

Abstract
Acetylcholinesterase (AChE) inhibitors are widely used for the symptomatic treatment of Alzheimer’s disease and other dementias. More recent use is for myasthenia gravis. Many of these inhibitors interact with the second known cholinesterase, butyrylcholinesterase (BChE). Further, evidence shows that acetylcholine plays a role in suppression of cytokine release through a “cholinergic anti-inflammatory pathway” which raises questions about the role of these inhibitors in the immune system. This review covers research and discussion of the role of the inhibitors in modulating the immune response using as examples the commonly available drugs, donepezil, galantamine, huperzine, neostigmine and pyridostigmine. Major attention is given to the cholinergic anti-inflammatory pathway, a well-described link between the central nervous system and terminal effector cells in the immune system.
Keywords: cholinergic anti-inflammatory pathway, acetylcholinesterase, butyrylcholinesterase, Alzheimer disease, neuroimmunity, neuromodulation
1. Introduction
Inhibitors of AChE (EC 3.1.1.7.) and BChE (EC 3.1.1.8) are neurotoxic compounds capable of causing central, peripheral or both central and peripheral cholinergic crises. A number of these compounds have also found application as drugs developed for the treatment of Alzheimer’s disease (AD) and myasthenia gravis [1,2]. These are based on the premise that increasing the availability of acetylcholine (ACh) at acetylcholine receptors in the brain, results in better neuron to neuron transport that will improve cognitive function. Cholinergic nerves, however, can be found in both the central (CNS) and peripheral (PNS) nervous systems and disparate body tissues [3]. Drugs that cross the blood brain barrier do not have dissociable groups as can be seen in the case of AD drugs [4]. Some of these drugs cannot penetrate the CNS and this property makes them suitable for use in myasthenia gravis [5]. For a long time, regulation of immunity was not considered an effect of AChE inhibitors. However, recent evidence casts new light on the subject. In this review, we explore the link between immunity and the AChE inhibitors as currently available AChE inhibiting drugs for AD.
2. The Cholinergic Anti-Inflammatory Pathway
Ach is a ubiquitous neurotransmitter [6,7] and found even in the roundworm Caenorhabditis elegans, one of the simplest organisms with a nervous system [8,9]. In the roundworm one third of the nervous system is cholinergic [10]. Humans have a large percentage of nervous system that is cholinergic including the CNS. Cholinergic nerves also form a major part of the parasympathetic and sympathetic nervous systems [11,12]. The wider significance of Ach is in understanding the biological effects of tested toxins and/or medical drugs: as any immunological effects of AChE inhibitors can involve both CNS and PNS, this has to be taken into consideration in interpreting any findings. For this reason, vagotomy, used to study the cholinergic anti-inflammatory pathway in animal experiments or selecting compounds that do not cross the blood brain barrier should be considered carefully before drawing any conclusions as to which pathway is involved in the proposed mechanism.
The cholinergic system is tightly associated with the cholinergic anti-inflammatory pathway dominantly located in blood and mucosa. This pathway is a regulatory link between nerve terminations in blood and macrophages expressing the α7 nicotinic acetylcholine receptor (α7 nAChR) on their surface [11,13,14]. For a long time, the mechanisms of inflammatory regulation remained unclear. Discovery of the cholinergic anti-inflammatory pathway, however, allowed us to understand how the CNS is involved in the regulation of innate immunity [15,16,17,18,19,20,21,22,23,24]. AChE bound on erythrocytes plays an important role in termination of cholinergic anti-inflammatory pathway activation [11,25]. AChE activity is typically low in AD patients treated with AChE inhibitors [26]. Compared to AChE, BChE is constituted in the liver and secreted into the plasma where the enzyme is dissolved [27]. Apart from the fact that the conversion rate of Ach by BChE is lower than the conversion by AChE, BChE can substitute for AChE and split the neurotransmitter once they make contact [28,29]. The effect of BChE became relevant once the cholinergic anti-inflammatory pathway was studied as BChE plays a greater role in the blood than in the nervous system.
The cholinergic anti-inflammatory pathway is one-way: the CNS can attenuate inflammation mediated by macrophages or any other immune cells having α7 nAChR. Ach released from the vagus nerve termination, agonizes α7 nAChR, which responds by opening a central channel allowing an influx of Ca2+ into macrophages [11,30,31]. Increased levels of Ca2+ activate the nuclear factor κ B (NF κB) resulting in suppression of inflammatory cytokine production including tumor necrosis factor α (TNFα), high mobility group box of proteins and interleukin 6 (IL-6) [32,33]. These blood AChE and plasma BChE are able to terminate the stimulation of the cholinergic anti-inflammatory pathway due to splitting ACh. The principle of the pathway is depicted in Figure 1.
Figure 1.

Principle of the cholinergic anti-inflammatory pathway; abbreviations: ACh-acetylcholine; AChE-acetylcholinesterase; BChE-butyrylcholinesterase; HMGB-high mobility group box; IL-6-interleukin 6; NFκB-nuclear factor kappa B; TNFα-tumor necrosis factor alpha.
The primary purpose of using AChE inhibitors in pharmacology is not modulation of immunity related pathologies. However, recent studies indicate that these inhibitors can cause a significant modulation of immunity as a side effect [29,34,35]. As seen from Figure 1, they can modulate the cholinergic anti-inflammatory pathway via protection of Ach from splitting by cholinesterases and thus enhancing the pathway. The mechanism of action is probably less effective than the standard mode but it becomes relevant when someone is using an inhibitor of cholinesterases in large amounts and/or for a long time such as patients suffering from AD.
Apart from the regulation processes, some inhibitors can influence immunity via forming antigens by reaction with e.g., plasma proteins. The immune system is thus activated and the stimulation counteracts the anti-inflammatory action. This effect is, however, very weak but it can play a role in forming antibody proteins modified by nerve agents [36].
3. Division of Inhibitors
The structure of AChE and BChE has been extensively reviewed in the following publications [7,29,37,38]. In brief, AChE has a more developed peripheral anionic site and narrower aromatic gorge than BChE. Aromatic compounds have higher affinity for AChE than BChE. Some aromatic inhibitors of AChE do not inhibit BChE, for example aflatoxins [39,40]. When the role of the cholinesterases is evaluated in humans, their genomic diversity and posttranslational modifications have to be taken into account [41,42].
Organophosphorus compounds are irreversible inhibitors of both AChE and BChE. They bind to the active site of the cholinesterases and easily cross the blood brain barrier [6,43]. Nerve agents, e.g., sarin, soman, tabun, and some highly toxic compounds, formerly used as pesticides (paraoxon, parathion, malaoxon, malathion), are examples [44,45,46,47]. High toxicity characterizes organophosphorus inhibitors that are used in chemical warfare or as pesticides. Their pharmacological importance is relatively small; Metrifonate (trichlorfon) was chosen for AD treatment and became an exception but it was withdrawn because of adverse effects [48,49,50]. Though organophosphorus compounds typically inhibit both AChE and BChE, tetraisopropyl pyrophosphoramide, also known as iso-OMPA, is dissimilar to the other inhibitors. It does not penetrate to the active site of AChE and it inhibits BChE only. It is typically used as a reagent for rapid distinction between AChE and BChE activity in biological samples [51,52].
Carbamate inhibitors bind to the active site of both cholinesterases like organophosphorus inhibitors; however, the covalent bound is not stable and the carbamate moiety is hydrolytically split from the active site after some time [29,53,54]. The mechanism of carbamate binding is sometimes called pseudo-irreversible because of carbamate moiety spontaneous hydrolysis and resurrection of cholinesterase activity. From a pharmacological point of view, there is a big difference between carbamates and organophosphorus inhibitors. Many carbamates do not cross the blood brain barrier and the carbamate moiety has to be modified or encapsulated to cross it [55,56,57]. Quarternary ammonium containing pyridostigmine and neostigmine are examples. On the other hand, the blood brain barrier is not impenetrable by all carbamates e.g., rivastigmine (see further text) and physostigmine easily reach AChE in the brain [58,59,60].
AChE and BChE can be inhibited by a group of secondary metabolites from plants and fungi. Galantamine and huperzine are examples of plant alkaloids used in pharmacology. Alkaloids α-chaconine, α-solanine, tomatine, berberine, palmatine and jatrorrhizine are other metabolites that inhibit cholinesterases [29,61,62,63,64,65]. Aflatoxins too, can be introduced as secondary metabolites from fungi.
Disparate synthetic drugs not belonging to carbamates and organophosphates can be mentioned last but not least. Donepezil is the most relevant compound of this group. (See the next chapter). Tacrine (1,2,3,4-tetrahydroacridin-9-amine) is another synthetic drug which easily crosses the blood brain barrier and is used as a highly effective drug for ameliorating Alzheimer disease manifestation by inhibition AChE and by lower but still effective inhibition of BChE [53,66,67]. It was withdrawn from clinical use because of adverse effects. Hepatotoxicity was the main pathological consequence of tacrine intake [68,69]. The basic facts about groups of inhibitors are summarized in Table 1.
Table 1.
Summarization of facts about cholinesterases’ inhibitors.
| Group of Compounds | Compounds (Examples) | Mechanism of Inhibition | Inhibition of AChE and BChE | Penetration through Blood Brain Barrier | Importance as Drugs | References |
|---|---|---|---|---|---|---|
| Organophosphates | sarin, soman, tabun, malaoxon | irreversible | equal to AChE and BChE | Good | Low | [6,43,44,45,46,47,48,49,50] |
| Carbamates | pyridostigmine, physostigmine neostigmine, rivastigmine | pseudo-irreversible | equal to AChE and BChE | low (pyridostigmine, neostigmine), good (physostigmine, rivastigmine) | High | [29,53,54,55,56,57,58,59,60] |
| - | Tacrine | non-competitive | AChE > BChE | Good | former drug, discontinued now | [53,66,68,69] |
| - | Galantamine | competitive | AChE | Good | High | [73,74] |
| - | Donepezil | non-competitive | AChE | Good | High | [75,76] |
| - | huperzine A | non-competitive | AChE >> BChE | Good | will increase | [29,75,77,78,79] |
In considering the role of the inhibitors in modulating immunity, other factors such as the environment and genetic disposition should be taken into account. This conclusion is based on blood cholinesterase activity in male volunteers working with pesticides [70]. The activity varied in infected patients with proven bacterial meningitis [71]. The sensitivity of humans to the inhibitors can also significantly be affected because detoxification mechanisms have unequal efficacy. This idea was, e.g., demonstrated by Sonali et al.on AD patients treated with rivastigmine [72]. Variability in inhibitor effects should be considered when conclusions are drawn from animal models and cell lines, extrapolated to humans.
4. Inhibitors that Cross the Blood Brain Barrier
The following text focuses on galantamine, donepezil, huperzine and rivastigmine. Galantamine (Figure 2) is a drug used for treating AD and related dementias. Currently, galantamine is an alternative to rivastigmine and can be given to patients with similar stages of dementia. On the market, it is sold under the trade names Razadyne™, Razadyne™ER, Reminyl™ER, and Reminyl® The drug was firstly isolated by soviet scientists Mashkovsky and Kruglikova-Lvova from bulbs of Caucasian snowdrops Galanthus sp. in the early 1950s and chemical synthesis was introduced in the following decades [80]. After marketing of the drug, it drove out the more toxic tacrine [81] and has become one of the best drugs for Alzheimer disease treatment [73].
Figure 2.

Structure of cited compounds that cross the blood brain barrier.
Galantamine is a competitive inhibitor of AChE and an allosteric modulator of nAChR [74]. Its dual action on both AChE and nAChR is an advantage and unlike other marketed drugs that inhibit AChE. It is believed that galantamine may interact with the cholinergic anti-inflammatory pathway via direct modulation of the α7 nAChR [82]. The anti-inflammatory pathway modulation may explain the activation of microglia followed by amyloid beta clearance [83]. In a rat model, galantamine was also approved as effective in reducing circulating TNFα which was, in the past, initiated by administration of bacterial lipopolysaccharide [84]. For this reason, galantamine could act not only as a drug for symptomatic treatment but as a compound for slowing Alzheimer disease progression. This conclusion is not, however, commonly accepted and more detailed evidence of the process is needed.
Donepezil is a selective, noncompetitive inhibitor of AChE. The inhibition is quite effective as the equilibrium constant is reported to be 12.5 nmol/L for AChE from rat erythrocytes [75]. Donepezil is available under the trade name Aricept as a highly effective drug for Alzheimer disease, originally developed by Eisai and Pfizer. The structure of donepezil is depicted in Figure 3. Clinical experience with donepezil is good: it is well tolerated and slowly eliminated so that the drug can be taken over long periods [76]. Compared to other drugs for Alzheimer disease, donepezil works via a simple pathway based on AChE inhibition. It is not involved in other pathways and does not involve BChE inhibition [29].
Figure 3.
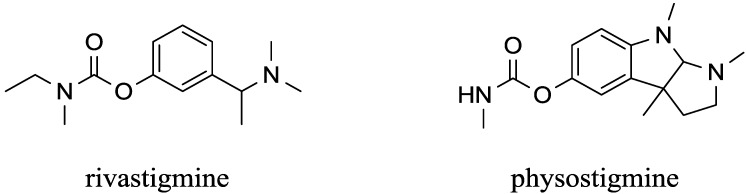
Structure of carbamates that cross the blood brain barrier.
We can assume that donepezil activates the cholinergic anti-inflammatory pathway via inhibition of AChE and increased availability of ACh. However, Hwang and coworkers found anti-inflammatory effects on microglia cell lines where no AChE was present [85]. These authors showed significant attenuation of TNFα, IL-1, and NF-κB release. From these results, we can infer that donepezil is either able to directly stimulate α7 nAChR and not act as an inhibitor only or it has some other unknown pathway. Beside the anti-inflammatory pathway, donepezil was proven to be able to modulate viral progression and the modulation is probably based on a mechanism other than agonism of α7 nAChR [86,87]. This fact would support the results on the microglia cells. Unfortunately, the antiviral effect of donepezil was not studied further, though the issue deserves greater attention.
Huperzine is a secondary metabolite from the lycopods, Huperzia from which it is isolated for pharmacological purposes. The upper production of huperzine alkaloids can be found in H. serata. More types of huperzine are known. However, huperzine A (Figure 2) is the most likely to be used in Alzeheimer disease [75]. Elaborative isolation of huperzine from plant biomass is the main disadvantage of huperzine as chemical synthesis is problematic due to expensive isolation of the (−) huperzine A from the (+) isomer which is not pharmacologically relevant [88]. Though advanced chemical synthesis protocols [89] and biotechnology processes [90] for the (−) huperzine A production are described, they are far from practical use. Huperzine is used in traditional Chinese medicine and it is available as a supplement in the country. It can be taken as a mechanically milled plant tissue or as an extract prepared by chromatographic isolation [77]. In countries of the European Union and United States, pro-drug ZT-1 derived from huperzine A is clinically tested and introduction for therapy purposes is expected in the near future [91].
Huperzine A is a selective inhibitor of AChE, acting by a non-competitive or mixed mechanism [75]. Huperzine A can bind to the peripheral anionic site of AChE and the effect is reportedly responsible for amelioration of the amyloidogenic process [92]. Besides AChE inhibition, huperzine A is a potent non-competitive inhibitor of the N-methyl-d-aspartate receptor [78,79].
Huperzine A was proven to reduce neuroinflammation in experimental autoimmune encephalomyelitis in mice [93]. The authors reported a decrease in the number of inflammatory cells, interferon gamma (IFNγ), IL-17, MCP-1, RANTES, TWEAK and an increase in IL-4 and IL-10 in the course of treatment. In another experiment, huperzine inhibited activation of NF-κB, inducible nitric oxide synthase and cyclooxygenase 2 in C6 rat glioma cells [94]. Though direct molecular evidence is missing, agonism of α7 nAChR with subsequent activation of the cholinergic anti-inflammatory pathway is presumed to be involved in huperzine’s mechanism of action [95].
Rivastigmine (Figure 3) is a carbamate inhibitor of AChE as well as BChE and it easily crosses the blood brain barrier. Rivastigmine is probably the most marketed carbamate in pharmacology. The drug is sold under the trade name Exelon for the treatment of Alzheimer and Parkinson disease in early and mild stages. Currently, it is the only available drug for these diseases, which is not a reversible inhibitor of AChE since it inhibits the cholinesterases in a pseudo-irreversible manner [96]. Slow elimination of rivastigmine because of the covalent bond in the active site of the enzyme is an advantage over other drugs. The effect of rivastigmine can last until the rivastigmine moiety spontaneously splits from the cholinesterase’s active site by a decarbonylation process [4,54].
The long-term effects of rivastigmine remain unclear. It was proven that rivastigmine causes significant up-regulation of AChE expression [97]. The molecular mechanism is, however, unrevealed. In clinical tests, rivastigmine was not found to generate inflammation or have any other adverse effect. For this reason, rivastigmine is considered as a quite safe drug [98]. On the contrary, more detailed examination showed that rivastigmine can suppress inflammation [99]. Namely, decreased reactivity of encephalitogenic T lymphocytes and production of pro-inflammatory cytokines was reported [100]. More experiments on this issue will be necessary as the exact mechanism is not clear. It can be assumed that the cholinergic anti-inflammatory pathway can be activated as Ach becomes available for the α7 nAChR on macrophages and microglial cells. Direct proof is, however, missing.
Like rivastigmine, physostigmine (or eserine in some sources) crosses the blood brain barrier and can inhibit AChE and BChE in the both central and peripheral nervous systems. Physostigmine is a carbamate of natural origin that can be found in the seeds from a plant Physostigma venosum known as the Calabar bean. At the current time, cheap and reliable protocols for physostigmine synthesis are available and preferred over isolation from plants [101]. Physostigmine can be used for alleviation of glaucoma manifestation [102] and it is suitable for the treatment of Alzheimer disease [103,104]. As discussed later in the text, physostigmine can inhibit protein kinase C. This ability is not common to other carbamate inhibitors of cholinesterases [105]. Physostigmine was shown to increase the availability of ACh and stimulate the cholinergic anti-inflammatory pathway in experimental endotoxemia by lipopolysaccharide [106]. In another experiment, physostigmine regulated early inflammation and oxidative stress as the superoxide radical in rats with induced forebrain ischemia [107].
5. Peripherally Acting Carbamates-Parasympathomimetics
Most carbamates have limited ability to pass the blood brain barrier. Rivastigmine is an exception from this point of view. The poor ability to cross the blood brain barrier can be an advantage when we need a compound to regulate the PNS and not the CNS. Drugs that do this are used for two main purposes: in the treatment of myasthenia gravis [108] and in anesthesia [109]. Myasthenia gravis is an autoimmune disease where antibodies against acetylcholine receptors are created in the body. The treatment the disease can be based on administration of immunosuppressant and/or an AChE inhibitor [110]. In surgical interventions, there is a necessity to give muscle relaxants such as parasympathomimetics. The parasympathomimetics can act as polarizing agents via direct stimulation of receptors (e.g., succinylcholine) or as non-polarizing agents where the peripherally acting carbamates belong [111,112]. Pyridostigmine and neostigmine are well known examples of carbamates acting as parasympathomimetics [113]. Beside the carbamates, a quaternary nitrogen containing inhibitor edrophonium is also used as a parasympathomimetic [114]. The chemical structures of neostigmine and pyridostigmine are shown in Figure 4. Though the blood brain barrier seems to be impenetrable for compounds like pyridostigmine and neostigmine, the contrary is true. For example, Friedman et al. showed that stress conditions can cause penetrability of blood brain barrier by pyridostigmine [115]. This factor has to be considered in critical evaluation of peripherally acting carbamates. Strong stressogenic conditions can cause peripherally acting carbamates to have the same effect as a centrally acting one.
Figure 4.
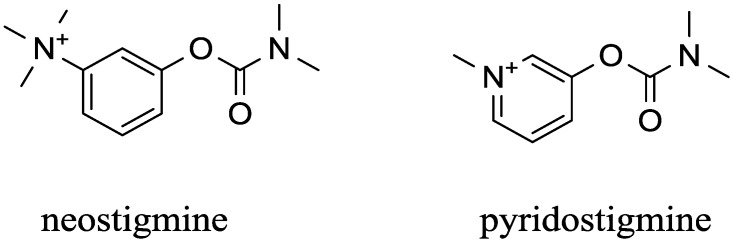
Structure of carbamate inhibitors that do not cross the blood brain barrier.
The peripherally acting carbamates have similar action to rivastigmine. They are pseudo-irreversible inhibitors of both AChE and BChE [29]. Selective effect on the PNS is the main difference between the carbamates such as neostigmine and pyridostigmine and the centrally acting rivastigmine. The inability to cross blood brain barrier discriminates the peripherally acting carbamates from being used for regulation of neuroinflammation and other immunity related disorders within the CNS. On the other hand, they can be expected to be favored in the generalized immunity disorder regulation whenever brain functions should be spared.
Plausible anti-inflammatory effects of the peripherally acting carbamates have been described in some papers. In an example, physostigmine (crossing blood brain barrier) and neostigmine (not crossing) reduced inflammation which was previously initiated by a bacterial lipopolysaccharide [116,117]. Similar results were reported in an experiment where mice were infected with tularemia and treated with neostigmine [118]. In this experiment, neostigmine worsened the tularemia pathology. Sun et al.tested neostigmine effects in mice and Beagle dogs [119]. They showed that neostigmine acts via the cholinergic anti-inflammatory pathway. This conclusion is supported by the fact that mice with knockout gene for α7 nAChR are not sensitive to the immunomodulatory effect. On the other hand, more experiments on the issue will be needed as some did not confirm any significant anti-inflammatory effect of carbamates [120]. Regulation of immunity can be based on pathways far from the cholinergic system. e.g., Bitzinger et al. revealed the ability of physostigmine to inhibit protein kinase C while neostigmine did not have this ability [105].
6. Conclusions
Currently available drugs for inhibiting either AChE alone or in combination with BChE are available for the treatment of AD, myasthenia gravis and other conditions. These drugs however, are not used for immunomodulation purposes at this time. This review highlights the fact that these inhibitors may affect not only the cholinergic anti-inflammatory pathway but also other unknown pathways involved in regulating immunity. A simplified mechanism for how these inhibitors may be involved in regulating immunity is depicted schematically in Figure 5. This issue deserves greater attention due to its pharmacological relevance.
Figure 5.

Simplified scheme for showing how inhibitors of cholinesterases may be involved in regulation of immunity using the cholinergic anti-inflammatory pathway. Abbreviations used in the figure: ACh—acetylcholine; CAP—cholinergic anti-inflammatory pathway; ChE—cholinesterase; HMGB—high mobility group box proteins.
Acknowledgments
The Ministry of Education, Youth and Sports of the Czech Republic is gratefully acknowledged for project LH11023. A long-term organization development plan 1011 (Faculty of Military Health Sciences, University of Defence, Czech Republic) is acknowledged as well.
Conflicts of Interest
The author declares no conflict of interest.
References
- 1.Ofek K., Soreq H. Cholinergic involvement and manipulation approaches in multiple system disorders. Chem. Biol. Interact. 2013;203:113–119. doi: 10.1016/j.cbi.2012.07.007. [DOI] [PubMed] [Google Scholar]
- 2.De los Rios C. Cholinesterase inhibitors: A patent review (2007–2011) Expert Opin. Ther. Pat. 2012;22:853–869. doi: 10.1517/13543776.2012.701619. [DOI] [PubMed] [Google Scholar]
- 3.Silman I., Sussman J.L. Acetylcholinesterase: “Classical” and “non-classical” functions and pharmacology. Curr. Opin. Pharmacol. 2005;5:293–302. doi: 10.1016/j.coph.2005.01.014. [DOI] [PubMed] [Google Scholar]
- 4.Tayeb H.O., Yang H.D., Price B.H., Tarazi F.I. Pharmacotherapies for Alzheimer’s disease: Beyond cholinesterase inhibitors. Pharmacol. Ther. 2012;134:8–25. doi: 10.1016/j.pharmthera.2011.12.002. [DOI] [PubMed] [Google Scholar]
- 5.Haines S.R., Thurtell M.J. Treatment of ocular myasthenia gravis. Curr. Treat. Option Neurol. 2012;14:103–112. doi: 10.1007/s11940-011-0151-8. [DOI] [PubMed] [Google Scholar]
- 6.Marrs T.C., Maynard R.L. Neurotranmission systems as targets for toxicants: A review. Cell Biol. Toxicol. 2013;29:381–396. doi: 10.1007/s10565-013-9259-9. [DOI] [PubMed] [Google Scholar]
- 7.Colovic M.B., Krstic D.Z., Lazarevic-Pasti T.D., Bondzic A.M., Vasic V.M. Acetylcholinesterase inhibitors: Pharmacology and toxicology. Curr. Neuropharmacol. 2013;11:315–335. doi: 10.2174/1570159X11311030006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Rakowski F., Srinivasan J., Sternberg P.W., Karbowski J. Synaptic polarity of the interneuron circuit controlling c. Elegans locomotion. Front. Comput. Neurosci. 2013;7:128. doi: 10.3389/fncom.2013.00128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kosinski R.A., Zaremba M. Dynamics of the model of the caenorhabditis elegans neural network. Acta Phys. Pol. B. 2007;38:2201–2210. [Google Scholar]
- 10.Rand J.B. Acetylcholine. WormBook. 2007 doi: 10.1895/wormbook1.131.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Pohanka M. Alpha7 nicotinic acetylcholine receptor is a target in pharmacology and toxicology. Int. J. Mol. Sci. 2012;13:2219–2238. doi: 10.3390/ijms13022219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Bellier J.P., Kimura H. Peripheral type of choline acetyltransferase: Biological and evolutionary implications for novel mechanisms in cholinergic system. J. Chem. Neuroanat. 2011;42:225–235. doi: 10.1016/j.jchemneu.2011.02.005. [DOI] [PubMed] [Google Scholar]
- 13.Wessler I., Kirkpatrick C.J. Acetylcholine beyond neurons: The non-neuronal cholinergic system in humans. Br. J. Pharmacol. 2008;154:1558–1571. doi: 10.1038/bjp.2008.185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Rosas-Ballina M., Tracey K.J. Cholinergic control of inflammation. J. Intern. Med. 2009;265:663–679. doi: 10.1111/j.1365-2796.2009.02098.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Bernik T.R., Friedman S.G., Ochani M., DiRaimo R., Ulloa L., Yang H., Sudan S., Czura C.J., Ivanova S.M., Tracey K.J. Pharmacological stimulation of the cholinergic antiinflammatory pathway. J. Exp. Med. 2002;195:781–788. doi: 10.1084/jem.20011714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Borovikova L.V., Ivanova S., Zhang M.H., Yang H., Botchkina G.I., Watkins L.R., Wang H.C., Abumrad N., Eaton J.W., Tracey K.J. Vagus nerve stimulation attenuates the systemic inflammatory response to endotoxin. Nature. 2000;405:458–462. doi: 10.1038/35013070. [DOI] [PubMed] [Google Scholar]
- 17.Ofek K., Krabbe K.S., Evron T., Debecco M., Nielsen A.R., Brunnsgaad H., Yirmiya R., Soreq H., Pedersen B.K. Cholinergic status modulations in human volunteers under acute inflammation. J. Mol. Med. 2007;85:1239–1251. doi: 10.1007/s00109-007-0226-x. [DOI] [PubMed] [Google Scholar]
- 18.Andersson U., Tracey K.J. Neural reflexes in inflammation and immunity. J. Exp. Med. 2012;209:1057–1068. doi: 10.1084/jem.20120571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Pavlov V.A., Ochani M., Gallowitsch-Puerta M., Ochani K., Huston J.M., Czura C.J., Al-Abed Y., Tracey K.J. Central muscarinic cholinergic regulation of the systemic inflammatory response during endotoxemia. Proc. Natl. Acad. Sci. USA. 2006;103:5219–5223. doi: 10.1073/pnas.0600506103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Pavlov V.A., Parrish W.R., Rosas-Ballina M., Ochani M., Puerta M., Qchani K., Chavan S., Al-Abed Y., Tracey K.J. Brain acetylcholinesterase activity controls systemic cytokine levels through the cholinergic anti-inflammatory pathway. Brain Behav. Immun. 2009;23:41–45. doi: 10.1016/j.bbi.2008.06.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Tracey K.J. The inflammatory reflex. Nature. 2002;420:853–859. doi: 10.1038/nature01321. [DOI] [PubMed] [Google Scholar]
- 22.Tracey K.J. Reflex control of immunity. Nat. Rev. Immunol. 2009;9:418–428. doi: 10.1038/nri2566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Wang H., Yu M., Ochani M., Amella C.A., Tanovic M., Susarla S., Li J.H., Wang H.C., Yang H., Ulloa L., et al. Nicotinic acetylcholine receptor alpha 7 subunit is an essential regulator of inflammation. Nature. 2003;421:384–388. doi: 10.1038/nature01339. [DOI] [PubMed] [Google Scholar]
- 24.Wang H., Liao H., Ochani M., Justiniani M., Lin X.C., Yang L.H., Al-Abed Y., Wang H.C., Metz C., Miller E.J., et al. Cholinergic agonists inhibit hmgb1 release and improve survival in experimental sepsis. Nat. Med. 2004;10:1216–1221. doi: 10.1038/nm1124. [DOI] [PubMed] [Google Scholar]
- 25.Silva-Herdade A.S., Saldanha C. Effects of acetylcholine on an animal mode of inflammation. Clin. Hemorheol. Microcirc. 2013;53:209–216. doi: 10.3233/CH-121646. [DOI] [PubMed] [Google Scholar]
- 26.Coin A., Perissinotto E., Catanzaro S., Mosele M., de Rui M., Girardi A., Inelmen E.M., Toffanello E.D., Manzato E., Sergi G. Effects of 21 months of cholinesterase inhibitors on cognitive and functional decline in demented patients. Aging Clin. Exp. Res. 2012;24:14–16. [PubMed] [Google Scholar]
- 27.Iwasaki T., Yoneda M., Nakajima A., Terauchi Y. Serum butyrylcholinesterase is strongly associated with adiposity, the serum lipid profile and insulin resistance. Intern. Med. 2007;46:1633–1639. doi: 10.2169/internalmedicine.46.0049. [DOI] [PubMed] [Google Scholar]
- 28.Karlsson D., Fallarero A., Brunhofer G., Mayer C., Prakash O., Mohan C.G., Vuorela P., Erker T. The exploration of thienothiazines as selective butyrylcholinesterase inhibitors. Eur. J. Pharm. Sci. 2012;47:190–205. doi: 10.1016/j.ejps.2012.05.014. [DOI] [PubMed] [Google Scholar]
- 29.Pohanka M. Cholinesterases, a target of pharmacology and toxicology. Biomed. Pap. 2011;155:219–229. doi: 10.5507/bp.2011.036. [DOI] [PubMed] [Google Scholar]
- 30.Noelker C., Stuckenholz V., Reese J.P., Alvarez-Fischer D., Sankowski R., Rausch T., Oertel W.H., Hartmann A., van Patten S., Al-Abed Y., et al. Cni-1493 attenuates neuroinflammation and dopaminergic neurodegeneration in the acute mptp mouse model of Parkinson’s disease. Neurodegener. Dis. 2013;12:103–110. doi: 10.1159/000342714. [DOI] [PubMed] [Google Scholar]
- 31.Lee L., Kosuri P., Arancio O. Picomolar amyloid-beta peptides enhance spontaneous astrocyte calcium transients. J. Alzheimers Dis. 2014;38:49–62. doi: 10.3233/JAD-130740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Sun P., Zhou K.W., Wang S., Li P., Chen S.J., Lin G.P., Zhao Y., Wang T.H. Involvement of mapk/NF-κB signaling in the activation of the cholinergic anti-inflammatory pathway in experimental colitis by chronic vagus nerve stimulation. PLoS One. 2013;8:e69424. doi: 10.1371/journal.pone.0069424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.De Haan J.J., Hadfoune M., Lubbers T., Hodin C., Lenaerts K., Ito A., Verbaeys I., Skynner M.J., Cailotto C., van der Vliet J., et al. Lipid-rich enteral nutrition regulates mucosal mast cell activation via the vagal anti-inflammatory reflex. Am. J. Physiol.-Gastroint. Liver Physiol. 2013;305:G383–G391. doi: 10.1152/ajpgi.00333.2012. [DOI] [PubMed] [Google Scholar]
- 34.Starec M., Sinet M., Kodym P., Rosina J., Fiserova A., Desforges B., Rouveix B. The effect of drugs on the mortality of mice inoculated with friend leukaemia virus or toxoplasma gondii. Physiol. Res. 1997;46:107–111. [PubMed] [Google Scholar]
- 35.Pohanka M. Role of oxidative stress in infectious diseases. A review. Folia Microbiol. 2013;58:503–513. doi: 10.1007/s12223-013-0239-5. [DOI] [PubMed] [Google Scholar]
- 36.Smirnov I., Belogurov A., Friboulet A., Masson P., Gabibov A., Renard P.Y. Strategies for the selection of catalytic antibodies against organophosphorus nerve agents. Chem.-Biol. Interact. 2013;203:196–201. doi: 10.1016/j.cbi.2012.10.011. [DOI] [PubMed] [Google Scholar]
- 37.Da Silva V.B., de Andrade P., Kawano D.F., Morais P.A.B., de Almeida J.R., Carvalho I., Taft C.A., da Silva C. In silico design and search for acetylcholinesterase inhibitors in Alzheimer’s disease with a suitable pharmacokinetic profile and low toxicity. Future Med. Chem. 2011;3:947–960. doi: 10.4155/fmc.11.67. [DOI] [PubMed] [Google Scholar]
- 38.Krall W.J., Sramek J.J., Cutler N.R. Cholinesterase inhibitors: A therapeutic strategy for alzheimer disease. Ann. Pharmacother. 1999;33:441–450. doi: 10.1345/aph.18211. [DOI] [PubMed] [Google Scholar]
- 39.Arduini F., Errico I., Amine A., Micheli L., Palleschi G., Moscone D. Enzymatic spectrophotometric method for aflatoxin b detection based on acetylcholinesterase inhibition. Anal. Chem. 2007;79:3409–3415. doi: 10.1021/ac061819j. [DOI] [PubMed] [Google Scholar]
- 40.Arduini F., Amine A., Moscone D., Palleschi G. Biosensors based on cholinesterase inhibition for insecticides, nerve agents and aflatoxin b-1 detection (review) Microchim. Acta. 2010;170:193–214. doi: 10.1007/s00604-010-0317-1. [DOI] [Google Scholar]
- 41.Brazzolotto X., Wandhammer M., Ronco C., Trovaslet M., Jean L., Lockridge O., Renard P.Y., Nachon F. Human butyrylcholinesterase produced in insect cells: Huprine-based affinity purification and crystal structure. FEBS J. 2012;279:2905–2916. doi: 10.1111/j.1742-4658.2012.08672.x. [DOI] [PubMed] [Google Scholar]
- 42.Massoulie J. The origin of the molecular diversity and functional floating of cholinesterases. Neurosignals. 2002;11:130–143. doi: 10.1159/000065054. [DOI] [PubMed] [Google Scholar]
- 43.Marrs T.C. Organophosphate poisoning. Pharmacol. Ther. 1993;58:51–66. doi: 10.1016/0163-7258(93)90066-M. [DOI] [PubMed] [Google Scholar]
- 44.Furtado M.D., Rossetti F., Chanda S., Yourick D. Exposure to nerve agents: From status epilepticus to neuroinflammation, brain damage, neurogenesis and epilepsy. Neurotoxicology. 2012;33:1476–1490. doi: 10.1016/j.neuro.2012.09.001. [DOI] [PubMed] [Google Scholar]
- 45.Dorandeu F., Foquin A., Briot R., Delacour C., Denis J., Alonso A., Froment M.T., Renault F., Lallement G., Masson P. An unexpected plasma cholinesterase activity rebound after challenge with a high dose of the nerve agent vx. Toxicology. 2008;248:151–157. doi: 10.1016/j.tox.2008.03.013. [DOI] [PubMed] [Google Scholar]
- 46.Raghu P., Madhusudana Reddy T., Reddaiah K., Swamy B.E., Sreedhar M. Acetylcholinesterase based biosensor for monitoring of malathion and acephate in food samples: A voltammetric study. Food Chem. 2014;142:188–196. doi: 10.1016/j.foodchem.2013.07.047. [DOI] [PubMed] [Google Scholar]
- 47.Ahmed M., Rocha J.B., Mazzanti C.M., Morsch A.L., Cargnelutti D., Correa M., Loro V., Morsch V.M., Schetinger M.R. Malathion, carbofuran and paraquat inhibit bungarus sindanus (krait) venom acetylcholinesterase and human serum butyrylcholinesterase in vitro. Ecotoxicology. 2007;16:363–369. doi: 10.1007/s10646-007-0137-1. [DOI] [PubMed] [Google Scholar]
- 48.Tian Y., Zhou S.F., Gao Y., Zhou Y.J., Shi R., Heiger-Bernays W., Ding Y., Wang P., Han S., Shen X.M. Effects of repeated maternal oral exposure to low levels of trichlorfon on development and cytogenetic toxicity in 3-day mouse embryos. Food Chem. Toxicol. 2011;49:2655–2659. doi: 10.1016/j.fct.2011.07.011. [DOI] [PubMed] [Google Scholar]
- 49.Becker R.E., Unni L.K., Greig N.H. Resurrecting clinical pharmacology as a context for alzheimer disease drug development. Curr. Alzheimer Res. 2009;6:79–81. doi: 10.2174/156720509787313916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.López-Arrieta J.M., Schneider L. Metrifonate for Alzheimer’s disease. Cochrane Database Syst. Rev. 2006:CD003155. doi: 10.1002/14651858.CD003155.pub3. [DOI] [PubMed] [Google Scholar]
- 51.Yang Y.X., Niu L.Z., Li S.N. Purification and studies on characteristics of cholinesterases from daphnia magna. J. Zhejiang Univ.-Sci. B. 2013;14:325–335. doi: 10.1631/jzus.B1200113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Oropesa A.L., Gravato C., Sanchez S., Soler F. Characterization of plasma cholinesterase from the white stork (ciconia ciconia) and its in vitro inhibition by anticholinesterase pesticides. Ecotoxicol. Environ. Saf. 2013;97:131–138. doi: 10.1016/j.ecoenv.2013.07.022. [DOI] [PubMed] [Google Scholar]
- 53.Zawadzka A., Lózinska I., Moleda Z., Panasiewicz M., Czarnocki Z. Highly selective inhibition of butyrylcholinesterase by a novel melatonin-tacrine heterodimers. J. Pineal Res. 2013;54:435–441. doi: 10.1111/jpi.12006. [DOI] [PubMed] [Google Scholar]
- 54.Bartolucci C., Stojan J., Yu Q.S., Greig N.H., Lamba D. Kinetics of torpedo californica acetylcholinesterase inhibition by bisnorcymserine and crystal structure of the complex with its leaving group. Biochem. J. 2012;444:269–277. doi: 10.1042/BJ20111675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Jokanovic M. Medical treatment of acute poisoning with organophosphorus and carbamate pesticides. Toxicol. Lett. 2009;190:107–115. doi: 10.1016/j.toxlet.2009.07.025. [DOI] [PubMed] [Google Scholar]
- 56.Ballatore C., Zhang B., Trojanowski J.Q., Lee V.M.Y., Smith A.B. In situ blood-brain barrier permeability of a c-10 paclitaxel carbamate. Bioorg. Med. Chem. Lett. 2008;18:6119–6121. doi: 10.1016/j.bmcl.2008.10.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Prades R., Guerrero S., Araya E., Molina C., Salas E., Zurita E., Selva J., Egea G., Lopez-Iglesias C., Teixido M., et al. Delivery of gold nanoparticles to the brain by conjugation with a peptide that recognizes the transferrin receptor. Biomaterials. 2012;33:7194–7205. doi: 10.1016/j.biomaterials.2012.06.063. [DOI] [PubMed] [Google Scholar]
- 58.Lee N.Y., Kang Y.S. The inhibitory effect of rivastigmine and galantamine on choline transport in brain capillary endothelial cells. Biomol. Ther. 2010;18:65–70. doi: 10.4062/biomolther.2010.18.1.065. [DOI] [Google Scholar]
- 59.Di Stefano A., Iannitelli A., Laserra S., Sozio P. Drug delivery strategies for Alzheimer’s disease treatment. Expert Opin. Drug Deliv. 2011;8:581–603. doi: 10.1517/17425247.2011.561311. [DOI] [PubMed] [Google Scholar]
- 60.Beilin B., Bessler H., Papismedov L., Weinstock M., Shavit Y. Continuous physostigmine combined with morphine-based patient-controlled analgesia in the postoperative period. Acta Anaesthesiol. Scand. 2005;49:78–84. doi: 10.1111/j.1399-6576.2004.00548.x. [DOI] [PubMed] [Google Scholar]
- 61.Arkypova V.N., Dzyadevych S.V., Soldatkin A.P., El’skaya A.V., Martelet C., Jaffrezic-Renault N. Development and optimisation of biosensors based on ph-sensitive field effect transistors and cholinesterases for sensitive detection of solanaceous glycoalkaloids. Biosens. Bioelectron. 2003;18:1047–1053. doi: 10.1016/S0956-5663(02)00222-1. [DOI] [PubMed] [Google Scholar]
- 62.Dzyadevich S.V., Arkhypova V.N., Soldatkin A.P., El’skaya A.V., Martelet C., Jaffrezic-Renault N. Enzyme biosensor for tomatine detection in tomatoes. Anal. Lett. 2004;37:1611–1624. doi: 10.1081/AL-120037591. [DOI] [Google Scholar]
- 63.Benilova I.V., Arkhypova V.N., Dzyadeviych S.V., Jaffrezic-Renault N., Martelet C., Soldatkin A.P. Kinetics of human and horse sera cholinesterases inhibition with solanaceous glycoalkaloids: Study by potentiometric biosensor. Pest. Biochem. Physiol. 2006;86:203–210. doi: 10.1016/j.pestbp.2006.04.002. [DOI] [Google Scholar]
- 64.Ingkaninan K., Phengpa P., Yuenyongsawad S., Khorana N. Acetylcholinesterase inhibitors from stephania venosa tuber. J. Pharm. Pharmacol. 2006;58:695–700. doi: 10.1211/jpp.58.5.0015. [DOI] [PubMed] [Google Scholar]
- 65.Xiao H.T., Peng J., Liang Y., Yang J., Bai X., Hao X.Y., Yang F.M., Sun Q.Y. Acetylcholinesterase inhibitors from corydalis yanhusuo. Nat. Prod. Res. 2011;25:1418–1422. doi: 10.1080/14786410802496911. [DOI] [PubMed] [Google Scholar]
- 66.Jann M.W., Shirley K.L., Small G.W. Clinical pharmacokinetics and pharmacodynamics of cholinesterase inhibitors. Clin. Pharmacokinet. 2002;41:719–739. doi: 10.2165/00003088-200241100-00003. [DOI] [PubMed] [Google Scholar]
- 67.Pohanka M. Acetylcholinesterase inhibitors: A patent review (2008-present) Expert Opin. Ther. Pat. 2012;22:871–886. doi: 10.1517/13543776.2012.701620. [DOI] [PubMed] [Google Scholar]
- 68.Alfirevic A., Mills T., Carr D., Barratt B.J., Jawaid A., Sherwood J., Smith J.C., Tugwood J., Hartkoorn R., Owen A., et al. Tacrine-induced liver damage: An analysis of 19 candidate genes. Pharmacogenet. Genomics. 2007;17:1091–1100. doi: 10.1097/FPC.0b013e3282f1f12b. [DOI] [PubMed] [Google Scholar]
- 69.Carr D.F., Alfirevic A., Tugwood J.D., Barratt B.J., Sherwood J., Smith J., Pirmohamed M., Park B.K. Molecular and genetic association of interleukin-6 in tacrine-induced hepatotoxicity. Pharmacogenet. Genomics. 2007;17:961–972. doi: 10.1097/FPC.0b013e3282f00919. [DOI] [PubMed] [Google Scholar]
- 70.Pathak M.K., Fareed M., Srivastava A.K., Pangtey B.S., Bihari V., Kuddus M., Kesavachandran C. Seasonal variations in cholinesterase activity, nerve conduction velocity and lung function among sprayers exposed to mixture of pesticides. Environ. Sci. Pollut. Res. 2013;20:7296–7300. doi: 10.1007/s11356-013-1743-5. [DOI] [PubMed] [Google Scholar]
- 71.Berg R.M.G., Ofek K., Qvist T., Tofteng F., Soreq H., Moller K. Cholinesterase modulations in patients with acute bacterial meningitis. Scand. J. Clin. Lab. Investig. 2011;71:350–352. doi: 10.3109/00365513.2011.558107. [DOI] [PubMed] [Google Scholar]
- 72.Sonali N., Tripathi M., Sagar R., Velpandian T., Subbiah V. Clinical effectiveness of rivastigmine monotherapy and combination therapy in Alzheimer’s patients. CNS Neurosci. Ther. 2013;19:91–97. doi: 10.1111/cns.12036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Bond M., Rogers G., Peters J., Anderson R., Hoyle M., Miners A., Moxham T., Davis S., Thokala P., Wailoo A., et al. The effectiveness and cost-effectiveness of donepezil, galantamine, rivastigmine and memantine for the treatment of Alzheimer’s disease (review of technology appraisal no. 111): A systematic review and economic model. Health Technol. Asses. 2012;16:1–470. doi: 10.3310/hta16210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Lilienfeld S. Galantamine—A novel cholinergic drug with a unique dual mode of action for the treatment of patients with Alzheimer’s disease. CNS Drug Rev. 2002;8:159–176. doi: 10.1111/j.1527-3458.2002.tb00221.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Bai D.L., Tang X.C., He X.C. Huperzine a, a potential therapeutic agent for treatment of Alzheimer’s disease. Curr. Med. Chem. 2000;7:355–374. doi: 10.2174/0929867003375281. [DOI] [PubMed] [Google Scholar]
- 76.Cheewakriengkrai L., Gauthier S. A 10-year perspective on donepezil. Expert Opin. Pharmacother. 2013;14:331–338. doi: 10.1517/14656566.2013.760543. [DOI] [PubMed] [Google Scholar]
- 77.Zhang H.C., Liang H., Kuang P.Q., Yuan Q.P., Wang Y. Simultaneously preparative purification of huperzine a and huperzine b from huperzia serrata by macroporous resin and preparative high performance liquid chromatography. J. Chromatogr. B. 2012;904:65–72. doi: 10.1016/j.jchromb.2012.07.019. [DOI] [PubMed] [Google Scholar]
- 78.Zhang J.M., Hu G.Y. Huperzine a, a nootropic alkaloid, inhibits n-methyl-d-aspartate-induced current in rat dissociated hippocampal neurons. Neuroscience. 2001;105:663–669. doi: 10.1016/S0306-4522(01)00206-8. [DOI] [PubMed] [Google Scholar]
- 79.Zhang Y.H., Zhao X.Y., Chen X.Q., Wang Y., Yang H.H., Hu G.Y. Spermidine antagonizes the inhibitory effect of huperzine a on h-3 dizocilpine (mk-801) binding in synaptic membrane of rat cerebral cortex. Neurosci. Lett. 2002;319:107–110. doi: 10.1016/S0304-3940(01)02565-4. [DOI] [PubMed] [Google Scholar]
- 80.Heinrich M., Teoh H.L. Galanthamine from snowdrop—The development of a modern drug against Alzheimer’s disease from local caucasian knowledge. J. Ethnopharmacol. 2004;92:147–162. doi: 10.1016/j.jep.2004.02.012. [DOI] [PubMed] [Google Scholar]
- 81.Rainer M. Galanthamine in Alzheimer’s disease-A new alternative to tacrine? CNS Drugs. 1997;7:89–97. doi: 10.2165/00023210-199707020-00001. [DOI] [PubMed] [Google Scholar]
- 82.Pavlov V.A., Ochani M., Parrish W.R., Rosas-Ballina M., Ochani K., Al-Abed Y., Tracey K.J. The anti-inflammatory efficacy of galantamine is dependent on the integrity of the cholinergic anti-inflammatory pathway. Shock. 2007;27:23. [Google Scholar]
- 83.Takata K., Kitamura Y., Saeki M., Terada M., Kagitani S., Kitamura R., Fujikawa Y., Maelicke A., Tomimoto H., Taniguchi T., et al. Galantamine-induced amyloid-beta clearance mediated via stimulation of microglial nicotinic acetylcholine receptors. J. Biol. Chem. 2010;285:40180–40191. doi: 10.1074/jbc.M110.142356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Liu Z.H., Ma Y.F., Wu J.S., Gan J.X., Xu S.W., Jiang G.Y. Effect of cholinesterase inhibitor galanthamine on circulating tumor necrosis factor alpha in rats with lipopolysaccharide-induced peritonitis. Chin. Med. J. 2010;123:1727–1730. [PubMed] [Google Scholar]
- 85.Hwang J., Hwang H., Lee H.W., Suk K. Microglia signaling as a target of donepezil. Neuropharmacology. 2010;58:1122–1129. doi: 10.1016/j.neuropharm.2010.02.003. [DOI] [PubMed] [Google Scholar]
- 86.Sochocka M., Zaczynska E., Leszek J., Siemieniec W., Blach-Olszewska Z. Effect of donepezil on innate antiviral immunity of human leukocytes. J. Neurol. Sci. 2008;273:75–80. doi: 10.1016/j.jns.2008.06.021. [DOI] [PubMed] [Google Scholar]
- 87.Sochocka M., Zaczynska E., Tabol A., Czarny A., Leszek J., Sobczynski M. The influence of donepezil and egb 761 on the innate immunity of human leukocytes effect on the NF-κB system. Int. Immunopharmacol. 2010;10:1505–1513. doi: 10.1016/j.intimp.2010.08.024. [DOI] [PubMed] [Google Scholar]
- 88.Tang X.C., Kindel G.H., Kozikowski A.P., Hanin I. Comparison of the effects of natural and synthetic huperzine-a on rat brain cholinergic function in vitro and in vivo. J. Enthnopharmacol. 1994;44:147–155. doi: 10.1016/0378-8741(94)01182-6. [DOI] [PubMed] [Google Scholar]
- 89.White J.D., Li Y., Kim J., Terinek M. A novel synthesis of (−)-huperzine a via tandem intramolecular aza-prins cyclization-cyclobutane fragmentation. Org. Lett. 2013;15:882–885. doi: 10.1021/ol400012s. [DOI] [PubMed] [Google Scholar]
- 90.Ma X., Gang D.R. In vitro production of huperzine a, a promising drug candidate for Alzheimer’s disease. Phytochemistry. 2008;69:2022–2028. doi: 10.1016/j.phytochem.2008.04.017. [DOI] [PubMed] [Google Scholar]
- 91.Leman L., Kitson S.L., Brown R.T., Cairns J., Watters W., McMordie A., Murrell V.L., Marfurt J. Synthesis of isotopically labelled c-14 zt-1 (debio-9902), d(3) zt-1 and (−)-d(3) huperzinea, a new generation of acetylcholinesterase inhibitors. J. Labelledc Compd. Rad. 2011;54:720–730. doi: 10.1002/jlcr.1918. [DOI] [Google Scholar]
- 92.Hedberg M.M., Clos M.V., Ratia M., Gonzalez D., Lithner C.U., Camps P., Munoz-Torrero D., Badia A., Gimenez-Llort L., Nordberg A. Effect of huprine x on beta-amyloid, synaptophysin and alpha 7 neuronal nicotinic acetylcholine receptors in the brain of 3xtg-ad and appswe transgenic mice. Neurodegener. Dis. 2010;7:379–388. doi: 10.1159/000287954. [DOI] [PubMed] [Google Scholar]
- 93.Wang J., Chen F., Zheng P., Deng W.J., Yuan J., Peng B., Wang R.C., Liu W.J., Zhao H., Wang Y.Q., et al. Huperzine a ameliorates experimental autoimmune encephalomyelitis via the suppression of t cell-mediated neuronal inflammation in mice. Exp. Neurol. 2012;236:79–87. doi: 10.1016/j.expneurol.2012.03.024. [DOI] [PubMed] [Google Scholar]
- 94.Wang Z.F., Tang X.C. Huperzine a protects c6 rat glioma cells against oxygen-glucose deprivation-induced injury. FEBS Lett. 2007;581:596–602. doi: 10.1016/j.febslet.2007.01.016. [DOI] [PubMed] [Google Scholar]
- 95.Wang Z.F., Wang J., Zhang H.Y., Tang X.C. Huperzine a exhibits anti-inflammatory and neuroprotective effects in a rat model of transient focal cerebral ischemia. J. Neurochem. 2008;106:1594–1603. doi: 10.1111/j.1471-4159.2008.05504.x. [DOI] [PubMed] [Google Scholar]
- 96.Darreh-Shori T., Soininen H. Effects of cholinesterase inhibitors on the activities and protein levels of cholinesterases in the cerebrospinal fluid of patients with Alzheimer’s disease: A review of recent clinical studies. Curr. Alzheimer Res. 2010;7:67–73. doi: 10.2174/156720510790274455. [DOI] [PubMed] [Google Scholar]
- 97.Darreh-Shori T., Hellstrom-Lindahl E., Flores-Flores C., Guan Z.Z., Soreq H., Nordberg A. Long-lasting acetylcholinesterase splice variations in anticholinesterase-treated Alzheimer’s disease patients. J. Neurochem. 2004;88:1102–1113. doi: 10.1046/j.1471-4159.2003.02230.x. [DOI] [PubMed] [Google Scholar]
- 98.Bastiat G., Plourde F., Motulsky A., Furtos A., Dumont Y., Quirion R., Fuhrmann G., Leroux J.C. Tyrosine-based rivastigmine-loaded organogels in the treatment of Alzheimer’s disease. Biomaterials. 2010;31:6031–6038. doi: 10.1016/j.biomaterials.2010.04.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Field R.H., Gossen A., Cunningham C. Prior pathology in the basal forebrain cholinergic system predisposes to inflammation-induced working memory deficits: Reconciling inflammatory and cholinergic hypotheses of delirium. J. Neurosci. 2012;32:6288–6294. doi: 10.1523/JNEUROSCI.4673-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Nizri E., Irony-Tur-Sinai M., Faranesh N., Lavon I., Lavi E., Weinstock M., Brenner T. Suppression of neuroinflammation and immunomodulation by the acetylcholinesterase inhibitor rivastigmine. J. Neuroimmunol. 2008;203:12–22. doi: 10.1016/j.jneuroim.2008.06.018. [DOI] [PubMed] [Google Scholar]
- 101.Chiou W.H., Kao C.L., Tsai J.C., Chang Y.M. Domino rh-catalyzed hydroformylation-double cyclization of o-amino cinnamyl derivatives: Applications to the formal total syntheses of physostigmine and physovenine. Chem. Commun. 2013;49:8232–8234. doi: 10.1039/c3cc43257b. [DOI] [PubMed] [Google Scholar]
- 102.Realini T. A history of glaucoma pharmacology. Optom. Vis. Sci. 2011;88:36–38. doi: 10.1097/OPX.0b013e3182058ead. [DOI] [PubMed] [Google Scholar]
- 103.Greig N.H., Becker R.E., Yu Q., Holloway H.W., Tweedie D., Luo W., Utsuki T., Ingram D.K., Maccecchini M.L., Rogers J.T., et al. From natural products to alzheimer experimental therapeutics, eserine based drugs as symptomatic and disease altering agents. Prog. Nutr. 2010;12:58–63. [Google Scholar]
- 104.Russo P., Frustaci A., del Bufalo A., Fini M., Cesario A. Multitarget drugs of plants origin acting on Alzheimer’s disease. Curr. Med. Chem. 2013;20:1686–1693. doi: 10.2174/0929867311320130008. [DOI] [PubMed] [Google Scholar]
- 105.Bitzinger D.I., Zausig Y.A., Paech C., Gruber M., Busse H., Sinner B., Graf B.M., Trabold B. Modulation of immune functions in polymorphonuclear neutrophils induced by physostigmine, but not neostigmine, independent of cholinergic neurons. Immunobiology. 2013;218:1049–1054. doi: 10.1016/j.imbio.2013.01.003. [DOI] [PubMed] [Google Scholar]
- 106.Peter C., Schmidt K., Hofer S., Stephan M., Martin E., Weigand M.A., Walther A. Effects of physostigmine on microcirculatory alterations during experimental endotoxemia. Shock. 2010;33:405–411. doi: 10.1097/SHK.0b013e3181b77e82. [DOI] [PubMed] [Google Scholar]
- 107.Kutsuna S., Tsuruta R., Fujita M., Todani M., Yagi T., Ogino Y., Igarashi M., Takahashi K., Izumi T., Kasaoka S., et al. Cholinergic agonist physostigmine suppresses excessivesuperoxide anion radical generation in blood, oxidative stress, early inflammation, and endothelial injury in rats with forebrain ischemia/reperfusion. Brain Res. 2010;1313:242–249. doi: 10.1016/j.brainres.2009.11.077. [DOI] [PubMed] [Google Scholar]
- 108.Benatar M., Kaminski H. Medical and surgical treatment for ocular myasthenia. Cochrane Database Syst. Rev. 2012;12:CD005081. doi: 10.1002/14651858.CD005081.pub3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Chambers D., Paulden M., Paton F., Heirs M., Duffy S., Craig D., Hunter J., Wilson J., Sculpher M., Woolacott N. Sugammadex for the reversal of muscle relaxation in general anaesthesia: A systematic review and economic assessment. Health Technol. Assess. 2010;14:1–211. doi: 10.3310/hta14390. [DOI] [PubMed] [Google Scholar]
- 110.Cossins J., Belaya K., Zoltowska K., Koneczny I., Maxwell S., Jacobson L., Leite M.I., Waters P., Vincent A., Beeson D. The search for new antigenic targets in myasthenia gravis. Ann. N. Y. Acad. Sci. 2012;1275:123–128. doi: 10.1111/j.1749-6632.2012.06833.x. [DOI] [PubMed] [Google Scholar]
- 111.Vyskocil F., Malomouzh A.I., Nikolsky E.E. Non-quantal acetylcholine release at the neuromuscular junction. Physiol. Res. 2009;58:763–784. doi: 10.33549/physiolres.931865. [DOI] [PubMed] [Google Scholar]
- 112.Walker S.M., Yaksh T.L. Neuraxial analgesia in neonates and infants: A review of clinical and preclinical strategies for the development of safety and efficacy data. Anesth. Analg. 2012;115:638–662. doi: 10.1213/ANE.0b013e31826253f2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Fisher D.M. Clinical pharmacology of neuromuscular blocking agents. Am. J. Health Syst. Pharm. 1999;56:S4–S9. doi: 10.1093/ajhp/56.S4. [DOI] [PubMed] [Google Scholar]
- 114.Yang L.P., Keam S.J. Sugammadex: A review of its use in anaesthetic practice. Drugs. 2009;69:919–942. doi: 10.2165/00003495-200969070-00008. [DOI] [PubMed] [Google Scholar]
- 115.Friedman A., Kaufer D., Shemer J., Hendler I., Soreq H., TurKaspa I. Pyridostigmine brain penetration under stress enhances neuronal excitability and induces early immediate transcriptional response. Nat. Med. 1996;2:1382–1385. doi: 10.1038/nm1296-1382. [DOI] [PubMed] [Google Scholar]
- 116.Kalb A., von Haefen C., Sifringer M., Tegethoff A., Paeschke N., Kostova M., Feldheiser A., Spies C.D. Acetylcholinesterase inhibitors reduce neuroinflammation and -degeneration in the cortex and hippocampus of a surgery stress rat model. PLoS One. 2013;8:e62679. doi: 10.1371/journal.pone.0062679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Kalb A., von Haefen C., Sifringer M., Tegethoff A., Paeschke N., Kostova M., Feldheiser A., Spies C.D. Physostigmine and neostigmine reduce the increased expression of il-1beta in the hippocampus and cortex after surgery combined with lps-treatment. Eur. J. Neurol. 2012;19:756. [Google Scholar]
- 118.Pohanka M., Pavlis O. Neostigmine modulates tularemia progression in balb/c mice. Afr. J. Pharm. Pharmacol. 2012;6:1317–1322. [Google Scholar]
- 119.Sun L., Zhang G.F., Zhang X., Liu Q., Liu J.G., Su D.F., Liu C. Combined administration of anisodamine and neostigmine produces anti-shock effects: Involvement of alpha 7 nicotinic acetylcholine receptors. Acta Pharm. Sin. 2012;33:761–766. doi: 10.1038/aps.2012.26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Kox M., Pompe J.C., Peters E., Vaneker M., van der Laak J.W., van der Hoeven J.G., Scheffer G.J., Hoedemaekers C.W., Pickkers P. Alpha 7 nicotinic acetylcholine receptor agonist gts-21 attenuates ventilator-induced tumour necrosis factor-alpha production and lung injury. Br. J. Anaesth. 2011;107:559–566. doi: 10.1093/bja/aer202. [DOI] [PubMed] [Google Scholar]