility prediction tool [30] of the B cell epitope prediction tools of The Immune Epitope Database (IEDB) [28] was used for this purpose using default threshold level 1.0.
Prediction of epitope conservancy
The epitope conservancy analysis tool from the IEDB analysis resource was employed for epitope conservancy prediction [31] of all predicted epitopes. The conservancy level of the epitopes were calculated by searching for identities in the given protein sequence.
Prediction of epitope hydrophilicity
The conserved epitope was then also analyzed to determine the hydrophilicity of the predicted epitopes. Parker hydrophilicity prediction tool [32] of Immune Epitope Database (IEDB) [28] was used for this purpose and default threshold 3.448 was used.
Prediction and evaluation protein 3D model
As the experimental structure of RNA directed RNA polymerase protein of any human coronavirus isolate was not found in protein data bank (PDB), a 3D structure was predicted using I-TASSER server [33]. I-TASSER server gives protein 3D structure by multiple threading alignments [33]. I-TASSER provided top models quality was then verified by PROCHECK software [34]. The model for which G factor was highest, and amino acid residues in favorable region was higher in PROCHECK analysis was selected as the best model. This model was then used to locate the epitope by using UCSF Chimera [35] visualization tool.
Abbreviations
HCoV: Human coronavirus; SARS: Severe acute respiratory syndrome; SARS-CoV: SARS coronavirus; MERS-CoV: Middle east respiratory syndrome coronavirus; HCoV-229E: Human coronavirus 229E; HCoV-OC43: Human coronavirus OC43; HCoV-NL63: Human coronavirus NL63; HCoV-HKU1: Human coronavirus HKU1; ACE2: AngiotensinI-converting enzyme 2; DPP4: Dipeptyl peptidae 4.
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
RS & AI performed the analysis. AI conceived the project idea. RS and AI wrote the manuscript. Both authors read and approved the final manuscript.
Supplementary Material
Multiple sequence alignment of Spike (S) protein: Multiple sequence alignment of total 17 numbers of sequences of human coronaviruse isolates indicate that there is no conservation in their spike protein. This alignment was visualized by Jalview 2.8 [25] and color scheme used is Clustalx. Conservation showed here is based on 11 base scales where yellow color bar and star sign indicates the full conservation. Alignment quality was based on BLOSUM 62 substitution matrix score where yellow color indicates good quality. All the colors changes according to the conservation and alignment quality. Black bars showed the consensus sequence.
{kind=link}
Multiple sequence alignment of envelope (E) protein: Figure legend as in supplementary Figure S1.
Multiple sequence alignment of membrane (M) protein: Figure legend as in supplementary Figure S1.
Multiple sequence alignment of nucleocapsid (N) protein: Figure legend as in supplementary Figure S1.
Conserved peptide found in RNA directed RNA polymerase by multiple sequence alignment of replicase polyprotein 1ab: All human coronaviruses are found to be conserved in their replicase polyprotein 1ab.
Ramachandran plot for RNA directed RNA polymerase protein: Red colored region is the most favored region, brown and yellow colored regions are additionally allowed region and generously allowed regions respectively.
Sequence sources and other sequence related information.
Contributor Information
Refat Sharmin, Email: refat519@gmail.com.
Abul Bashar Mir Md Khademul Islam, Email: khademul@du.ac.bd.
Acknowledgement
RS is a recipient of NST fellowship from Ministry of Science and Technology, Govt. of Bangladesh.
References
- Luis E, Isabel S, Sonia Z, Fernando A. Animal Viruses: Molecular Biology. UK: Caister Academic Press; 2008. Coronavirus Replication and Interaction with Host; pp. 149–202. [Google Scholar]
- De Groot RJ, Baker SC, Baric R, Enjuanes L, Gorbalenya AE, Holmes KV, Perlman S, Poon L, Rottier PJM, Talbot PJ, Woo PCY, Ziebuhr J. International Committee on Taxonomy of Viruses. (ninth report) Oxford: Elsevier; 2011. Family Coronaviridae; pp. 806–828. [Google Scholar]
- Lu R, Yu X, Wang W, Duan X, Zhang L, Roujian L, Xiaoyan Y, Wen W, Xijie D, Linglin Z, Weimin Z, Jin X, Lingjie X, Qin H, Jianxin L, Li R, Zhong W, Wenjie T. Characterization of human coronavirus etiology in Chinese adults with acute upper respiratory tract infection by real-timeRT-PCR assays. PLoS One. 2012;7(6):e38638. doi: 10.1371/journal.pone.0038638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Global alert and response (GAR) Middle East respiratory syndrome coronavirus (MERS-CoV) – update. World Health Organization; 2014. [ http://www.who.int/csr/don/2014_01_09/en/] [Google Scholar]
- Boheemen S, Sander B, Miranda G, Chris L, Theo MB, Stalin R, Ali MZ, Albert DME, Bart LH, Osterhaus AD, Alexander EG, Eric JS, Ron AMF. Genomic characterization of a newly discovered coronavirus associated with acute respiratory distress syndrome in humans. mBio. 2012;3(6):e00473–12. doi: 10.1128/mBio.00473-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wever BA, Van der Hoek L. Recently discovered human coronaviruses. Clin Lab Med. 2009;29(4):715–724. doi: 10.1016/j.cll.2009.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan-Yeung M, Xu RH. SARS: epidemiology. Respirology. 2003;8:S9–S14. doi: 10.1046/j.1440-1843.2003.00518.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hofmann H, Pyrc K, Van der Hoek L, Geier M, Ben B, Stefan P. Human coronavirus NL63 employs the severe acute respiratory syndrome coronavirus receptor for cellular entry. Proc Natl Acad Sci U S A. 2005;102(22):7988–7993. doi: 10.1073/pnas.0409465102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ali Mohamed Z, Van Boheemen S, Bestebroer TM, Albert DME, Osterhaus AD, Ron AM, Fouchier RA. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N Engl J Med. 2012;367:1814. doi: 10.1056/NEJMoa1211721. [DOI] [PubMed] [Google Scholar]
- Sørensen MD, Sørensen B, Gonzalez-Dosal R, Melchjorsen CJ, Weibel J, Wang J, Jun CW, Huanming Y, Kristensen P. Severe acute respiratory syndrome (SARS): development of diagnostics and antivirals. Ann N Y Acad Sci. 2006;1067:500–505. doi: 10.1196/annals.1354.072. PMID 16804033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng VC, Lau SK, Woo PC, Yuen KY. Severe acute respiratory syndrome coronavirus as an agent of emerging and reemerging infection. Clin Microbiol Rev. 2007;20:660e94. doi: 10.1128/CMR.00023-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pierre S. First coronavirus sufferer in France dies in hospital Reuters. 2013. [ http://uk.reuters.com/article/2013/05/28/uk-coronavirus-france-idUKBRE94R0Y320130528]
- Robert R. Coronavirus cases, deaths reported in Tunisia, Saudi Arabia. 2013. [ http://www.cidrap.umn.edu/news-perspective/2013/05/coronavirus-cases-deaths-reported-tunisia-saudi-arabia]
- Helen B. Spain reports its first MERS case; woman travelled to Saudi Arabia for Hajj. 2013. [ http://www.vancouversun.com/health/Spain+reports+first+MERS+case+woman+travelled+Saudi+Arabia+Hajj/9133754/story.html]
- Middle East respiratory syndrome coronavirus (MERS-CoV) 2014. [ http://www.cdc.gov/coronavirus/MERS/faq.html]
- Falzarano D, Darryl F, De Emmie W, Cynthia M, Julie C, Vincent J, Heinz F, Munster VJ. Inhibition of novel beta coronavirus replication by a combination of interferon-alpha2b and ribavirin. Sci Rep. 2013;3:1686. doi: 10.1038/srep01686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wong SS, Yuen KY. The management of coronavirus infections with particular reference to SARS. J Antimicrob Chemother. 2008;62:437–441. doi: 10.1093/jac/dkn243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coronavirus. [ http://www.cdc.gov/coronavirus/about/index.html]
- Xianfu W, Smith TG, Rupprecht CE. From brain passage to cell adaptation: the road of human rabies vaccine development. Exper Rev Vaccines. 2011;10(11):1597–1608. doi: 10.1586/erv.11.140. [DOI] [PubMed] [Google Scholar]
- Plotkin SA. Vaccines: Past, present and future. Nat Med. 2005;11(4):S5–S11. doi: 10.1038/nm1209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matthew C, Lam TT, Watson SJ, Annie LP, Velilava P, Paul G, Oliver GP, Andrew R, Yi G, Deenan P, Paul K, Eleni K. Full-genome deep sequencing and phylogenetic analysis of novel human betacoronavirus. Emerg Infect Dis. 2013;19:5. doi: 10.3201/eid1905.130057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Islam R, Sadman S, Aubhishek Z. A computational assay to design an epitope-based peptide vaccine against Chikungunya virus. Future Virol. 2012;7(10):1029–1042. doi: 10.2217/fvl.12.95. [DOI] [Google Scholar]
- Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994;22(22):4673–4680. doi: 10.1093/nar/22.22.4673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garcia-Boronat M, Diez-Rivero CM, Reinherz EL, Pedro AR. PVS: a web server for protein sequence variability analysis tuned to facilitate conserved epitope discovery. Nucleic Acids Res. 2008;36(Web Server issue):W35–W41. doi: 10.1093/nar/gkn211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Waterhouse AM, Procter JB, Martin DM, Clamp M, Barton GJ. Jalview Version 2-a multiple sequence alignment editor and analysis workbench. Bioinformatics. 2009;25:1189–1191. doi: 10.1093/bioinformatics/btp033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caoili SEC. B-cell epitope prediction for peptide-based vaccine design: towards a paradigm of biological outcomes for global health. Immunome Res. 2011;7:2. 2. [Google Scholar]
- Kolaskar A, Tongaonkar PC. A semiempirical method for prediction of antigenic determinants on protein antigens. FEBS Lett. 1990;276(1):172–174. doi: 10.1016/0014-5793(90)80535-q. [DOI] [PubMed] [Google Scholar]
- Vita R, Zarebski L, Greenbaum JA, Emami H, Hoof I, Salimi N, Damle R, Sette A, Peters B. The immune epitope database 2.0. Nucleic Acids Res. 2010;38(Database issue):D854–D862. doi: 10.1093/nar/gkp1004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jens Erik PL, Pontoppidan L, Ole L, Morten N. Improved method for predicting linear B-cell epitopes. Immunome Res. 2006;2:2. doi: 10.1186/1745-7580-2-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Emini EA, Hughes JV, Perlow DS, Boger J. Induction of hepatitis A virus-neutralizing antibody by a virus-specific synthetic peptide. J Virol. 1985;55(3):836–839. doi: 10.1128/jvi.55.3.836-839.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bui HH, Sidney J, Li W, Fusseder N, Sette A. Development of an epitope conservancy analysis tool to facilitate the design of epitope-based diagnostics and vaccines. BMC Bioinformatics. 2007;8(1):361. doi: 10.1186/1471-2105-8-361. PMID: 17897458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parker JM, Guo D, Hodges RS. New hydrophilicity scale derived from high performance liquid chromatography peptide retention data: correlation of predicted surface residues with antigenicity and X-ray-derived accessible sites. Biochemistry. 1986;23:25. doi: 10.1021/bi00367a013. [DOI] [PubMed] [Google Scholar]
- Zhang Y. I-TASSER server for protein 3D structure prediction. BMC Bioinformatics. 2008;9:40. doi: 10.1186/1471-2105-9-40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laskowski RA, MacArthur MW, Moss DS, Thornton JM. PROCHECK: a program to check the stereochemical qualit of protein structures. J Appl Cryst. 2001;26:283–291. [Google Scholar]
- Pettersen EF, Goddard TD, Huang CC. UCSF Chimera- a visualization system for exploratory research and analysis. J Comput Chem. 2004;25:1605–1612. doi: 10.1002/jcc.20084. [DOI] [PubMed] [Google Scholar]
- To KKW, Hung IFN, Chan JFW, Yuen KY. From SARS coronavirus to novel animal and human coronaviruses. J Thorac Dis. 2013;5((Suppl 2)):S103–S108. doi: 10.3978/j.issn.2072-1439.2013.06.02. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muller MA, Muller MA, Stalin RV, Doreen M, Benjamin M, Stephan K, Saskia L, Robert W, Sabine S, Tasnim S, Katrin Z, Tabea B, Isabella E, Marco T, Ali MZ, Albert DME, Ron AM, Bart LH, Christian D. Human Coronavirus EMC Does Not Require the SARS-Coronavirus Receptor and Maintains Broad Replicative Capability in Mammalian Cell Lines. mBio. 2012;3:e00515–12. doi: 10.1128/mBio.00515-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang ZY, Kong WP, Huang Y, Roberts A, Murphy BR, Subbarao K, Nabel GJ. A DNA vaccine induces SARS coronavirus neutralization and protective immunity in mice. Nature. 2004;428(6982):561–564. doi: 10.1038/nature02463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sudhakar A, Robin G, Boyd L, Agnihothram S, Gopal R, Yount BL Jr, Donaldson EF, Menachery VD, Graham RL, Scobey TD, Gralinski LE, Denison MR, Zambon M, Baric R. Platform strategies for rapid response against emerging coronaviruses: MERS-CoV serologic and antigenic relationships in vaccine design. J Infect Dis. 2013;10:1093. [Google Scholar]
- Fernando A, Marta L, Isabel S, Sonia Z, Jose L, Silvia LJ, German A, Luis E. Engineering a replication-competent, propagation defective middle east respiratory syndrome coronavirus as a vaccine candidate. mBio. 2013;4(5):e00650–13. doi: 10.1128/mBio.00650-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pica N, Palese P. Toward a universal influenza virus vaccine: prospects and challenges. Annu Rev Med. 2013;64:189–202. doi: 10.1146/annurev-med-120611-145115. [DOI] [PubMed] [Google Scholar]
- Elaine VJ, Sharon K, Timothy JM, Albert PR, Smithkline BC. Universal coronavirus vaccine. WIPO Patentscope. 1993. WO1993023421 A1. [ http://www.google.com/patents/WO1993023421A1?cl=en]
- Arnon R. A novel approach to vaccine design–epitope-based vaccines. FEBS J. 2006;273:33–34. [Google Scholar]
- Olesen OF, Lonnoroth A, Mulligan B. Human vaccine research in the European Union. Vaccine. 2009;27(5):640–645. doi: 10.1016/j.vaccine.2008.11.064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benson DA, Cavanaugh M, Clark K, Karsch-Mizrachi I, Lipman DJ, Ostell J, Sayers EW. GenBank. Nucleic Acids Res. 2008;41(Database issue):D36–D42. doi: 10.1093/nar/gks1195. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Multiple sequence alignment of Spike (S) protein: Multiple sequence alignment of total 17 numbers of sequences of human coronaviruse isolates indicate that there is no conservation in their spike protein. This alignment was visualized by Jalview 2.8 [25] and color scheme used is Clustalx. Conservation showed here is based on 11 base scales where yellow color bar and star sign indicates the full conservation. Alignment quality was based on BLOSUM 62 substitution matrix score where yellow color indicates good quality. All the colors changes according to the conservation and alignment quality. Black bars showed the consensus sequence.
Multiple sequence alignment of envelope (E) protein: Figure legend as in supplementary Figure S1.
Multiple sequence alignment of membrane (M) protein: Figure legend as in supplementary Figure S1.
Multiple sequence alignment of nucleocapsid (N) protein: Figure legend as in supplementary Figure S1.
Conserved peptide found in RNA directed RNA polymerase by multiple sequence alignment of replicase polyprotein 1ab: All human coronaviruses are found to be conserved in their replicase polyprotein 1ab.
Ramachandran plot for RNA directed RNA polymerase protein: Red colored region is the most favored region, brown and yellow colored regions are additionally allowed region and generously allowed regions respectively.
Sequence sources and other sequence related information.