

Abstract
We aim to investigate the effects of curcumin on preventing diabetes-induced vascular inflammation in association with its actions on Txnip, ICAM-1, and NOX2 enzyme expressions. Male Wistar rats were divided into four groups: control (CON), diabetic (DM; streptozotocin (STZ), i.v. 55 mg/kg BW), control-treated with curcumin (CONCUR; 300 mg/kg BW), and diabetes treated with curcumin (DMCUR; 300 mg/kg BW). 12th week after STZ injection, iris blood perfusion, leukocyte adhesion, Txnip, p47phox, and malondialdehyde (MDA) levels were determined by using laser Doppler, intravital fluorescent confocal microscopy, Western Blot analysis, and TBAR assay, respectively. The iris blood perfusion of DM and DMCUR was decreased significantly compared to CON and CONCUR (P < 0.001). Plasma glucose and HbA1c of DM and DMCUR were increased significantly compared to CON and CONCUR (P < 0.001). Leukocyte adhesion, ICAM-1, p47phox expression, and MDA levels in DM were increased significantly compared to CON, CONCUR, and DMCUR (P < 0.05). Txnip expression in DM and DMCUR was significantly higher than CON and CONCUR (P < 0.05). From Pearson's analysis, the correlation between the plasma MDA level and the endothelial functions was significant. It suggested that curcumin could ameliorate diabetic vascular inflammation by decreasing ROS overproduction, reducing leukocyte-endothelium interaction, and inhibiting ICAM-1 and NOX2 expression.
1. Introduction
Diabetes-induced hyperglycemia can disturb the vascular homeostasis that is characterized by decreased vascular blood perfusion, increased vascular permeability, and enhanced vascular inflammation, leading to diabetic vascular complications. Several studies [1–4] reported that hyperglycemia-induced reactive oxygen species (ROS) overproduction are continuously produced during metabolic processes through the pathways including NADPH oxidase, xanthine oxidase, and mitochondria respiratory chain. ROS overproduction above the physiological level can overcome the functions of cellular antioxidant system leading to oxidative stress.
Hyperglycemia could induce high-level thioredoxin-interacting protein (Txnip) expression [5, 6] by stimulating carbohydrate response element binding protein and forkhead box01 transcription factor [7, 8]. Txnip is an endogenous inhibitor of thioredoxin (TRX) system which is one of the important cellular antioxidant systems. The Txnip-TRX interaction can produce multiple processes consequently including increasing intracellular ROS [9–11], activating stress signaling pathway of apoptosis signal-regulating kinase 1 (ASK-1), increasing intercellular adhesion molecule 1 (ICAM-1) expression, and finally resulting in endothelial dysfunction [10, 12–14].
Through the other pathway, hyperglycemia-induced ROS can stimulate NF-κB activation which then causes the increase in vascular adhesion molecule expression. Therefore, it implies that both pathways of Txnip-TRX interaction and NF-κB activation refer to promoting leukocyte-endothelium interaction and play a central role in diabetic vascular inflammation [12, 15, 16]. Moreover, the severity of diabetic vascular inflammation reproduced after the plugged leukocyte in microcirculation via the process of phagocytic NADPH oxidase (NOX2 enzyme) mediated superoxide radical formation [17–19].
Recently, the use of natural plant products has gained more attention among scientists in order to prevent diabetic vascular complications. Curcumin is one of the polyphenol compounds which exhibit antioxidant, anti-inflammatory, antitumorigenic, and antimicrobial properties. With its antioxidant activity, curcumin has direct effect to scavenge free radicals by redox activity [20–22] and indirect effect to enhance cellular antioxidant system by activating Nrf2 transcription factor, a cytoprotective gene [23–26]. With its anti-inflammatory activity, curcumin can attenuate proinflammatory cytokine levels such as TNF-α, IL-1β, and IL-6 by interfering with the activation of NF-κB, c-Jun, and JNK [27–30]. Several studies [31, 32] found that curcumin can reduce adhesion molecule by inhibiting proinflammatory cytokine secretion in in vitro model. Also, our previous study found that curcumin can attenuate leukocyte-endothelium interaction and ROS overproduction represented by MDA levels in STZ-induced diabetic rat model [33, 34].
Based on these reviews, we hypothesized that the curcumin supplementation which is able to reduce leukocytes adhesion may be achieved by its antioxidative effects on Txnip and ICAM-1 expression and also the inhibition of phagocytic/leukocyte action of NOX2-generated O2 −. Therefore, the present study aims to investigate the mechanism of curcumin on diabetes-induced vascular complications related to these molecular biomarkers including Txnip, ICAM-1, and NOX2 expressions using STZ-induced diabetic rat model.
2. Materials and Methods
2.1. Treatment of Animals
Male Wistar rats (220–250 g) were obtained from National Laboratory Animal Center, Salaya Campus, Mahidol University, Thailand. All rats were randomly divided into diabetic and nondiabetic groups. The diabetic group was induced by streptozotocin (STZ, Sigma Co., USA) at dose 55 mg/kg BW, i.v. [33, 34]. The diabetic condition was defined as a glucose concentration more than 200 mg/dL [35] and verified at 48 hours after STZ injection. The diabetic rats were divided into two groups: diabetes treated with corn oil (DM; n = 6) and diabetes treated with curcumin (DMCUR, n = 6, 300 mg/kg BW, Cayman, USA). In the control groups, they will receive the same volume of citrate buffer. They were divided into two groups: control treated with corn oil (CON, n = 6) and control treated with curcumin (CONCUR, n = 6). The dairy gavage feeding of curcumin dissolved in corn oil was started on 10th day after the STZ injection.
2.2. Experiment of Animals
On the 12th week after STZ injection, the rat was anesthetized with pentobarbital sodium (60 mg/kg BW i.p.), the rats were kept warm at 37°C using warming pad, and a tracheotomy was performed. A jugular vein and carotid artery were cannulated with polyethylene tube for injection of fluorescence tracers and for recording of systolic and diastolic blood pressure, respectively. The blood pressure was measured by using Statham pressure transducer connected to the Polygraph system (Nihon Koden, Japan). Mean arterial blood pressure was calculated from diastolic pressure + 1/3(systolic-diastolic).
2.3. Iris Blood Perfusion Measurement
The right iris blood perfusion was measured using the Laser Doppler Blood Perfusion Monitoring (Perimed AB, Sweden) with the optic needle probe (0.1 millimeter). The needle probe was fixed perpendicularly about 1 mm above iris. Eight different measurement points of iris around pupil were performed at each time and the mean of iris blood perfusion was determined for each rat.
2.4. Evaluation of Leukocyte Adhesion
The iris blood vessel was used to observe instead of retina because the microcirculation in the iris is quite properly to get the good image of leukocyte-endothelial cell interaction and to assess blood perfusion. Besides, the diabetic iridopathy is commonly used clinically to indicate the progression of the diabetic microvascular complications, particularly, in association with advanced proliferative diabetic retinopathy by using fluorescein angiography of the iris [36].
During the experiment, the real time image of iris blood vessel was recorded by an epi-illumination fluorescence video microscopy system (Optiphot 2, Nikon, Japan) equipped with a 100 W mercury lamp, CCD camera (Hamamatsu C2400, Japan), a video recorder (VC-S5, Sharp, Japan) with a video timer (VTG-33, For-A, Japan), and 20x objective lens (CF Plan Fluor, Nikon, Japan).
Leukocyte adhesion in postcapillary venules of iris was determined by rhodamine-6G (R6G) which can label mitochondria especially in leukocyte (Sigma, USA) at 0.15 mg/kg BW following the method described by Jariyapongskul et al. [37]. The emission wavelength of R6G lies between 530 and 540 nm. rhodamine-6G was injected intravenously for visualizing the leukocyte adhesion. Five observation views were chosen and recorded to further determine leukocyte adhesion [38]. The leukocytes which remained stationary to postcapillary venules for 30 seconds were considered to be leukocyte adhesion [33, 39]. The number of leukocyte adhesions in each view was manually counted and reported by the mean number of cells per field of view.
2.5. Biochemical Parameters
The parameters for metabolic changes were blood glucose and HbA1c. All these were determined at the end of the experiments by collected blood sample from abdominal aorta under anesthesia. Blood glucose and HbA1c were measured using enzymatic method and turbidimetric immunoinhibition method, respectively (Bangkok RIA Laboratory Co., Bangkok, Thailand).
In this study fundus malondialdehyde level (MDA), product of lipid peroxidation, was used as indicator of oxygen free radicals in fundus. Thiobarbituric acid (TBA) assay is a commonly used method to determine malondialdehyde. This method is based on the reaction of TBA and MDA. The lipid material is simply heated with TBA under acidic condition, and the formation of a pink color is measured at or close to 532 nm. The pink color is formed by reaction of one molecule of MDA with two molecules of TBA [37].
2.6. Protein Assay
After collecting blood, the eyeball was collected immediately by enucleating from the orbital cavity. Lens and eye fluid were excised from the eye ball; the eyeball without lens and fluids is called fundus. The fundus was washed in phosphate buffer (0.1 M, pH 7.4). Then, the fundus was chopped with fine scissors and was directly in lysis buffer containing 1X RIPA buffer (250 μL of lysis solution per 25 mg of tissue, Cell Signaling, Beverly, MA), 1X phosphatase inhibitor cocktails (1 : 100, Sigma Co., USA), and 1X protease inhibitor (1 : 100, Sigma Co., USA) for 30 minutes [40]. After that, the chopped fundus was sonicated for 3 times for 10 seconds each and was centrifuged at 15,000 g for 10 minutes to spin down cellular debris. The supernatant was collected as the whole cell lysate to determine protein concentration [41]. After protein quantification, the samples were used to determine MDA level by method of Ohkawa et al. [42] and Txnip, ICAM-1, and p47phox expressions by Western blot analysis [6, 7, 21, 24].
2.7. Western Blot Analysis
The protein expressions of Txnip, ICAM-1, and p47phox were performed with 80 μg of fundus protein extraction per lane by 10% SDS-PAGE gel (Bio-Rad Laboratories), transferred to polyvinylidene difluoride (PVDF) membrane, and blocked with blocking solution (5% BSA (Sigma Co., USA) in TTBS). After blocking, the PVDF membrane was incubated with either anti-mouse Txnip antibody (1 μg/mL, MBL, IL), anti-mouse ICAM-1 antibody (1 : 500, BD Biosciences, CA), anti-mouse p47phox antibody (1 : 200, Santa Cruz, CA), or anti-mouse β-actin antibody (1 : 200, Santa Cruz, CA) and followed by horseradish-peroxidase conjugated goat anti-mouse secondary antibody at 1 : 2000 dilution (Santa Cruz, CA). The PVDF membrane was incubated with chemiluminescence substrate solution (GE, USA) and the bands were detected using ChemiDoc system (Bio-Rad Laboratories) with Quantity One program version 4.6.9. Then, densitometry was performed using Image J analysis software (NIH) and the results of Txnip, ICAM-1, and p47phox expressions were quantified as a ratio to β-actin expression.
2.8. Statistical Analysis
All data were presented as means ± SEM (standard error of mean). For comparison among groups of rats, two-way analysis of variance (two-way ANOVA) was used and followed by unpaired t-test. P < 0.05 was considered statistically significant. The association of each parameter was analyzed by using two-tailed Pearson's correlation analysis with P < 0.05 being considered statistically significant. The data was analyzed using the SPSS program (version 16.0) for windows.
3. Results
3.1. The Effects of Curcumin on Physiological Characteristics
As shown in Table 1, the body weights (BW) of 12-week DM and DMCUR rats were significantly decreased (35.93% and 39.15%) as compared to CON and CONCUR groups (P < 0.001). Mean arterial blood pressure (MAP) of 12-week DM and DMCUR groups was not significantly different as compared to CON and CONCUR groups (Table 1).
Table 1.
Values are means ± SEM.
| Group | BW (g) | MAP (mmHg) |
|---|---|---|
| 12-week CON (n = 6) | 424.00 ± 28.28 | 124.77 ± 9.60 |
| 12-week CONCUR (n = 6) | 480.67 ± 17.26 | 132.58 ± 5.50 |
| 12-week DM (n = 6) | 271.67 ± 13.22** | 113.56 ± 6.98 |
| 12-week DMCUR (n = 6) | 258.00 ± 17.64** | 112.80 ± 4.99 |
**P < 0.001 versus CON.
3.2. The Effects of Curcumin on Biochemical Parameters
Blood glucose levels (BG) of DM group and DMCUR groups were significantly elevated as compared to CON group and CONCUR group (P < 0.001).
Plasma glycosylated hemoglobin levels of DM group and DMCUR group were significantly elevated as compared to CON group and CONCUR group (P < 0.001) (Table 2). The results seem to indicate that the curcumin (300 mg/kg BW) could not inhibit high BG significantly.
Table 2.
Values are means ± SEM.
| Group | BG (mg/dL) | Hemoglobin A1c (%) |
|---|---|---|
| 12-week CON (n = 6) | 172.83 ± 11.08 | 4.00 ± 0.06 |
| 12-week CONCUR (n = 6) | 192.50 ± 13.05 | 4.08 ± 0.05 |
| 12-week DM (n = 6) | 410.33 ± 16.77** | 9.47±0.47∗∗ |
| 12-week DMCUR (n = 6) | 390.33 ± 11.59** | 9.30 ± 0.21** |
**P < 0.001 versus CON.
3.3. The Effects of Curcumin on Hemodynamic Changes
By using laser Doppler flowmetry, the regional iris blood perfusion of each rat was evaluated from eight different points measured around the pupil as described previously. Mean regional iris blood perfusion in each group was summarized in Figure 1. Mean iris blood perfusion (IBP) of DM group was significantly reduced as compared to CON and CONCUR groups (P < 0.001). IBP in DMCUR group showed a trend to increase but not significantly different when compared to DM group.
Figure 1.
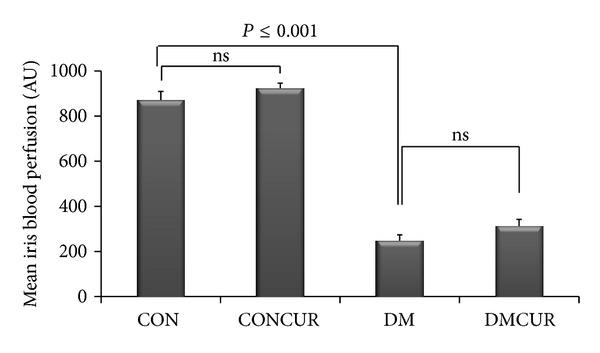
The effects of curcumin on mean regional iris blood perfusion in each group. Control = CON, control treated with curcumin = CONCUR; curcumin 300 mg/kg BW, started at 10 days after STZ injection, diabetic = DM; streptozotocin (STZ), i.v. 55 mg/kg BW, diabetes treated with curcumin = DMCUR; curcumin 300 mg/kg BW. Data are means ± SEM (n = 6 for each group).
3.4. The Effects of Curcumin on Leukocyte-Endothelium Interaction
The number of leukocytes was counted as adherent one that remained stationary for equal to or longer than 30 seconds. The leukocyte adhesion was counted per field of view totally of postcapillary venule (diameter 20–30 μm) as described previously.
In the present video microscopic visualization showed clear image of leukocyte adhering to the endothelium of postcapillary venule in the different monitored views of iris of each rat (Figure 2). Five different monitored views of leukocyte adhesion were manually counted and the mean of leukocyte adhesion per field of views were summarized in the graph as shown in Figure 3.
Figure 2.

Intravital microscopic demonstration of leukocyte adhesion in the postcapillary venule of iris in CON group (a), CONCUR group (b), DM group (c), and DMCUR group (d). White dots represent leukocyte stained by rhodamine-6G i.v. injection. (Scale bar represents 100 μm.)
Figure 3.
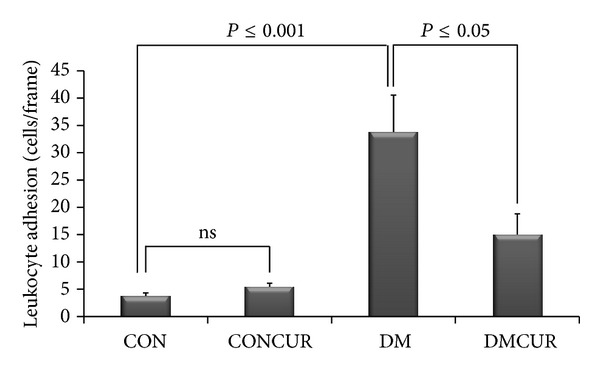
The effects of curcumin on leukocyte-endothelium interaction. Data are means ± SEM (n = 5 for each group). Control = CON, control treated with curcumin = CONCUR; curcumin 300 mg/kg BW, started at 10 days after STZ injection, diabetic = DM; streptozotocin (STZ), i.v. 55 mg/kg BW, diabetes treated with curcumin = DMCUR; curcumin 300 mg/kg BW.
The number of leukocyte adhesions was significantly increased in DM group as compared to CON group and CONCUR group (P < 0.001). Interestingly, the number of leukocyte adhesions of DMCUR group was significantly reduced as compared to DM group (P < 0.05) (Figure 3).
3.5. The Effects of Curcumin on Free Radicals by Products
In this study fundus malondialdehyde level (MDA), product of lipid peroxidation, was used as indicator of oxygen free radicals in fundus. MDA level was significantly elevated in DM group as compared to CON group and CONCUR group (P < 0.001). Interestingly, MDA level of DMCUR group was significantly reduced as compared to DM group (P < 0.05) (Figure 4).
Figure 4.
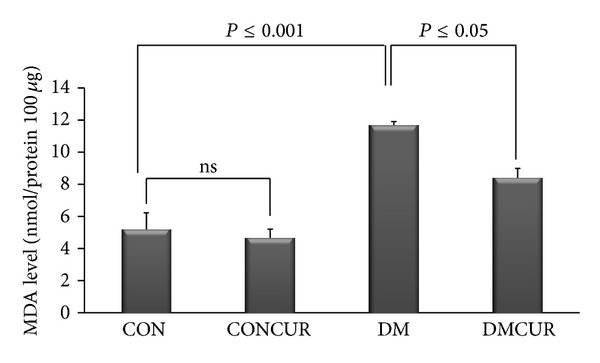
The effects of curcumin on fundus MDA level. Control = CON, control treated with curcumin = CONCUR; curcumin 300 mg/kg BW, started at 10 days after STZ injection, diabetic = DM; streptozotocin (STZ), i.v. 55 mg/kg BW, diabetes treated with curcumin = DMCUR; curcumin 300 mg/kg BW. Data are means ± SEM (n = 5 for each group).
3.6. The Effects of Curcumin on Txnip Expression
The level of Txnip was significantly elevated in DM group and DMCUR group as compared to CON group (P < 0.05). However, the results showed that curcumin could not reduce Txnip expression significantly when compared to the DM group. (P < 0.05) (Figure 5).
Figure 5.
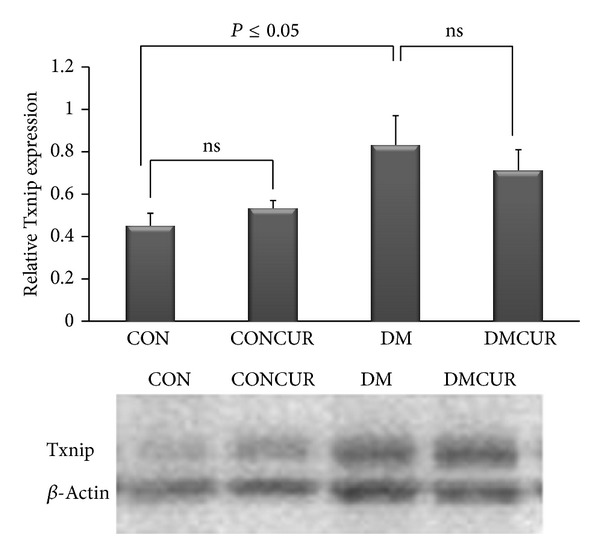
The effect of curcumin on Txnip expression. Control = CON, control treated with curcumin = CONCUR; curcumin 300 mg/kg BW, started at 10 days after STZ injection, diabetic = DM; streptozotocin (STZ), i.v. 55 mg/kg BW, diabetes treated with curcumin = DMCUR; curcumin 300 mg/kg BW. Data are means ± SEM (CON and DMCUR groups; n = 5, CONCUR and DM groups; n = 6).
3.7. The Effects of Curcumin on ICAM-1 Expression
The level of ICAM-1 was significantly elevated in DM group as compared to CON group (P < 0.05). The level of ICAM-1 was significantly decreased in DMCUR compared to DM group (P < 0.05) (Figure 6).
Figure 6.
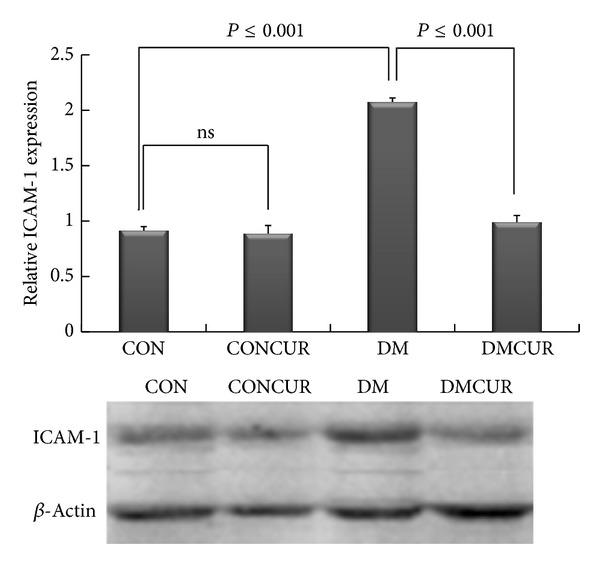
The effect of curcumin on ICAM-1 expression. Control = CON, control treated with curcumin = CONCUR; curcumin 300 mg/kg BW, started at 10 days after STZ injection, diabetic = DM; streptozotocin (STZ), i.v. 55 mg/kg BW, diabetes treated with curcumin = DMCUR; curcumin 300 mg/kg BW. Data are means ± SEM (n = 4).
3.8. The Effects of Curcumin on p47phox Expression
The level of p47phox was significantly elevated in DM group as compared to CON group and CONCUR group (P < 0.05). Interestingly, the level of p47phox of DMCUR group was significantly reduced as compared to DM group, approximately 1.71-fold (P < 0.05) (Figure 7).
Figure 7.
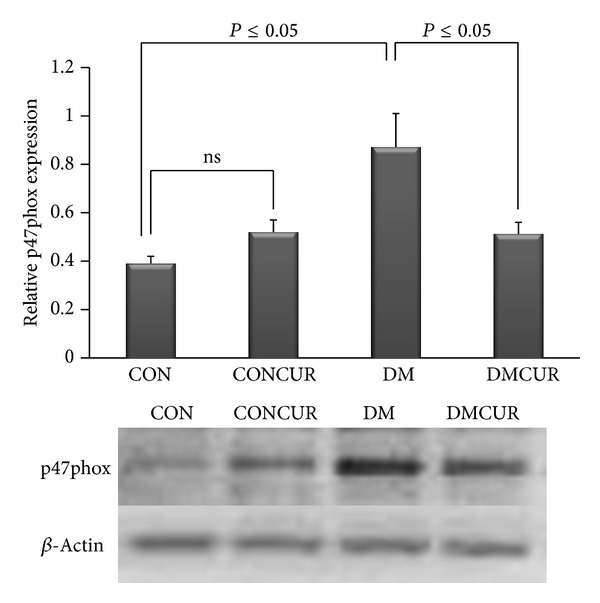
The effect of curcumin on p47phox expression. Control = CON, control treated with curcumin = CONCUR; curcumin 300 mg/kg BW, started at 10 days after STZ injection, diabetic = DM; streptozotocin (STZ), i.v. 55 mg/kg BW, diabetes treated with curcumin = DMCUR; curcumin 300 mg/kg BW. Data are means ± SEM (CON, CONCUR, and DM groups; n = 5, and DMCUR group; n = 4).
From Figure 8, Pearson's correlation between the leukocyte adhesion and MDA level, p47phox and MDA level, and Txnip andMDA level was obtained together with Pearson's correlation and significance: r = 0.705 (P = 0.001), r = 0.729 (P = 0.0001), and r = 0.636 (P = 0.0003), respectively.
Figure 8.
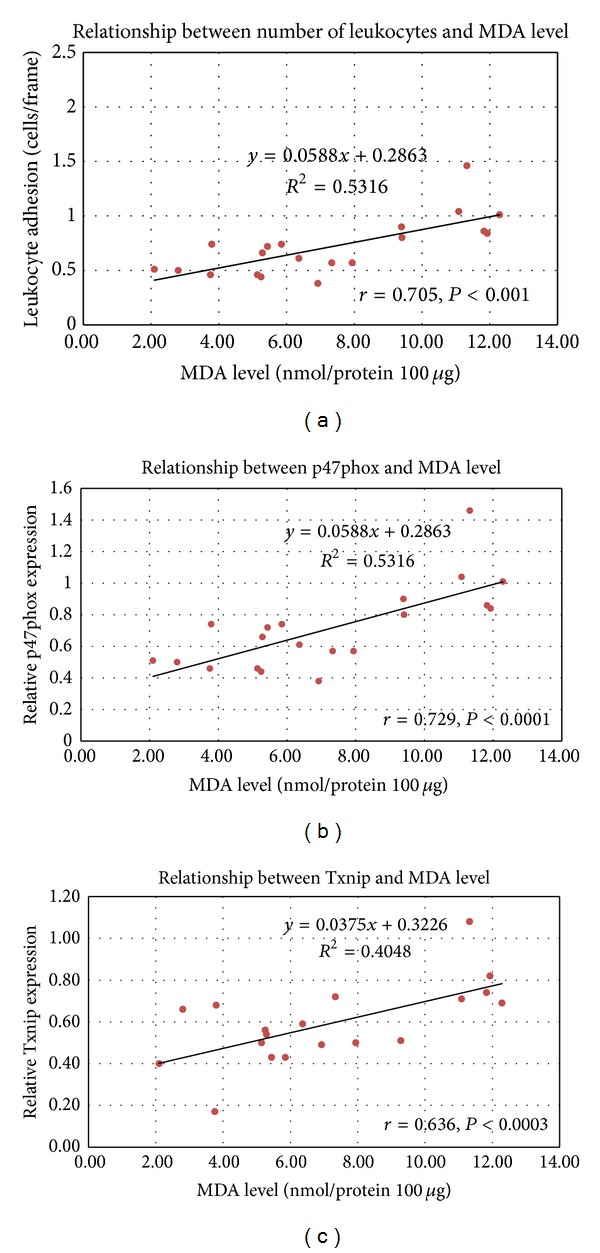
The association between the biochemical and functional data parameters was analyzed by using two-tailed Pearson's correlation analysis with P < 0.05 being considered statistically significant.
4. Discussion
4.1. The Mechanisms of Curcumin on Leukocyte-Endothelium Interaction: Roles of Txnip, ICAM-1, and p47phox Expressions
This experiment demonstrated that curcumin supplementation can attenuate leukocyte-endothelium interaction in association with the decreased ROS overproduction. In addition, curcumin can also suppress NOX2 enzyme expression in vascular inflammatory process in diabetic rats. However, the curcumin supplementation (300 mg/kg BW) could not show the significant inhibition of the hyperglycemia-enhanced Txnip expression when compared to the DM group.
From the previous studies, hyperglycemia was reported as an important factor to induce Txnip expression to increase. Moreover, the hyperglycemia-enhanced Txnip expression has been noted to play a critical role in progression of diabetic vascular complications as well [6, 16, 43, 44]. As shown in our data, Txnip expression in DM group was significantly increased up to 84.44% when compared to the CON group (P = 0.045). The results showed that curcumin could not reduce Txnip expression as that it conforms to the results shown no hypoglycemic effects for curcumin in DMCUR. Since curcumin could not inhibit high BG, therefore, hyperglycemia-enhanced Txnip expression was still existent. Actually, the hypoglycemic effect of curcumin is still controversy. Some studies have reported that curcumin supplementation can ameliorate hyperglycemia in both type 1 [33, 45, 46] and type 2 diabetic models [47] but some did not support this [21, 48, 49].
It is noted that Txnip expression in DMCUR group has tended to slightly reduce down to 14.46% compared to DM group (Figure 5). It may explain that at dose of 300 mg/kg BW curcumin used in this study may not be able to decrease BG and HbA1c; therefore, it was not enough to prevent the alteration of Txnip expression in response to hyperglycemia.
In addition to hyperglycemia-regulated Txnip overexpression, hyperglycemia also can activate many sources to generate ROS such as mitochondria electron transport chain, NADPH oxidase, xanthine oxidase, endothelial NO synthase, and cytochrome p450 [1–4].
The experiments confirmed that hyperglycemia promotes ROS overproduction resulting oxidative stress to damage macromolecules such as lipid which is in apparently the increasing of fundus MDA level in DM group. However, the fundus MDA level in DMCUR group was significantly decreased down to 28% as compared to DM group (P = 0.005). The results of MDA level implied the potential of curcumin as an effective antioxidant.
Both Txnip expression and ROS overproduction could impair endothelial function that causes vascular homeostasis alteration including the abnormal vascular blood flow resistance. In this experiment, blood perfusion in iris was chosen to represent one of the alterations of vascular homeostasis. The results showed that iris blood perfusion (IBP) in DM and DMCUR groups was significantly decreased as compared to CON group, though IBP in DMCUR had tended to increase up to 4.78% as compared to DM group. However, these results did not show the significant difference similar to the previous studies [33, 34]. There are several studies that reported that curcumin could increase cerebral blood flow and improve endothelium-dependent relaxation in mesentery of diabetes rats [50–52]. Therefore, the antioxidant potential of curcumin should be different depending on doses and also the pathological condition of those organs including the severity of cellular antioxidant system [52, 53].
Furthermore, our results have shown that the low dose of curcumin supplementation could significantly decrease ICAM-1 expression down to 52.17% and the number of leukocyte-endothelium interactions was down to 55.68% as compared to the CON group (Figures 3 and 6). Several studies indicated that the enhancing of adhesion molecule expression such as ICAM-1 and VCAM-1 is associated with Txnip-activated ASK-1 stress signaling pathway and ROS-suppressed nitric oxide bioavailability resulting in vascular inflammation [12–14, 54]. Therefore our results may imply that the anti-inflammatory effect of curcumin contributed to prevention of leukocyte adhesion partly by reducing ICAM-1 expression. The effect of curcumin on proinflammatory cytokines such as IL-1 and IL-6 via blockade of Akt, p38MAPK, and NF-κB pathways might be the major processes that explained our observation [28, 29, 33, 55, 56].
From Figure 7, the results of p47phox expressions indicated that the expression of NOX2 enzyme was significantly increased in DM group when compared to the CON group (P = 0.05). The increased p47phox, one of the NOX2 multienzyme complexes in diabetes, was demonstrated by other reports as well [19, 57–59]. It is believed that this phagocytic NADPH oxidase (NOX2 enzyme) is expressed by both phagocytic and vascular endothelial cells. Through this phagocyte NADPH oxidase, ROS could be produced at high levels after leukocyte and endothelial cell activations and then it will cause more diabetic vascular injury [18, 19, 60]. Thus, it seems likely that both phagocyte- and endothelial cell-derived ROS are critically involved in the diabetes-induced vascular pathologic complications.
Our findings indicated that the p47phox expression in DMCUR group was significantly decreased down to 41.38% as compared to DM group. At this point, it may imply that when curcumin could decrease ICAM-1 and NOX2 enzyme expressions, it will inhibit the leukocyte adhesion and prevent vascular inflammation in consequence. Moreover the data from our study also demonstrated the correlation between the plasma MDA level and the endothelial functions significantly (Figure 8). By this significant correlation, it further confirmed the possible protective effects of curcumin against diabetes-induced vascular pathologic complications via the antioxidant mechanism.
5. Conclusions
In summary, our experiments demonstrated that low dose curcumin supplementation (300 mg/kg BW) can ameliorate diabetic vascular inflammation through the decrease in ROS overproduction, reducing leukocyte-endothelium interaction associated with decreased ICAM-1 and NOX2 enzyme expressions in diabetic rat model. Therefore the findings of this study should be a good evidence to suggest that the low dose of curcumin might offer a novel therapeutic strategy for treatment of the diabetic retinopathy associated with vascular inflammation in the future.
Acknowledgments
This study was supported by Ratchadapiseksompotch Fund of the Faculty of Medicine and the 90th Anniversary Research Fund, the Graduate School, Chulalongkorn University, Thailand.
Abbreviations
- DM:
Diabetes mellitus
- IBP:
Iris blood perfusion
- ICAM-1:
Intercellular adhesion molecule 1
- MDA:
Malondialdehyde levels
- MAP:
Mean arterial blood pressure
- NOX2:
NADPH oxidase enzyme
- R6G:
Rhodamine-6G
- ROI:
Region of interest
- ROS:
Reactive oxygen species
- STZ:
Streptozotocin
- O2−:
Superoxide
- Txnip:
Thioredoxin-interacting protein.
Conflict of Interests
The authors have no conflict of interests to report.
References
- 1.Fatehi-Hassanabad Z, Chan CB, Furman BL. Reactive oxygen species and endothelial function in diabetes. European Journal of Pharmacology. 2010;636(1–3):8–17. doi: 10.1016/j.ejphar.2010.03.048. [DOI] [PubMed] [Google Scholar]
- 2.Perez-Matute P, Zulet MA, Martinez JA. Reactive species and diabetes: counteracting oxidative stress to improve health. Current Opinion in Pharmacology. 2009;9(6):771–779. doi: 10.1016/j.coph.2009.08.005. [DOI] [PubMed] [Google Scholar]
- 3.Soldatos G, Cooper ME. Diabetic nephropathy: important pathophysiologic mechanisms. Diabetes Research and Clinical Practice. 2008;82(supplement 1):S75–S79. doi: 10.1016/j.diabres.2008.09.042. [DOI] [PubMed] [Google Scholar]
- 4.Turrens JF. Mitochondrial formation of reactive oxygen species. The Journal of Physiology. 2003;552, part 2:335–344. doi: 10.1113/jphysiol.2003.049478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Schulze PC, Liu H, Choe E, et al. Nitric oxide-dependent suppression of thioredoxin-interacting protein expression enhances thioredoxin activity. Arteriosclerosis, Thrombosis, and Vascular Biology. 2006;26(12):2666–2672. doi: 10.1161/01.ATV.0000248914.21018.f1. [DOI] [PubMed] [Google Scholar]
- 6.Schulze PC, Yoshioka J, Takahashi T, He Z, King GL, Lee RT. Hyperglycemia promotes oxidative stress through inhibition of thioredoxin function by thioredoxin-interacting protein. The Journal of Biological Chemistry. 2004;279(29):30369–30374. doi: 10.1074/jbc.M400549200. [DOI] [PubMed] [Google Scholar]
- 7.Li X, Rong Y, Zhang M, et al. Up-regulation of thioredoxin interacting protein (Txnip) by p38 MAPK and FOXO1 contributes to the impaired thioredoxin activity and increased ROS in glucose-treated endothelial cells. Biochemical and Biophysical Research Communications. 2009;381(4):660–665. doi: 10.1016/j.bbrc.2009.02.132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Cha-Molstad H, Saxena G, Chen J, Shalev A. Glucose-stimulated expression of Txnip is mediated by carbohydrate response element-binding protein, p300, and histone H4 acetylation in pancreatic beta cells. The Journal of Biological Chemistry. 2009;284(25):16898–16905. doi: 10.1074/jbc.M109.010504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Nishiyama A, Matsui M, Iwata S, et al. Identification of thioredoxin-binding protein-2/vitamin D3 up-regulated protein 1 as a negative regulator of thioredoxin function and expression. Journal of Biological Chemistry. 1999;274(31):21645–21650. doi: 10.1074/jbc.274.31.21645. [DOI] [PubMed] [Google Scholar]
- 10.Junn E, Han SH, Im JY, et al. Vitamin D3 up-regulated protein 1 mediates oxidative stress via suppressing the thioredoxin function. Journal of Immunology. 2000;164(12):6287–6295. doi: 10.4049/jimmunol.164.12.6287. [DOI] [PubMed] [Google Scholar]
- 11.Spindel ON, World C, Berk BC. Thioredoxin interacting protein: redox dependent and independent regulatory mechanisms. Antioxidants & Redox Signaling. 2012;16(6):587–596. doi: 10.1089/ars.2011.4137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Harrison CB, Drummond GR, Sobey CG, Selemidis S. Evidence that nitric oxide inhibits vascular inflammation and superoxide production via a p47phox-dependent mechanism in mice. Clinical and Experimental Pharmacology and Physiology. 2010;37(4):429–434. doi: 10.1111/j.1440-1681.2009.05317.x. [DOI] [PubMed] [Google Scholar]
- 13.Yamawaki H, Berk BC. Thioredoxin: a multifunctional antioxidant enzyme in kidney, heart and vessels. Current Opinion in Nephrology & Hypertension. 2005;14(2):149–153. doi: 10.1097/00041552-200503000-00010. [DOI] [PubMed] [Google Scholar]
- 14.Yamawaki H, Pan S, Lee RT, Berk BC. Fluid shear stress inhibits vascular inflammation by decreasing thioredoxin-interacting protein in endothelial cells. Journal of Clinical Investigation. 2005;115(3):733–738. doi: 10.1172/JCI200523001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Perrone L, Devi TS, Hosoya K, Terasaki T, Singh LP. Thioredoxin interacting protein (TXNIP) induces inflammation through chromatin modification in retinal capillary endothelial cells under diabetic conditions. Journal of Cellular Physiology. 2009;221(1):262–272. doi: 10.1002/jcp.21852. [DOI] [PubMed] [Google Scholar]
- 16.Perrone L, Devi TS, Hosoya KI, Terasaki T, Singh LP. Inhibition of TXNIP expression in vivo blocks early pathologies of diabetic retinopathy. Cell Death & Disease. 2010;1(8, article e65) doi: 10.1038/cddis.2010.42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Szabo ME, Droy-Lefaix MT, Doly M, Braquet P. Free radical-mediated effects in reperfusion injury: a histologic study with superoxide dismutase and EGB 761 in rat retina. Ophthalmic Research. 1991;23(4):225–234. doi: 10.1159/000267107. [DOI] [PubMed] [Google Scholar]
- 18.Omori K, Ohira T, Uchida Y, et al. Priming of neutrophil oxidative burst in diabetes requires preassembly of the NADPH oxidase. Journal of Leukocyte Biology. 2008;84(1):292–301. doi: 10.1189/jlb.1207832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Al-Shabrawey M, Rojas M, Sanders T, et al. Role of NADPH oxidase in retinal vascular inflammation. Investigative Ophthalmology & Visual Science. 2008;49(7):3239–3244. doi: 10.1167/iovs.08-1755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Jain SK, Rains J, Jones K. Effect of curcumin on protein glycosylation, lipid peroxidation, and oxygen radical generation in human red blood cells exposed to high glucose levels. Free Radical Biology & Medicine. 2006;41(1):92–96. doi: 10.1016/j.freeradbiomed.2006.03.008. [DOI] [PubMed] [Google Scholar]
- 21.Kowluru RA, Kanwar M. Effects of curcumin on retinal oxidative stress and inflammation in diabetes. Nutrition and Metabolism. 2007;4, article 8 doi: 10.1186/1743-7075-4-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Meghana K, Sanjeev G, Ramesh B. Curcumin prevents streptozotocin-induced islet damage by scavenging free radicals: a prophylactic and protective role. European Journal of Pharmacology. 2007;577(1–3):183–191. doi: 10.1016/j.ejphar.2007.09.002. [DOI] [PubMed] [Google Scholar]
- 23.Kanitkar M, Bhonde RR. Curcumin treatment enhances islet recovery by induction of heat shock response proteins, Hsp70 and heme oxygenase-1, during cryopreservation. Life Sciences. 2008;82(3-4):182–189. doi: 10.1016/j.lfs.2007.10.026. [DOI] [PubMed] [Google Scholar]
- 24.Mandal MNA, Patlolla JMR, Zheng L, et al. Curcumin protects retinal cells from light-and oxidant stress-induced cell death. Free Radical Biology & Medicine. 2009;46(5):672–679. doi: 10.1016/j.freeradbiomed.2008.12.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Jung K-A, Kwak M-K. The Nrf2 system as a potential target for the development of indirect antioxidants. Molecules. 2010;15(10):7266–7291. doi: 10.3390/molecules15107266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Molina-Jijón E, Tapia E, Zazueta C, et al. Curcumin prevents Cr(VI)-induced renal oxidant damage by a mitochondrial pathway. Free Radical Biology and Medicine. 2011;51(8):1543–1557. doi: 10.1016/j.freeradbiomed.2011.07.018. [DOI] [PubMed] [Google Scholar]
- 27.Gao X, Kuo J, Jiang H, et al. Immunomodulatory activity of curcumin: suppression of lymphocyte proliferation, development of cell-mediated cytotoxicity, and cytokine production in vitro. Biochemical Pharmacology. 2004;68(1):51–61. doi: 10.1016/j.bcp.2004.03.015. [DOI] [PubMed] [Google Scholar]
- 28.Binion DG, Heidemann J, Li MS, Nelson VM, Otterson MF, Rafiee P. Vascular cell adhesion molecule-1 expression in human intestinal microvascular endothelial cells is regulated by PI 3-kinase/Akt/MAPK/NF- κB: inhibitory role of curcumin. American Journal of Physiology: Gastrointestinal and Liver Physiology. 2009;297(2):G259–G268. doi: 10.1152/ajpgi.00087.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Deeb D, Jiang H, Gao X, et al. Curcumin [1,7-bis(4-hydroxy-3-methoxyphenyl)-1-6-heptadine-3,5-dione; C21H20O6] sensitizes human prostate cancer cells to tumor necrosis factor-related apoptosis-inducing ligand/Apo2L-induced apoptosis by suppressing nuclear factor-κB via inhibition of the prosurvival Akt signaling pathway. The Journal of Pharmacology and Experimental Therapeutics. 2007;321(2):616–625. doi: 10.1124/jpet.106.117721. [DOI] [PubMed] [Google Scholar]
- 30.Banerjee M, Tripathi LM, Srivastava VML, Puri A, Shukla R. Modulation of inflammatory mediators by ibuprofen and curcumin treatment during chronic inflammation in rat. Immunopharmacology and Immunotoxicology. 2003;25(2):213–224. doi: 10.1081/iph-120020471. [DOI] [PubMed] [Google Scholar]
- 31.Kumar A, Dhawan S, Hardegen NJ, Aggarwal BB. Curcumin (diferuloylmethane) inhibition of tumor necrosis factor (TNF)- mediated adhesion of monocytes to endothelial cells by suppression of cell surface expression of adhesion molecules and of nuclear factor-κB activation. Biochemical Pharmacology. 1998;55(6):775–783. doi: 10.1016/s0006-2952(97)00557-1. [DOI] [PubMed] [Google Scholar]
- 32.Gupta B, Ghosh B. Curcuma longa inhibits TNF-α induced expression of adhesion molecules on human umbilical vein endothelial cells. International Journal of Immunopharmacology. 1999;21(11):745–757. doi: 10.1016/s0192-0561(99)00050-8. [DOI] [PubMed] [Google Scholar]
- 33.Patumraj S, Wongeakin N, Sridulyakul P, Jariyapongskul A, Futrakul N, Bunnag S. Combined effects of curcumin and vitamin C to protect endothelial dysfunction in the iris tissue of STZ-induced diabetic rats. Clinical Hemorheology and Microcirculation. 2006;35(4):481–489. [PubMed] [Google Scholar]
- 34.Wongeakin N, Sridulyakul P, Jariyapongskul A, Futrakula N, Bunnag SC, Patumraj S. Effects of curcumin and tetrahydrocurcumin on stz-induced diabetic rats. Journal of Vascular Research. 2006;43:p. 32. [PubMed] [Google Scholar]
- 35.American Diabetes Association. Diagnosis and classification of diabetes mellitus. Diabetes Care. 2011;34(supplement 1):S62–S69. doi: 10.2337/dc11-S062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Bandello F, Brancato R, Lattanzio R, Galdini M, Falcomata B. Relation between iridopathy and retinopathy in diabetes. British Journal of Ophthalmology. 1994;78(7):542–545. doi: 10.1136/bjo.78.7.542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Jariyapongskul A, Rungjaroen T, Kasetsuwan N, Patumraj S, Seki J, Niimi H. Long-term effects of oral vitamin C supplementation on the endothelial dysfunction in the iris microvessels of diabetic rats. Microvascular Research. 2007;74(1):32–38. doi: 10.1016/j.mvr.2007.03.002. [DOI] [PubMed] [Google Scholar]
- 38.Stuehmer C, Schumann P, Bormann K, et al. A new model for chronic in vivo analysis of the periosteal microcirculation. Microvascular Research. 2009;77(2):104–108. doi: 10.1016/j.mvr.2008.09.008. [DOI] [PubMed] [Google Scholar]
- 39.Saetzler RK, Jallo J, Lehr HA, et al. Intravital fluorescence microscopy: impact of light-induced phototoxicity on adhesion of fluorescently labeled leukocytes. Journal of Histochemistry and Cytochemistry. 1997;45(4):505–513. doi: 10.1177/002215549704500403. [DOI] [PubMed] [Google Scholar]
- 40.Byfield G, Budd S, Hartnett ME. The role of supplemental oxygen and JAK/STAT signaling in intravitreous neovascularization in a ROP rat model. Investigative Ophthalmology & Visual Science. 2009;50(7):3360–3365. doi: 10.1167/iovs.08-3256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Trichonas G, Manola A, Morizane Y, et al. A novel nonradioactive method to evaluate vascular barrier breakdown and leakage. Investigative Ophthalmology & Visual Science. 2010;51(3):1677–1682. doi: 10.1167/iovs.09-4193. [DOI] [PubMed] [Google Scholar]
- 42.Ohkawa H, Ohishi N, Yagi K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Analytical Biochemistry. 1979;95(2):351–358. doi: 10.1016/0003-2697(79)90738-3. [DOI] [PubMed] [Google Scholar]
- 43.Chen L, Zhang J, Zhang Y, Wang Y, Wang B. Improvement of inflammatory responses associated with NF-κB pathway in kidneys from diabetic rats. Inflammation Research. 2008;57(5):199–204. doi: 10.1007/s00011-006-6190-z. [DOI] [PubMed] [Google Scholar]
- 44.Shelev A. Lack of TXNIP protects β-cells against glucotoxicity. Biochemical Society Transactions. 2008;36, part 5:963–965. doi: 10.1042/BST0360963. [DOI] [PubMed] [Google Scholar]
- 45.Rungseesantivanon S, Thenchaisri N, Ruangvejvorachai P, Patumraj S. Curcumin supplementation could improve diabetes-induced endothelial dysfunction associated with decreased vascular superoxide production and PKC inhibition. BMC Complementary and Alternative Medicine. 2010;10, article 57 doi: 10.1186/1472-6882-10-57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Rungseesantivanon S, Thengchaisri N, Ruangvejvorachai P, Patumraj S. Curcumin improves prostanoid ratio in diabetic mesenteric arteries associated with cyclooxygenase-2 and NF-kappaB suppression. Diabetes, Metabolic Syndrome and Obesity: Targets and Therapy. 2010;3:421–429. doi: 10.2147/DMSOTT.S14882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Seo K, Choi M, Jung UJ, et al. Effect of curcumin supplementation on blood glucose, plasma insulin, and glucose homeostasis related enzyme activities in diabetic db/db mice. Molecular Nutrition and Food Research. 2008;52(9):995–1004. doi: 10.1002/mnfr.200700184. [DOI] [PubMed] [Google Scholar]
- 48.Suryanarayana P, Saraswat M, Mrudula T, Krishna TP, Krishnaswamy K, Reddy GB. Curcumin and turmeric delay streptozotocin-induced diabetic cataract in rats. Investigative Ophthalmology & Visual Science. 2005;46(6):2092–2099. doi: 10.1167/iovs.04-1304. [DOI] [PubMed] [Google Scholar]
- 49.Chanpoo M, Petchpiboonthai H, Panyarachun B, Anupunpisit V. Effect of curcumin in the amelioration of pancreatic islets in streptozotocin-induced diabetic mice. Journal of the Medical Association of Thailand. 2010;93(supplement 6):S152–S159. [PubMed] [Google Scholar]
- 50.Nam SM, Lee MY, Koh JH, et al. Effects of NADPH oxidase inhibitor on diabetic nephropathy in OLETF rats: the role of reducing oxidative stress in its protective property. Diabetes Research and Clinical Practice. 2009;83(2):176–182. doi: 10.1016/j.diabres.2008.10.007. [DOI] [PubMed] [Google Scholar]
- 51.Awasthi H, Tota S, Hanif K, Nath C, Shukla R. Protective effect of curcumin against intracerebral streptozotocin induced impairment in memory and cerebral blood flow. Life Sciences. 2010;86(3-4):87–94. doi: 10.1016/j.lfs.2009.11.007. [DOI] [PubMed] [Google Scholar]
- 52.Gupta SK, Kumar B, Nag TC, et al. Curcumin prevents experimental diabetic retinopathy in rats through its hypoglycemic, antioxidant, and anti-inflammatory mechanisms. Journal of Ocular Pharmacology and Therapeutics. 2011;27(2):123–130. doi: 10.1089/jop.2010.0123. [DOI] [PubMed] [Google Scholar]
- 53.Menon VP, Sudheer AR. Antioxidant and anti-inflammatory properties of curcumin. Advances in Experimental Medicine and Biology. 2007;595:105–125. doi: 10.1007/978-0-387-46401-5_3. [DOI] [PubMed] [Google Scholar]
- 54.Joussen AM, Poulaki V, Le ML, et al. A central role for inflammation in the pathogenesis of diabetic retinopathy. FASEB Journal. 2004;18(12):1450–1452. doi: 10.1096/fj.03-1476fje. [DOI] [PubMed] [Google Scholar]
- 55.Pemp B, Schmetterer L. Ocular blood flow in diabetes and age-related macular degeneration. Canadian Journal of Ophthalmology. 2008;43(3):295–301. doi: 10.3129/i08-049. [DOI] [PubMed] [Google Scholar]
- 56.Aoki H, Takada Y, Kondo S, Sawaya R, Aggarwal BB, Kondo Y. Evidence that curcumin suppresses the growth of malignant gliomas in vitro and in vivo through induction of autophagy: role of akt and extracellular signal-regulated kinase signaling pathways. Molecular Pharmacology. 2007;72(1):29–39. doi: 10.1124/mol.106.033167. [DOI] [PubMed] [Google Scholar]
- 57.Yang J, Lane PH, Pollock JS, Carmines PK. Protein kinase C-dependent NAD(P)H oxidase activation induced by type 1 diabetes in renal medullary thick ascending limb. Hypertension. 2010;55(2):468–473. doi: 10.1161/HYPERTENSIONAHA.109.145714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Asaba K, Tojo A, Onozato ML, et al. Effects of NADPH oxidase inhibitor in diabetic nephropathy. Kidney International. 2005;67(5):1890–1898. doi: 10.1111/j.1523-1755.2005.00287.x. [DOI] [PubMed] [Google Scholar]
- 59.Tsai KH, Wang WJ, Lin CW, et al. NADPH oxidase-derived superoxide Anion-induced apoptosis is mediated via the JNK-dependent activation of NF-κB in cardiomyocytes exposed to high glucose. Journal of Cellular Physiology. 2012;227(4):1347–1357. doi: 10.1002/jcp.22847. [DOI] [PubMed] [Google Scholar]
- 60.Szabó A, Farkas A, Vámos R, Bajor T, Hrabák A. The inhibition of retinal inducible nitric oxide synthase overexpression and the attenuation of experimental uveitis by anti-inflammatory drugs in rats. Inflammation Research. 2004;53(6):262–267. doi: 10.1007/s00011-004-1259-z. [DOI] [PubMed] [Google Scholar]