

SUMMARY
We show that OsSERK2 is a regulator of innate immune signaling mediated by multiple non-RD receptor kinases (RKs) including XA21, XA3, and OsFLS2. OsSerk2-silenced rice lines are impaired in XA21-mediated immunity to Xoo PXO99, XA3-mediated immunity to Xoo PXO86, and OsFLS2-mediated defense responses. Thus, OsSERK2 is broadly involved in PRR-mediated immunity in rice.
Key words: immune receptor kinases, somatic embryogenesis receptor kinase (SERK), immunity, Xanthomonas oryzae pv. oryzae, rice.
Abstract
The rice XA21 immune receptor kinase and the structurally related XA3 receptor confer immunity to Xanthomonas oryzae pv. oryzae (Xoo), the causal agent of bacterial leaf blight. Here we report the isolation of OsSERK2 (rice somatic embryogenesis receptor kinase 2) and demonstrate that OsSERK2 positively regulates immunity mediated by XA21 and XA3 as well as the rice immune receptor FLS2 (OsFLS2). Rice plants silenced for OsSerk2 display altered morphology and reduced sensitivity to the hormone brassinolide. OsSERK2 interacts with the intracellular domains of each immune receptor in the yeast two-hybrid system in a kinase activity-dependent manner. OsSERK2 undergoes bidirectional transphosphorylation with XA21 in vitro and forms a constitutive complex with XA21 in vivo. These results demonstrate an essential role for OsSERK2 in the function of three rice immune receptors and suggest that direct interaction with the rice immune receptors is critical for their function. Taken together, our findings suggest that the mechanism of OsSERK2-meditated regulation of rice XA21, XA3, and FLS2 differs from that of AtSERK3/BAK1-mediated regulation of Arabidopsis FLS2 and EFR.
INTRODUCTION
The XA21 receptor kinase confers broad-spectrum resistance to Xanthomonas oryzae pv. oryzae (Xoo) (Song et al., 1995). Animals and other plant species also carry membrane-anchored receptors with striking structural similarities to XA21 (Ronald and Beutler, 2010). Many of these receptors play key roles in recognition of conserved microbial signatures (also called pathogen-associated molecular patterns (PAMPs)) and host defense (Song et al., 1995; Lemaitre et al., 1996; Medzhitov et al., 1997; Poltorak et al., 1998; Gomez-Gomez and Boller, 2000; Zipfel et al., 2006; Ronald and Beutler, 2010). XA21 and structurally similar immune receptors activate defense signaling via membrane-associated complexes that include non-RD (arginine-aspartic acid) kinases to induce a core set of defense responses (Ronald and Beutler, 2010; Dardick et al., 2012). The non-RD kinases are either associated with the receptor via adaptor proteins (animals) or integral to the receptor (plants) (Ronald and Beutler, 2010; Schwessinger and Ronald, 2012). In rice, the immune receptors XA21, XA3, Pid2, and FLS2 all belong to the non-RD subclass of kinases (Dardick et al., 2012; Schwessinger and Ronald, 2012).
In contrast to non-RD kinases, which are associated with the immune response, most RD kinases appear to regulate non-immune responses or serve as co-regulators of receptor kinase-mediated immunity (Chinchilla et al., 2006; Heese et al., 2007; Roux et al., 2011; Schwessinger et al., 2011; Schwessinger and Ronald, 2012) with the notable exception of the RD-kinase CERK1 in Arabidopsis, which directly binds chitin (Liu et al., 2012b).
In Arabidopsis, members of the somatic embryogenesis receptor kinase (SERK) regulate the function of multiple plasma-membrane-localized receptor kinases (RKs) including hormone receptors and immune RKs (Chinchilla et al., 2009; Li, 2010), and are members of the RD subclass of kinases (Schwessinger and Ronald, 2012). The best-studied member SERK3 is also referred to as BAK1 (brassinosteroid-insensitive 1 (BRI1) associated kinase 1), as it was initially identified as a key regulator of BRI1-mediated signaling (Li et al., 2002; Nam and Li, 2002). BRI1 is the main receptor of brassinosteroids (BR), an important class of plant hormones regulating growth and development (Li and Chory, 1997; Clouse, 2011). SERK3 and its closest paralog SERK4 are critical co-regulators of the immune response triggered by the ligand-activated Arabidopsis immune receptor kinase FLS2 (flagellin insensitive 2), EFR (EF-TU receptor), and PEPR1/2 (PEP receptors 1 and 2) (Chinchilla et al., 2006; Heese et al., 2007; Postel et al., 2009; Krol et al., 2010; Roux et al., 2011; Schwessinger et al., 2011). The pattern-recognition receptors (PRRs) FLS2 and EFR recognize the bacterial proteins (or derived epitopes) flagellin (flg22) or EF-TU (elf18), respectively (Zipfel et al., 2006; Chinchilla et al., 2007). In contrast, PEPR1/2 are paralogous receptors for the endogenously produced small danger-associated peptides, AtPeps (Yamaguchi et al., 2006, 2010). Arabidopsis FLS2 and EFR do not constitutively interact with SERK3 (or any other SERK-family member) (Chinchilla et al., 2007; Heese et al., 2007; Schulze et al., 2010; Roux et al., 2011; Schwessinger et al., 2011). Only upon ligand binding, FLS2 and EFR undergo a nearly instantaneous complex formation with SERK3 and potentially with additional co-regulatory RKs (Chinchilla et al., 2009; Schulze et al., 2010; Roux et al., 2011). The FLS2/EFR–SERK3 complex formation is independent of the kinase activity of either interaction partner or any other associated kinase (Schulze et al., 2010; Schwessinger et al., 2011). Indeed, the co-crystal structure of the FLS2–SERK3 ectodomains and flg22 suggests that flg22 acts as molecular glue by stabilizing the interaction between both receptors (Sun et al., 2013). This ligand-induced heterodimer formation is the molecular switch-on for transmembrane signaling of these Arabidopsis RKs (Albert et al., 2013). The tight association of the intracellular kinase domains is hypothesized to induce downstream signaling activation via specific structurally guided auto- and transphosphorylation events. SERK3 and FLS2/EFR undergo unidirectional phosphorylation in vitro—that is, SERK3 is able to transphosphorylate FLS2 or EFR but not vice versa (Schwessinger et al., 2011).
In rice, XA21 confers robust resistance to Xoo (Song et al., 1995). XA21 biogenesis occurs in the endoplasmic reticulum (ER) (Park et al., 2010, 2013). After processing and transit to the plasma membrane, XA21 binds to XB24 (XA21 binding protein 24) (Chen et al., 2010b). XB24 physically associates with the XA21 juxtamembrane (JM) domain and catalyzes the autophosphorylation of serine and threonine residue(s) on XA21, keeping XA21 in an inactive state (Chen et al., 2010b). Upon pathogen recognition, XA21 kinase disassociates from XB24 and is activated (Chen et al., 2010b). This activation triggers a series of downstream events resulting in a robust resistance response. XA21-mediated signaling is attenuated by the XB15 protein phosphatase 2C, which dephosphorylates XA21 (Park et al., 2008). Despite these advances, the early events governing XA21 activation have not yet been fully elucidated.
Based on the structural similarity of the XA3 immune receptor, which also confers immunity to Xoo (Sun et al., 2004; Xiang et al., 2006), and OsFLS2, which recognizes bacterial flagellin (Takai et al., 2008), with XA21, we hypothesized that XA3 and OsFLS2 transduce their responses through the same components that transduce the XA21-mediated response.
We have also identified an XA21 paralog lacking the transmembrane (TM) and kinase domains (called XA21D) (Wang et al., 1998). Based on the partial resistance phenotype conferred by XA21D and its predicted exclusively extracellular location, we hypothesized that XA21 and XA21D would partner with a co-regulatory receptor kinase (Wang et al., 1998). Based on recent findings (Chinchilla et al., 2007; Heese et al., 2007; Roux et al., 2011), we hypothesize that this hypothetical co-regulatory receptor kinase might be orthologous to Arabidopsis SERK proteins.
We therefore investigated the function of rice SERK-family members in XA21-, XA3-, and OsFLS2-mediated immunity. We identify the RD receptor kinase OsSERK2 (Os04g38480) and demonstrate its requirement for both XA21- and XA3-mediated immunity as well as rice FLS2 signaling. We also show that OsSERK2 is involved in BR-regulated plant growth. The kinase domain of OsSERK2 directly interacts with XA3, XA21, OsFLS2, and OsBRI1 in yeast two-hybrid assays in an enzymatic activity-dependent manner. Consistently with these results, OsSERK2 and XA21 form constitutive heterodimeric complexes in planta. OsSERK2 and XA21 undergo bidirectional transphosphorylation in vitro, which is influenced by the domain architecture of both. These results demonstrate an essential role for OsSERK2 in regulating development and receptor kinase-mediated immunity and suggest that direct interaction of OsSERK2 with rice immune receptors is critical for function.
RESULTS
Phylogenetic Analysis of Rice SERK-Family Members
Previous studies in Arabidopsis demonstrated that SERK-family members, and in particular SERK3 (also known as BAK1), are essential for both BR signaling mediated by BRI1 (Li et al., 2002; Nam and Li, 2002) and immunity mediated by FLS2, EFR, and PEPR1/2 (Chinchilla et al., 2006; Heese et al., 2007; Krol et al., 2010; Roux et al., 2011; Schwessinger et al., 2011). In the case of rice XA21 and XA21D (an XA21 paralog lacking transmembrane and kinase domain), a co-regulatory receptor kinase has been hypothesized but its identification has remained elusive (Wang et al., 1998). Because XA21 is structurally similar to FLS2 and EFR and belongs to the same subfamily XII of LRR-RKs (Chen and Ronald, 2011), we hypothesized that one or more rice SERK-family members serve as co-regulatory receptor kinase for rice immune receptors.
To identify such a co-regulator, we carried out phylogenetic analysis on the two rice SERK proteins, OsSERK1 (Os08g07760) and OsSERK2 (Os04g38480) (Supplemental Figure 1) (Singla et al., 2009), and five Arabidopsis SERK proteins (Li, 2010). The two rice SERKs are most closely related to Arabidopsis SERK1 and SERK2 (Figure 1A and Supplemental Figure 1). In contrast to Arabidopsis SERK3 (BAK1) and SERK4, which are the main SERK-family members involved in Arabidopsis immune signaling (Li, 2010; Roux et al., 2011; Schwessinger et al., 2011), SERK1 and SERK2 are known for their role in developmental processes (Li, 2010). Recently, it was shown that Arabidopsis SERK1 is also involved in immune signaling in transgenic plants expressing the tomato immune receptor Ve1 (Fradin et al., 2011). Non-specific silencing of the two rice SERK proteins and several closely related proteins in rice compromises resistance against the fungal pathogen Magnaporthe oryzae (Park et al., 2011). Conversely, overexpression of OsSERK2 (referred to as OsSERK1 in the original publication (Hu et al., 2005)) enhances resistance against Magnaporthe oryzae (Hu et al., 2005). These results suggest that one or both rice SERK proteins are involved in the rice immune response.
Figure 1.
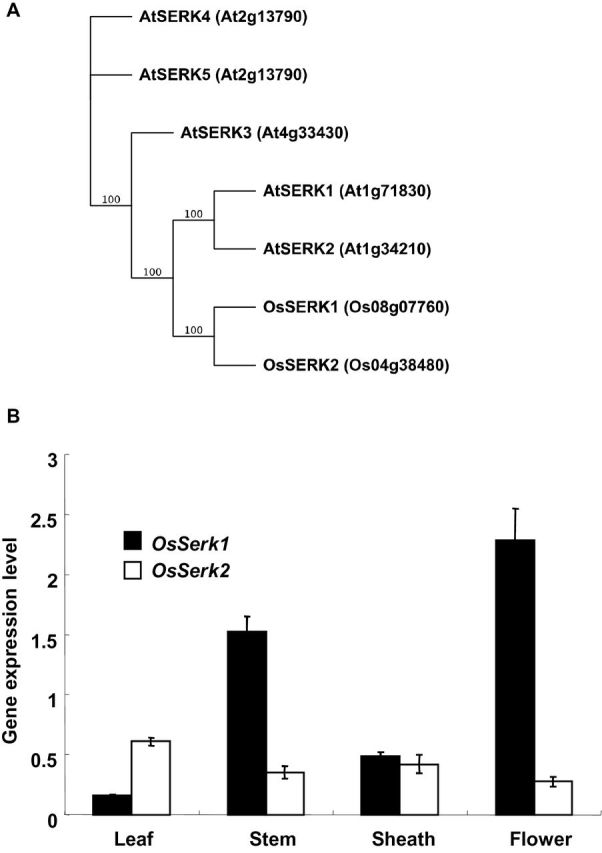
OsSERK2 Is the Only Rice SERK-Family Member Highly Expressed in Leaf Tissue.
(A) Phylogenetic analysis of the two rice and five Arabidopsis SERK proteins. Rice SERK1 and SERK2 were grouped with their five Arabidopsis homologous SERK proteins. Full-length amino acid sequences of all SERK proteins were analyzed using Geneious Tree builder. The phylogenetic tree was generated using a bootstrap neighbor-joining tree applying 1000 replicates. Protein identifiers are given in brackets.
(B) OsSERK2 is the most highly expressed SERK-family member in mature rice leaves. Quantitative real-time PCR was performed on cDNA synthesized from RNA samples extracted from rice cultivar Nipponbare tissue as indicated. Gene expression levels of OsSerk1 and OsSerk2 were normalized to the expression of the actin reference gene. Data shown represent average expression level of one out three biological experiments with error bars indicating SD of three technical replicates. The experiment was repeated three times, with similar results.
OsSerk2 Is Preferentially Expressed in Leaves whereas OsSerk1 Is Expressed in Flowers
Because Xa21 confers resistance to Xoo in rice leaves (Song et al., 1995), we analyzed the expression patterns of OsSerk1 and OsSerk2 in leaves, stems, sheaths, and flowers by performing quantitative RT–PCR. OsSerk1 and OsSerk2 were expressed in all tissues tested. OsSerk2 is mainly expressed in leaves whereas OsSerk1 is mainly expressed in flowers and stems (Figure 1B). The expression level of OsSerk2 is much higher than that of OsSerk1 in rice leaves (Figure 1B). These results suggest that OsSerk2 rather than OsSerk1 regulates Xa21-mediated immunity.
Silencing of OsSerk2 Compromises Xa21-Mediated Immunity to Xoo
To test the function of OsSERK2 in rice XA21-mediated immunity, we carried out OsSerk2 silencing experiments in the Xa21 genetic background. For these experiments, we isolated a 383-bp OsSerk2 cDNA fragment, which is unique to the OsSerk2 gene, and introduced it into the pANDA vector, which carries a hygromycin selection marker, to generate the double-stranded RNA-based interference (dsRi) construct pANDA–OsSerk2Ri (Miki and Shimamoto, 2004). We then introduced the OsSerk2Ri construct into Xa21 and ProA-tagged Xa21 (ProAXa21) homozygous rice lines carrying the mannose selectable marker (Chen et al., 2010b). Three Xa21–OsSerk2Ri (abbreviated as XOsSerk2Ri) and one ProAXa21–OsSerk2Ri (abbreviated as ProAXOsSerk2Ri) double transgenic lines were assayed for silencing of OsSerk2 using quantitative RT–PCR. Among these four double transgenic lines, two XOsSerk2Ri lines (XOsSerk2Ri2 (B-2) and XOsSerk2Ri3 (B-3)) and the ProAXOsSerk2Ri line (ProAXa21/X-B-1) display specific reduction in the expression of OsSerk2 (Supplemental Figures 2 and 3). The expression levels of Xa21 and OsSerk1 in these lines are similar to the control Xa21 or ProAXa21 lines (Supplemental Figures 2 and 3).
We then analyzed the response of the T1 plants from the double transgenic lines, A814 derived from B-2, A815 derived from B-3, and A804 derived from ProAXa21/X-B-1, to infection by the Xoo strain, PXO99AZ. Whereas the Xa21 control line is highly resistant to Xoo, the double transgenic plants carrying Xa21 and silenced for OsSerk2 are susceptible, showing typical long water-soaked lesions (Figure 2). The susceptibility phenotype of the OsSerk2-silenced lines co-segregates with the presence of the OsSerk2Ri transgene. Segregants from double transgenic lines carrying Xa21 but lacking OsSerk2Ri are fully resistant (Supplemental Figures 4 and 5), demonstrating that silencing of OsSerk2 compromises Xa21-mediated immunity in rice.
Figure 2.
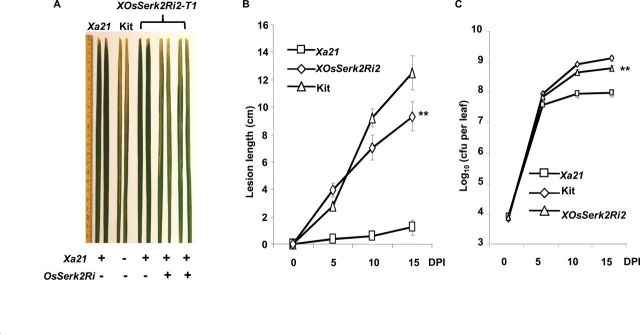
Silencing of OsSerk2 Compromises Xa21-Mediated Resistance to Xoo PXO99AZ.
Six-week-old plants of Xa21–OsSerk2Ri-2 (A814), Xa21 (resistant control), and Kitaake (Kit) (susceptible control) were inoculated with Xoo strain PXO99AZ.
(A) A814 plants in the presence of OsSerk2Ri develop long water-soaking lesions. Photograph depicts representative symptom development in leaves 14 d post inoculation. ‘+’ and ‘–‘ indicate absence or presence of the Xa21 and OsSerk2Ri transgene, respectively.
(B) XOsSerk2Ri2 plants (A814–178) homozygous for silenced OsSerk2 develop long water-soaked lesions. Lesion length was measured 0, 5, 10, and 15 d post inoculation. Graph shows average lesion length ± SD of at least 21 leaves from 7 independent plants. Statistical significance comparing A814–178 with Xa21 plants is indicated by asterisk (** P ≤ 0.05, ANOVA analysis, Tukey’s test).
(C) A814–178 is susceptible to Xoo PXO99AZ. Bacterial populations were counted 0, 5, 10, and 15 d post inoculation. Each data point represents the average ± SD of six leaves from two independent plants. Statistical significance comparing A814–178 with Xa21 plants is indicated by asterisk (** P ≤ 0.05, ANOVA analysis, Tukey’s test). These experiments were repeated at least three times, with similar results.
To further quantify the effect of OsSerk2 silencing, we generated Xa21 plants homozygous for the OsSerk2Ri (A814–178) transgene and performed more detailed infection studies. After 15 d of infection with Xoo PXO99AZ, the A814–178 plants display long lesions, similar to the Kitaake control plants (Figure 2A). At 15 d post inoculation, the average lesion length (9.30±1.04cm) of A814–178 plants is more than seven-fold greater than that of the Xa21 plants (1.23±0.55cm). The observed lesion length difference between A814–178 and Xa21 plants is highly significant with a p-value less than 0.0003. The average disease lesion length of A814–178 plants is closer to that of the susceptible parental control, Kitaake (12.5±1.26cm) (Figure 2B). Bacterial growth curve analysis revealed that the Xoo bacterial population in A814–178 plants (6.51×108 ± 1.07×108) is approximately nine-fold greater than in Xa21 lines (7.47×107 ± 1.67×107) and half of that observed for Kitaake (1.07×109 ± 2.01×108) at 15 d post inoculation (Figure 2C). These results are consistent with the leaf lesion phenotype described above. We also performed a similar experiment on an additional T2 homozygous double transgenic line (A804–55) developed from the ProAXOsSerk2Ri parent A804 and obtained similar results (Supplemental Figure 6). These results demonstrate that silencing of OsSerk2 compromises Xa21-mediated resistance. Using the same approach, we silenced OsSerk1 in the Xa21-Kitaake genetic background and analyzed the progeny for resistance. The silencing of OsSerk1 did not affect Xa21-mediated immunity (Supplemental Table 1). These results indicate that OsSerk2 but not OsSerk1 is a key player in Xa21-mediated immunity.
OsSerk2 Is Essential for Xa3-Mediated Immunity
Like XA21, the rice XA3 resistance protein belongs to subfamily XII of the LRR RKs, XA3 also functions as an immune receptor, conferring broad-spectrum resistance to most Xoo strains including PXO86 but not PXO99AZ (Sun et al., 2004; Xiang et al., 2006). Because of the structural and functional similarity of XA3 and XA21, we hypothesized that OsSerk2 may also be required for Xa3-mediated innate immunity. To test this hypothesis, we crossed Xa3 plants (IRBB3) with the homozygous Kitaake-OsSerk2Ri-4 (Kit-B-4) plants and obtained four F1 progeny called Xa3OsSerk2Ri F1 plants (Supplemental Figure 7). We inoculated these F1 plants with Xoo strain PXO86. As a control, we also inoculated the F1 progeny from a cross of Xa3 and Kitaake plants. We found that F1 progeny carrying both Xa3 and OsSerk2Ri displayed much longer lesions at 14 and 21 d post inoculation compared with F1 progeny of the control cross carrying Xa3 but lacking OsSerk2RiXa3 (ANOVA analysis: p-value less than 0.0001) (Figure 3A and 3B). To confirm the disease phenotype, we monitored bacterial growth over time (Figure 3C). Fourteen days after inoculation, bacterial populations of Xoo strain PXO86 accumulated to nearly 100-fold higher levels in Xa3OsSerk2Ri plants when compared to F1 plants from the Xa3-Kitaake control cross (Figure 3C). These results show that OsSERK2 is also critical for resistance mediated by the XA3 immune receptor.
Figure 3.
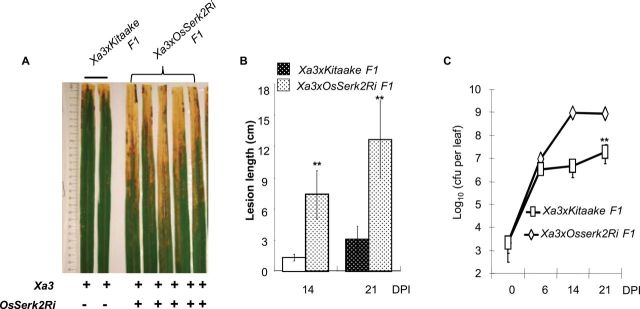
Silencing of OsSerk2 Compromises Xa3-Mediated Resistance to Xoo PXO86.
Eight-week-old plants of the F1 progeny of a cross between IRBB3 and Kitaake ((Xa3/Kitaake) F1, resistant control) and a cross between IRBB3 and Kitaake OsSerk2RNAi(X-B-4–2) homozygous for OsSerk2RNAi(Xa3/X-B-4–2) F1) were inoculated with Xoo strain PXO86.
(A) (Xa3/X-B-4–2) F1 plants develop long water-soaking lesions. Photograph depicts representative symptom development in leaves 21 d post inoculation. ‘+’ and ‘–‘ indicate absence or presence of the Xa3 gene and OsSerk2Ri transgene, respectively.
(B) (Xa3/X-B-4–2) F1 plants develop long water-soaked lesions. Lesion length was measured 14 and 21 d post inoculation. Graph shows average lesion length ± SD of at least 21 leaves from 7 independent plants. Statistical significance comparing (Xa3/X-B-4–2) F1 plants with (Xa3/Kitaake) F1 plants is indicated by asterisk (** P ≤ 0.05, ANOVA analysis, Tukey’s test).
(C) (Xa3/X-B-4–2) F1 plants are susceptible to Xoo PXO86. Bacterial populations were counted 0, 6, 14, and 21 d post inoculation. Each data point represents the average ± SD of six leaves from two independent plants from the same cross of Xa3/X-B-4–2. Statistical significance comparing (Xa3/X-B-4–2) F1 plants with (Xa3/Kitaake) F1 plants is indicated by asterisk (** P ≤ 0.05, ANOVA analysis, Tukey’s test). These experiments were repeated twice, with similar results.
OsSERK2 Is Involved in Rice FLS2-mediated Immune Signaling
SERK3 and SERK4 associate with the PRR FLS2 in vivo and are important for FLS2-mediated signaling in Arabidopsis (Chinchilla et al., 2006; Heese et al., 2007; Roux et al., 2011; Schwessinger et al., 2011). Because flagellin is also able to trigger OsFLS2-mediated signaling in rice (Takai et al., 2008), we tested whether OsSERK2 is involved in defense gene expression triggered by the application of flg22, a conserved peptide sequence derived from flagellin that is able to trigger FLS2-dependent defense signaling in many plant species including rice (Albert et al., 2010; Ding et al., 2012). We treated mature leaf strips of Kitaake or Kitaake plants silenced for OsSerk2 (Kit-OsSerk2Ri-4) (Supplemental Figure 7) with 1 μM flg22 and measured the gene expression changes of two independent marker genes by quantitative RT–PCR. The expression of both PR10b and Os4g10010 was dramatically reduced in plants silenced for OsSerk2 (Figure 4A and 4B). Plants silenced for OsSerk2 appeared to be fully sensitive to chitin application (Supplemental Figure 8) indicating that this reduction in defense gene expression was specific to flg22-triggered responses. These results suggest that chitin perception in rice is independent of OsSERK2 and is similar to SERK3-indpendent chitin perception in Arabidopsis (Chinchilla et al., 2009).
Figure 4.

OsSERK2 Regulates flg22-Triggered Defense Gene Expression in Rice and Directly Interacts with the Intracellular Domain of FLS2 in the Yeast Two-Hybrid System in a Kinase Catalytic Activity-Dependent Manner.
Leaf strips of 4-week-old Kitaake control or OsSerk2Ri (X-B-4–2) plants were treated with 1 μM flg22 peptide for 2 or 12h. Expression levels of the two defense marker genes of PR10b (A) and Os04g10010 (B) were measured by quantitative RT–PCR. Expression levels for each gene were normalized to actin reference gene expression. Data shown are normalized to the Kitaake mock-treated (2h) sample. Bars depict average expression level ± SD of two technical replicates. This experiment was repeated four times, with similar results.
(C) OsFLS2 and OsSERK2 intracellular domains interact in a kinase-dependent manner in the yeast two-hybrid system. Upper left panel: two representative colonies for each co-transformation. The blue color indicates nuclear interaction between the two co-expressed proteins. Numbers indicate the specific co-transformations. Upper right panel: legend for the specific co-transformation events encoded by numbers. Lower panel: Western blot with anti-LexA or anti-HA antibodies to confirm expression of LexA and B42 fusion proteins, respectively, for each co-transformation event. The Matchmaker LexA two-hybrid system (Clontech) was used for the yeast two-hybrid assay.
Using the yeast two-hybrid system, we found that the intracellular domain of OsSERK2 interacts with the intracellular domain of OsFLS2 (#1 in Figure 4C) suggesting, that OsSERK2 may directly regulate OsFLS2 function in rice. Mutations in residues required for full enzymatic activity in OsSERK2, OsFLS2, or both proteins compromised the interaction observed in the yeast two-hybrid systems (#2–#4 in Figure 4C). This result suggests that full enzymatic activity of both OsFLS2 and OsSERK2 is required for complex formation.
Rice Plants Silenced for OsSerk2 Display Morphological Features of BR-Insensitive Mutant Plants and Show Reduced Sensitivity to Brassinolide
In Arabidopsis, all four functional SERK-family members are involved in BRI1-mediated brassinosteroid signal transduction (Gou et al., 2012). We therefore hypothesized that OsSERK2 would regulate BR signaling in rice. Indeed, we found that OsSerk2Ri plants are semi-dwarf (Figure 5A), similar to the Osbri1 mutant plants (Nakamura et al., 2006). XOsSerk2Ri2 plants are reduced in size compared with the Xa21 control plants (Figure 5A). The leaf sheath, panicle, and internodes of each tiller of XOsSerk2Ri2 plants are shorter than those in Xa21 control plants (Figure 5B–5D). The lamina joint angle line is much reduced (2.8±0.5°) compared to that of Xa21 control plants (30±3.2°) (P = 1.12×10–29, Student’s two-tailed t-test) (Figure 5E and 5F). The culm length of XOsSerk2Ri2 is significantly shorter than that of Xa21 control plants (Figure 5G). The relative lengths of internodes III and IV in the XOsSerk2Ri2 plants are much reduced compared with those of Xa21 control plants (Figure 5H). OsSerk2Ri plants exhibit shorter coleoptiles and show reduced sensitivity to brassinolide hormone (Supplemental Figure 9). These results demonstrate that OsSERK2 is also involved in rice BR hormone signaling.
Figure 5.
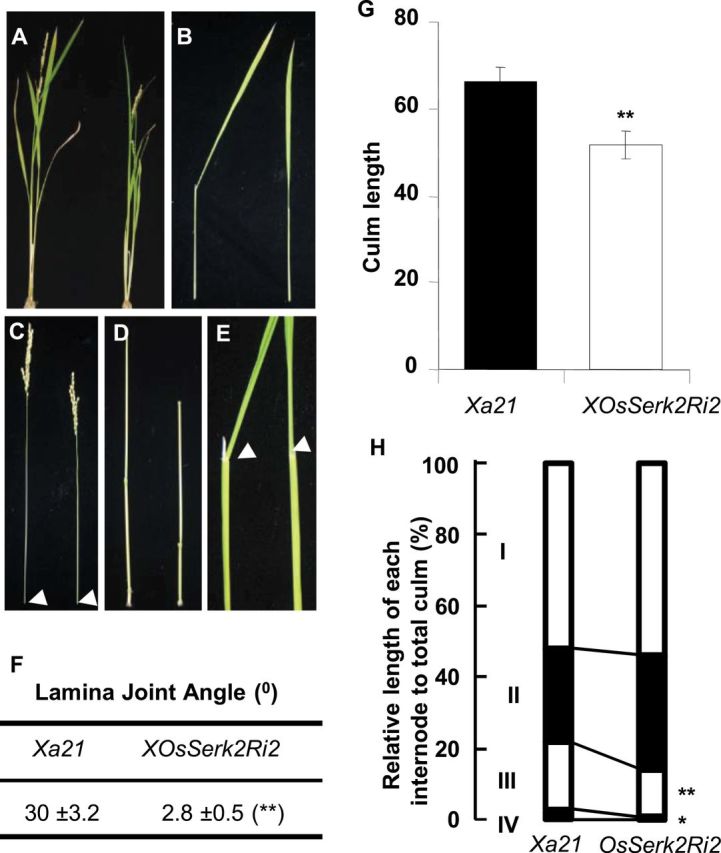
Plants Silenced for OsSerk2 Display Morphological Features Associated with Compromised Brassinosteroid Signaling.
(A) Gross morphology of Xa21 control plants (left) and A814–178 plants homozygous for silenced OsSerk2 (right).
(B) Leaf sheath morphology: the leaf sheath of A814–178 (right) is shorter than in Xa21 control plants (left).
(C) Panicle structure: A814–178 plants (right) have shorter panicle when compared to Xa21 control (left) plants. The arrow heads indicate nodes.
(D) Elongation pattern of internodes: the Xa21 plants (left) show an N-type elongation pattern, whereas A814–178 plants (right) show the typical dn-type pattern (Takeda, 1977).
(E) Leaf morphology: leaves of Xa21 control plants (left) are bent at the lamina joint indicated by the white arrowhead, whereas the leaves of A814–178 plants (right) are erect.
(F) Average degree of lamina joint angels of Xa21 control and A814–178 plants, respectively.
(G) Measurement of the culm length from Xa21 and A814–178 plants, respectively.
(H) Relative distance between internodes relative to total culm length in Xa21 control and A814–178 plants. In (F)–(H), the average ± SD of each parameter was determined from 12 plants of each genotype Xa21 control and A814–178 (* P ≤ 0.1, ** P ≤ 0.05, Student’s t-test).
OsSERK2 Interacts with XA21, XA3, and OsBRI1 in a Kinase-Dependent Manner in Yeast
Next, we investigated whether OsSERK2 directly regulates XA21-, XA3-, and OsBRI1-mediated signaling. The rationale for this experiment was that it had previously been shown that SERK3 interacts with BRI1, EFR, FLS2, and PEPR1/2 (Li et al., 2002; Nam and Li, 2002; Postel et al., 2009; Roux et al., 2011). We performed yeast two-hybrid assays using XA21K668, a truncated version of XA21 containing the whole intracellular domain and part of the TM domain, which was previously shown to interact with several key XA21 binding proteins (Park et al., 2008; Chen et al., 2010b), as bait for interaction with OsSERK2. We found that OsSERK2JMK carrying part of the TM, as well as the JM and kinase (K) domain, interacts with XA21K668 (Figure 6A). Similarly, OsSERK2JMK is also able to interact with XA3JMK in the yeast two-hybrid assay (Figure 6B). However, this interaction appears to be weaker than the interaction with XA21K668. OsSERK2JMK also interacts with OsBRI1JMK, the rice ortholog of Arabidopsis BRI1 (Figure 6C). In Arabidopsis, the interactions between SERK3 and the ligand-binding receptor FLS2, EFR, and BRI1 are independent of the catalytic activity of SERK3 when tested by in planta co-immunoprecipitation assays (Chinchilla et al., 2006; Wang et al., 2008; Schwessinger et al., 2011). To examine whether this is also the case for OsSERK2, we tested the interaction between the catalytic inactive OsSERK2JMKKE and XA21K668 or BRI1JMK in our yeast two-hybrid system. Both interactions were compromised by the catalytic inactivation of OsSERK2 (Figure 6A and 6C).
Figure 6.
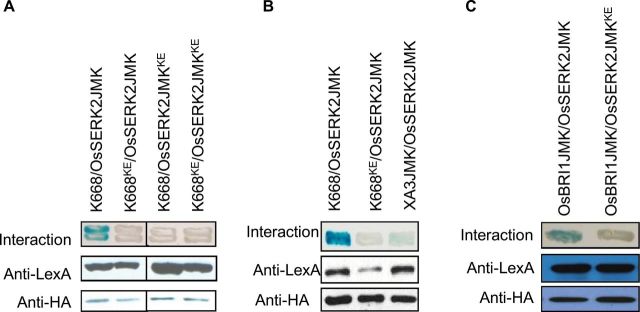
The OsSERK2 Intracellular Domain Interacts in a Kinase-Activity-Dependent Manner with the Intracellular Domain of the Three Predicted Ligand-Binding Receptors XA21, XA3, and OsBRI1 in Yeast Two-Hybrid System.
The blue color indicates nuclear interaction between the two co-expressed proteins.
(A) OsSERK2 and XA21 directly interact in a kinase catalytic activity-dependent manner. Part of the transmembrane (TM) and the whole intracellular domain ofOsSERK2 (OsSERK2JMK) and its kinase catalytically inactive mutant OsSERK2JMKK334E (OsSERK2JMKKE) were fused with the HA epitope in the vector pB42ADgc to obtain HA–OsSERK2JMK (abbreviated as OsSERK2JMK) and HA–OsSERK2JMKKE (abbreviated as OsSERK2JMKKE). HA–OsSERK2JMK and HA–OsSERK2JMKKE were co-transformed with LexA–XA21K668 (K668) or the catalytically inactive mutant LexA–K668K736E (abbreviated as K668KE), respectively.
(B) OsSERK2 directly interacts with XA3. HA–OsSERK2JMK was co-transformed XA3, containing part of the TM and the whole intracellular domain used with LexA (LexA–XA3JMK (abbreviated as XA3JMK)). HA–OsSERK2JMK was also co-transformed with LexA–K668 and LexA–K668KE for positive and negative interaction controls, respectively.
(C) OsSERK2 and OsBRI1 directly interact in a kinase catalytic activity-dependent manner. HA–OsSERK2JMK and HA–OsSERK2JMKKE were co-transformed with OsBRI1, respectively, containing part of the transmembrane and the whole intracellular domain, fused with LexA (LexA–OsBRI1JMK (abbreviated as OsBRI1JMK)). In (A)–(C), the expression of LexA-fused proteins, LexA–K668, LexA–K668KE, LexA–XA3JMK, and LexA–OsBRI1JMK, was confirmed by Western blotting using an anti-LexA antibody. The expression of proteins, HA–OsSERK2JMK and HA–OsSERK2JMKKE, was confirmed by Western blotting using an anti-HA antibody. Yeast two-hybrid experiments were performed using a Matchmaker LexA two-hybrid system (Clontech). This experiment was repeated three times, with similar results.
To assess whether phosphorylation is critical for the interaction between XA21 and OsSERK2, we generated a suite of catalytically inactive protein variants including XA21JK, XA21JKDN, XA21K668KE, XA21K668DN, OsSERK2JMKKE, OsSERK2JK, OsSERK2JKDN, and OsSERK2TJKDN. The catalytically compromised protein variants were generated by either mutating the conserved lysine (K) required for ATP binding and catalytic activity or the aspartate (D) required for phospho-transfer (Nolen et al., 2004). We tested the interaction between these different protein variants in the yeast two-hybrid system (Figure 6A and Supplemental Figure 10). All mutant XA21 protein variants were compromised in the interaction with OsSERK2JMK and OsSERK2JK (Figure 6A and Supplemental Figure 10). Similarly, no catalytically inactive protein variant of OsSERK2 was able to interact with XA21K688 (Supplemental Figure 9). Taken together, we conclude based on these results that the association of OsSERK2 with XA21, XA3, and OsBRI1 is dependent on the integrity of important catalytic residues and therefore most likely on the catalytic kinase activity of each protein in our yeast two-hybrid system.
OsSERK2 Forms a Constitutive Heterodimeric Complex with XA21 In Planta
Next, we aimed to confirm the interaction between OsSERK2 and XA21 in planta. It was recently reported that the addition of fusion peptides to the carboxy terminus of Arabidopsis SERK3 interferes with its function in innate immune signaling (Ntoukakis et al., 2011). For this reason, instead of tagging OsSERK2, we raised an antibody (anti-OsSERK2) against a unique peptide consisting of 10 amino acids (602–611) at the C-terminus of OsSERK2. Initially, we tested the specificity of anti-OsSERK2 using the Escherichia coli-produced GST–OsSERK2JMK protein. Because the protein encoded by OsSerk1 is the closest paralog to OsSERK2, we included the E. coli-produced GST–OsSERK1JMK protein in our experiment as a control. The anti-OsSERK2 antibody specifically recognizes OsSERK2 but not OsSERK1 (Figure 7A). The presence of two bands corresponding to GST–OsSERK2JMK suggests that GST–OsSERK2JMK is strongly phosphorylated during heterologous protein production in E. coli (Wang et al., 2005). Indeed, when we treated GST–OsSERK2JMK with the highly active lambda-phosphatase, the upper bands corresponding to (hyper-)phosphorylated GST–OsSERK2JMK disappeared and GST–OsSERK2JMK was detected as a discrete band of a single molecular size (Supplemental Figure 11). We also tested the specificity of the anti-OsSERK2 antibody on total protein extracted from Xa21 rice plants and Xa21 rice plants silenced for OsSerk2 (XOsSerk2Ri2). The anti-OsSERK2 antibody specifically recognized a protein band of the approximate size of 70kDa very close to the predicted molecular mass of OsSERK2. This band was only present in the total protein extract of Xa21 plants but not in the total protein extracts of plants silenced for OsSerk2 (XOsSerk2Ri2) (Figure 7B and Supplemental Figure 12). This observation suggests that the anti-OsSERK2 antibody specifically recognizes the OsSERK2 protein in total rice protein extracts.
Figure 7.

OsSERK2 and XA21 Form Constitutive Complexes In Planta.
(A) The newly developed anti-OsSERK2 antibody raised against a specific 10 amino acid epitope at the carboxy terminus specifically recognizes OsSERK2 but not OsSERK1. Top panel shows an anti-OsSERK2 Western blot on in Escherichia coli expressed and purified GST–OsSERK1JMK, GST–OsSERK2JMK, and GST–control proteins. The lower panel shows the Coomassie Brilliant Blue (CBB) staining of the corresponding region to assess equal quantities of protein were loaded.
(B) The anti-OsSERK2 antibody recognizes a specific protein of approximately 70kDa in size in total rice protein extracts from Xa21 plants but not from OsSerk2-silenced Xa21 plants (XOsSerkRi2) (upper panel). 75 μg of total protein for each genotype were separated by SDS–PAGE gel electrophoresis and subjected to immunoblot analysis with anti-OsSERK2 antibody (upper panel) or anti-Actin antibody (lower panel) as loading control. Full membranes for each immunoblot are shown in Supplemental Figure 12.
(C) OsSERK2 and XA21 form constitutive ligand-independent complexes in vivo. Immuno-complexes were precipitated from leaf material of Myc–Xa21-expressing rice plants using agarose-conjugated anti-Myc antibody. Kitaake rice leaves were used for the negative control. Components of the immuno-precipitated complexes were separated by SDS–PAGE gel followed by immuno-detection with anti-Myc (for Myc–XA21) and anti-OsSERK2 (for OsSERK2), separately. Myc–XA21 gives a band at about 130kDa. OsSERK2 (~70 kDa) was co-immunoprecipitated with XA21 in the absence of any treatment. The lower panel shows equal amounts of OsSERK2 in both total protein fractions before immunoprecipitation. This experiment was repeated four times, with similar results.
To further investigate the association between XA21 and OsSERK2 in vivo, we carried out co-immunoprecipitation experiments on protein extracts from mature leaves of 4–5-week-old greenhouse-grown plants at a stage when XA21 signaling is fully functional and Xoo infection assays are usually performed (Century et al., 1999). We previously generated transgenic plants carrying fully functional Myc–Xa21 under the control of its native promoter (Park et al., 2008). In co-immunoprecipitation experiments with anti-Myc-conjugated agarose beads, we detected a ~130-kDa polypeptide using the anti-Myc antibody only in transgenic plants (Figure 7C, top panel). This is consistent with our previous reports (Park et al., 2008; Chen et al., 2010b).
We next used the anti-OsSERK2 antibody to test whether OsSERK2 is present in the immunoprecipitated complexes. We successfully detected a band corresponding to the predicted size of OsSERK2 of ~70kDa in plants expressing Myc–XA21 but not in the wild-type Kitaake control (Figure 7C, middle panel). OsSERK2 was readily detected in the absence of any treatment indicating that XA21 and OsSERK2 can be found in constitutive complexes in planta (Figure 7C, middle panel).
OsSERK2 and XA21 Undergo Bidirectional Transphosphorylation Events In Vitro, Depending on Their Domain Architecture
As both OsSERK2 and XA21 are RKs, we tested whether transphosphorylation occurs between these two kinases. For this purpose, we expressed and purified a suite of GST– or His-NUS-tagged truncated protein variants of OsSERK2 and XA21, respectively (Figure 8A). We first investigated the phosphorylation capacities of XA21K668 and OsSERK2JMK, both containing part of the TM, full JM, and kinase domains (Figure 8A). Either kinase incubated on its own in the presence of [32P]-γ–ATP is able to undergo autophosphorylation (Figure 8B, 8C, and Supplemental Figure 13). This phosphorylation is abolished by mutations in the ATP-binding site, demonstrating that the observed effect was not due to a co-purified kinase (Figure 8B, 8C, and Supplemental Figure 13). Next we incubated each kinase with a catalytically inactive counterpart. Using protein variants in which both kinases contain part of the TM domain, we found that OsSERK2JMK was able to transphosphorylate XA21K668 but not the reverse (Figure 8B). The inability of XA21K668 to transphosphorylate OsSERK2 is independent of the domain structure of OsSERK2 or of the residue mutated to compromise catalytic activity (Figure 8B). XA21K668 is also unable to transphosphorylate OsSERK2JK variants that lack the TM domain and consist exclusively of the entire intracellular domain (Supplemental Figure 13).
Figure 8.

OsSERK2 and XA21 Undergo Bidirectional Transphosphorylation, Depending on Their Domain Architecture In Vitro.
(A) Depiction of protein domain architecture used for the transphosphorylation assays. OsSERK2JMK, XA21K668, and their respective kinase inactive variants OsSERK2JMKK334E (OsSERK2JMKKE) and XA21K668K736E (XA21K668KE) proteins contain partial sequences of their TM domain and full juxtamembrane (JM) and kinase domains. XA21JK, and its kinase inactive variant XA21JKD841N (XA21JKDN), contains the full JM and kinase domain but lacks the partial TM domain.
(B) OsSERK2JMK is able to transphosphorylate XA21K668 but not vice versa. The assay was performed by incubating GST–OsSERK2JMK (abbreviated as OsSERK2JMK) and GST–OsSERK2JMKK334E (abbreviated as OsSERK2JMKKE) in the presence or absence of His-Nus–XA21K668 (abbreviated as XA21K668), and His-Nus–XA21K668 and His-Nus–XA21K668K736E (abbreviated as XA21K668KE) in the presence or absence of GST–OsSERK2JMK using radioactive labeled [32P]-γ-ATP. Proteins were separated by SDS–PAGE and analyzed by autoradiography in the top panel and the protein loading control by CBB staining is shown in the lower panel, respectively.
(C) XA21JK is able to transphosphorylate OsSERK2JMK but not vice versa. The assay was performed by incubating GST–OsSERK2JMK (abbreviated as OsSERK2JMK) and GST–OsSERK2JMKK334E (abbreviated as OsSERK2JMKKE) in presence or absence of His-Nus–XA21JK (abbreviated as XA21JK), and His-Nus–XA21JK and His-Nus–XA21JKD841N (abbreviated as XA21JKDN) in the presence or absence of GST–OsSERK2JMK using radioactive labeled [32P]-γ-ATP. Proteins were separated by SDS–PAGE and analyzed by autoradiography in the top panel and the protein loading control is shown by Ponceau S in lower panel, respectively. This experiment was repeated twice, with similar results.
Next we tested the catalytic capacity of XA21JK, a XA21 protein variant exclusively consisting of the entire intracellular domain (Figure 8A). In the absence of any TM domain, XA21JK is able to transphosphorylate the catalytic inactive version of OsSERK2JMK (Figure 8C). In this set-up, OsSERK2JMK is unable to phosphorylate XA21JKD841N (Figure 8C).
These observations suggest that XA21 and OsSERK2 undergo bidirectional phosphorylation in vitro. In addition, the capacity of XA21 kinase to function as phospho-group-acceptor and donor is influenced by the presence of the TM domain. A similar observation has been recently made for the human epidermal growth factor receptor (EGFR) (Wang et al., 2011).
OsSERK2 Is an Active Kinase that Undergoes Autophosphorylation at Multiple Serine and Threonine Residues In Vitro
We previously demonstrated that XA21 exhibits relatively low autophosphorylation activity in vitro and is mostly autophosphorylated in the juxtamembrane region, which is important for its function (Chen et al., 2010a, 2010b). In contrast, OsSERK2 exhibits a much stronger autophosphorylation activity (Figure 8 and Supplemental Figure 13) and potentially undergoes multiple autophosphorylation events similar to its Arabidopsis ortholog (Karlova et al., 2006; Wang et al., 2008; Oh et al., 2010). To identify autophosphorylation sites of OsSERK2, we performed mass spectrometry on OsSERK2JK after incubation with cold ATP. We identified 12 unique phosphorylation events on serine and threonine residues (Table 1, Supplemental Figure 14, and Supplemental Data 1). The phosphorylation sites are evenly distributed over the entire intracellular domain of OsSERK2. Comparison with previously published phosphorylation sites of Arabidopsis SERK1 to SERK3 (Karlova et al., 2009; Wang et al., 2008; Oh et al., 2010) revealed that the in vitro phosphorylation pattern of OsSERK2 is most closely related to AtSERK1 rather than AtSERK3 (Table 1), further validating the phylogenetic analysis (Figure 1). Overall, the phosphorylation sites within the activation segments are conserved between all SERK proteins (residues T459, T463, T464, and T468 in OsSERK2). In contrast, residues predicted to be involved in protein–protein interactions and downstream signaling (all other residues of OsSERK2) appear to be specific to each individual SERK protein (Table 1).
Table 1.
Identification of In Vitro OsSERK2 Phosphorylation Sites by Q-ToF LC/MS/MS and Their Conservation in Arabidopsis SERK1, SERK2, and SERK3.
| Peptide sequencea | Measured [M+H]+ | Charge state | Actual minus calculated peptide mass (AMU) | Mascot Ion Scoreb | Identified sitec | Location within the protein | Orthologous phosphorylated site ind | ||
|---|---|---|---|---|---|---|---|---|---|
| Single phosphorylated peptides | AtSERK1 | AtSERK2 | AtSERK3 | ||||||
| 297-R.ELQVApTDNFSNK.N310 | 1444.62 | 2 | –0.002 | 42.6 | T303 | Juxtamembrane region | S299 | S302 | n/a |
| 324-R.LADGpSLVAVK.R-335 | 1051.55 | 2 | 0.015 | 48.5 | S329 | Nterminal lobe | T325 | n/a | T312 |
| 376-R. LLVYPYMANGpSVASR.L-392 | 1719.81 | 2 | 0.0045 | 54.8 | S387 | Predicted substrate binding pocket | S383 | n/a | n/a |
| 461-K.DTHVpTTAVR.G-471 | 1078.48 | 2 | 0.0014 | 41.3 | T466 (or T467) | Activation segment | T462 | T465 | T449 |
| 461-K.DTHVTpTAVR.G-471 | 1078.48 | 2 | 0.0014 | 43.9 | T467 (or T466) | Activation segment | T463 | T466 | T450 |
| 470-R.GpTIGHIAPEYLSTGK.S-486 | 1622.78 | 2 | 0.0048 | 68.6 | T472 | Activation segment | T468 | n/a | T455 |
| 607-R.HNDWIVDpSTYNLR.A-621 | 1711.74 | 2 | 0.0033 | 87.2 | S615 | Cterminal tail | S612 | n/a | n/a |
| 620-R.AMELpSGPR.-628 | 939.39 | 2 | 0.0005 | 43.4 | S625 | Cterminal tail | S622 | S625 | S612 |
| Double phosphorylated peptides | AtSERK1 | AtSERK2 | AtSERK3 | ||||||
| 340-R. TPGGELQFQpTEVEMIpSMAVHR. N362 | 2519.07 | 3 | 0.0031 | 66.7 | T350, S356 | Nterminal lobe | T346, S352 | n/a | n/a |
| 456-K. LMDYKDpTHVpTTAVR.G-471 | 1808.77 | 2 | 0.018 | 46.4 | T463, T466 | Activation segment | T459, T462 | T462, T465 | T446, T449 |
| 607-R.HNDWIVDpSpTYNLR.A-621 | 1791.71 | 2 | 0.0062 | 62.8 | S615, T616 | Cterminal tail | S612, T613 | –, T616 | n/a |
For each identified phosphorylation site, the highest scoring peptide, its specific parameters, and its conservation in three Arabidopsis SERK proteins are given. The specific MS2 spectra can be found in Supplemental Figure 12 and in Supplemental Data 1.
aThe numbers appearing before and after the amino acid sequence correspond to the preceding and following amino acids, respectively. pS and pT indicate phosphoseryl and phosphothreonyl residues, respectively. bBest Mascot Ion score for the indicated peptide. All peptides were inspected manually. cAt least two peptides were obtained and manually inspected for each identified phosphorylation. dAccording to Karlova et al. (2009), Oh et al. (2010), and Wang et al. (2008).
DISCUSSION
It was previously reported that OsSERK2 is involved in BR signaling and in resistance against the fungal pathogen M. oryzae (Hu et al., 2005; Park et al., 2011). Silencing of OsSerk2 in combination with OsSerk1 and other related genes leads to stunted growth, reduced sensitivity to exogenous BR application, and compromised resistance to M. oryzae (Park et al., 2011). In these studies, it was consistently shown that overexpression of OsSerk2 enhances resistance to M. oryzae (Hu et al., 2005). A detailed molecular mechanism of how the altered expression of OsSerk2 leads to these phenotypes has not been previously provided. Here we show that OsSERK2 is required for immune signaling pathways controlled by three immune RKs: XA21, XA3, and OsFLS2 (Figures 2–4 and Supplemental Figures 4–6), but is not required for CeBIP-mediated chitin signaling (Supplemental Figure 8). In addition, we demonstrate that OsSerk2 is required for BR signaling in rice (Figure 5 and Supplemental Figure 9). Because OsSERK2 interacts with the intracellular domain of these immune receptors in the yeast two-hybrid system, OsSERK2 most likely exerts its regulatory function by directly interacting with and phosphorylating these receptors (Figures 4 and 5, and Supplemental Figure 10). In yeast, the interaction between XA21 and OsSERK2 requires the catalytic activity of both kinases (Figure 5A and Supplemental Figure 10).
These observations suggest that the catalytic activity of each interaction partner is required for formation of stable constitutive heterodimeric complexes between XA21 and OsSERK2 in yeast. However, if the catalytic activity of either kinase is compromised, these proteins might still be able to transiently interact . This hypothesis is supported by the observed transphosphorylation between active and catalytic impaired kinases in vitro (Figure 8 and Supplemental Figure 10). A transient interaction between XA21 and OsSERK2 might explain how a catalytically impaired variant of XA21 is able to confer a partial resistance phenotype (Andaya and Ronald, 2003). Several of the newly identified autophosphorylation sites of OsSERK2 might be important for a stable interaction with XA21 and downstream signaling (Table 1). To fully test these hypotheses, it will be critical to analyze the contribution of individual phosphorylation sites of OsSERK2 on its role in BR and immune receptor kinase signaling in vivo.
Multiple Functional Roles of Rice OsSERK2
Our domain structure and phylogenetic analysis indicate that the rice genome encodes only two SERK proteins: OsSERK1 and OsSERK2 (Figure 1A and Supplemental Figures 1 and 15A and 15B). Although previous reports hypothesized the presence of several additional SERK-like proteins (Li et al., 2009; Singla et al., 2009), our analysis shows that these additional candidates lack at least one of the characteristic structural features of SERK proteins: five extracellular LRR domains, a proline-rich region, a transmembrane domain, and an intracellular kinase domain (Supplemental Figure 15A and 15B) (Hecht et al., 2001). In addition, only OsSERK1 and OsSERK2 cluster with the five Arabidopsis SERKs whereas the next 10 closest rice homologs that contain five extracellular LRR domains do not (Supplemental Figure 15A and 15B).
In Arabidopsis, the five SERK proteins are involved in diverse signaling pathways and are often functionally redundant (Li, 2010). SERK1, SERK2, SERK3, and SERK4 interact with BRI1 and function as positive regulators of BL signaling (Li et al., 2002; Nam and Li, 2002; Karlova et al., 2006; Albrecht et al., 2008; Gou et al., 2012). SERK1 and SERK2 play redundant roles in male sporogenesis (Albrecht et al., 2005). SERK1 has recently been shown to be involved in organ separation in flowers (Lewis et al., 2010). Both SERK3 and SERK4 regulate cell death and senescence (He et al., 2007; Kemmerling et al., 2007). Importantly, SERK3 and SERK4 are also both required for FLS2-, EFR-, and PEPR1/2-mediated innate immune responses (Roux et al., 2011; Schwessinger et al., 2011).
These observations suggest that the SERK proteins in Arabidopsis may have undergone functional diversification/specification. The fact that the overexpression of the two rice SERK-like proteins Os02g18320 and Os06g12120 is able to partially rescue the BR-insensitive phenotype of the bri1-5 mutation in Arabidopsis suggests that rice SERK-like proteins (Supplemental Figure 15A and 15B) can also fulfill functions previously only attributed to SERK proteins (Li et al., 2009).
In Arabidopsis, SERK3 and SERK4 are required for innate immunity to biotrophic, hemibiotrophic, and necotrophic pathogens (Kemmerling et al., 2007; Roux et al., 2011; Schwessinger et al., 2011). In contrast, only OsSERK2, but not OsSERK1, significantly contributes to rice immunity to the biotrophic bacterial pathogen Xoo (Figures 2–4 and Supplemental Figures 4–6) and the hemibiotrophic fungal pathogen M. oryzae (Hu et al., 2005; Park et al., 2011). OsSERK2 may mediate its immunity against M. oryzae through a yet-to-be characterized immune receptor. Suppression of OsSerk2 expression in transgenic calli by RNA interference results in a significant reduction in the rate of shoot regeneration indicating that OsSERK2 is a positive regulator of somatic embryogenesis in rice (Hu et al., 2005). In contrast, overexpression of OsSerk2 increases the rate of shoot regeneration (Hu et al., 2005). OsSERK2 is also involved in BR hormone signaling in rice (Figures 5, 6C, and Supplemental Figure 9). OsSerk2-silenced rice plants show a similar morphology to the Osbri1 mutant (Nakamura et al., 2006) (Figure 5), are less sensitive to exogenous BR application (Supplemental Figure 8), and OsSERK2 directly interacts with OsBRI1 (Figure 6C). The fact that transgenic overexpression of OsSerk2 in the Arabidopsis bri1-5 mutant partially rescues its BR-insensitive phenotype (Li et al., 2009) also supports that OsSERK2 functions in BR hormone signaling. It is clear that OsSERK2 functions in signaling pathways regulating multiple developmental programs and rice innate immune responses. How OsSERK2 regulates these multiple signaling pathways and whether these pathways are cross-co-regulated remain to be determined. Recent studies investigating the crosstalk between BR-mediated growth and innate immune signaling in Arabidopsis reached conflicting conclusions on the requirement of SERK3 (Albrecht et al., 2012; Belkhadir et al., 2012).
OsSERK2 is phylogenetically most closely related to Arabidopsis SERK1 and SERK2 (Figure 1) and its in vitro autophosphorylation pattern is closest to that of SERK1 (Table 1) but not SERK3 and SERK4. It would be informative to test whether OsSERK2 is able to complement immune related phenotypes of SERK3 mutants in Arabidopsis and whether any of the differential autophosphorylation sites are involved in this process. These experiments might determine whether the phylogenetic diversification of the SERK gene family in Arabidopsis is driven by functional specification of certain family members and their specific phosphorylation pattern.
Mechanistic Differences between Rice and Arabidopsis in the Perception of Conserved Microbial Signatures
Here, we report the functional conservation of one rice SERK protein and investigate it function in innate immune signaling mediated by three PRRs. SERK proteins are involved in the immune response towards a plethora of distinct pathogens in multiple plant species including Nicotiana benthamiana, Solanum lycopersicum, Lactuca sativa L., Oryza sativa, and Arabidopsis thaliana (Hu et al., 2005; Kemmerling et al., 2007; Fradin et al., 2009; Santos and Aragao, 2009; Chaparro-Garcia et al., 2011; Mantelin et al., 2011; Park et al., 2011; Roux et al., 2011; Schwessinger et al., 2011). The molecular mechanism of how SERK proteins exert their function in these immune pathways is well studied only in the case of SERK3/BAK1 in Arabidopsis. SERK3/BAK1 was shown to interact with multiple PRRs such as FLS2 and EFR in a ligand-dependent and kinase-independent manner in planta using whole 2-week-old seedlings grown in sterile media, in sterile Arabidopsis cell cultures, or when transiently overexpressed in N. benthamiana leaves (Schulze et al., 2010; Roux et al., 2011; Schwessinger et al., 2011). This heterodimeric complex formation was shown to be quasi-instantaneous in the case of SERK3/BAK1 and FLS2 in Arabidopsis cell cultures (Schulze et al., 2010) suggesting that these proteins constitutively co-localize in plasma-membrane subdomains ready to signal. Recent crystallographic studies show that flg22 serves as molecular glue between FLS2 and BAK1 and stabilizes the complex between both ectodomains (Sun et al., 2013). This ligand-dependent rapid heterodimeric complex formation is thought to be a key molecular switch for activating plant immune receptor-mediated signaling in Arabidopsis (Albert et al., 2013).
In rice, the interaction between OsSERK2 and XA21 occurs in the absence of any ligand treatment in fully mature leaves of 4–5-week-old greenhouse-grown plants (Figure 7B) indicating a relatively strong constitutive heterodimeric complex formation. Another mechanistic difference between rice and Arabidopsis is that it appears that, in rice, the complex formation between OsSERK2 and distinct PRRs, XA21, and FLS2 requires the catalytic activity of each interacting kinase (at least in the yeast two-hybrid assay) (Figures 4C, 6, and Supplemental Figure 10). To our knowledge, direct interaction between the kinase domains of Arabidopsis SERK proteins and FLS2 or EFR in the yeast two-hybrid system have not been reported to date. The observed ability of the kinase domains of rice SERK proteins and their respective rice PRR counterparts to form constitutive heterodimeric complexes in yeast in the absence of the ligand (or any ectodomain) could be a further indication that the interaction is more strongly mediated by their kinase domains in rice when compared to Arabidopsis.
The mechanistic differences of microbial signature perception in rice and Arabidopsis are not restricted to the interaction between OsSERK2, XA21, and other PRRs. Several recent reports demonstrate the differential involvement of homologous proteins in chitin and peptidoglycan perception (PGN) when comparing rice and Arabidopsis. For example, both plant species utilize LysM-containing proteins in chitin and PGN perception. However, the mechanism with which these proteins directly bind to the corresponding conserved microbial signatures to trigger signal transduction is clearly distinct. In rice, the GPI-anchored LysM-containing proteins CEBIP, LYP4, and LYP6 directly bind to chitin and the latter two also bind bacterial PGNs (Kaku et al., 2006; Liu et al., 2012a). The rice LysM receptor-like kinase CERK1 forms direct heteromeric complexes with CeBIP and is required for chitin signaling but is most likely not involved in direct chitin binding (Shimizu et al., 2010). Therefore, rice CERK1 appears to be a downstream receptor-like kinase relaying the extracellular chitin (and potentially PGN) perception into an intracellular defense response.
In contrast, in Arabidopsis, CERK1 is the major chitin-binding protein and is required for chitin perception (Miya et al., 2007; Wan et al., 2008; Petutschnig et al., 2010). Even though Arabidopsis CEBIP is able to biochemically bind chitin, it is not involved in chitin perception (Shinya et al., 2012). This clearly suggests that, in Arabidopsis, CERK1 is the sole functional chitin receptor. In addition, CERK1 is also required for PGN perception in Arabidopsis but does not directly bind to PGN (Willmann et al., 2011). Instead, Arabidopsis LYM1 and LYM3, the two closest homologs of rice LYP4 and LYP6, specifically bind to and are required for PGN signaling but not chitin (Willmann et al., 2011).
These examples of chitin and PGN perception and the data presented in this study demonstrate that homologous proteins are involved in the perception of conserved microbial signatures in rice and Arabidopsis. Yet, the molecular mechanisms and the specific involvement of each protein can be distinct.
Model for XA21 Signal Transduction from the Plasma Membrane to the Nucleus
Our in vitro phosphorylation assays show that OsSERK2 directly transphosphorylates XA21 only if it contains parts of its TM domain. In contrast, an XA21 variant carrying the TM domain is unable to phosphorylate OsSERK2 (Figure 8 and Supplemental Figure 13), whereas an XA21 truncated protein lacking the TM domain (XA21JK) is still capable of transphosphorylating OsSERK2 (Figure 8 and Supplemental Figure 13). These results have several implications. First, they demonstrate that XA21 has the capacity to transphosphorylate OsSERK2 under the appropriate conditions in vitro, but not under conditions where XA21 contains part of the TM domain. Second, they suggest a possible mechanism by which XA21 is activated: the XA21 kinase is kept inactive by structural features mediated by its TM domain. In this scenario, ligand binding to the XA21 extracellular domain would induce conformational changes in the XA21–OsSERK2 complex and subsequently trigger transphosphorylation of XA21 by OsSERK2. These XA21-phosphorylated residues may serve as binding sites for downstream signaling components. Signaling might also be transduced by OsSERK2-mediated phosphorylation of yet to be identified downstream signaling components.
METHODS
Plant Growth, Xoo Inoculation, and Disease Resistance Determination
Transgenic Xa21, cMycXa21, and ProAXa21 plants were generated in the Kitaake genetic background (Park et al., 2008; Chen et al., 2010b). Rice IRBB3 carrying the LRR receptor kinase XA3 (Sun et al., 2004; Xiang et al., 2006) was used for Xa3-related experiments. The rice Nipponbare genetic background was used for analysis of the transcriptional expression of OsSerk1 and OsSerk2. All transgenic plants in Kitaake were grown in the greenhouse until 6 weeks of age and transferred to the growth chamber before Xoo (PXO99AZ) inoculation. IRBB3 plants, which carry the endogenous Xa3 and confer resistance to Xoo strain PXO86 (Sun et al., 2004; Xiang et al., 2006), were grown in the greenhouse until 2 months of age and transferred to the growth chamber before Xoo (PXO86) inoculation. In the greenhouse, the light intensity in photosynthetic photon flux across the spectrum from 400 to 700nm was approximately 250 μmol m−2 s−1 in spring. The growth chamber was set on a 14-h daytime period, a 28/26°C temperature cycle, and at 90% humidity. The chamber was equipped with metal halide and incandescent lights. The light intensity in the growth chamber was approximately 100 μmol m−2 s−1. Bacterial suspensions (OD600 of 0.5) of Xoo were used to inoculate rice by the scissors-dip method (Song et al., 1995). The disease lesion length and bacterial population accumulated in rice leaf were evaluated as reported previously (Chern et al., 2005). Statistical analysis was performed using the appropriate statistical analyses.
Generation of Rice Transgenic Plants and F1 Progeny
RNAi constructs OsSerk2Ri and OsSerk1Ri were introduced into Xa21, ProAXa21, or Kitaake plants through Agrobacterium-mediated transformation according to the method described previously (Chern et al., 2005). Because the Xa21 and ProAxa21 transgenic plants are mannose resistant, transgenes OsSerk2Ri and OsSerk1Ri were selected with hygromycin in our studies. The plants of transgenic line X-B-4–2 homozygous for OsSerk2Ri in the Kitaake genetic background (abbreviated as OsSerk2Ri) were used for crossing with IRBB3 to obtain Xa3OsSerk2Ri plants. The cross was performed using IRBB3 as the pollen donor. PCR-based genotyping on Xa21, OsSerk2Ri, and OsSerk1Ri was performed as described previously (Chen et al., 2010b).
RNA Extraction and Quantitative RT–PCR Analyses
Total RNA was isolated from rice plant tissues using TRIzol (Invitrogen), following the manufacturer’s instructions. Total RNA was treated with DNase I (NEB) and used for first-strand cDNA synthesis using the Invitrogen reverse transcription kit (Invitrogen) following the provided manual. Quantitative real-time PCR (qRT–PCR) was performed on a Bio-Rad CFX96 Real-Time System coupled to a C1000 Thermal Cycler (Bio-Rad). For qRT–PCR reactions, the Bio-Rad SsoFast EvaGreen Supermix was used. qRT–PCR primer pairs used were as follows: OsSerk2-Q1/Q2 (5′-TAGTCTGCGCCAAAGTCTGA-3′/5′-GCACCT GACAGTTGTGCATT-3′) for the OsSerk2 gene, OsSerk1-Q1/-Q2 (5′-TGCATTGCATAGCTTGAGGA-3′/5′-GCAGCATTC CCAAGATC AAC-3′) for the OsSerk1 gene, Xa21-Q1/-Q2 (5′-TGACACG AAG CTCATTTTGG-3′/5′-TTGATGGCATTCAGTTCGTC-3′) for the Xa21 gene, Os04g10010-Q1/-Q2 (5′-AAATGATTTGGGACCAGTCG-3′/5′-GATG GAATGTCCTCGCAAAC-3′) for Os04g10010 gene, PR10b-Q1/-Q2 (5′-GTCGCGGTGTCGGTGGAGAG-3′, 5′-ACGGC GTCGATGAATCCGGC-3′) for PR10b gene, Actin-Q1/-Q2 (5′-TCGG CTCTGAATGTACCTCCTA-3′/5′-CAC TTGAGTAAAGACTGTCACT TG-3′) for the reference gene actin. qRT–PCR reactions were run for 40 cycles with annealing at 620C for 5 s and denaturation at 95ºC for 5 s. The expression levels of OsSerk2, OsSerk1, Os04g10010, PR10b, and Xa21 were normalized to the actin gene expression level.
Constructions
All constructs were made according to supplemental experimental procedures (Supplemental Text 1).
Purification of Recombinant Proteins and Kinase Assay
Purification of GST– or His-Nus-fusion proteins and in vitro kinase and transphosphorylation assays were performed as described previously (Liu et al., 2002; Schwessinger et al., 2011).
Defense Gene Expression Analysis
Fully developed leaves of 6-week-old rice plants were cut into 2-cm-long strips and incubated for at least 12h in ddH2O to reduce residual wound signal. Leaf strips were treated with 1 μM flg22 peptide (Felix et al., 1999), purchased from Pacific Immunology, or 50 μg ml–1 chitin, purchased from Sigma, for the indicated time. Leaf tissue was snap-frozen in liquid nitrogen and processed as described above.
Yeast Two-Hybrid Assays
The Matchmaker LexA two-hybrid system (Clontech) was used for yeast two-hybrid assays. Yeast pEGY48/p8op-lacZ (Clontech) was co-transformed with the BD and AD vectors by using the Frozen-EZ yeast transformation II kit (Zymo Research) and spread on an appropriate medium following the procedures described previously (Chen et al., 2010b).
Immunoblotting
Total protein extraction from yeast, E. coli, and rice plants and immunoblotting (Western blotting) were performed as previously described (Chen et al., 2010b). The anti-OsSERK2 antibody against the synthetic peptide AELAPRHNDW-Cys of OsSERK2 (amino acids 602–611) was provided as a service by Pacific Immunology. Detailed information about their methods can be obtained at Pacific Immunology (www.pacificimmunology.com/). Anti-OsSERK2 for detection of OsSERK2, anti-LexA (Clontech) for detection of LexA-fused protein produced from BD vectors, anti-HA (Covance) for detection of HA-fused protein produced from AD vectors, and anti-Myc (Santa Cruze Biotech) for detection of XA21 with Myc tag were used as primary antibodies.
Co-Immunoprecipitation of Rice Proteins
Detached rice leaves from 6-week-old cMyc–Xa21 or Kit plants were cut into 4-cm-long pieces and snap-frozen. Myc–XA21 complex was immunoprecipitated using the agarose-conjugated anti-Myc antibody (Santa Cruz) following the method described previously (Roux et al., 2011; Schwessinger et al., 2011) with slight adaptation. The immunoprecipitates were then probed with anti-Myc and anti-OsSERK2, respectively, after being separated by SDS–PAGE.
Brassinolide (BL) Treatment
Seeds from Xa21, A814–178, and A814–186 were sterilized with 30% bleach for 20min. After rinsing with distilled ddH2O four times, they were germinated in a growth chamber at 30°C on MS agar in the presence or absence of 0 μM, 0.001 μM, 0.01 μM, and 0.1 μM of 24-epiBL (Sigma). Seedlings were examined 5 d after germination.
Phylogenetic and Molecular Evolutionary Analyses
Phylogenetic and molecular evolutionary analyses were conducted using MUSCLE in the Geneious (Biomatters) environment. One thousand bootstraps were adopted to infer the statistical support for the tree.
Tandem Mass Spectrometry (LC–MS/MS)
Samples were analyzed on an Agilent 6550 iFunnel Q-TOF mass spectrometer (Agilent Technologies) coupled to an Agilent 1290 LC system (Agilent). Peptide samples were loaded onto a Ascentis Peptides ES-C18 column (2.1mm × 100mm, 2.7-μm particle size; Sigma-Aldrich, St. Louis, MO) via an Infinity Autosampler (Agilent Technologies) with buffer A (2% Acetonitrile, 0.1% Formic Acid) flowing at 0.400ml min–1. Peptides were eluted into the mass spectrometer via a gradient with initial starting conditions of 5% buffer B (98% acetonitrile, 0.1% formic acid) increasing to 35% B over 5.5min. Subsequently, B was increased to 90% over 30 s and held for 3min at a flow rate of 0.6ml min–1 followed by a ramp back down to 5% B over 1 min, where it was held for 2.5min to re-equilibrate the column. Peptides were introduced to the mass spectrometer from the LC by using a Jet Stream source (Agilent Technologies) operating in positive-ion mode (5000V). The data were acquired with the Agilent MassHunter Workstation Software, LC/MS Data Acquisition B.05.00 (Build 5.0.5042.2) operating in Auto MS/MS mode whereby the five most intense ions (charge states 2–5) within a 300–1400-m/z mass range above a threshold of 1000 counts were selected for MS/MS analysis. MS/MS spectra were collected with the quadrupole set to ‘Medium’ resolution and collision energy dependent on the m/z to optimize fragmentation (3.6 × (m/z)/100 – 4.8). MS/MS spectra were scanned from 100 to 1700 m/z and were acquired until 45000 total counts were collected or for a maximum accumulation time of 333ms. Former parent ions were excluded for 0.1min following selection for MS/MS acquisition.
Analysis of Tandem Mass Spectrometry Data
Mass spectral data were initially examined in the Agilent MassHunter Workstation Software, Qualitative Analysis B.05.00 (Build 5.0.519.13 Service Pack 1). All MSMS data were exported in MGF format from the Qualitative Analysis software using; Absolute height ≥ 20 counts, Relative height ≥ 0.100% of largest peak, Maximum number of peaks (limited by height) to the largest 300, Peak spacing tolerance 0.0025 m/z plus 7.0 p.p.m., Isotope model: Peptides, Limit of assigned charge states to a maximum of 5. Resultant.mgf files were used to interrogate the Mascot search engine version 2.3.02 (Matrix Science) with a peptide tolerance of ±20 p.p.m. and MS/MS tolerance of ±0.1Da; variable modifications were Oxidation (M), Phospho (ST), Phospho (Y); up to one missed cleavage for trypsin; require bold red; and the instrument type was set to ESI-QUAD-TOF. Searches were performed against the current protein set (all.pep, release 7.0) from the MSU Rice Genome Annotation Project (Kawahara et al., 2013) including standard contaminants (keratin, trypsin, GFP, BSA, etc.) resulting in a database with 66506 sequences and 29649083 residues. An initial ions score or expect cut-off of 20 was applied to filter low-scoring phosphopeptide matches. All phosphopeptide matches were manually inspected and annotated to confirm the modification. A minimum of two independent spectra were inspected for each phosphorylation site.
SUPPLEMENTARY DATA
Supplementary Data are available at Molecular Plant Online.
FUNDING
This work was supported by NIH GM59962 to P.C.R. Dr. X.C. was also supported by the National Natural Science Foundation of China (NSFC 31171622), the Sichuan ‘Hundred Talents Plan’ fund, and Sichuan Agricultural University ‘High Talents’ start-up fund in China. B.S. was supported by an EMBO long-term fellowship. B.S. is supported by a Human Frontiers Science Program fellowship.
Supplementary Material
ACKNOWLEDGMENTS
We are grateful to Dr. Wenming Wang (Sichuan Agricultural University, Chengdu, Sichuan, China) for his helpful discussion on this manuscript. Conceived and designed the experiments: X.C., B.S., S.Z., and P.C.R. Performed the experiments: X.C., B.S., S.Z., P.E.C., D.R., X.Z., A.D., C.P., J.H., and J.W. Analyzed the data: B.S., X.C, S.Z., M.C., C.P., J.H., and P.C.R. Wrote the paper: B.S., X.C., M.C., and P.C.R. No conflict of interest declared.
REFERENCES
- Albert M., Jehle A.K., Furst U., Chinchilla D., Boller T., Felix G. (2013). A two-hybrid-receptor assay demonstrates heteromer formation as switch-on for plant immune receptors. Plant Physiol. 163, 1504–1509 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Albert M., Jehle A.K., Lipschis M., Mueller K., Zeng Y., Felix G. (2010). Regulation of cell behaviour by plant receptor kinases: pattern recognition receptors as prototypical models. Eur. J. Cell Biol. 89, 200–207 [DOI] [PubMed] [Google Scholar]
- Albrecht C., Boutrot F., Segonzac C., Schwessinger B., Gimenez-Ibanez S., Chinchilla D., Rathjen J.P., de Vries S.C., Zipfel C. (2012). Brassinosteroids inhibit pathogen-associated molecular pattern-triggered immune signaling independent of the receptor kinase BAK1. Proc. Natl Acad. Sci. U S A. 109, 303–308 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Albrecht C., Russinova E., Hecht V., Baaijens E., de Vries S. (2005). The Arabidopsis thaliana SOMATIC EMBRYOGENESIS RECEPTOR-LIKE KINASES1 and 2 control male sporogenesis. Plant Cell. 17, 3337–3349 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Albrecht C., Russinova E., Kemmerling B., Kwaaitaal M., de Vries S.C. (2008). Arabidopsis SOMATIC EMBRYOGENESIS RECEPTOR KINASE proteins serve brassinosteroid-dependent and -independent signaling pathways. Plant Physiol. 148, 611–619 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andaya C.B., Ronald P.C. (2003). A catalytically impaired mutant of rice Xa21 receptor kinase confers partial resistance to Xanthomonas oryzae pv oryzae . Physiological and Molecular Plant Pathology. 62, 203–208 [Google Scholar]
- Belkhadir Y., Jaillais Y., Epple P., Balsemao-Pires E., Dangl J.L., Chory J. (2012). Brassinosteroids modulate the efficiency of plant immune responses to microbe-associated molecular patterns. Proc. Natl Acad. Sci. U S A. 109, 297–302 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Century K.S., Lagman R.A., Adkisson M., Morlan J., Tobias R., Schwartz K., Smith A., Love J., Ronald P.C., Whalen M.C. (1999). Short communication: developmental control of Xa21-mediated disease resistance in rice. Plant J. 20, 231–236 [DOI] [PubMed] [Google Scholar]
- Chaparro-Garcia A., Wilkinson R.C., Gimenez-Ibanez S., Findlay K., Coffey M.D., Zipfel C., Rathjen J.P., Kamoun S., Schornack S. (2011). The receptor-like kinase SERK3/BAK1 is required for basal resistance against the late blight pathogen phytophthora infestans in Nicotiana benthamiana . PLoS One. 6, e16608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen X., Ronald P.C. (2011). Innate immunity in rice. Trends Plant Sci. 16, 451–459 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen X., Chern M., Canlas P.E., Jiang C., Ruan D., Cao P., Ronald P.C. (2010a). A conserved threonine residue in the juxtamembrane domain of the XA21 pattern recognition receptor is critical for kinase autophosphorylation and XA21-mediated immunity. J. Biol. Chem. 285, 10454–10463 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen X., Chern M., Canlas P.E., Ruan D., Jiang C., Ronald P.C. (2010b). An ATPase promotes autophosphorylation of the pattern recognition receptor XA21 and inhibits XA21-mediated immunity. Proc. Natl Acad. Sci. U S A. 107, 8029–8034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chern M.S., P.E. C., Fitzgerald H., Ronald P.C. (2005). NRR, a negative regulator of disease resistance in rice that interacts with Arabidopsis NPR1 and rice NH1. Plant J. 43, 623–635 [DOI] [PubMed] [Google Scholar]
- Chinchilla D., Bauer Z., Regenass M., Boller T., Felix G. (2006). The Arabidopsis receptor kinase FLS2 binds flg22 and determines the specificity of flagellin perception. Plant Cell. 18, 465–476 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chinchilla D., Shan L., He P., de Vries S., Kemmerling B. (2009). One for all: the receptor-associated kinase BAK1. Trends Plant Sci. 14, 535–541 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chinchilla D., Zipfel C., Robatzek S., Kemmerling B., Nurnberger T., Jones J.D., Felix G., Boller T. (2007). A flagellin-induced complex of the receptor FLS2 and BAK1 initiates plant defence. Nature. 448, 497–500 [DOI] [PubMed] [Google Scholar]
- Clouse S.D. (2011). Brassinosteroid signal transduction: from receptor kinase activation to transcriptional networks regulating plant development. Plant Cell. 23, 1219–1230 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dardick C., Schwessinger B., Ronald P. (2012). Non-arginine-aspartate (non-RD) kinases are associated with innate immune receptors that recognize conserved microbial signatures. Curr. Opin. Plant Biol. 15, 358–366 [DOI] [PubMed] [Google Scholar]
- Ding B., Bellizzi, Mdel R., Ning Y., Meyers B.C., Wang G.L. (2012). HDT701, a histone H4 deacetylase, negatively regulates plant innate immunity by modulating histone H4 acetylation of defense-related genes in rice. Plant Cell. 24, 3783–3794 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Felix G., Duran J.D., Volko S., Boller T. (1999). Plants have a sensitive perception system for the most conserved domain of bacterial flagellin. Plant J. 18, 265–276 [DOI] [PubMed] [Google Scholar]
- Fradin E., Adb-El-Haliem A., Masini L., van den Berg G., Joosten M., Thomma B. (2011). Interfamily transfer of tomato Ve1 mediates Verticillium resistance in Arabidopsis . Plant Physiol. 156, 2255–2265 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fradin E.F., Zhang Z., Juarez, Ayala J.C., Castroverde C.D., Nazar R.N., Robb J., Liu C.M., Thomma B.P. (2009). Genetic dissection of Verticillium wilt resistance mediated by tomato Ve1. Plant Physiol. 150, 320–332 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gomez-Gomez L., Boller T. (2000). FLS2: an LRR receptor-like kinase involved in the perception of the bacterial elicitor flagellin in Arabidopsis . Mol. Cell. 5, 1003–1011 [DOI] [PubMed] [Google Scholar]
- Gou X., Yin H., He K., Du J., Yi J., Xu S., Lin H., Clouse S.D., Li J. (2012). Genetic evidence for an indispensable role of somatic embryogenesis receptor kinases in brassinosteroid signaling. PLoS Genet. 8, e1002452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- He K., Gou X., Yuan T., Lin H., Asami T., Yoshida S., Russell S.D., Li J. (2007). BAK1 and BKK1 regulate brassinosteroid-dependent growth and brassinosteroid-independent cell-death pathways. Curr. Biol. 17, 1109–1115 [DOI] [PubMed] [Google Scholar]
- Hecht V., Vielle-Calzada J.-P., Hartog M.V., Schmidt E.D.L., Boutilier K., Grossniklaus U., de Vries S.C. (2001). The Arabidopsis Somatic Embryogenesis Receptor Kinase 1 gene is expressed in developing ovules and embryos and enhances embryogenic competence in culture. Plant Physiol. 127, 803–816 [PMC free article] [PubMed] [Google Scholar]
- Heese A., Hann D.R., Gimenez-Ibanez S., Jones A.M., He K., Li J., Schroeder J.I., Peck S.C., Rathjen J.P. (2007). The receptor-like kinase SERK3/BAK1 is a central regulator of innate immunity in plants. Proc. Natl Acad. Sci. U S A. 104, 12217–12222 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu H., Xiong L., Yang Y. (2005). Rice SERK1 gene positively regulates somatic embryogenesis of cultured cell and host defense response against fungal infection. Planta. 222, 107–117 [DOI] [PubMed] [Google Scholar]
- Kaku H., Nishizawa Y., Ishii-Minami N., Akimoto-Tomiyama C., Dohmae N., Takio K., Minami E., Shibuya N. (2006). Plant cells recognize chitin fragments for defense signaling through a plasma membrane receptor. Proc. Natl Acad. Sci. U S A. 103, 11086–11091 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karlova R., Boeren S., Russinova E., Aker J., Vervoort J., de Vries S. (2006). The Arabidopsis SOMATIC EMBRYOGENESIS RECEPTOR-LIKE KINASE1 protein complex includes BRASSINOSTEROID-INSENSITIVE1. Plant Cell. 18, 626–638 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karlova R., Boeren S., van Dongen W., Kwaaitaal M., Aker J., Vervoot J., de Vries S. (2009). Identification of in vitro phosphorylation sites in the Arabidopsis thaliana somatic embryogenesis receptor-like kinases. Proteomics. 9, 368–379 [DOI] [PubMed] [Google Scholar]
- Kawahara Y., de la Bastide M., Hamilton J.P., Kanamori H., McCombie W.R., Ouyang S., Schwartz D.C., Tanaka T., Wu J., Zhou S., et al. (2013). Improvement of the Oryza sativa Nipponbare reference genome using next generation sequence and optical map data. Rice. 6, 1–10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kemmerling B., Schwedt A., Rodriguez P., Mazzotta S., Frank M., Qamar S.A., Mengiste T., Betsuyaku S., Parker J.E., Mussig C., et al. (2007). The BRI1-associated kinase 1, BAK1, has a brassinolide-independent role in plant cell-death control. Curr. Biol. 17, 1116–1122 [DOI] [PubMed] [Google Scholar]
- Krol E., Mentzel T., Chinchilla D., Boller T., Felix G., Kemmerling B., Postel S., Arents M., Jeworutzki E., Al-Rasheid K.A., et al. (2010). Perception of the Arabidopsis danger signal peptide 1 involves the pattern recognition receptor AtPEPR1 and its close homologue AtPEPR2. J. Biol. Chem. 285, 13471–13479 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lemaitre B., Nicolas E., Michaut L., Reichhart J.-M., Hoffmann J.A. (1996). The dorsoventral regulatory gene cassette spatzle/toll/cactus controls the potent antifungal response in Drosophila adults. Cell. 86, 973–983 [DOI] [PubMed] [Google Scholar]
- Lewis M.W., Leslie M.E., Fulcher E.H., Darnielle L., Healy P.N., Youn J.Y., Liljegren S.J. (2010). The SERK1 receptor-like kinase regulates organ separation in Arabidopsis flowers. Plant J. 62, 817–828 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li D., Wang L., Wang M., Xu Y.Y., Luo W., Liu Y.J., Xu Z.H., Li J., Chong K. (2009). Engineering OsBAK1 gene as a molecular tool to improve rice architecture for high yield. Plant Biotechnol. J. 7, 791–806 [DOI] [PubMed] [Google Scholar]
- Li J. (2010). Multi-tasking of somatic embryogenesis receptor-like protein kinases. Curr. Opin. Plant Biol. 13, 509–514 [DOI] [PubMed] [Google Scholar]
- Li J., Chory J. (1997). A putative leucine-rich repeat receptor kinase involved in brassinosteroid signal transduction. Cell. 90, 929–938 [DOI] [PubMed] [Google Scholar]
- Li J., Wen J., Lease K.A., Doke J.T., Tax F.E., Walker J.C. (2002). BAK1, an Arabidopsis LRR receptor-like protein kinase, interacts with BRI1 and modulates brassinosteroid signaling. Cell. 110, 213–222 [DOI] [PubMed] [Google Scholar]
- Liu B., Li J.F., Ao Y., Qu J., Li Z., Su J., Zhang Y., Liu J., Feng D., Qi K., et al. (2012a). Lysin motif-containing proteins LYP4 and LYP6 play dual roles in peptidoglycan and chitin perception in rice innate immunity. Plant Cell. 24, 3406–3419 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu G.Z., Pi L.Y., Walker J.C., Ronald P.C., Song W.Y. (2002). Biochemical characterization of the kinase domain of the rice disease resistance receptor-like kinase XA21. J. Biol. Chem. 277, 20264–20269 [DOI] [PubMed] [Google Scholar]
- Liu T., Liu Z., Song C., Hu Y., Han Z., She J., Fan F., Wang J., Jin C., Chang J., et al. (2012b). Chitin-induced dimerization activates a plant immune receptor. Science. 336, 1160–1164 [DOI] [PubMed] [Google Scholar]
- Mantelin S., Peng H.C., Li B., Atamian H.S., Takken F.L., Kaloshian I. (2011). The receptor-like kinase SlSERK1 is required for Mi-1-mediated resistance to potato aphids in tomato. Plant J. 67, 459–471 [DOI] [PubMed] [Google Scholar]
- Medzhitov R., Preston-Hurlburt P., Janeway C.A., Jr (1997). A human homologue of the Drosophila Toll protein signals activation of adaptive immunity. Nature. 388, 394–397 [DOI] [PubMed] [Google Scholar]
- Miki D., Shimamoto K. (2004). Simple RNAi vectors for stable and transient suppression of gene function in rice. Plant Cell Physiol. 45, 490–495 [DOI] [PubMed] [Google Scholar]
- Miya A., Albert P., Shinya T., Desaki Y., Ichimura K., Shirasu K., Narusaka Y., Kawakami N., Kaku H., Shibuya N. (2007). CERK1, a LysM receptor kinase, is essential for chitin elicitor signaling in Arabidopsis . Proc. Natl Acad. Sci. U S A. 104, 19613–19618 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakamura A., Fujioka S., Sunohara H., Kamiya N., Hong Z., Inukai Y., Miura K., Takatsuto S., Yoshida S., Ueguchi-Tanaka M., et al. (2006). The role of OsBRI1 and its homologous genes, OsBRL1 and OsBRL3, in rice. Plant Physiol. 140, 580–590 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nam K.H., Li J. (2002). BRI1/BAK1, a receptor kinase pair mediating brassinosteroid signaling. Cell. 110, 203–212 [DOI] [PubMed] [Google Scholar]
- Nolen B., Taylor S., Ghosh G. (2004). Regulation of protein kinases; controlling activity through activation segment conformation. Mol. Cell. 15, 661–675 [DOI] [PubMed] [Google Scholar]
- Ntoukakis V., Schwessinger B., Segonzac C., Zipfel C. (2011). Cautionary notes on the use of C-terminal BAK1 fusion proteins for functional studies. Plant Cell. 23, 3871–3878 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oh M.H., Wang X., Wu X., Zhao Y., Clouse S.D., Huber S.C. (2010). Autophosphorylation of Tyr-610 in the receptor kinase BAK1 plays a role in brassinosteroid signaling and basal defense gene expression. Proc. Natl Acad. Sci. U S A. 107, 17827–17832 [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Park C.J., Bart R., Chern M., Canlas P.E., Bai W., Ronald P.C. (2010). Overexpression of the endoplasmic reticulum chaperone BiP3 regulates XA21-mediated innate immunity in rice. PLoS One. 5, e9262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park C.J., Peng Y., Chen X., Dardick C., Ruan D., Bart R., Canlas P.E., Ronald P.C. (2008). Rice XB15, a protein phosphatase 2C, negatively regulates cell death and XA21-mediated innate immunity. PLoS Biol. 6, e231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park C.J., Sharma R., Lefebvre B., Canlas P.E., Ronald P.C. (2013). The endoplasmic reticulum-quality control component SDF2 is essential for XA21-mediated immunity in rice. Plant Sci. 210, 53–60 [DOI] [PubMed] [Google Scholar]
- Park H.S., Ryu H.Y., Kim B.H., Kim S.Y., Yoon I.S., Nam K.H. (2011). A subset of OsSERK genes, including OsBAK1, affects normal growth and leaf development of rice. Mol. Cells. 32, 561–569 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petutschnig E.K., Jones A.M., Serazetdinova L., Lipka U., Lipka V. (2010). The LysM-RLK CERK1 is a major chitin binding protein in Arabidopsis thalianaand subject to chitin-induced phosphorylation. J. Biol. Chem. 285, 28902–28911 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poltorak A., He X., Smirnova I., Liu M.Y., Van, Huffel C., Du X., Birdwell D., Alejos E., Silva M., Galanos C., et al. (1998). Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: mutations in Tlr4 gene. Science. 282, 2085–2088 [DOI] [PubMed] [Google Scholar]
- Postel S., Kufner I., Beuter C., Mazzotta S., Schwedt A., Borlotti A., Halter T., Kemmerling B., Nurnberger T. (2009). The multifunctional leucine-rich repeat receptor kinase BAK1 is implicated in Arabidopsis development and immunity. Eur. J. Cell Biol. 89, 169. [DOI] [PubMed] [Google Scholar]
- Ronald P.C., Beutler B. (2010). Plant and animal sensors of conserved microbial signatures. Science. 330, 1061–1064 [DOI] [PubMed] [Google Scholar]
- Roux M., Schwessinger B., Albrecht C., Chinchilla D., Jones A., Holton N., Malinovsky F.G., Tor M., de Vries S., Zipfel C. (2011). The Arabidopsis leucine-rich repeat receptor-like kinases BAK1/SERK3 and BKK1/SERK4 are required for innate immunity to hemibiotrophic and biotrophic pathogens. Plant Cell. 23, 2440–2455 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santos M.O., Aragao F.J. (2009). Role of SERK genes in plant environmental response. Plant Signal Behav. 4, 1111–1113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schulze B., Mentzel T., Jehle A.K., Mueller K., Beeler S., Boller T., Felix G., Chinchilla D. (2010). Rapid heteromerization and phosphorylation of ligand-activated plant transmembrane receptors and their associated kinase BAK1. J. Biol. Chem. 285, 9444–9451 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwessinger B., Ronald P.C. (2012). Plant innate immunity: perception of conserved microbial signatures. Annu. Rev. Plant Biol. 63, 451–482 [DOI] [PubMed] [Google Scholar]
- Schwessinger B., Roux M., Kadota Y., Ntoukakis V., Sklenar J., Jones A., Zipfel C. (2011). Phosphorylation-dependent differential regulation of plant growth, cell death, and innate immunity by the regulatory receptor-like kinase BAK1. PLoS Genet. 7, e1002046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shimizu T., Nakano T., Takamizawa D., Desaki Y., Ishii-Minami N., Nishizawa Y., Minami E., Okada K., Yamane H., Kaku H., et al. (2010). Two LysM receptor molecules, CEBiP and OsCERK1, cooperatively regulate chitin elicitor signaling in rice. Plant J. 64, 20–14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shinya T., Motoyama N., Ikeda A., Wada M., Kamiya K., Hayafune M., Kaku H., Shibuya N. (2012). Functional characterization of CEBiP and CERK1 homologs in Arabidopsis and rice reveals the presence of different chitin receptor systems in plants. Plant Cell Physiol. 53, 1696–1706 [DOI] [PubMed] [Google Scholar]
- Singla B., Khurana J.P., Khurana P. (2009). Structural characterization and expression analysis of the SERK/SERL gene family in rice (Oryza sativa). Int. J. Plant Genomics. 2009, 539402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song W.Y., Wang G.L., Chen L.L., Kim H.S., Pi L.Y., Holsten T., Gardner J., Wang B., Zhai W.X., Zhu L.H., et al. (1995). A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science. 270, 1804–1806 [DOI] [PubMed] [Google Scholar]
- Sun X., Cao Y., Yang Z., Xu C., Li X., Wang S., Zhang Q. (2004). Xa26, a gene conferring resistance to Xanthomonas oryzae pv. oryzae in rice, encodes an LRR receptor kinase-like protein. Plant J. 37, 517–527 [DOI] [PubMed] [Google Scholar]
- Sun Y., Li L., Macho A.P., Han Z., Hu Z., Zipfel C., Zhou J.M., Chai J. (2013). Structural basis for flg22-induced activation of the Arabidopsis FLS2–BAK1 immune complex. Science. 342, 624–628 [DOI] [PubMed] [Google Scholar]
- Takai R., Isogai A., Takayama S., Che F.S. (2008). Analysis of flagellin perception mediated by flg22 receptor OsFLS2 in rice. Mol. Plant Microbe Interact. 21, 1635–1642 [DOI] [PubMed] [Google Scholar]
- Takeda K. (1977). Internode elongation and dwarfism in some gramineous plants. Gamma Field Symp, 16, 1–18 [Google Scholar]
- Wan J., Zhang X.C., Neece D., Ramonell K.M., Clough S., Kim S.Y., Stacey M.G., Stacey G. (2008). A LysM receptor-like kinase plays a critical role in chitin signaling and fungal resistance in Arabidopsis . Plant Cell. 20, 471–481 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang G.L., Ruan D.L., Song W.Y., Sideris S., Chen L., Pi L.Y., Zhang S., Zhang Z., Fauquet C., Gaut B.S., et al. (1998). Xa21D encodes a receptor-like molecule with a leucine-rich repeat domain that determines race-specific recognition and is subject to adaptive evolution. Plant Cell. 10, 765–779 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang X., Goshe M.B., Soderblom E.J., Phinney B.S., Kuchar J.A., Li J., Asami T., Yoshida S., Huber S.C., Clouse S.D. (2005). Identification and functional analysis of in vivo phosphorylation sites of the Arabidopsis BRASSINOSTEROID-INSENSITIVE1 receptor kinase. Plant Cell. 17, 1685–1703 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang X., Kota U., He K., Blackburn K., Li J., Goshe M.B., Huber S.C., Clouse S.D. (2008). Sequential transphosphorylation of the BRI1/BAK1 receptor kinase complex impacts early events in brassinosteroid signaling. Dev. Cell. 15, 220–235 [DOI] [PubMed] [Google Scholar]
- Wang Z., Longo P.A., Tarrant M.K., Kim K., Head S., Leahy D.J., Cole P.A. (2011). Mechanistic insights into the activation of oncogenic forms of EGF receptor. Nat. Struct. Mol. Biol. 18, 1388–1393 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Willmann R., Lajunen H.M., Erbs G., Newman M.A., Kolb D., Tsuda K., Katagiri F., Fliegmann J., Bono J.J., Cullimore J.V., et al. (2011). Arabidopsis lysin-motif proteins LYM1 LYM3 CERK1 mediate bacterial peptidoglycan sensing and immunity to bacterial infection. Proc. Natl Acad. Sci. U S A. 108, 19824–19829 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xiang Y., Cao Y., Xu C., Li X., Wang S. (2006). Xa3, conferring resistance for rice bacterial blight and encoding a receptor kinase-like protein, is the same as Xa26. Theor. Appl. Genet. 113, 1347–1355 [DOI] [PubMed] [Google Scholar]
- Yamaguchi Y., Huffaker A., Bryan A.C., Tax F.E., Ryan C.A. (2010). PEPR2 is a second receptor for the Pep1 and Pep2 peptides and contributes to defense responses in Arabidopsis . Plant Cell. 22, 508–522 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamaguchi Y., Pearce G., Ryan C.A. (2006). The cell surface leucine-rich repeat receptor for AtPep1, an endogenous peptide elicitor in Arabidopsis, is functional in transgenic tobacco cells. Proc. Natl Acad. Sci. U S A. 103, 10104–10109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zipfel C., Kunze G., Chinchilla D., Caniard A., Jones J.D., Boller T., Felix G. (2006). Perception of the bacterial PAMP EF-Tu by the receptor EFR restricts Agrobacterium-mediated transformation. Cell. 125, 749–760 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.