

Abstract
Notch intercellular signaling is critical for diverse developmental pathways and for homeostasis in various types of stem cells and progenitor cells. Because Notch gene products need to be precisely regulated spatially and temporally, epigenetics is likely to help control expression of Notch signaling genes. Reduced representation bisulfite sequencing (RRBS) indicated significant hypomethylation in myoblasts, myotubes, and skeletal muscle vs. many nonmuscle samples at intragenic or intergenic regions of the following Notch receptor or ligand genes: NOTCH1, NOTCH2, JAG2, and DLL1. An enzymatic assay of sites in or near these genes revealed unusually high enrichment of 5-hydroxymethylcytosine (up to 81%) in skeletal muscle, heart, and cerebellum. Epigenetics studies and gene expression profiles suggest that hypomethylation and/or hydroxymethylation help control expression of these genes in heart, brain, myoblasts, myotubes, and within skeletal muscle myofibers. Such regulation could promote cell renewal, cell maintenance, homeostasis, and a poised state for repair of tissue damage.
Keywords: DNA methylation, 5-hydroxymethylcytosine, notch signaling, enhancers, histone H3 methylation, DNaseI hypersensitivity, myoblasts, muscle, brain, heart
Introduction
Notch intercellular signaling is essential to cell fate decisions during differentiation, especially in stem cells and progenitor cells. It is also involved in controlling proliferation, survival, and homeostasis in many cell types. Notch signaling genes are targets for pathogenic mutations, including those associated with cancer.1,2 In the skeletal muscle lineage, Notch signaling is implicated in the quiescence of postnatal muscle stem cells (satellite cells), the proliferation of myoblasts (Mb)—which are activated satellite cells—and the transient inhibition of terminal differentiation of Mb into multinucleated myotubes (Mt).3-6 Among other cell types that Notch signaling guides into self-renewal or differentiation pathways are cardiac7 and neural stem cells.8 In canonical Notch signaling, the Notch receptor protein (NOTCH1, NOTCH2, NOTCH3, or NOTCH4 in human cells) spans the cell membrane as a heterodimer of the proteolytically cleaved precursor protein. The heterodimer is activated upon interaction with a Notch ligand (DLL1, DLL3, DLL4, JAG1, or JAG2) on the membrane of an adjacent cell and then cleaved to yield a NOTCH protein fragment that migrates to the nucleus. In the nucleus, this fragmented, active form of NOTCH1 or NOTCH2 converts a transcription regulatory protein (RBPJ) from being a repressor to being an activator, thereby inducing expression of key genes.5,9-11 Different Notch receptors and ligands play different cell-type specific roles. For example, NOTCH1, but not NOTCH2 or NOTCH3, is implicated in stimulating proliferation of porcine muscle satellite cells, even though all three genes are expressed in these cells.12 Many important details of Notch signaling pathways are yet to be elucidated.
Levels of Notch receptors and ligands have to be tightly regulated in a spatial and temporal manner to control different outcomes, even in a given cell type. For example, when a muscle satellite cell is induced to divide, different levels of active Notch receptor can promote quiescence in one progeny cell and division in the other.13 Posttranscriptional regulation and posttranslational regulation play major roles in controlling levels and cellular localization of active Notch receptor and ligand proteins.14-16 Cell-type specific differences in steady-state transcript levels17-19 suggest differential transcription of these genes. Despite the importance of Notch signaling to the generation and maintenance of skeletal muscle,9,20 only one previous report described DNA methylation in the vicinity of a Notch receptor gene in the muscle lineage, and that involved only small subregions around the transcription start site (TSS) of Notch1.17 A few CpG sites showed treatment-specific differences in C2C12 mouse Mb, but that differential methylation was of uncertain statistical significance.
Here we use genome-wide profiles of DNA methylation (reduced representation bisulfite sequencing, RRBS; ENCODE21,22) from large numbers of different types of human samples, including Mb, Mt, and postnatal skeletal muscle, to identify statistically significant, differential methylation of Notch signaling genes in the muscle lineage. In addition, we analyzed genomic profiles for histone modification, open chromatin (DNaseI hypersensitivity), and transcripts (RNA-seq) to compare genes encoding Notch receptors or ligands in Mb, Mt, and many nonmuscle cell cultures. We also assayed for myogenesis-associated increases in genomic 5-hydroxymethylcytosine (5hmC) in differentially methylated regions (DMRs) within Notch signaling genes. This sixth base in vertebrate DNA plays very different roles from 5mC, e.g., 5hmC at enhancers is often associated with their activation, while 5mC is usually associated with their repression.23,24 It was recently found that 5hmC is enriched in the body of NOTCH1 and DLL1 genes during neural differentiation of human embryonic stem cells (ESC)25 and within NOTCH1, NOTCH3, DLL1, and JAG2 in human brain tissue.26 Identifying and quantifying this base in genes from skeletal muscle DNA is of particular interest because we previously found that RNA levels for the 5hmC-generating TET1 and TET2 enzymes are particularly high in Mb and Mt vs. many other normal cell cultures and that global genomic 5hmC levels are higher in skeletal muscle than in Mb, Mt, and leukocyte samples.22
Results
Hypomethylation in or near NOTCH1, JAG2, DLL1, NOTCH2, and LFNG in the skeletal muscle lineage
Analysis of CpG methylation data obtained by RRBS27 from diverse human samples indicated that NOTCH1 has very many myogenic hypomethylated CpG sites within the gene body in both adult-derived myogenic progenitor cells (Mb and Mt) and skeletal muscle tissue (Fig. 1A and D). Given the importance of NOTCH1 receptor protein in many developmental, homeostasis, and disease pathways1,2 and to skeletal muscle formation in particular,3-6 we focused on the DNA and chromatin epigenetics of NOTCH1 and other Notch signaling genes. The determination of myogenic DM sites or DMRs involved comparison of 9 Mb or Mt samples as a set (MbMt) with 16 types of cell cultures derived from noncancerous tissue, and also skeletal muscle with 14 types of nonmuscle tissue (ENCODE21,22,28). Mb and Mt were considered as a set because they showed very modest numbers of differences in their DNA methylation between each other, compared with their differences from nonmuscle cell cultures.22 NOTCH1 had 16 MbMt-hypomethylated and 27 skeletal muscle-hypomethylated CpGs that were clustered in a 2.8-kb subregion of intron 2. Only six genes displaying myogenic hypomethylation (out of 2595 genes with associated MbMt-hypomethylated sites and 3165 genes with skeletal muscle-hypomethylated sites22) had more intragenic sites in both myogenic progenitor cells and muscle tissue (PRKAR1B, CBFA2T3, MAD1L1, HDAC4, KCNAB2, and PTPRN2 with 21–54 and 30–87 MbMt- and muscle-hypomethylated sites, respectively).

Figure 1. Myogenesis-associated hypomethylation and chromatin epigenetic marks in NOTCH1 intron 2: whole-gene view. (A) Significantly hypomethylated (green) or hypermethylated (red) DMRs for the set of Mb +Mt samples vs. 16 types of nonmuscle cell cultures and for skeletal muscle vs. 14 types of nonmuscle tissue. Wide green bars, the DMR encompassing MbMt- or muscle-hypomethylated CpG sites. (B)The predicted chromatin structure (enh, enhancer; prom, promoter; transcr, transcriptionally active) based mostly on histone modifications.57 (C) DNaseI-hypersensitivity mapping by DNaseI-seq.22 (D) Examples of RRBS data and the positions of CpG islands. Using an 11-color, semicontinuous scale (see color guide), the RRBS tracks indicate the average DNA methylation levels at each monitored CpG site from the quantitative sequencing data.22,28 The RRBS data are shown for four independent Mb cultures and for Mt preparations derived from them, three skin or foreskin fibroblast cultures, five independent LCLs, H1 ESC, skeletal muscle, brain, and leukocytes, as described previously.22 The orange boxes in (A–C) indicate overlapping epigenetic features and other boxes indicate other noteworthy epigenetic features. All tracks are aligned. The chr9:139,386,830–139,447,728 (hg19) region is shown. Mt exhibited histone modifications patterns similar to Mb in the regions shown in this figure and in those of subsequent figures (not shown). See Figure S1 for a close-up of the myogenic DMR and its features that cannot be seen well in this whole-gene view.
Further evidence for muscle-lineage specific, cis-acting transcription regulation from this subregion came from MyoD ChIP-seq profiles of C2C12 mouse Mb and Mt.29 Two sequences at either end of the enhancer-like DMR that we identified in Mb and Mt were orthologous to C2C12 sequences that were bound strongly in Mb and Mt by the myogenic transcription factor MyoD (Fig. S1C). Moreover, the DMR overlapped several peaks of DNaseI hypersensitivity (DHS) that were present in Mb and Mt but not in nonmuscle cell cultures (Fig. 1C, orange box; Fig. S1D). Importantly, the central DHS seen in Mb and Mt at this enhancer-like region (as determined by histone modifications; ENCODE, Broad Institute) was retained in adult skeletal muscle (Fig. S1D, orange boxes). In addition, the intergenic region upstream of NOTCH1 had a myogenic hypomethylated DMR observed at both the tissue and progenitor cell stages (Fig. S2). These results suggest the biological importance of NOTCH1 protein within the muscle fiber itself (probably only transiently or at low levels) in addition to the previously described roles of NOTCH1 in satellite cells and Mb.9,12 Skeletal muscle-associated epigenetic features should reflect the epigenetics of the nuclei within the syncytial myofibers (Fig. 2A) because satellite cells constitute only a few percent of the nuclei in adult muscle and because Mb and Mt are yet scarcer in normal mature muscle.13 Small DMRs with significant myogenic hypermethylation in the vicinity of NOTCH1 were also seen in Mb and Mt although not in muscle (Fig. 1A, pink box; Fig. S2A). Importantly, most of these myogenic hypermethylated DMRs overlapped sites displaying hypomethylation in lymphoblastoid cell lines (LCLs) compared with most non-LCL cell types (Fig. 1D, pink box). This suggests that the Mb and Mt hypermethylation is counteracting potential regulatory elements activated in LCLs.
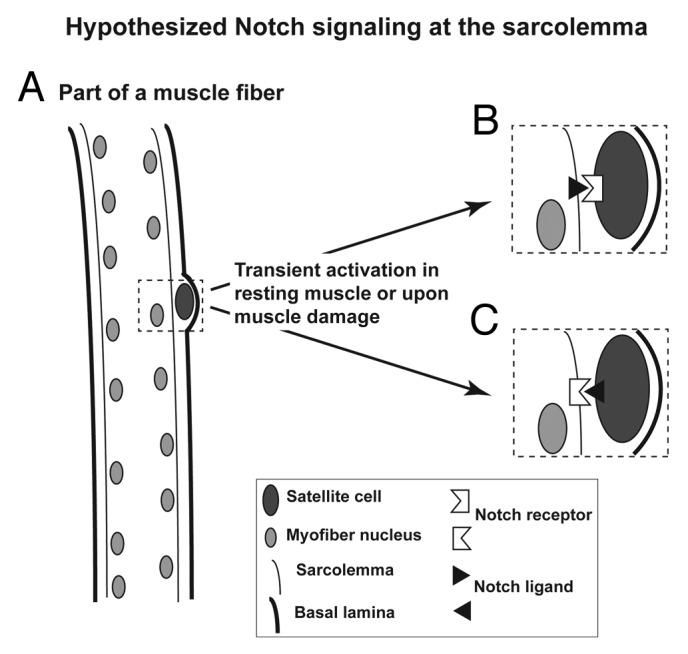
Figure 2. Cartoon illustrating the muscle satellite cell niche and the hypothesized Notch signaling involving the sarcolemma. (A) Illustration of how satellite cells are wedged between the basal lamina and the sarcolemma (plasma membrane of the myofiber). (B) Diagram of Notch signaling from a myofiber to an associated satellite cell. (C) Diagram of Notch signaling from the satellite cell to the myofiber.
Using the Cufflinks tool,30 we analyzed pre-existing RNA-seq profiles of Mb and five nonmyogenic cell cultures (ENCODE/California Institute of Technology, http://genome.ucsc.edu) and our newly generated profiles of Mb and Mt RNA-seq (Table S1). NOTCH1 canonical transcripts were present in Mb and Mt. Notch signaling inhibits differentiation of Mb to Mt 3 but the Mt used in all these analyses were not in the process of differentiating, but rather already maximally differentiated (4-to-5 d after induction of differentiation of Mb to Mt by serum restriction). NOTCH1 was expressed at higher levels in ESC, umbilical vein endothelial cells (HUVEC) and keratinocytes (NHEK) than in Mb. NOTCH1 RNA was reported to be present at only low steady-state levels in mature skeletal muscle, although exercise is known to induce the formation of more of this RNA.18 Many tissues, such as heart and lung, have much higher expression.31
JAG2, like NOTCH1, is implicated in myogenesis.32 We found MbMt hypomethylation and muscle hypomethylation in a JAG2 intron overlapping myogenic enhancer-type chromatin, DHS peaks, and C2C12-deduced MyoD binding29 (Fig. 3). In addition, DNA hypomethylation in skeletal muscle, Mb, and Mt was found in or near the following genes encoding a Notch ligand, receptor, or receptor-modifying enzyme: DLL1 (downstream, Fig. 4A), NOTCH2 (in the 3′ terminus of one of the gene isoforms, Fig. S3A), and the NOTCH1 glycosyltransferase-encoding LFNG8 (upstream of the promoter region, Fig. S3B). Some small intergenic regions with DLL1 and LFNG hypomethylation overlapped myogenic enhancer-like chromatin (Fig. 4B; Fig. S3B, orange boxes in chromatin state tracks). These genes were all appreciably expressed in Mb and Mt samples (Fig. 4E; Fig. S3; Table S1) and skeletal muscle.31NOTCH3, which had only one muscle-hypomethylated DM site in its last exon detectable by RRBS (data not shown), and NOTCH2 were expressed at moderately high levels in Mb and Mt (Table S1). Most of the MbMt hypomethylation in or near the Notch signaling genes did not overlap CpG islands.

Figure 3. Myogenesis-associated hypomethylation and chromatin epigenetic marks in JAG2 intron 5. (A) Myogenic hypomethylated CpG sites (green, 2 Mb -and-Mt sites and 10 muscle sites) and the absence of hypermethylated sites (red) in JAG2 intron 5. (B) The predicted type of chromatin is shown as in Figure 1B, with the addition of designations for polycomb-type H3K27 trimethylation (PcG), which is usually repressive, and poised promoter (poised prom), which usually contains both H3K27me3 and H3K4me3.57 (C) MyoD binding from C2C12 ChIP-seq29 and identification of orthologous human sequences. The relative binding strength is indicated. Sites shown in blue in this and subsequent figures overlapped variants of the MyoD consensus sequence (CAGCTG, V$MYOD_01, V$MYOD_Q6, or E47 sites from http://genome.ucsc.edu, Conserved TFBS). (D) DNase-seq. (E) RNA-seq data for the minus-strand (http://genome.ucsc.edu; vertical viewing range, 1-to-200). In this region, no specific signal was seen for the plus-strand nor for <200 nt RNA-seq (http://genome.ucsc.edu). (F) Examples of RRBS data tracks as in Figure 1. Orange boxes, overlapping epigenetic features in panels a, b, and c. The chr14:105,601,999–105,638,022 region is shown.

Figure 4. Myogenesis-associated hypomethylation from 16 to 230 kb downstream of DLL1. (A) Significantly hypomethylated (green) or hypermethylated (red) sites downstream of DLL1. A long intergenic non-coding RNA (linc RNA) of 0.98 kb, ENST00000610240, which is based on expressed sequence tags (ESTs) found in many tissues, including skeletal muscle (http://www.ensembl.org), overlaps the central cluster of MbMt- and muscle-hypomethylated sites (tall orange box in center) and might have long-distance enhancer function as demonstrated for some other lncRNAs.59 (B) and (C), predicted chromatin states and C2C12-deduced MyoD-binding sites as in Figure 3. Binding sites with signals >40 are indicated by a lollipop. (D) DNase-seq. (E) RNA-seq for the minus-strand with a vertical viewing range of 1 to 50. (F) An example of RRBS data tracks (Mb7) to indicate the coverage of the RRBS data. Thick- and thin-lined orange boxes, the positions of myogenic hypomethylated sites that overlapped enhancer-type chromatin and DHS in Mb and Mt and those that overlapped only a DHS seen in Mb and Mt, respectively. The region shown is chr6:170,357,260–170,608,718.
High levels of 5hmC at sites in NOTCH1 and JAG2 and near DLL1 in specific tissues
RRBS can detect both 5hmC and 5mC as “methylated.”23 Because of this and our desire to study more skeletal muscle samples than were profiled by RRBS (two biological replicates), we quantified 5mC and 5hmC at representative CCGG sites within myogenic DMRs. This was done with an enzyme assay that involves T4 phage β-glucosyltransferase, restriction endonucleases, and quantitative real-time PCR.22 We compared a variety of samples from skeletal muscle and myogenic progenitor cells, including multiple samples of cardiac muscle and brain. Brain is known to be much more enriched in 5hmC than most of the other studied human samples.33,34 Moreover, in NOTCH1 intron 2 at the subregion containing the myogenic DMR, brain tissue was one of the few other samples that shared very low methylation levels (Fig. 1D, blue box), so that it was of much interest to quantify methylation and hydroxymethylation in that subregion in multiple samples of brain.
Surprisingly, we found high levels of 5hmC in the central CCGG in NOTCH1‘s myogenic DMR in intron 2 in skeletal muscle (38% ± 12%, SD) and cerebellum (29% ± 13%), and extraordinarily high levels in heart (81% ± 3.8%), while only low levels were seen in leukocytes (4.2% ± 5.4%; Fig. 5A and B). The substantial levels of 5hmC in skeletal muscle, brain, and heart samples were often accompanied by much less 5mC (Fig. 5A). We also quantified 5hmC and 5mC at a CpG site in the myogenic DMR located 97 kb downstream of DLL1 (Fig. 4A and F) and at an exon 32 site in NOTCH1 (Fig. 1D) that was highly methylated in almost all 15 tissues examined by RRBS (average methylation in nonmuscle tissues, ~79%; average methylation in skeletal muscle, ~85%; >90% methylation in all examined cell cultures including Mb and Mt). We found that both sites were enriched in 5hmC in skeletal muscle, heart, and brain (Fig. 5B). Leukocytes again had little or no 5hmC at these sites. In contrast to the tested CCGG sites associated with NOTCH1 or DLL1, which were rich in 5hmC in skeletal muscle, heart, and brain, the assayed myogenic DMR site at intron 5 of JAG2 had an average of only 0.4% 5hmC in three skeletal muscle samples (Fig. 5B, arrow) although brain and heart had high levels of 5hmC at this site. Therefore, the assayed sites in myogenic intronic DMRs of JAG2 and NOTCH1 differed strongly in their 5hmC content in skeletal muscle tissue despite sharing epigenetic features in myogenic progenitor cells (Figs. 1, 3, and 5).

Figure 5. Quantification of 5hmC and 5mC at assayed CCGG sites in myogenic hypomethylated regions by enzymatic assay. (A) The percentages of 5hmC and 5mC for the assayed site in intron 2 of NOTCH1 in each tested sample and (B) and (C), average percentages of 5hmC and 5mC for each group of samples at the indicated subregions, determined as described in Methods. DMR, refers to a region of significant differential methylation between the set of Mb and Mt vs. nonmuscle cell cultures or skeletal muscle tissue vs. nonmuscle tissue. For the JAG2 site, the light green arrow in panel b denotes the lack of appreciable 5hmC at the assayed site in skeletal muscle in contrast to its abundance in heart and cerebellum; note the high levels of 5hmC for all three of these tissues at NOTCH1 intron 2 and exon32 sites and the DLL1 sites. The exact sites studied are given in Table S2 and their relative positions are illustrated in Figures 1D, 3F, 4F and Figure S3A.
We also examined three other CpG sites in the following locations: in exon 20 of NOTCH1 (by RRBS, the site was >90% methylated in most examined cell types and tissues), in a subregion 56 kb downstream of DLL1 (by RRBS, average methylation in skeletal muscle and in myogenic progenitor cells, ~76 and ~93%, respectively; in non-muscle tissues, ~53%, and in most non-muscle cell cultures, >90%), and in a myogenic hypomethylated DMR in exon 24 of NOTCH2. The enzymatic assay indicated that the DLL1 56-kb downstream CpG site had high levels of 5hmC in skeletal muscle, heart, and brain. The NOTCH2 site and the NOTCH1 exon 20 site had low levels of 5hmC in skeletal and cardiac muscle but more in brain (Fig. 5B). Our comparison of three distant subregions in NOTCH1 indicates that 5hmC can be present throughout this moderately large gene but with an inhomogeneous distribution, as was seen at NOTCH1 in neural progenitor cells obtained by in vitro differentiation of ESC.25 Most of the examined 5hmC-containing sites in skeletal muscle, heart, and brain had much more 5hmC than 5mC. This is especially noteworthy because 5mC levels are usually higher than 5hmC levels at a given CpG position that is positive for 5hmC (unpublished data).35,36 At all tested CCGG sites, there were negligible amounts of 5hmC in most Mb samples. Although the levels were low, a joint analysis of the data using a generalized linear model indicated that significantly more 5hmC was found in Mt than in Mb at these sites (P = 0.004). Therefore, we propose that at the assayed NOTCH1 and DLL1 sites, hydroxymethylation begins at the Mt stage and increases later.
Discussion
Much attention has been focused lately on Notch signaling, including its participation in the regeneration of injured skeletal muscle, its contribution to brain function, and its roles in cardiac disease and other diseases.2,19,37-39 Our study provides the first evidence that the multilayered control of Notch signaling in the skeletal and cardiac muscle lineages may include differential cytosine methylation and hydroxymethylation within or near Notch signaling genes. It also confirms the enrichment of 5hmC within the body of Notch signaling genes in brain.26 We propose that differential DNA methylation and hydroxymethylation help fine-tune expression of these genes in Mt, myofibers, cardiac muscle, and brain as well as in muscle satellite cells and Mb.13 The unusually high ratios of 5hmC to 5mC that we found at tested intragenic and intergenic CpG sites of various Notch signaling genes in skeletal muscle tissue, brain, and heart, but not in Mb or Mt, are consistent with the proposed tissue- and stage-specific role for 5hmC in regulating transcription of these genes.
The differential DNA methylation associated with NOTCH1, NOTCH2, and JAG2 in Mb and Mt vs. nonmuscle cells was observed mostly in the gene body rather than the promoter or upstream region. This is in accord with the genome-wide trends in the distribution of myogenic hypomethylation or hypermethylation among gene subregions that we previously described22 and with the general finding that hypomethylated intragenic DMRs are often related to cis-acting gene regulatory elements.23,24,27,40 It is likely that the MbMt DNA hypomethylation in introns of NOTCH1 and JAG2 contributes to the activity or stabilization of previously undescribed myogenic enhancers because each of these myogenic DMRs overlapped a chromatin segment that had histone modifications characteristic of active enhancers in Mb and Mt but not in the studied nonmuscle cell types. This finding and the myogenic DHS peaks suggest that despite the modest levels of expression of these genes in Mb and Mt, their transcription is regulated in a cell type-specific manner. Similarly, we found enhancer-like chromatin located in cell type-specific intragenic positions of NOTCH1 and JAG2 even in different cell types expressing these genes at similar levels (e.g., Mb and LCL cultures, Fig. 1; Table S1A). We hypothesize that the control of Notch signaling genes relies on using differential DNA methylation and, in certain tissues, cytosine hydroxymethylation, as one of the mechanisms to exactly adjust their RNA levels. Such fine-tuning could be necessary because Notch signaling guides diverse cell-fate decisions.2
Mature skeletal muscle relies on NOTCH1 for repair of injury12,41,42,13 but NOTCH1 protein was previously not detected in myofibers of murine skeletal muscle explants although it was observed in the explants’ satellite cells and Mb.43 Therefore, it was surprising that RRBS profiles revealed a cluster of skeletal muscle-hypomethylated DM sites at the same intronic subregion of NOTCH1 (and JAG2) in which hypomethylation was seen in Mb and Mt. Enzymatic assay of selected CCGG sites in NOTCH1 suggests that, after the Mt stage, 5hmC replaced much of the unmethylated C at a DMR site and much of the methylated C at one of two tested non-DMR sites (Fig. 5). That similar results were obtained for two intergenic regions of DLL1, a Notch ligand, is consistent with a role for DNA epigenetic changes in the regulation of expression of Notch signaling genes in skeletal muscle as well as in myogenic progenitor cells.
Our finding of unusually high levels of 5hmC, >75% of total C, in all three heart samples at the tested NOTCH1 DMR site could indicate that 5hmC at this site is especially important in postnatal cardiac muscle. Control of expression of this Notch receptor gene might be related to its involvement in the repair of injury to adult heart by increasing myocyte survival, decreasing myocyte hypertrophy, controlling cardiogenic differentiation, and inhibiting fibrosis.44 In mature brain, which also had high levels of 5hmC at this NOTCH1 site, NOTCH1 has been implicated in the control of cell fate decisions in adult stem and progenitor cells.45
Our previous studies of the TBX1, PAX3, and HOXB5 genes, which encode differentiation-determining transcription factors, revealed much 5hmC at the examined sites in skeletal muscle but little or none in brain and heart.22,46 This is a very different tissue-specificity than was seen in the present study for the four above-mentioned DLL1-downstream and NOTCH1 sites, where 5hmC levels were high in brain, heart, and skeletal muscle, and for the tested JAG2 site, where 5hmC levels were high in brain and heart but not in skeletal muscle (Fig. 5). Such differences indicate the high tissue- and gene-selectivity of genomic hydroxymethylation even within genes that encode proteins in the same signaling pathway.
For brain, partial loss of 5hmC throughout the genome is implicated in inherited diseases affecting cognitive function.47,48 Correspondingly, mammalian brain has a total of about 3–10 times more genomic 5hmC than other tissues, including skeletal muscle and heart.22,33 This makes our finding of higher levels of 5hmC in skeletal muscle than in cerebellum at the NOTCH1 intron 2 and exon 32 sites, the DLL1 97-kb-downstream site, and the previously examined sites at TBX1, PAX3, and HOXB5 especially noteworthy and suggests a transcription regulatory role for 5hmC that is separate from that of 5mC or unmodified C residues in DNA.49
One transcription regulatory role of tissue-specific 5hmC at certain intragenic or intergenic regions in specific tissues might be to serve as a long-lived intermediate in DNA demethylation50 creating DNA regions poised for upregulatory demethylation, which might increase expression beyond a low basal level. Hydroxymethylation of C residues can be a particularly dynamic DNA modification50 that could help explain the dynamic control of Notch gene expression in fiber type determination in skeletal muscle51 and memory acquisition in brain.38 Other possible roles for some of the tissue-specific 5hmC and hypomethylation at Notch signaling genes include contributing to biologically advantageous cell heterogeneity9 and helping to determine the relative amounts of alternative RNA isoforms that are produced (Fig. S3A, NOTCH2 alternative 3′ transcription termination sites near a hypomethylated MbMt DMR with small amounts of 5hmC in muscle).52
Lastly, our study of the epigenetics of Notch signaling genes leads us to new insights into the role of Notch signaling in the muscle lineage that were not obtained in expression studies. In a previous study, immunohistochemical analysis did not reveal the presence of Notch1 protein in myofibers themselves.43 This might have been due to insufficiently sensitive detection and a short half-life for Notch receptors.14 As described above, our epigenetic findings suggest that Notch signaling plays a role in the myofibers of muscle tissue (Fig. 2) as well as the known role in muscle progenitor cells and satellite cells.3-6 Although we did not analyze satellite cells, which are difficult to obtain from human tissue, previous studies indicate the importance of Notch signaling for muscle satellite cell maintenance, repopulation, differentiation, and niche localization.9,12,20,43,53 Our study suggests a model that helps bridge the gap between the intercellular nature of canonical Notch signaling and the large distances between satellite cells in mature, resting muscle. We propose that small amounts of Notch ligand are made within the myofiber, localize to the sarcolemma (myofiber cell membrane), and interact with a Notch receptor at satellite cell membranes43 to initiate Notch signaling within satellite cells (Fig. 2B). Transiently or at low concentrations, Notch receptor proteins may similarly reside at the sarcolemma where they can interact with satellite membrane-bound Notch ligand to activate Notch signaling within the myofiber (Fig. 2C). Such interactions between myofibers and attached satellite cells wedged under the basal lamina can help explain the role of Notch signaling in homing of satellite cells to their niche during murine embryogenesis, in maintaining the stable localization of satellite cells in this niche, and in securing satellite cell quiescence.9,20
The model presented in Figure 2B also elucidates how Notch signaling might occur in satellite cells upon disruption of the sarcolemma in damaged muscle13,54 before these cells have been induced to divide to give adjacent daughter cells (Fig. 2B). Consistent with a role for Notch ligands in the myofiber itself is the finding of DLL1 protein associated with myofibers per se, adjacent to a site of injury.13,55 Importantly, Nedd4/NEDD4, a ubiquitin ligase that helps control Notch receptor stability and muscle atrophy, is localized to the sarcolemma.51 This is consistent with the hypothesis that Notch receptors too are at the sarcolemma, albeit normally at low levels that are determined by epigenetically governed transcriptional regulation as well as by posttranscriptional and posttranslational controls.
Methods
DNA samples, RRBS data, and DHS profiles were obtained as previously described.22,28 DHS (except for tissue profiles) and RRBS databases are available at http://ucsc.genome.edu (DNA methylation by RRBS, Richard Myers, HudsonAlpha Institute for Biotechnology; Open Chromatin, DNaseI HS, Greg Crawford, Duke University). Quality control of Mb (70% confluent) and Mt used for RRBS, DHS, and 5hmC/5mC assays was previously described.22 Mt were harvested 4-to-5 d after initiation of differentiation and had >70% of nuclei in multinucleated cells. Myogenic hypomethylation refers to statistically significant differences between myogenic and nonmyogenic samples. Statistical analysis of myogenic differentially methylated sites (a change in methylation of at least 50% at a significance level of ≤0.01 from RRBS data on 18 types of cell culture or 15 types of tissues) was done as previously described.22 Myogenic DMRs from the same RRBS data sets were detected using our UPQ algorithm as recently described.56 Profiles of histone modifications and strand-specific RNA-seq are also from ENCODE data available at http://ucsc.genome.edu (Chromatin state segmentation by HMM, Bradley Bernstein, Broad Institute57; Strand-specific RNA-seq, long RNA-seq, >200 nt poly(A)+, Tom Gingeras, Cold Spring Harbor). For quantification of RNA-seq data by Cufflinks,58 we used non-strand-specific profiles from ENCODE (http://ucsc.genome.edu ; RNA-seq, Barbara Wold, California Institute of Technology) or our newly generated RNA-seq data (Mb and Mt RNA-seq libraries prepared from poly(A)+ RNA and processed on Illumina Hiseq 2000, 50-bp paired-end reads, Duke Genome Sequencing Core).
Both 5hmC and 5mC were quantified at specific CCGG sites by previously described methods (Epimark Kit, New England Biolabs22) using newly designed primers for real-time PCR given in Table S2. For the studied CCGG sites, three different tissue samples were tested except for NOTCH1 intron 2, where 3-to-5 were used (Fig. 5A), and NOTCH2 and DLL1, where 2-to-3 were used. For cell culture samples, 2-to-5 heterologous samples were tested. The samples used for this analysis were different from those used for RRBS. The ages of most of the tissue donors for these assays are unknown except that they were from adults and one donor (for skeletal muscle H, heart H, and cerebellum H) was a 68-y-old female, comparable in ages to the two RRBS skeletal muscle samples, a 71-y-old man and an 83 y-old female.
Supplementary Material
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
We thank Carl Baribault for help with mapping DM sites and DMRs and Melody Badoo and the Tulane Cancer Center (COBRE grant NIGMS P20GM103518) for help with the Cufflinks analysis of the ENCODE RNA-seq data, and Mary C. Phelan and Sufen Zheng for supplying several skin fibroblast stocks. This research was supported in part by a grant from the National Institutes of Health to ME (NS04885). Research in one of our laboratories (SP) was supported by 4R44GM096723–02 and internal funding for basic research at New England Biolabs.
Glossary
Abbreviations:
- 5hmC
5-hydroxymethylcytosine
- 5mC
5-methylcytosine
- Mb
myoblasts
- Mt
myotubes
- MbMt
the set of Mb plus Mt samples
- RRBS
reduced representation bisulfite sequencing
- DM
differentially methylated
- DMR
differentially methylated region
- RNA-seq
next-generation whole-genome sequencing analysis of cDNA
- ChIP-seq
next-generation whole-genome sequencing of a chromatin immunoprecipitate
- TSS
transcription start site
- DHS
DNaseI hypersensitive site
- LCL
lymphoblastoid cell line
- NHLF
human lung fibroblasts
- ESC
embryonic stem cells
- HMEC
human mammary epithelial cells
- HUVEC
human umbilical vein endothelial cells
- NHEK
normal human epidermal keratinocytes
- skin fib
skin or foreskin fibroblasts
References
- 1.Helbig C, Gentek R, Backer RA, de Souza Y, Derks IA, Eldering E, Wagner K, Jankovic D, Gridley T, Moerland PD, et al. Notch controls the magnitude of T helper cell responses by promoting cellular longevity. Proc Natl Acad Sci U S A. 2012;109:9041–6. doi: 10.1073/pnas.1206044109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Aster JC. In brief: Notch signalling in health and disease. J Pathol. 2014;232:1–3. doi: 10.1002/path.4291. [DOI] [PubMed] [Google Scholar]
- 3.Nofziger D, Miyamoto A, Lyons KM, Weinmaster G. Notch signaling imposes two distinct blocks in the differentiation of C2C12 myoblasts. Development. 1999;126:1689–702. doi: 10.1242/dev.126.8.1689. [DOI] [PubMed] [Google Scholar]
- 4.Dahlqvist C, Blokzijl A, Chapman G, Falk A, Dannaeus K, Ibâñez CF, Lendahl U. Functional Notch signaling is required for BMP4-induced inhibition of myogenic differentiation. Development. 2003;130:6089–99. doi: 10.1242/dev.00834. [DOI] [PubMed] [Google Scholar]
- 5.Tsivitse S. Notch and Wnt signaling, physiological stimuli and postnatal myogenesis. Int J Biol Sci. 2010;6:268–81. doi: 10.7150/ijbs.6.268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hindi SM, Shin J, Ogura Y, Li H, Kumar A. Matrix metalloproteinase-9 inhibition improves proliferation and engraftment of myogenic cells in dystrophic muscle of mdx mice. PLoS One. 2013;8:e72121. doi: 10.1371/journal.pone.0072121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Gude N, Sussman M. Notch signaling and cardiac repair. J Mol Cell Cardiol. 2012;52:1226–32. doi: 10.1016/j.yjmcc.2012.03.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kato TM, Kawaguchi A, Kosodo Y, Niwa H, Matsuzaki F. Lunatic fringe potentiates Notch signaling in the developing brain. Mol Cell Neurosci. 2010;45:12–25. doi: 10.1016/j.mcn.2010.05.004. [DOI] [PubMed] [Google Scholar]
- 9.Brack AS, Rando TA. Tissue-specific stem cells: lessons from the skeletal muscle satellite cell. Cell Stem Cell. 2012;10:504–14. doi: 10.1016/j.stem.2012.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Fan CM, Li L, Rozo ME, Lepper C. Making skeletal muscle from progenitor and stem cells: development versus regeneration. Wiley Interdiscip Rev Dev Biol. 2012;1:315–27. doi: 10.1002/wdev.30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lepper C, Low S, Partridge TA. The satellite cell builds its nest under Notch’s guidance. Cell Stem Cell. 2012;11:443–4. doi: 10.1016/j.stem.2012.09.010. [DOI] [PubMed] [Google Scholar]
- 12.Qin L, Xu J, Wu Z, Zhang Z, Li J, Wang C, Long Q. Notch1-mediated signaling regulates proliferation of porcine satellite cells (PSCs) Cell Signal. 2013;25:561–9. doi: 10.1016/j.cellsig.2012.11.003. [DOI] [PubMed] [Google Scholar]
- 13.Luo D, Renault VM, Rando TA. The regulation of Notch signaling in muscle stem cell activation and postnatal myogenesis. Semin Cell Dev Biol. 2005;16:612–22. doi: 10.1016/j.semcdb.2005.07.002. [DOI] [PubMed] [Google Scholar]
- 14.Beres BJ, George R, Lougher EJ, Barton M, Verrelli BC, McGlade CJ, Rawls JA, Wilson-Rawls J. Numb regulates Notch1, but not Notch3, during myogenesis. Mech Dev. 2011;128:247–57. doi: 10.1016/j.mod.2011.02.002. [DOI] [PubMed] [Google Scholar]
- 15.Bae Y, Yang T, Zeng HC, Campeau PM, Chen Y, Bertin T, Dawson BC, Munivez E, Tao J, Lee BH. miRNA-34c regulates Notch signaling during bone development. Hum Mol Genet. 2012;21:2991–3000. doi: 10.1093/hmg/dds129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Adam MG, Berger C, Feldner A, Yang WJ, Wüstehube-Lausch J, Herberich SE, Pinder M, Gesierich S, Hammes HP, Augustin HG, et al. Synaptojanin-2 binding protein stabilizes the Notch ligands DLL1 and DLL4 and inhibits sprouting angiogenesis. Circ Res. 2013;113:1206–18. doi: 10.1161/CIRCRESAHA.113.301686. [DOI] [PubMed] [Google Scholar]
- 17.Acharyya S, Sharma SM, Cheng AS, Ladner KJ, He W, Kline W, Wang H, Ostrowski MC, Huang TH, Guttridge DC. TNF inhibits Notch-1 in skeletal muscle cells by Ezh2 and DNA methylation mediated repression: implications in duchenne muscular dystrophy. PLoS One. 2010;5:e12479. doi: 10.1371/journal.pone.0012479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Carey KA, Farnfield MM, Tarquinio SD, Cameron-Smith D. Impaired expression of Notch signaling genes in aged human skeletal muscle. J Gerontol A Biol Sci Med Sci. 2007;62:9–17. doi: 10.1093/gerona/62.1.9. [DOI] [PubMed] [Google Scholar]
- 19.Kawaguchi D, Furutachi S, Kawai H, Hozumi K, Gotoh Y. Dll1 maintains quiescence of adult neural stem cells and segregates asymmetrically during mitosis. Nat Commun. 2013;4:1880. doi: 10.1038/ncomms2895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Bröhl D, Vasyutina E, Czajkowski MT, Griger J, Rassek C, Rahn HP, Purfürst B, Wende H, Birchmeier C. Colonization of the satellite cell niche by skeletal muscle progenitor cells depends on Notch signals. Dev Cell. 2012;23:469–81. doi: 10.1016/j.devcel.2012.07.014. [DOI] [PubMed] [Google Scholar]
- 21.Myers RM, Stamatoyannopoulos J, Snyder M, Dunham I, Hardison RC, Bernstein BE, Gingeras TR, Kent WJ, Birney E, Wold B, et al. ENCODE Project Consortium A user’s guide to the encyclopedia of DNA elements (ENCODE) PLoS Biol. 2011;9:e1001046. doi: 10.1371/journal.pbio.1001046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Tsumagari K, Baribault C, Terragni J, Varley KE, Gertz J, Pradhan S, Badoo M, Crain CM, Song L, Crawford GE, et al. Early de novo DNA methylation and prolonged demethylation in the muscle lineage. Epigenetics. 2013;8:317–32. doi: 10.4161/epi.23989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Stroud H, Feng S, Morey Kinney S, Pradhan S, Jacobsen SE. 5-Hydroxymethylcytosine is associated with enhancers and gene bodies in human embryonic stem cells. Genome Biol. 2011;12:R54. doi: 10.1186/gb-2011-12-6-r54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Szulwach KE, Li X, Li Y, Song CX, Han JW, Kim S, Namburi S, Hermetz K, Kim JJ, Rudd MK, et al. Integrating 5-hydroxymethylcytosine into the epigenomic landscape of human embryonic stem cells. PLoS Genet. 2011;7:e1002154. doi: 10.1371/journal.pgen.1002154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kim M, Park YK, Kang TW, Lee SH, Rhee YH, Park JL, Kim HJ, Lee D, Lee D, Kim SY, et al. Dynamic changes in DNA methylation and hydroxymethylation when hES cells undergo differentiation toward a neuronal lineage. Hum Mol Genet. 2014; ;23:657, 67. doi: 10.1093/hmg/ddt453. [DOI] [PubMed] [Google Scholar]
- 26.Jin SG, Wu X, Li AX, Pfeifer GP. Genomic mapping of 5-hydroxymethylcytosine in the human brain. Nucleic Acids Res. 2011;39:5015–24. doi: 10.1093/nar/gkr120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Meissner A, Mikkelsen TS, Gu H, Wernig M, Hanna J, Sivachenko A, Zhang X, Bernstein BE, Nusbaum C, Jaffe DB, et al. Genome-scale DNA methylation maps of pluripotent and differentiated cells. Nature. 2008;454:766–70. doi: 10.1038/nature07107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Varley KE, Gertz J, Bowling KM, Parker SL, Reddy TE, Pauli-Behn F, Cross MK, Williams BA, Stamatoyannopoulos JA, Crawford GE, et al. Dynamic DNA methylation across diverse human cell lines and tissues. Genome Res. 2013;23:555–67. doi: 10.1101/gr.147942.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Cao Y, Yao Z, Sarkar D, Lawrence M, Sanchez GJ, Parker MH, MacQuarrie KL, Davison J, Morgan MT, Ruzzo WL, et al. Genome-wide MyoD binding in skeletal muscle cells: a potential for broad cellular reprogramming. Dev Cell. 2010;18:662–74. doi: 10.1016/j.devcel.2010.02.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Mortazavi A, Williams BA, McCue K, Schaeffer L, Wold B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat Methods. 2008;5:621–8. doi: 10.1038/nmeth.1226. [DOI] [PubMed] [Google Scholar]
- 31.Kupershmidt I, Su QJ, Grewal A, Sundaresh S, Halperin I, Flynn J, Shekar M, Wang H, Park J, Cui W, et al. Ontology-based meta-analysis of global collections of high-throughput public data. PLoS One. 2010;5:e1306. doi: 10.1371/journal.pone.0013066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Luo B, Aster JC, Hasserjian RP, Kuo F, Sklar J. Isolation and functional analysis of a cDNA for human Jagged2, a gene encoding a ligand for the Notch1 receptor. Mol Cell Biol. 1997;17:6057–67. doi: 10.1128/mcb.17.10.6057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Terragni J, Bitinaite J, Zheng Y, Pradhan S. Biochemical characterization of recombinant β-glucosyltransferase and analysis of global 5-hydroxymethylcytosine in unique genomes. Biochemistry. 2012;51:1009–19. doi: 10.1021/bi2014739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kriaucionis S, Heintz N. The nuclear DNA base 5-hydroxymethylcytosine is present in Purkinje neurons and the brain. Science. 2009;324:929–30. doi: 10.1126/science.1169786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Lister R, Mukamel EA, Nery JR, Urich M, Puddifoot CA, Johnson ND, Lucero J, Huang Y, Dwork AJ, Schultz MD, et al. Global epigenomic reconfiguration during mammalian brain development. Science. 2013;341:1237905. doi: 10.1126/science.1237905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Sun Z, Terragni J, Borgaro JG, Liu Y, Yu L, Guan S, Wang H, Sun D, Cheng X, Zhu Z, et al. High-resolution enzymatic mapping of genomic 5-hydroxymethylcytosine in mouse embryonic stem cells. Cell Rep. 2013;3:567–76. doi: 10.1016/j.celrep.2013.01.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Tran T, Andersen R, Sherman SP, Pyle AD. Insights into skeletal muscle development and applications in regenerative medicine. Int Rev Cell Mol Biol. 2013;300:51–83. doi: 10.1016/B978-0-12-405210-9.00002-3. [DOI] [PubMed] [Google Scholar]
- 38.Guo JU, Ma DK, Mo H, Ball MP, Jang MH, Bonaguidi MA, Balazer JA, Eaves HL, Xie B, Ford E, et al. Neuronal activity modifies the DNA methylation landscape in the adult brain. Nat Neurosci. 2011;14:1345–51. doi: 10.1038/nn.2900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Penton AL, Leonard LD, Spinner NB. Notch signaling in human development and disease. Semin Cell Dev Biol. 2012;23:450–7. doi: 10.1016/j.semcdb.2012.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kozlenkov A, Roussos P, Timashpolsky A, Barbu M, Rudchenko S, Bibikova M, Klotzle B, Byne W, Lyddon R, Di Narzo AF, et al. Differences in DNA methylation between human neuronal and glial cells are concentrated in enhancers and non-CpG sites. Nucleic Acids Res. 2014;42:109–27. doi: 10.1093/nar/gkt838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Akiho M, Nakashima H, Sakata M, Yamasa Y, Yamaguchi A, Sakuma K. Expression profile of Notch-1 in mechanically overloaded plantaris muscle of mice. Life Sci. 2010;86:59–65. doi: 10.1016/j.lfs.2009.11.011. [DOI] [PubMed] [Google Scholar]
- 42.Tsivitse SK, Peters MG, Stoy AL, Mundy JA, Bowen RS. The effect of downhill running on Notch signaling in regenerating skeletal muscle. Eur J Appl Physiol. 2009;106:759–67. doi: 10.1007/s00421-009-1077-7. [DOI] [PubMed] [Google Scholar]
- 43.Conboy IM, Rando TA. The regulation of Notch signaling controls satellite cell activation and cell fate determination in postnatal myogenesis. Dev Cell. 2002;3:397–409. doi: 10.1016/S1534-5807(02)00254-X. [DOI] [PubMed] [Google Scholar]
- 44.Zhou XL, Liu JC. Role of Notch signaling in the mammalian heart. Braz J Med Biol Res. 2014;47:1–10. doi: 10.1590/1414-431X20133177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Pierfelice T, Alberi L, Gaiano N. Notch in the vertebrate nervous system: an old dog with new tricks. Neuron. 2011;69:840–55. doi: 10.1016/j.neuron.2011.02.031. [DOI] [PubMed] [Google Scholar]
- 46.Tsumagari K, Baribault C, Terragni J, Chandra S, Renshaw C, Sun Z, Song L, Crawford GE, Pradhan S, Lacey M, et al. DNA methylation and differentiation: HOX genes in muscle cells. Epigenetics Chromatin. 2013;6:25. doi: 10.1186/1756-8935-6-25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Wang F, Yang Y, Lin X, Wang JQ, Wu YS, Xie W, Wang D, Zhu S, Liao YQ, Sun Q, et al. Genome-wide loss of 5-hmC is a novel epigenetic feature of Huntington’s disease. Hum Mol Genet. 2013;22:3641–53. doi: 10.1093/hmg/ddt214. [DOI] [PubMed] [Google Scholar]
- 48.Yao B, Lin L, Street RC, Zalewski ZA, Galloway JN, Wu H, Nelson DL, Jin P. Genome-wide alteration of 5-hydroxymethylcytosine in a mouse model of fragile X-associated tremor/ataxia syndrome. Hum Mol Genet. 2014;23:1095–107. doi: 10.1093/hmg/ddt504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Spruijt CG, Gnerlich F, Smits AH, Pfaffeneder T, Jansen PW, Bauer C, Münzel M, Wagner M, Müller M, Khan F, et al. Dynamic readers for 5-(hydroxy)methylcytosine and its oxidized derivatives. Cell. 2013;152:1146–59. doi: 10.1016/j.cell.2013.02.004. [DOI] [PubMed] [Google Scholar]
- 50.Guo JU, Su Y, Zhong C, Ming GL, Song H. Hydroxylation of 5-methylcytosine by TET1 promotes active DNA demethylation in the adult brain. Cell. 2011;145:423–34. doi: 10.1016/j.cell.2011.03.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Koncarevic A, Jackman RW, Kandarian SC. The ubiquitin-protein ligase Nedd4 targets Notch1 in skeletal muscle and distinguishes the subset of atrophies caused by reduced muscle tension. FASEB J. 2007;21:427–37. doi: 10.1096/fj.06-6665com. [DOI] [PubMed] [Google Scholar]
- 52.Ehrlich M, Lacey M. DNA methylation and differentiation: silencing, upregulation and modulation of gene expression. Epigenomics. 2013;5:553–68. doi: 10.2217/epi.13.43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Wen Y, Bi P, Liu W, Asakura A, Keller C, Kuang S. Constitutive Notch activation upregulates Pax7 and promotes the self-renewal of skeletal muscle satellite cells. Mol Cell Biol. 2012;32:2300–11. doi: 10.1128/MCB.06753-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Chargé SB, Rudnicki MA. Cellular and molecular regulation of muscle regeneration. Physiol Rev. 2004;84:209–38. doi: 10.1152/physrev.00019.2003. [DOI] [PubMed] [Google Scholar]
- 55.Conboy IM, Conboy MJ, Smythe GM, Rando TA. Notch-mediated restoration of regenerative potential to aged muscle. Science. 2003;302:1575–7. doi: 10.1126/science.1087573. [DOI] [PubMed] [Google Scholar]
- 56.Lacey MR, Baribault C, Ehrlich M. Modeling, simulation and analysis of methylation profiles from reduced representation bisulfite sequencing experiments. Stat Appl Genet Mol Biol. 2013;12:723–42. doi: 10.1515/sagmb-2013-0027. [DOI] [PubMed] [Google Scholar]
- 57.Ernst J, Kheradpour P, Mikkelsen TS, Shoresh N, Ward LD, Epstein CB, Zhang X, Wang L, Issner R, Coyne M, et al. Mapping and analysis of chromatin state dynamics in nine human cell types. Nature. 2011;473:43–9. doi: 10.1038/nature09906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Trapnell C, Roberts A, Goff L, Pertea G, Kim D, Kelley DR, Pimentel H, Salzberg SL, Rinn JL, Pachter L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat Protoc. 2012;7:562–78. doi: 10.1038/nprot.2012.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Ørom UA, Shiekhattar R. Long noncoding RNAs usher in a new era in the biology of enhancers. Cell. 2013;154:1190–3. doi: 10.1016/j.cell.2013.08.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.