

Abstract
Purpose
Programmed death ligand 1 (PD-L1) is an immunomodulatory molecule expressed by antigen-presenting cells and select tumors that engage receptors on T cells to inhibit T-cell immunity. Immunotherapies targeting the PD-1/PD-L1 pathway have shown durable anti-tumor effects in a subset of patients with solid tumors. PD-L1 can be expressed by Reed-Sternberg cells comprising classical Hodgkin lymphoma (CHLs) and by malignant B cells comprising EBV-positive post-transplant lymphoproliferative disorders (PTLDs). We sought to determine whether the expression of PD-L1 represents a general strategy of immune evasion among aggressive B-cell lymphomas and virus- and immunodeficiency-associated tumors.
Experimental Design
Using novel antibodies and formalin-fixed, paraffin-embedded (FFPE) tissue biopsies, we examined 237 primary tumors for expression of PD-L1 protein.
Results
Robust PD-L1 protein expression was found in the majority of nodular sclerosis CHL, mixed cellularity CHL, primary mediastinal large B-cell lymphoma, T-cell/histiocyte-rich B-cell lymphoma, EBV-positive and -negative PTLD, and EBV-associated diffuse large B-cell lymphoma (DLBCL), plasmablastic lymphoma, extranodal NK/T cell lymphoma, nasopharyngeal carcinoma, and HHV8-associated primary effusion lymphoma. Within these tumors, PD-L1 was highly expressed by malignant cells and tumor-infiltrating macrophages. In contrast, neither the malignant nor the non-malignant cells comprising nodular lymphocyte-predominant Hodgkin lymphoma, DLBCL-not otherwise specified, Burkitt lymphoma, and HHV8-associated Kaposi sarcoma expressed detectable PD-L1.
Conclusion
Certain aggressive B-cell lymphomas and virus- and immunodeficiency-associated malignancies associated with an ineffective T-cell immune response express PD-L1 on tumor cells and infiltrating macrophages. These results identify a group of neoplasms that should be considered for PD-1/PD-L1-directed therapies, and validate a method to detect PD-L1 in FFPE tissue biopsies.
Introduction
Programmed cell death ligand 1 (PD-L1, also known as B7-H1) is an immunomodulatory cell-surface glycoprotein and a member of the B7 family of costimulatory molecules primarily expressed by antigen-presenting cells and that serve to regulate the cellular immune response.1 Binding of PD-L1 to its cognate receptor PD-1 inhibits proliferation of activated T cells in peripheral tissues leading to “T-cell exhaustion,” a functional phenotype that can be reversed by PD-1 blockade.2 In clinical trials of anti-PD-1 and anti-PD-L1 antibodies, patients with solid tumors have achieved long-lasting clinical responses and thus emphasizing the importance of the PD-1/PD-L1 axis in regulating anti-tumor immunity.3,4 Moreover, in the limited number of cases examined, clinical responsiveness to PD-1 blockade correlated with tumor-cell specific expression of PD-L1 as detected by immunohistochemistry (IHC) using a proprietary antibody.4
We have shown that Hodgkin lymphoma cell lines express high levels of PD-L1 transcript and protein due to multiple mechanisms including amplification of chromosome 9p24.1 and constitutive AP1 signaling. The copy number of 9p24.1, a genomic region that includes CD274 (encoding PD-L1), PDCD1LG2 (encoding PD-L2), and JAK2, correlates with cell surface PD-L1 protein expression as a result of direct amplification of CD274, and due to increased JAK2 protein expression, enhanced JAK-STAT signaling, and activation of an interferon (IFN)-stimulated regulatory element/IFN-regulatory factor 1 (ISRE/IRF1) motif in the CD274 promoter in cell lines.5,6 In a limited series of genetically annotated primary classical Hodgkin lymphoma (CHL) cases, we confirmed that high copy numbers of 9p24.1 correlated with increased PD-L1 expression in Reed-Sternberg (RS) cells.5 We have further shown that: 1) PD-L1 expression is regulated by an AP1-dependent enhancer in CD274; 2) AP1 signaling components JunB/cJun constitutively bind this enhancer in Hodgkin cell lines; and 3) JunB and cJun are overexpressed in the RS cells of primary CHL.5
Moreover, a subset of CHL is Epstein-Barr virus (EBV)-positive and aberrant signaling through EBV-encoded gene products provides alternative mechanisms to upregulate PD-L1. Using EBV-transformed B cells, we demonstrated that the expression of the EBV-encoded latent membrane protein (LMP)-1 promotes both AP1-signaling and JAK-STAT signaling to activate the enhancer and promoter elements of CD274, respectively.6 Consistent with these findings, we have detected PD-L1 in the majority of examined EBV-positive post-transplant lymphoproliferative disorders (PTLD).6 Thus, CHL and EBV-positive PTLD can utilize several complementary mechanisms to upregulate PD-L1 in malignant tumor cells.
CHL is also notable for its prominent inflammatory infiltrate which includes a skewed T-cell population and abundant macrophages. In CHL, increased numbers of CD68+ tumor-associated macrophages have been associated with inferior clinical outcome.8,9 However, the biological basis for this association remains undefined.
CHL is genetically, phenotypically, and/or histologically related to additional large cell lymphomas including primary mediastinal large B-cell lymphoma (PMLBCL), nodular lymphocyte predominant Hodgkin lymphoma (NLPHL), and T-cell rich, histiocyte-rich large B-cell lymphoma (TCRLBCL).10,11 Similarly, EBV and the closely related virus human herpes virus-8 (HHV8, also known as Kaposi sarcoma-associated herpesvirus) are postulated or known drivers of additional malignancies that include HIV/immunodeficiency-related diffuse large B-cell lymphoma (DLBCL), EBV-associated DLBCL of the elderly, extranodal NK/T cell lymphoma (ENKTCL), nasopharyngeal carcinoma (NPC), plasmablastic lymphoma (PBL), endemic Burkitt lymphoma (BL), and the HHV8-associated primary effusion lymphoma (PEL) and Kaposi sarcoma (KS).12,13 A comprehensive analysis of PD-L1 expression in these tumor types has not been completed.
In this study, we validate two novel anti-PD-L1 antibodies for the detection of PD-L1 protein in formalin-fixed, paraffin-embedded (FFPE) tissue sections. We then evaluate PD-L1 expression in a range of aggressive lymphoid malignancies, including those related to CHL and those associated with EBV and/or HHV8 infection, as well as NPC and KS. We find that robust expression of PD-L1 is characteristic of the malignant cells and a large proportion of non-malignant cells, primarily tumor infiltrating macrophages, within the tumor microenvironment of CHL, PMLBCL, TCRLBCL, and a broad range of virus- and immunodeficiency-associated malignancies that are characterized by an ineffective immune response and suggest a biological basis for the prognostic significance of tumor-infiltrating macrophages in CHL. Finally, our methods establish a template for screening FFPE tumor biopsies for PD-L1 expression by IHC using a generally available monoclonal antibody.
Materials and Methods
Case Selection
Cases were retrieved from the surgical pathology files of Brigham and Women's Hospital, Boston, MA; Yale School of Medicine, New Haven CT; UMass Memorial Medical Center, Worcester, MA; and from the consult files of one of the authors (C.D.M.F.) with institutional review boards' approvals. All tumors were diagnosed and classified according to standard WHO criteria. Representative hematoxylin and eosin stained slides were reviewed to confirm the presence of adequate amounts of tumor. Cases included whole tissue sections and tissue microarrays. Cases included nodular sclerosis CHL (NSCHL, 25 cases); mixed-cellularity CHL (MCCHL, 8 cases); CHL, not otherwise specified (CHL-NOS, 5 cases); nodular lymphocyte-predominant Hodgkin lymphoma (NLPHL, 15 cases); primary mediastinal large B-cell lymphoma (PMBCL, 21 cases); T-cell/histiocyte-rich large B-cell lymphoma (TCHRBCL, 11 cases); EBV-positive DLBCL of the elderly (9 cases); EBV-positive immunodeficiency-related DLBCL (HIV-associated and iatrogenic; excluding patients with a history of solid organ transplantation, 7 cases); EBV-positive PTLD (10 cases); EBV-negative PTLD (7 cases); DLBCL, not otherwise specified (DLBCL-NOS, 66 cases); PBL (9 cases); PEL (4 cases); ENKTCL (6 cases); EBV-positive BL (7 cases); NPC (18 cases); KS (9 cases). All cases of EBV-positive DLBCL, EBV-positive PTLD, EBV-positive BL, NPC, 4 of 6 cases of ENKTCL, and 7 of 9 cases of PBL were shown previously to be positive for EBV-encoded RNA (EBER) by in situ hybridization study. All EBV-negative PTLD and DLBCL-NOS cases were negative for EBER. All PEL and KS cases were shown previously to be positive for HHV8 by immunohistochemistry.
Cell culture and Western blot
Hodgkin lymphoma cell lines (L428, SUP-HD1, and HDLM2) and DLBCL cell lines (SUDHL4 and OCI-Ly1) were cultured as previously described.14 Preparation of cell lysates, SDS-PAGE, and Western blot were performed as previously described14 with the following antibody conditions: PD-L1 was detected with a rabbit monoclonal antibody (#10084-R015, 1:1500, 1 ug/ml final concentration, Sino Biological, Beijing, China) in 3% non-fat dry milk (sc-2325, Santa Cruz Biotechnologies, Santa Cruz, CA). Blots were stripped and reprobed with anti-GAPDH antibody (FL-335, sc-25778, 1:2000, Santa Cruz) in 3% non-fat dry milk.
Immunohistochemistry
IHC using a rabbit anti-PD-L1 monoclonal antibody (clone 15, #10084-R015, 6.2ug/ml final concentration, Sino Biological, Beijing, China) was performed using 4-μm-thick, FFPE tissue sections on a Benchmark XT autostainer (Ventana Medical System, Tuscon, AZ) with standard antigen retrieval methods (CC1 buffer, pH8.0, #950-124, Ventana). UltraView Universal DAB Detection kit (#760-500, Ventana) was used according to the manufacturer's instructions. Counterstaining was performed as part of the automated staining protocol using hematoxylin (#760-2021, Ventana). IHC using the mouse anti-PD-L1 monoclonal antibody (IgG1, generated in the laboratory of G. Freeman, clone 339.7G11, 69 ug/ml final concentration) was performed using the same protocol as above. After staining, slides were then washed in soap water and distilled water, dehydrated in graded alcohol and xylene, mounted and coverslipped.
Double IHC staining for CD68 and PD-L1 was performed on selected cases within the cohort. PD-L1 staining using the rabbit anti-PD-L1 monoclonal antibody was performed first using the same protocol as above, except counterstaining was omitted at this stage. The slides were taken off the autostainer and washed in soap water and distilled water, and antigen retrieval was performed with 10mM citrate buffer, pH6.0 (Invitrogen, Carlsbad, CA) in a steam pressure cooker (Decloaking Chamber, Biocare Medical, Concord, CA) according to the manufacturer's instruction. All further steps were carried out at room temperature in a hydrated chamber. Slides were incubated with monoclonal mouse anti-human CD68 (clone PG-M1, #M0876, 0.3 μg/ml final concentration, Dako) in diluent (Dako) for 1 hour. Slides were then washed in Tris buffer and treated with MACH2 Mouse AP-Polymer (#MALP521L, Biocare Medical) for 30 minutes. After further washing, slides were incubated with Vulcan Fast Red Chromogen (#FR805H, Biocare Medical) for 20 minutes. Slides were counterstained with hematoxylin, washed in distilled water, and dehydrated in 60°C oven for 20 minutes, and briefly washed in xylene, mounted and coverslipped.
Double IHC staining for PAX5 (BSAP) and PD-L1 was performed on selected cases within the cohort. PD-L1 staining using the mouse anti-PD-L1 monoclonal antibody was performed first using the same protocol described above, except counterstaining was omitted at this stage. The slides were taken off the autostainer and washed in soap water and distilled water, and antigen retrieval was performed with 1mM EDTA, pH8.0 (Invitrogen) in a steam pressure cooker. Slides were incubated with mouse anti-BSAP/PAX5 monoclonal antibody (clone 24/Pax-5, #610863, 2.5 μg/ml final concentration, BD Transduction Laboratories, Franklin Lakes, NJ) in diluent (Dako, Carpinteria, CA) for 1 hour. Slides were then washed in Tris buffer and treated with MACH2 Mouse AP-Polymer for 30 minutes, followed by Vulcan Fast Red Chromogen for 20 minutes. Slides were counterstained with hematoxylin, washed in distilled water, and dehydrated in 60°C oven for 20 minutes, and briefly washed in xylene, mounted and coverslipped.
Case Evaluation
IHC stained sections were evaluated by two hematopathologists (BJC and SJR) with concurrence. The percentage of tumor cells present in tissue sections was estimated for each case. The percentage of the total cellularity staining for PD-L1, including malignant and non-malignant cells, and the percentage of the tumor cell population staining for PD-L1 were scored independently. Staining intensity was scored as follows: 0 (no staining), 1+ (weak or equivocal staining), 2+ (moderate staining), or 3+ (strong staining). Cases with membrane staining, with or without cytoplasmic staining, and cases with only cytoplasmic staining were tabulated separately. Cytoplasmic cellular staining had to unequivocally exceed background to be considered positive. Tumor staining for PD-L1 was considered positive if ≥5% of the tumor cell population showed 2+ or 3+ membrane staining. A case was considered to have a microenvironment positive for PD-L1 if ≥20% of the total tissue cellularity showed 2+ or 3+ membrane or cytoplasmic staining in malignant and/or non-malignant cells. The threshold for positive PD-L1 staining in malignant cells used here is comparable to that used in a prior publication, using a distinct antibody.4,15,16 Appropriate external positive (placenta) and negative (tonsil) controls were also included with each staining run.
Results
Validation of PD-L1 antibodies for IHC
After testing a variety of commercially available antibodies, we identified a rabbit monoclonal antibody that demonstrated both sensitive and specific staining for PD-L1 on a set of well-characterized cell lines and tissues (Figure 1, Materials and Methods). In addition, we validated a novel mouse monoclonal antibody (clone 339.7G11, Materials and Methods) recognizing PD-L1 that was generated by one of the co-authors on this study (GF, data not shown). By western blot analysis (Figure 1A), the rabbit anti-PD-L1 monoclonal antibody recognized a protein of ∼55 kDa—the size of glycosylated PD-L1—in lysates derived from Hodgkin cell lines (L428, SUP-HD1, HDLM2). The relative amount of PD-L1 in the Hodgkin cell lines as detected by western blot analysis was consistent with previously reported levels of PD-L1 expression detected by flow cytometry, and also correlated with chromosome 9p24 copy number status.5 Importantly, no band was detected in lysates derived from DLBCL cell lines (SUDHL4, OCI-Ly1), consistent with the previously reported lack of detectable PD-L1 in these lines (Figure 1A).5 Similarly, IHC analysis of FFPE cell pellets using the rabbit monoclonal antibody revealed robust staining of the Hodgkin cell line HDLM2 in a strong membranous pattern (Figure 1B) but not in the DLBCL line SUDHL4 (Figure 1C). The staining pattern was identical using the mouse monoclonal antibody (Figures 1B and 1C insets). Additional IHC analysis of transfected cell lines showed specific staining by both rabbit and mouse PD-L1 antibodies of cell lines expressing human PD-L1 but not human PD-L2 (data not shown). We conclude that IHC analysis using both the rabbit and mouse monoclonal antibodies is specific for PD-L1 protein.
Figure 1.
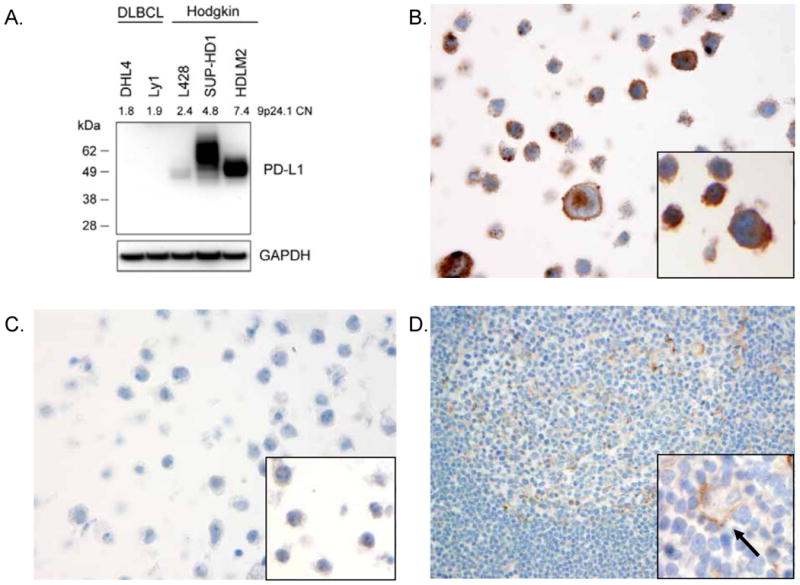
PD-L1 antibody validation. (A) Western blot analysis of cell lysates derived from genetically characterized diffuse large B-cell lymphoma (DLBCL) cell lines (DHL4, Ly1) and the Hodgkin lymphoma cell lines (L428, SUP-HD1, HDLM2). Lysates were probed with a rabbit monoclonal antibody recognizing PD-L1 (top panel) showing a band at the expected size of the fully glycosylated form of PD-L1 (∼55 kDa). The gene copy number (CN) for the CD274 (PD-L1) locus, 9p24.1, is shown, as reported previously, for each cell line.5 Equal loading was demonstrated by probing for GAPDH. IHC analysis of formalin-fixed, paraffin embedded (FFPE) Hodgkin cell line HDLM2 (B) and DLBCL cell line DHL4 (C) stained with the rabbit anti-PD-L1 antibody showing membranous staining of the HDLM2 cells but no staining of DHL4 cells. Insets show similar staining patterns with the mouse monoclonal anti-PD-L1 antibody. (D) FFPE human tonsil stained with rabbit anti-PD-L1 antibody showing little staining of lymphoid cells and weak membranous staining of occasional macrophages (inset, arrow).
IHC analysis of human tonsil revealed little to no specific staining of the vast majority of lymphocytes within the interfollicular T cell-rich regions and in the B cell-rich secondary follicles using the rabbit monoclonal PD-L1 antibody (Figure 1D). Rare lymphoid cells within reactive germinal centers did show weak membranous staining. High-power examination of human tonsil also revealed distinct membranous staining of the tonsilar epithelium and weak, membranous staining of scattered cells morphologically consistent with macrophages (Figure 1D inset).1,16 IHC analysis of tonsil using the mouse monoclonal PD-L1 antibody resulted in identical findings (Supplemental Figure S1). As an external positive control, syncytiotrophoblasts in human placental tissue demonstrated strong membranous expression of PD-L1, as described previously (Supplemental Figure S1).1
We noted that with the rabbit monoclonal antibody, a mild degree of general, nonspecific background staining was observed in tissue sections. Background staining varied according to the degree of dilution necessary to achieve the final antibody concentration optimized for IHC (Supplemental Figure S1). Less background was associated with the more concentrated lots of antibody (6.2 ug/ml final concentration used; lot concentrations ranged from 0.22 to 1.55 mg/ml). Very little background was noted with the mouse monoclonal antibody (Supplemental Figure S1; S2).
Expression of PD-L1 in HL and variants
Upon validation of two novel antibodies directed against PD-L1, we next surveyed a large cohort of Hodgkin lymphomas, including NSCHL, MCCHL, CHL-NOS, and NLPHL for PD-L1 expression by IHC. Tissue sections were comprised of a varying amount of RS cells. The median estimated percent malignant cells was 5% (range 2-20%) for NSCHL, 2% (range 2-10%) for MCCHL, 50% (range 2-90%) for CHL-NOS, and 2% (range 2-5%) for NLPHL. Twenty-one of 25 (84%) cases of NSCHL, 7/8 (88%) cases of MCCHL, and 5/5 (100%) cases of CHL-NOS showed strong (2+ or 3+) membranous staining of the RS cells (Figure 2A, Table 1, Supplemental Table S1). Of the 33 cases of CHL showing positive staining in malignant cells, 32 cases (97%) had at least 50% of the RS cells positive for PD-L1 and 27 cases (82%) had at least 90% of the RS cells positive for PD-L1 (Supplemental Table S1, Figure 3). By contrast, only two cases of NLPHL (13%) were scored as positive for PD-L1 with only 5% of the LP cells positive in each case (Figure 2B and 3, Supplemental Table S1).
Figure 2.
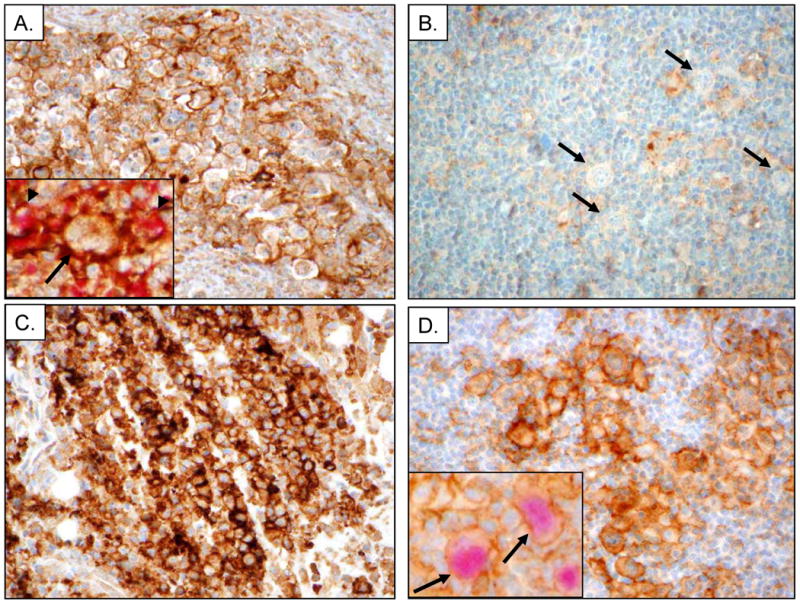
Immunohistochemical analysis of PD-L1 in CHL, NLPHL, PMLBCL and TCRLBCL. (A) Representative example nodular sclerosis classical Hodgkin lymphoma (NSCHL) stained with the rabbit anti-PD-L1 antibody showing distinct membranous staining of Reed-Sternberg (RS) cells and intra-tumoral macrophages. Inset shows staining with PD-L1 (brown) highlighting the cell membrane of RS cells (arrow) as well as non-malignant cells, and macrophages (arrowheads) double stained with PD-L1 and the macrophage marker CD68 (red). (B) Representative case of nodular lymphocyte-predominant Hodgkin lymphoma (NLPHL) stained with rabbit anti-PD-L1 showing LP cells (arrows) that are negative for PD-L1. (C) Representative case of primary mediastinal large B-cell lymphoma (PMBCL) stained with rabbit anti-PD-L1 antibody showing predominantly membranous staining of lymphoma cells. (D) Representative case of T-cell/histiocyte-rich large B-cell lymphoma (TCHRBCL) stained with rabbit anti-PD-L1 antibody. Inset shows staining with PD-L1 (brown) highlighting macrophages adjacent to scattered malignant cells (arrow) that are double stained for PD-L1 and the B-cell lineage marker PAX5 (red).
Table 1.
| Diagnosis | Cases N |
Cases with ≥5% malignant cells positive* N (%) |
Cases with ≥20% total cellularity positive** N (%) |
|---|---|---|---|
| Classical Hodgkin lymphoma (CHL) | 38 | 33 (87) | 31 (82) |
| Nodular sclerosis CHL | 25 | 21 (84) | 19 (76) |
| Mixed cellularity CHL | 8 | 7 (88) | 7 (88) |
| CHL-not otherwise specified (NOS) | 5 | 5 (100) | 5 (100) |
| Nodular lymphocyte predominant Hodgkin lymphoma | 15 | 2 (13) | 1 (7) |
| Primary mediastinal large B-cell lymphoma | 21 | 15 (71) | 19 (90) |
| T-cell/Histiocyte-rich large B-cell lymphoma | 11 | 10 (91) | 11 (100) |
| EBV+ diffuse large B-cell lymphoma (DLBCL) | 16 | 16 (100) | 16 (100) |
| EBV+ DLBCL of elderly | 9 | 9 (100) | 9 (100) |
| EBV+ immunodeficiency associated DLBCL | 7 | 7 (100) | 7 (100) |
| EBV+ posttransplant lymphoproliferative disorder (PTLD) | 10 | 6 (60) | 7 (70) |
| EBV-negative PTLD | 7 | 4 (57) | 4 (57) |
| DLBCL-NOS | 66 | 7 (11) | 9 (14) |
| Plasmablastic lymphoma | 9 | 4 (44) | 4 (44) |
| Primary effusion lymphoma | 4 | 2 (50) | 3 (75) |
| Extranodal NK/T-cell lymphoma | 6 | 4(67) | 5(83) |
| EBV+ Burkitt lymphoma | 7 | 0 (0) | 0 (0) |
| EBV+ Nasopharyngeal carcinoma | 18 | 16 (89) | nd |
| HHV8+ Kaposi sarcoma | 9 | 0 (0) | nd |
≥2+ membranous staining;
≥2+ membranous and/or cytoplasmic staining; nd - not determined.
Figure 3.
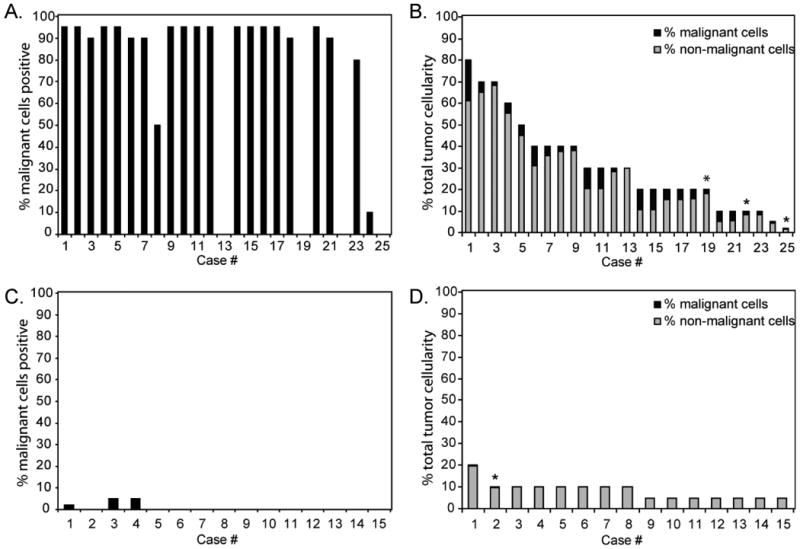
Graphical representation of the percentage of the malignant cells (A, C) and the percentage of the total tumor cellularity (malignant and non-malignant cells) (B, D) staining positive for PD-L1 in each case of NSCHL (A, B) and NLPHL (C, D) examined. * indicates cases exhibiting predominantly cytoplasmic staining.
Whereas the RS cells comprised a relatively small proportion of the overall tumor cellularity in CHL cases, non-malignant cells—predominantly tissue macrophages—contributed to the total tumor cellularity and consistently demonstrated strong membranous staining for PD-L1 (Supplemental Table S1). We found that 19/25 (76%) cases of NSCHL, 7/8 (88%) cases of MCCHL, and 5/5 (100%) cases of CHL-NOS showed strong (2+ or 3+) membranous and/or cytoplasmic staining for PD-L1 in at least 20% of the total tumor cellularity (Figure 3), including non-malignant cells (Figure 3, Supplemental Table S1). Double immunostaining for PD-L1 and CD68 on a set of cases confirmed this analysis and revealed strong membranous PD-L1 staining of CD68+ macrophages in close proximity to PD-L1 positive RS cells (Figure 2A inset). Only one case of NLPHL (7%) showed PD-L1 positive staining in 20% of the total tumor cellularity. However, malignant cells in this case were estimated to comprise 5% of the cellularity and only rare LP cells (∼2%) were positive for PD-L1 (Figure 3, Supplemental Table S1). Overall, the expression of PD-L1 on the malignant tumor cells was well correlated with the expression of PD-L1 on the tumor infiltrating CD68+ macrophages in individual tumors (Supplemental Table S3).
We have previously shown that EBV-encoded LMP1 can promote PD-L1 expression in tumor cells.6 Therefore, we looked to see if the EBV status of the Hodgkin lymphoma cases correlated with PD-L1 expression. Eleven of 12 (92%) cases of Hodgkin lymphoma that were positive for EBV showed strong (2+ or 3+) membranous PD-L1 staining in malignant RS cells. One case of EBV-positive NSCHL showed weak (1+) membranous staining in 90% of RS cells (Supplemental Table S1). Twenty of 29 (69%) Hodgkin lymphoma cases that were negative for EBV had malignant cells that scored positive for PD-L1 (Supplemental Table S1). Overall, EBV status did not predict a presence or absence of PD-L1 expression in Hodgkin lymphoma.
Two entities that lie on the histologic and genetic spectrum with CHL and NLPHL are PMBCL and TCHRBCL. Strong membranous PD-L1 expression was observed in 15/21 (71%) cases of PMBCL (90% median percent malignant cells, Figure 2C) and 10/11 (91%) cases of TCHRBCL (10% median percent malignant cells, Figure 2D). Double immunostaining for PD-L1 and PAX5 in cases of TCHRBCL demonstrated strong membranous PD-L1 staining of the scattered, PAX5-positive malignant B-cells (Figure 2D inset). PAX5-negative cells, consistent with macrophages, adjacent to the malignant B-cells also demonstrated strong PD-L1 staining. The patterns of PD-L1 expression observed using the rabbit monoclonal antibody was identical using the mouse monoclonal antibody for IHC (Supplemental Figure S2, data not shown).
Expression of PD-L1 in EBV- and HHV8-positive malignancies
Previously we reported a small series of EBV-positive PTLD cases that express PD-L1.6 Using our newly characterized PD-L1 antibodies, we extended our analysis to include additional aggressive EBV-positive and -negative B-cell lymphomas (Table 1). All EBV-positive DLBCLs, including 9/9 (100%) EBV-positive DLBCLs of the elderly and 7/7 (100%) EBV-positive immunodeficiency-related DLBCLs, demonstrated strong PD-L1 membrane staining (Figure 4A). The percentages of malignant cells staining positive for PD-L1 ranged from 5-90%, with 7 of 16 EBV-positive DLBCLs having at least 60% PD-L1-positive cells (Supplemental Table S2). Six of 10 (60%) EBV-positive PTLD cases demonstrated strong PD-L1 membrane staining on at least 80% of malignant cells, consistent with our previous study (data not shown).6 Four of 7 (57%) EBV-negative PTLD cases also showed strong membranous PD-L1 tumor staining (Figure 4B). By contrast, only 7/66 (11%) EBV-negative DLBCL-NOS cases showed strong membranous PD-L1 tumor staining (range 10-90% of malignant cells) (Figure 4C). The median tumor composition for all of the DLBCL categories ranged from 70-90%.
Figure 4.
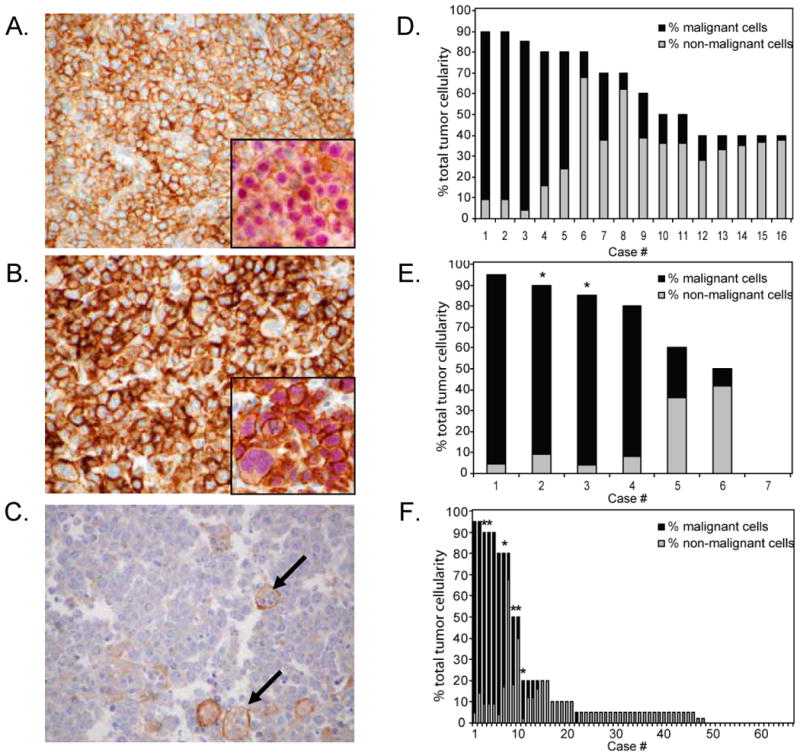
PD-L1 expression in EBV-positive DLBCLs, EBV-negative PTLD and DLBCL-NOS. (A) Representative case of an EBV-positive immunodeficiency-associated diffuse large B-cell lymphoma (DLBCL) stained with rabbit anti-PD-L1 showing distinct membranous staining of the lymphoma cells. Inset shows a case of EBV-positive DLBCL of the elderly double stained for PD-L1 (brown) and PAX5 (red). (B) Representative case of an EBV-negative post-transplant lymphoproliferative disorder (PTLD) stained with rabbit anti-PD-L1 showing distinct membranous staining of the lymphoma cells. Inset shows double staining of the same case for PD-L1 and PAX5. (C) Case of DLBCL-NOS stained with rabbit anti-PD-L1 showing no staining of the lymphoma cells but scattered intra-tumoral macrophages filled with apoptotic debris showing weak membrane positivity for PD-L1. (D-F) Graphical representation of the contribution of malignant and non-malignant cells staining positive for PD-L1 to total PD-L1 staining in each case of EBV+ DLBCL of the elderly/immunodeficiency-associated, EBV-negative PTLD, and DLBCL-NOS, respectively. * indicates cases exhibiting predominantly cytoplasmic staining.
PD-L1 expression on the variable numbers of non-malignant cells in EBV-positive DLBCL and other lymphomas consisted primarily of tumor-infiltrating macrophages (Figures 4D-F, Supplemental Table S2). Careful morphologic review and double immunostaining for CD68 or PAX5 and PD-L1 (e. g. Figure 4A and 4B insets) were used to support our histologic assessment. As for the Hodgkin lymphomas, expression of PD-L1 by malignant cells and non-malignant tumor infiltrating macrophages was correlated (Supplemental Table S3).
Analysis of additional EBV- and HHV8-positive malignancies showed that 4/9 (44%) PBL, 2/4 (50%) PEL, 4/6 (67%) ENKTCL, 16/18 (89%) NPC cases showed positive PD-L1 staining in malignant cells (Figure 5, Table 1, Supplemental Table S2). The majority of cases had greater than 50% malignant cells positive for PD-L1 and all cases in this group had at least 10% malignant cells positive for PD-L1. By contrast, no cases of EBV-positive BL or HHV8-positive KS were positive for PD-L1 (Figure 5, Table 1).
Figure 5.
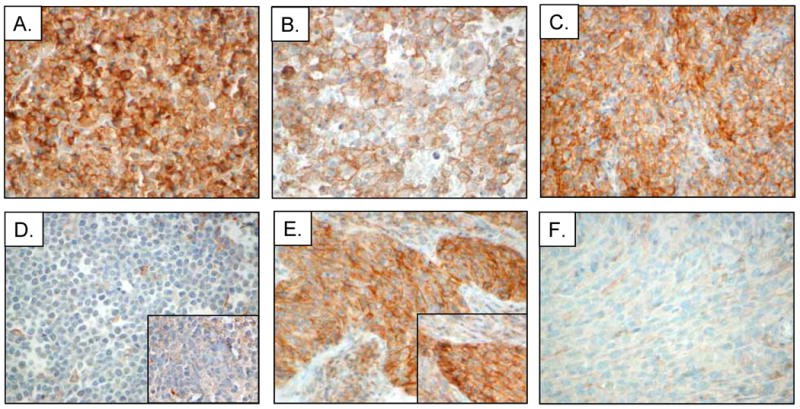
Immunohistochemical analysis of PD-L1 expression in viral-associated lymphomas and additional cancers. Representative cases of (A) EBV-positive plasmablastic lymphoma (PBL), (B) HHV8-positive primary effusion lymphoma (PEL), (C) EBV-positive extranodal NK/T-cell lymphoma (ENKTCL), (D) EBV-positive Burkitt lymphoma (BL), (E) EBV-positive nasopharyngeal carcinoma (NPC), and (F) HHV8-positive Kaposi sarcoma (KS) stained with rabbit anti-PD-L1 or mouse anti-PD-L1 (insets).
For cases with available tissue, we confirmed the patterns of PD-L1 expression obtained using the rabbit monoclonal antibody with those using the mouse monoclonal antibody. We observed no differences in tumor staining between these two reagents (Figure 5D and 5E insets; Supplemental Figure S2).
For the majority of tumors showing positive staining, the PD-L1 expression was distinctly membranous. However, in a few cases (2 NSCHL, 1 MCCHL, 4 PMBCL, 1 EBV-positive PTLD, 2 EBV-negative PTLD, 6 DLBCL-NOS, 1 PEL, 1 ENKTCL, and 2 KS), we observed cytoplasmic staining that unequivocally exceeded background (in at least 5% tumor cells) but with little to no distinct staining of cell membranes (Figure 4D-F, cases designated in graphs by *, Supplemental Table S2). Although this pattern was observed with both the rabbit and mouse anti-PD-L1 antibodies, it is unclear whether such staining represents exclusively cytoplasmic PD-L1 expression, or both cytoplasmic and low-level membrane expression.
In total, we observed robust PD-L1 expression by the malignant cells and tumor infiltrating macrophages in the majority of CHL regardless of subtype and EBV-status, PMLBCL, TCRLBCL, EBV+ DLBCL, EBV+ and EBV- PTLD, HHV8+ PEL, and EBV+ NPC. We observed that nearly one-half of PBL and ENKTCL expressed PD-L1 as well. In contrast, only rare cases of NLPHL, DLBCL NOS, BL, and no cases of KS expressed detectable PD-L1.
Discussion
EBV-associated DLBCL, PTLD, ENKTCL, PBL, NPC, and HHV8-associated PEL are rare, but often aggressive, malignancies that are largely resistant to current chemotherapeutic regimens.17–22 Given their association with oncogenic viruses and with immunodeficiency, these tumors are attractive targets for immune-based therapies. Several novel immunomodulatory agents have shown great promise in clinical trials involving patients with advanced solid tumors, especially those using human antibodies directed against critical immune checkpoint molecules. Specifically, anti-PD-1 and anti-PD-L1 directed therapies have resulted in durable clinical responses in patients with nonsmall cell lung cancer, renal cell cancer and melanoma.3,4 Furthermore, expression of PD-L1 in a subset of tumors appeared to correlate with treatment response.4 However the efficacy of such immune targeted therapies in virus- and immunodeficiency-related malignancies has not been tested. Moreover, there remains a need for reliable methods to identify the critical immunoregulatory molecules expressed by individual tumors to improve patient selection.
In this study, we validated two novel anti-PD-L1 antibodies, one of which is commercially available, and demonstrated robust PD-L1 expression in the majority of CHL/NSCHL, MCCHL, PMBCL, TCHRBCL, EBV-positive DLBCL of the immunocompromised and elderly, PTLD, NPC, and ENKTCL. A subset of PBL and PEL cases revealed unequivocal PD-L1 expression, as well. These results establish PD-L1 expression as a common feature in these malignancies, which include both viral- and immunodeficiency-associated tumors, and provide a clinically-applicable method for screening FFPE tumor samples on a routine basis.
PD-L1 expression is often driven by intrinsic genetic aberrations and disregulated signaling pathways within malignant cells. We have shown that RS cells of up to 40% of NSCHLs harbor amplification of 9p24.1, a genomic region that includes CD274, PDCD1LG2 (encoding PD-L2), and JAK2.5 In cell lines, increased 9p24 copy number is associated with increased PD-L1 expression due to both direct amplification of CD274, and through increased JAK2 and enhanced JAK-STAT signaling.6 Additional, intrinsic mechanisms driving PD-L1 expression in the RS cells of CHL include EBV infection (32% of all CHL cases in this series) and constitutive AP1 activity. Similarly, approximately 70% of primary PMLBCLs also show gains in 9p24.1,5 and thus the robust PD-L1 protein expression we observe in the majority of PMLBCLs is likely attributable, in part, to this genetic lesion. Taken together, the percentage of CHL and PMLBCL with membranous PD-L1 on the malignant cells (87% and 71%, respectively) is consistent with these prior findings.
We have shown that EBV-transformed human B-cell lines (lymphoblastoid cell lines, LCLs) and EBV-positive PTLDs upregulate PD-L1. The results in the current study both confirm the PD-L1 protein expression by EBV-positive PTLDs using newly available, more robust antibodies and establish that the majority of additional EBV-associated malignancies express PD-L1, with the exception of BL.
In LCLs, PD-L1 induction is dependent upon constitutive signaling through the EBV-encoded protein LMP1 via its effects on both the PD-L1 enhancer and promoter.6 In both LCLs and EBV-positive PTLDs, EBV maintains a type III latency program characterized by expression of LMP1 in addition to LMP2A, the nuclear proteins EBNA-1, -2,- 3 and -LP, and a series of non-coding RNAs (EBERs).12,17 Similarly, in EBV-positive CHL, EBV maintains a type II latency program that is characterized by expression of LMP1, LMP2A, EBNA1, and EBERs.23 In EBV-positive DLBCL, the virus maintains type II or type III latency program,24 and suggesting that LMP1-mediated signaling is likely to directly contribute to tumorigenesis and to the immune evasion signature for these tumors.
In contrast, the virus in endemic BL resembles that of quiescent, EBV-infected memory B cells in immunocompetent hosts and is characterized by a minimal gene expression program that includes EBNA1 and EBERs, but not LMP1.25 Consistent with a lack of signaling by EBV-encoded proteins in this tumor type, we found BL to be consistently negative for PD-L1. The EBV-encoded proteins, including LMP1, seem to be more heterogeneously expressed in ENKTCL and PBL than in EBV-positive PTLD, CHL, and DLBCL.12,20,26 Therefore, the direct contribution of EBV-dependent and independent signaling pathways to the immune signature will require further analyses using cell culture models of these tumor types. Similarly, the contribution of viral-mediated signaling to PD-L1 expression by NPC and PEL remains to be defined.
Surprisingly, we observed robust PD-L1 protein expression in EBV-negative PTLD. Very little is known about this tumor type, which comprises up to 30% of post-transplant lymphoproliferative disorders.27,28 Because patients with these tumors can still respond to a reduction in immunosuppression, there is a likely role for immune surveillance in preventing or eradicating these tumors. In our cohort, EBV-negative PTLD was similar to EBV-positive PTLD in expressing PD-L1. Because we have shown that PD-L1 can be upregulated by mechanisms independently of viral-encoded gene products in EBV-negative CHL, we suggest that this tumor type is a good candidate for further genetic studies of the CD274 locus.
Given the multiple, intrinsic mechanisms malignant cells exploit to upregulate PD-L1, an important additional result of this study is the recognition that non-malignant cells, especially tissue macrophages, contribute to the PD-L1 expression in certain aggressive B-cell, immunodeficiency-related, and viral-related malignancies. In this regard, these tumors resemble a subset of T-cell lymphoproliferative disorders in which PD-L1 expression on dendritic cells can functionally inhibit T-cell proliferation.29 We find that the intensity of PD-L1 expression in macrophages varies from absent to weak in normal tonsil and in tumors with little PD-L1 expression in malignant cells, such as NLPHL, DLBCL-NOS, and BL, to moderate and often, very strong in PD-L1-positive tumors. For several tumor types, especially CHL and other tumors with a marked inflammatory infiltrate, a major component of the PD-L1 expression within the total tumor cellularity is derived from the tumor-infiltrating macrophages. Overall, we found a high correlation between PD-L1 expression by the malignant cells and by the non-malignant cells within an individual tumor, regardless of tumor type. This correlation extends to tumors such as DLBCL-NOS, which are only rarely positive for PD-L1, and suggests coordinate regulation of PD-L1 among malignant and tumor infiltrating macrophages (Supplemental Table S3).
Recently, it has been recognized that the number of tumor infiltrating macrophages in CHL constitute a major biomarker that is predictive of inferior failure free survival and inferior overall survival.8,9 Several studies have since confirmed this observation.30–33 However, the biological basis for this association has remained undefined. PD-L1, as a major ligand for PD-1, serves to suppress T-cell immunity- including immunity directed towards virally-encoded antigens expressed in EBV-positive CHL.6 Thus our observation that macrophages constitute the major source of PD-L1 in CHL suggests that macrophage-mediated upregulation of the PD-1/PD-L1 signaling axis and suppression of anti-tumor immunity are critical factors in determining clinical outcome.
The mechanisms responsible for the recruitment of macrophages to tumors and the origin of PD-L1 upregulation on these cells require further definition. Cytokines, such as interferon-gamma, can upregulate PD-L1 on macrophages, potentially via the ISRE/IRF1 motif in the CD274 promoter.5,34 Given the cytokine-rich inflammatory microenvironment of CHL, TCRLBCL, and additional viral-associated lymphomas, secreted immunomodulatory factors may be responsible for the upregulation of PD-L1 on tumor-infiltrating macrophages in these diseases. Of note, we also observed high PD-L1 expression on the tumor-infiltrating macrophages in tumors, such as EBV-positive DLBCL and EBV-positive and -negative PTLD, in which the cytokine milieu is not well described. Further studies correlating the cytokine profiles of the microenvironment and the relative expression of PD-L1 are needed for these additional tumor types.
Given our development and validation of novel reagents for the detection of PD-L1 protein in FFPE tissue sections, it will be of interest to more broadly survey expression of this protein in additional hematological and non-hematological malignancies. Others have noted that PD-L1 is expressed by a subset of T-cell lymphoproliferative disorders, especially those at cutaneous sites.29 In contrast, low grade B-cell lymphoproliferative disorders are thought to rarely express PD-L1.35 Given our finding that PD-L1 expression can be limited to discrete and rare tumor types, it will be important to establish a comprehensive survey of hematological neoplasms based on detailed pathological classification.
Moreover, it will be of interest to survey additional virus-associated malignancies. A very recent publication detailing the expression of PD-L1 in the tumor associated macrophages of human papillomavirus (HPV)-positive squamous cell carcinoma of the head and neck provides a rationale for targeting PD-1/PD-L1 signaling in HPV associated tumors.36 Investigations of PD-L1 expression in HPV-positive cervical carcinoma, polyomavirus-positive Merkel cell carcinoma, and hepatitis B and C virus-positive hepatocellular carcinomas are warranted.
The EBV-, HHV8-, and immunodeficiency-associated malignancies are uncommon but often aggressive, life-threatening neoplasms with limited treatment options. Our data indicate that PD-L1 expression is a common feature of these tumors. Among these tumors, EBV-positive DLBCL of the elderly is associated with a particularly dismal prognosis with a median survival of 2 years.22 As this patient population is characterized by co-morbidities that complicate the use of standard chemotherapy, it is likely to be an ideal candidate for novel therapies that enhance anti-tumor immune responses. Although it will be important to interrogate larger tumor cohorts to confirm our findings, we suggest that patients with CHL, PMLBCL, TCHRBCL, EBV-positive DLBCL, PTLD, ENKTCL, PBL, PEL, and NPC may be appropriate candidates for clinical trials targeting the PD-1 pathway.
Supplementary Material
Figure S1. (A) Human placenta stained with the rabbit PD-L1 monoclonal antibody (original lot concentration of 0.22 mg/ml) and showing membrane staining of syncytiotrophoblasts and, (B) the same case stained with the mouse PD-L1 monoclonal antibody (original lot concentration of 0.65 mg/ml). (C) Human tonsil stained with the rabbit PD-L1 monoclonal antibody (original lot concentration of 1.55 mg/ml) and, (D) stained with the rabbit PD-L1 monoclonal antibody (original lot concentration of 0.22 mg/ml) and, (E) stained with the mouse PD-L1 monoclonal antibody (original lot concentration of 0.65 mg/ml).
Figure S2. (A-C) Case of classical Hodgkin lymphoma stained with (A) the rabbit PD-L1 monoclonal antibody (original lot concentration of 1.55 mg/ml), (B) the rabbit PD-L1 monoclonal antibody (original lot concentration of 0.22 mg/ml, and (C) the mouse PD-L1 monoclonal antibody (original lot concentration of 0.65 mg/ml). (E-F) Case of EBV-negative PTLD stained with (D) the rabbit PD-L1 monoclonal antibody (original lot concentration of 1.55 mg/ml), (E) the rabbit PD-L1 monoclonal antibody (original lot concentration of 0.22 mg/ml, and (F) the mouse PD-L1 monoclonal antibody (original lot concentration of 0.65 mg/ml).
Supplemental Table 1. Pathological Characteristics and PD-L1 Expression in Hodgkin Lymphomas and Related Cases
Supplemental Table 2. Pathological Characteristics and PD-L1 Expression in Non-Hodgkin Lymphoma Cases
Supplemental Table 3. Correlative Expression of PD-L1 by Malignant Cells and the Cellular Microenvironment of Examined Tumor Types
Statement of Translational Relevence.
Therapies targeting the immunomodulatory PD-1/PD-L1 signaling pathway promote marked anti-tumor immunity in a subset of patients with solid tumors. However, routine methods to detect PD-L1 expression in formalin-fixed paraffin embedded (FFPE) tissue biopsy sections are lacking. Herein, we validate novel immunohistochemical assays amenable to FFPE tissue sections and analyze a broad group of aggressive B-cell and virus-associated malignancies for PD-L1 protein expression. We find that upregulation of PD-L1 is a general feature of both the malignant cells and the non-malignant tumor infiltrating macrophages of classical Hodgkin lymphoma, primary mediastinal large B-cell lymphoma, T-cell/histiocyte-rich large B-cell lymphoma, and a broad group of virus- and immunodeficiency-associated malignancies, including EBV-positive diffuse large B-cell lymphoma, post-transplant lymphoproliferative disorder, plasmablastic lymphoma, primary effusion lymphoma, and extranodal NK/T-cell lymphoma. In addition, we detected PD-L1 expression on the malignant cells of nearly all EBV-associated nasopharyngeal carcinomas. These results indicate that PD-L1 positive tumor cells and tumor-infiltrating macrophages likely contribute to immune evasion in multiple lymphoid and virus-associated malignancies and suggest that these tumors may be amenable to PD-1 blockade.
Acknowledgments
This work was supported, in part, by R01 CA161026 (to MAS), a Specialized Center of Research (SCOR) grant from the Leukemia and Lymphoma Society (MAS; SJR), and NIH grants AI56299, AI080192, and U54CA163125 (to GJF).
Footnotes
Conflicts of interest: GJF has patents and receives patent roylaties on the PD-1 pathway. There are no other conflicts of interest from the authors to disclose.
References
- 1.Keir ME, Butte MJ, Freeman GJ, Sharpe AH. PD-1 and its ligands in tolerance and immunity. Annu Rev Immunol. 2008;26:677–704. doi: 10.1146/annurev.immunol.26.021607.090331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Freeman GJ, Long AJ, Iwai Y, Bourque K, Chernova T, Nishimura H, et al. Engagement of the PD-1 immunoinhibitory receptor by a novel B7 family member leads to negative regulation of lymphocyte activation. J Exp Med. 2000;192:1027–34. doi: 10.1084/jem.192.7.1027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Brahmer JR, Tykodi SS, Chow LQM, Hwu WJ, Topalian SL, Hwu P, et al. Safety and activity of anti-PD-L1 antibody in patients with advanced cancer. N Engl J Med. 2012;366:2455–65. doi: 10.1056/NEJMoa1200694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Topalian SL, Hodi FS, Brahmer JR, Gettinger SN, Smith DC, McDermott DF, et al. Safety, activity, and immune correlates of anti-PD-1 antibody in cancer. N Engl J Med. 2012;366:2443–54. doi: 10.1056/NEJMoa1200690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Green MR, Monti S, Rodig SJ, Juszczynski P, Currie T, O'Donnell E, et al. Integrative analysis reveals selective 9p24.1 amplification, increased PD-1 ligand expression, and further induction via JAK2 in nodular sclerosing Hodgkin lymphoma and primary mediastinal large B-cell lymphoma. Blood. 2010;116:3268–77. doi: 10.1182/blood-2010-05-282780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Green MR, Rodig S, Juszczynski P, Ouyang J, Sinha P, O'Donnell E, et al. Constitutive AP-1 activity and EBV infection induce PD-L1 in Hodgkin lymphomas and posttransplant lymphoproliferative disorders: implications for targeted therapy. Clin Cancer Res. 2012;18:1611–8. doi: 10.1158/1078-0432.CCR-11-1942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Marzec M, Zhang Q, Goradia A, Raghunath PN, Liu X, Paessler M, et al. Oncogenic kinase NPM/ALK induces through STAT3 expression of immunosuppressive protein CD274 (PD-L1, B7-H1) Proc Natl Acad Sci USA. 2008;105:20852–7. doi: 10.1073/pnas.0810958105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Tan KL, Scott DW, Hong F, Kahl BS, Fisher RI, Bartlett NL, et al. Tumor-associated macrophages predict inferior outcomes in classic Hodgkin lymphoma: a correlative study from the E2496 Intergroup trial. Blood. 2012;120:3280–7. doi: 10.1182/blood-2012-04-421057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Steidl C, Lee T, Shah SP, Farinha P, Han G, Nayar T, et al. Tumor-Associated Macrophages and Survival in Classic Hodgkin's Lymphoma. New England Journal of Medicine. 2010;362:875–85. doi: 10.1056/NEJMoa0905680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Savage KJ, Monti S, Kutok JL, Cattoretti G, Neuberg D, De Leval L, et al. The molecular signature of mediastinal large B-cell lymphoma differs from that of other diffuse large B-cell lymphomas and shares features with classical Hodgkin lymphoma. Blood. 2003;102:3871–9. doi: 10.1182/blood-2003-06-1841. [DOI] [PubMed] [Google Scholar]
- 11.Harris NL. Shades of gray between large B-cell lymphomas and Hodgkin lymphomas: differential diagnosis and biological implications. Mod Pathol. 2013;26(Suppl 1):S57–70. doi: 10.1038/modpathol.2012.182. [DOI] [PubMed] [Google Scholar]
- 12.Kutok JL, Wang F. Spectrum of Epstein-Barr virus-associated diseases. Annu Rev Pathol. 2006;1:375–404. doi: 10.1146/annurev.pathol.1.110304.100209. [DOI] [PubMed] [Google Scholar]
- 13.Taylor GS, Blackbourn DJ. Infectious agents in human cancers: lessons in immunity and immunomodulation from gammaherpesviruses EBV and KSHV. Cancer Lett. 2011;305:263–78. doi: 10.1016/j.canlet.2010.08.019. [DOI] [PubMed] [Google Scholar]
- 14.Juszczynski P, Ouyang J, Monti S, Rodig SJ, Takeyama K, Abramson J, et al. The AP1-dependent secretion of galectin-1 by Reed Sternberg cells fosters immune privilege in classical Hodgkin lymphoma. Proc Natl Acad Sci USA. 2007;104:13134–9. doi: 10.1073/pnas.0706017104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Taube JM, Anders RA, Young GD, Xu H, Sharma R, McMiller TL, et al. Colocalization of inflammatory response with B7-h1 expression in human melanocytic lesions supports an adaptive resistance mechanism of immune escape. Sci Transl Med. 2012;4:127–37. doi: 10.1126/scitranslmed.3003689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Dong H, Strome SE, Salomao DR, Tamura H, Hirano F, Flies DB, et al. Tumor-associated B7-H1 promotes T-cell apoptosis: a potential mechanism of immune evasion. Nat Med. 2002;8:793–800. doi: 10.1038/nm730. [DOI] [PubMed] [Google Scholar]
- 17.Knecht H, Berger C, Rothenberger S, Odermatt BF, Brousset P. The role of Epstein-Barr virus in neoplastic transformation. Oncology. 2001;60:289–302. doi: 10.1159/000058523. [DOI] [PubMed] [Google Scholar]
- 18.Perri F, Bosso D, Buonerba C, Lorenzo GD, Scarpati GDV. Locally advanced nasopharyngeal carcinoma: Current and emerging treatment strategies. World J Clin Oncol. 2011;2:377–83. doi: 10.5306/wjco.v2.i12.377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Nador RG, Cesarman E, Chadburn A, Dawson DB, Ansari MQ, Sald J, et al. Primary effusion lymphoma: a distinct clinicopathologic entity associated with the Kaposi's sarcoma-associated herpes virus. Blood. 1996;88:645–56. [PubMed] [Google Scholar]
- 20.Delecluse HJ, Anagnostopoulos I, Dallenbach F, Hummel M, Marafioti T, Schneider U, et al. Plasmablastic lymphomas of the oral cavity: a new entity associated with the human immunodeficiency virus infection. Blood. 1997;89:1413–20. [PubMed] [Google Scholar]
- 21.Chan JK, Sin VC, Wong KF, Ng CS, Tsang WY, Chan CH, et al. Nonnasal lymphoma expressing the natural killer cell marker CD56: a clinicopathologic study of 49 cases of an uncommon aggressive neoplasm. Blood. 1997;89:4501–13. [PubMed] [Google Scholar]
- 22.Oyama T, Yamamoto K, Asano N, Oshiro A, Suzuki R, Kagami Y, et al. Age-related EBV-associated B-cell lymphoproliferative disorders constitute a distinct clinicopathologic group: a study of 96 patients. Clin Cancer Res. 2007;13:5124–32. doi: 10.1158/1078-0432.CCR-06-2823. [DOI] [PubMed] [Google Scholar]
- 23.Herbst H, Dallenbach F, Hummel M, Niedobitek G, Pileri S, Müller-Lantzsch N, et al. Epstein-Barr virus latent membrane protein expression in Hodgkin and Reed-Sternberg cells. Proc Natl Acad Sci USA. 1991;88:4766–70. doi: 10.1073/pnas.88.11.4766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hamilton-Dutoit SJ, Rea D, Raphael M, Sandvej K, Delecluse HJ, Gisselbrecht C, et al. Epstein-Barr virus-latent gene expression and tumor cell phenotype in acquired immunodeficiency syndrome-related non-Hodgkin's lymphoma. Correlation of lymphoma phenotype with three distinct patterns of viral latency. Am J Pathol. 1993;143:1072–85. [PMC free article] [PubMed] [Google Scholar]
- 25.Vereide DT, Sugden B. Lymphomas differ in their dependence on Epstein-Barr virus. Blood. 2011;117:1977–85. doi: 10.1182/blood-2010-05-285791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Elenitoba-Johnson KS, Zarate-Osorno A, Meneses A, Krenacs L, Kingma DW, Raffeld M, et al. Cytotoxic granular protein expression, Epstein-Barr virus strain type, and latent membrane protein-1 oncogene deletions in nasal T-lymphocyte/natural killer cell lymphomas from Mexico. Mod Pathol. 1998;11:754–61. [PubMed] [Google Scholar]
- 27.Nelson BP, Nalesnik MA, Bahler DW, Locker J, Fung JJ, Swerdlow SH. Epstein-Barr virus-negative post-transplant lymphoproliferative disorders: a distinct entity? Am J Surg Pathol. 2000;24:375–85. doi: 10.1097/00000478-200003000-00006. [DOI] [PubMed] [Google Scholar]
- 28.Leblond V, Davi F, Charlotte F, Dorent R, Bitker MO, Sutton L, et al. Posttransplant lymphoproliferative disorders not associated with Epstein-Barr virus: a distinct entity? J Clin Oncol. 1998;16:2052–9. doi: 10.1200/JCO.1998.16.6.2052. [DOI] [PubMed] [Google Scholar]
- 29.Wilcox RA, Feldman AL, Wada DA, Yang ZZ, Comfere NI, Dong H, et al. B7-H1 (PD-L1, CD274) suppresses host immunity in T-cell lymphoproliferative disorders. Blood. 2009;114:2149–58. doi: 10.1182/blood-2009-04-216671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Panico L, Ronconi F, Lepore M, Tenneriello V, Cantore N, Dell'angelo AC, et al. The prognostic role of tumor associated macrophages and angiogenesis in classical Hodgkin lymphoma. Leuk Lymphoma. 2013 doi: 10.3109/10428194.2013.778405. Epub ahead of print. [DOI] [PubMed] [Google Scholar]
- 31.Jones K, Vari F, Keane C, Crooks P, Nourse JP, Seymour LA, et al. Serum CD163 and TARC as disease response biomarkers in classical Hodgkin lymphoma. Clin Cancer Res. 2013;19:731–42. doi: 10.1158/1078-0432.CCR-12-2693. [DOI] [PubMed] [Google Scholar]
- 32.Kamper P, Bendix K, Hamilton-Dutoit SJ, Honoré B, Nyengaard JR, D' Amore F. Tumor-infiltrating macrophages correlate with adverse prognosis and Epstein-Barr virus status in classical Hodgkin's lymphoma. Haematologica. 2011;96:269–76. doi: 10.3324/haematol.2010.031542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Tzankov A, Matter MS, Dirnhofer S. Refined prognostic role of CD68-positive tumor macrophages in the context of the cellular micromilieu of classical Hodgkin lymphoma. Pathobiology. 2010;77:301–8. doi: 10.1159/000321567. [DOI] [PubMed] [Google Scholar]
- 34.Zou W, Chen L. Inhibitory B7-family molecules in the tumour microenvironment. Nat Rev Immunol. 2008;8:467–77. doi: 10.1038/nri2326. [DOI] [PubMed] [Google Scholar]
- 35.Andorsky DJ, Yamada RE, Said J, Pinkus GS, Betting DJ, Timmerman JM. Programmed death ligand 1 is expressed by non-hodgkin lymphomas and inhibits the activity of tumor-associated T cells. Clin Cancer Res. 2011;17:4232–44. doi: 10.1158/1078-0432.CCR-10-2660. [DOI] [PubMed] [Google Scholar]
- 36.Lyford-Pike S, Peng S, Young GD, Taube JM, Westra WH, Akpeng B, et al. Evidence for a Role of the PD-1:PD-L1 Pathway in Immune Resistance of HPV-Associated Head and Neck Squamous Cell Carcinoma. Cancer Res. 2013;73:1733–41. doi: 10.1158/0008-5472.CAN-12-2384. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1. (A) Human placenta stained with the rabbit PD-L1 monoclonal antibody (original lot concentration of 0.22 mg/ml) and showing membrane staining of syncytiotrophoblasts and, (B) the same case stained with the mouse PD-L1 monoclonal antibody (original lot concentration of 0.65 mg/ml). (C) Human tonsil stained with the rabbit PD-L1 monoclonal antibody (original lot concentration of 1.55 mg/ml) and, (D) stained with the rabbit PD-L1 monoclonal antibody (original lot concentration of 0.22 mg/ml) and, (E) stained with the mouse PD-L1 monoclonal antibody (original lot concentration of 0.65 mg/ml).
Figure S2. (A-C) Case of classical Hodgkin lymphoma stained with (A) the rabbit PD-L1 monoclonal antibody (original lot concentration of 1.55 mg/ml), (B) the rabbit PD-L1 monoclonal antibody (original lot concentration of 0.22 mg/ml, and (C) the mouse PD-L1 monoclonal antibody (original lot concentration of 0.65 mg/ml). (E-F) Case of EBV-negative PTLD stained with (D) the rabbit PD-L1 monoclonal antibody (original lot concentration of 1.55 mg/ml), (E) the rabbit PD-L1 monoclonal antibody (original lot concentration of 0.22 mg/ml, and (F) the mouse PD-L1 monoclonal antibody (original lot concentration of 0.65 mg/ml).
Supplemental Table 1. Pathological Characteristics and PD-L1 Expression in Hodgkin Lymphomas and Related Cases
Supplemental Table 2. Pathological Characteristics and PD-L1 Expression in Non-Hodgkin Lymphoma Cases
Supplemental Table 3. Correlative Expression of PD-L1 by Malignant Cells and the Cellular Microenvironment of Examined Tumor Types